
Abstract
A 30-year-old man presented with a left undescended testis, right testicular deficiency and azoospermia. Testicular biopsy revealed an absence of spermatocytes and increased numbers of Leydig cells in the undescended testis. Additional comparative analyses were undertaken to explore Sonic Hedgehog (Shh) immunostaining in the testis of juvenile and adult mice, in the testis of the patient with cryptorchidism, and in archival testicular tissue from a patient with obstructive azoospermia and a patient with prostate cancer. Shh immunostaining was demonstrated in spermatocytes in juvenile and adult mouse testis and in the patients with obstructive azoospermia and prostate cancer, suggesting that Shh signalling is involved in normal spermatogenesis. In the patient with cryptorchidism, Shh immunostaining was localized to the Leydig cells, which suggests that Shh might be involved in the abnormal expansion of the Leydig cell population in the testis. These preliminary data on the appearance of Shh protein during normal spermatogenesis might provide the basis for further investigations to clarify the role of Shh signalling in spermatogenesis during normal and pathogenic testis development.
Introduction
Cryptorchidism, in which at least one testicle is undescended, remains a major risk factor for male infertility, 1 with 10–20% of unilateral cryporchid cases resulting in male infertility, and a higher rate of morbidity (40–80%) being observed in men with bilateral cryprochidism. 2
Testicle descent consists of two basic stages: the transabdominal stage and the inguinal–scrotal stage. The developmental processes during testicle descent are controlled by various factors including growth factors, transcription factors, steroid hormone levels and environmental factors. 3 Numerous basic and clinical studies have identified some of the relevant genetic aetiologies of cryptorchidism, for example, disorders of the Y chromosome, and of genes including insulin-like 3 (INSL-3), peptide or leucine-rich repeat-containing G protein-coupled receptor 8/G protein-coupled receptor affecting testis descent G protein-coupled receptor (RXFP2), androgen receptor (AR) and homeobox A10 (HOXA10). 3 The Hedgehog (Hh) family of secreted proteins, which includes three subgroups in vertebrates, Sonic Hedgehog (Shh), Indian Hedgehog and Desert Hedgehog (Dhh), 4 controls the formation of tissues and organs during embryonic development and it also helps to maintain the structure and function of endocrine glands in postnatal life.5,6 Dhh has been demonstrated to play an important role in ovary and testis development, and it is mainly secreted in the Sertoli cells of testicles and ovarian granular cells.7,8 Shh signalling is thought to be the most extensively studied of the Hh family and it participates in multiple developmental processes. 9 Studies have suggested that the Shh signalling pathway serves as the key regulator of the cross talk between steroidogenic factor (Sf)-positive cells and Sf-negative cells in the capsule of the adrenal gland, and it is mainly secreted by the steroidogenic cells (Sf-negative). 9 Sf-1 (also known as NR5A1) is directly responsible for the secretion of steroid hormones, and the precise expression of the NR5A1 gene controls the normal development and function of adrenal glands and testicles. 10 The Shh-dependent synthesis of corticosteroid in the adrenal gland has confirmed the direct interaction between Shh signalling and Sf-1 in adrenal development. 10
In the testis, the levels of steroidogenic enzymes are also controlled by the NR5A1 gene, and thereby affect the production of testosterone and INSL-3 peptide in the testis. 8 A disruption in the synthesis of androgens can lead to cryptorchidism, as indicated by the correlation between cryptorchidism and expression of the AR gene. 3 It is reasonable to hypothesize that Shh signalling might play a role in testicular development, most probably by either regulating the synthesis of androgens (which is mainly produced by the Leydig cells) or by controlling the proliferation and differentiation of the testicular Leydig cell population, similarly to that observed in the adrenal gland. Some testicular diseases might also be attributed to a disorder of the Shh signalling pathway, but there is little evidence at present. Based on this hypothesis, this present study was designed to explore the Shh immunostaining patterns in a case of cryptorchidism with nonobstructive azoospermia compared with healthy mouse and human testis.
Case report
A 30-year-old man presented to the Sperm Development and Genetics Laboratory, Shanghai Human Sperm Bank, Shanghai Institute of Andrology, Department of Urology, Renji Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, China in April 2012 after several years of infertility and with a history of azoospermia and a left inguinal hernia. The patient showed no discomfort, he was without any history of infection or haematoma of the scrotum, and his family had no relevant hereditary diseases. The initial physical examination revealed a normally proportioned body without any sign of constitutional symptoms. However, the scrotum was small and no testes were palpated in it. The left testis was palpable in the inguinal region but smaller in size than normal (about 10 cm3) and soft in texture, whereas the penis was normal and asymptomatic. Ultrasonographic scanning of the scrotum was performed and showed the absence of testes. A heterogeneous mass with a regular border was observed in the left inguinal region, measuring 29 mm × 14 mm × 24 mm (about 9.8 cm3). Results of a pelvic computed tomography scan were consistent with the ultrasonography scan described above: it demonstrated an additional solid mass located in the inguinal region, which was thought to be the left testis. The patient presented a 46, XY, 14pstk+ karyotype, as determined by chromosome analysis of cultured lymphocytes from peripheral blood, and no microdeletions of the Y chromosome were detected. Finally, the patient was diagnosed as cryptorchidism with right testicular deficiency and nonobstructive azoospermia by clinical guidelines. He underwent a left orchiopexy. Finally, testicular biopsies were obtained from this patient to confirm the presence of the undescended left testicle.
Using routine clinical laboratory enzyme immunoassays, hormone analyses showed that the serum follicle stimulating hormone and luteinizing hormone concentrations were within the normal range: 11.7 and 3.8 mIU/ml, respectively (normal ranges: 1.3–11.8 mIU/ml and 1.8–8.4 mIU/ml, respectively). The serum level of total testosterone was 3.0 ng/ml (normal range: 2.6–7.4 ng/ml). Prolactin and 17β-oestradiol levels also appeared to be normal (11.5 µg/l and 16.6 pg/ml, respectively; normal range: 4.1–18.5 µg/l and 0.0–50.0 pg/ml, respectively). Three independent semen examinations were conducted and no sperm could be found, even after routine centrifugation.
This study was conducted following approval from the Ethics Committee of Renji Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, China. The 30-year-old male patient with cryptorchidism gave written informed consent to participate in this study. The two control patients gave written informed consent to allow their archival testicular samples to be used in this study for the additional analyses.
Additional analyses
Cellular localization of Shh proteins in juvenile and adult mouse testis
Immunofluorescence staining was performed to detect which cells synthesize Shh proteins in the testis of 2-week-old juvenile (n = 3; 3.5–4.5 g) and 9-week-old adult (n = 3; 24–26 g) C57BL/6 mice (specific-pathogen-free animals; Bio-X Centre, Shanghai Jiao Tong University, Shanghai, China). The mice were housed under a 12-h light–dark cycle (lights on at 08:00 hours) with free access to food and water. The protocol for the animal study was reviewed and approved by the Animal Experiment Committee, School of Medicine, Shanghai Jiao Tong University.
First, the testis tissues were frozen in optimum cutting temperature compound (Tissue-Tek® OCT Compound 4583; Sakura Finetek, Torrance, CA, USA). Then, frozen sections (8 µm) were cut and blocked with 1% sheep serum in 1.0 M phosphate-buffered saline (PBS; pH 7.2) for 30 min at room temperature. The sections were then incubated with primary rabbit antihuman Shh polyclonal antibody (1 : 100 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA) diluted in 1.0 M PBS (pH 7.2) for 12 h at 4℃. The tissue sections were then washed three times (5 min per wash) with 1.0 M PBS (pH 7.2), followed by incubation with fluorescein isothiocyanate-goat antirabbit immunoglobulin (Ig) G secondary antibody (1 : 200 dilution; Sigma-Aldrich, St Louis, MO, USA) for 1 h at 37℃. The slides were then washed three times (5 min per wash) with 1.0 M PBS (pH 7.2). The slides were stained with 5 µg/ml 2 -(4-amidinophenyl)-6-indolecarbamidine dihydrochloride for 5 min at 37℃ to stain the cell nuclei. Finally, the sections were examined under a confocal microscope (TCS SP5; Leica Microsystems, Mannheim, Germany) or an optical microscope (CX21; Olympus Optical, Tokyo, Japan).
The results showed that Shh protein immunostaining was only observed in primary or secondary spermatocytes at 2 weeks of age (Figure 1), which are located in the centre of the seminiferous tubules. By adulthood (9 weeks), the Shh proteins were still observed in the spermatocytes, but the Sertoli and Leydig cells were negative for Shh protein. The immunostaining of the Shh proteins was specifically located in the spermatocytes from juvenile and adult mice, which indicates the potentially important function of Shh signalling in the meiotic process of spermatogenesis. These findings in mice suggest that the Shh protein could be used as a distinctive marker of spermatocytes due to its strong and specific immunostaining in these cells within the testis.
Detection of Sonic Hedgehog (Shh) protein in mouse testis by immunofluorescence staining: (a) 2-week-old juvenile mouse testis; and (b) 9-week-old adult mouse testis. Haematoxylin and eosin (H&E) staining is shown on the left hand side. Cell nuclei have been stained with 2 -(4-amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI). The Shh proteins were strongly immunostained in the spermatocytes. Scale bar, 200 µm. The colour version of this figure is available at: http://imr.sagepub.com
Levels of Shh protein and mRNA in the testis of the patient with cryptorchidism, a patient with obstructive azoospermia, and a patient with prostate cancer
To investigate the immunostaining pattern of the Shh protein in human testis, immunofluorescence examinations of sections of testis from the 30-year-old male cryptorchid patient (i.e. from the undescended left testicle), one patient with obstructive azoospermia and one patient with prostate cancer (these two patients were used as controls due to their almost normal testicular spermatogenic function, as shown by pathology results) were undertaken using testicular biopsies obtained from the Department of Pathology, Renji Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, China. The immunofluorescence methodology was identical to that described above for the mouse testis.
For the cryptorchid patient, the results of haematoxylin and eosin (H&E) staining confirmed the absence of spermatocytes in the testis (Figure 2a,), while the other two male patients showed sufficient spermatocytes (Figures 2b and 2c; H&E). With regard to the immunostaining pattern of the Shh protein, the results were consistent with those of the mouse studies, with strong staining of spermatocytes observed in the two patients with normal spermatocytes (patient with obstructive azoospermia and patient with prostate cancer; Figures 2b and c, respectively). However, despite the lack of spermatocytes, the cryptorchid patient still showed an immunofluorescent signal in the testis that was mainly located in the expanded population of Leydig cells (Figure 2a).
Immunofluorescence staining of Sonic Hedgehog (Shh) protein in sections of the testis from: the 30-year-old male patient with cryptorchidism (a); a patient with obstructive azoospermia (OA) (b); and a patient with prostate cancer (Pca) (c). Haematoxylin and eosin (H&E) staining is shown on the left hand side. Cell nuclei have been stained with 2 -(4-amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI). The immunostaining of Shh proteins in the Leydig cells of the male cryptorchid patient is clearly shown. Scale bar, 100 µm. The colour version of this figure is available at: http://imr.sagepub.com
Quantitative real-time reverse transcription–polymerase chain reaction (RT–PCR) was used to measure the levels of Shh mRNA in samples of testis from the 30-year-old male cryptorchid patient, the patient with obstructive azoospermia and the patient with prostate cancer. Total RNA was extracted using TRIzol® reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The RT reaction was performed using the Reverse Transcription System A3500 kit provided by Promega (Madison, WI, USA). Quantitative real-time RT–PCR was conducted with 10 µl SYBR® Green Master Mix (Life Technologies, Paisley, UK) according to the manufacturer's instructions using the Applied Biosystems® 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). The primer sequences for the internal control β-actin mRNA were as follows: forward, 5′-AAGATCAAGATCATTGCTCCTC-3′, and reverse, 5′-GGGTGTAACGCAACTAAGTC-3′ (Shanghai Generay Biotech, Shanghai, China). The primer sequences for the Shh mRNA were as follows: forward, 5′-TTAGCCTACAAGCAGTTTATCC-3′, and reverse, 5′-TGTCCTTACACCTCTGAGTC-3′ (Shanghai Generay Biotech). The sizes of the PCR amplification products were as follows: β-actin 181 base pairs (bp); Shh 180 bp. The PCR cycling programme involved preliminary denaturation at 95℃ for 10 min, followed by 30 cycles of denaturation at 95℃ for 15 s and annealing at 60℃ for 1 min. The PCR products were subjected to agarose gel analysis (1.5% agarose gels) and visualized using ethidium bromide staining and ultraviolet light (Figures 3a and 3b). The threshold cycle (CT) values were measured and calculated using Microsoft® Excel software (Microsoft, Redmond, WA, USA). Higher Shh mRNA levels were seen in the patients with obstructive azoospermia (about 5.0-fold higher) and prostate cancer (about 6.2-fold higher) compared with the patient with cryptorchidism.
Analysis of Sonic Hedgehog (Shh) mRNA and protein levels using quantitative real-time reverse transcription–polymerase chain reaction (RT–PCR) (a, b) and Western blot analysis (c) of testis samples from the 30-year-old male patient with cryptorchidism, from a patient with obstructive azoospermia (OA), and from a patient with prostate cancer (Pca). The results of the RT–PCR are expressed as mean ± SD based on three analyses for each sample (a). The products of quantitative real-time RT–PCR were assessed by agarose gel analysis and compared with the internal control β-actin mRNA (b). The internal control β-actin was also used for the Western blot analysis (c).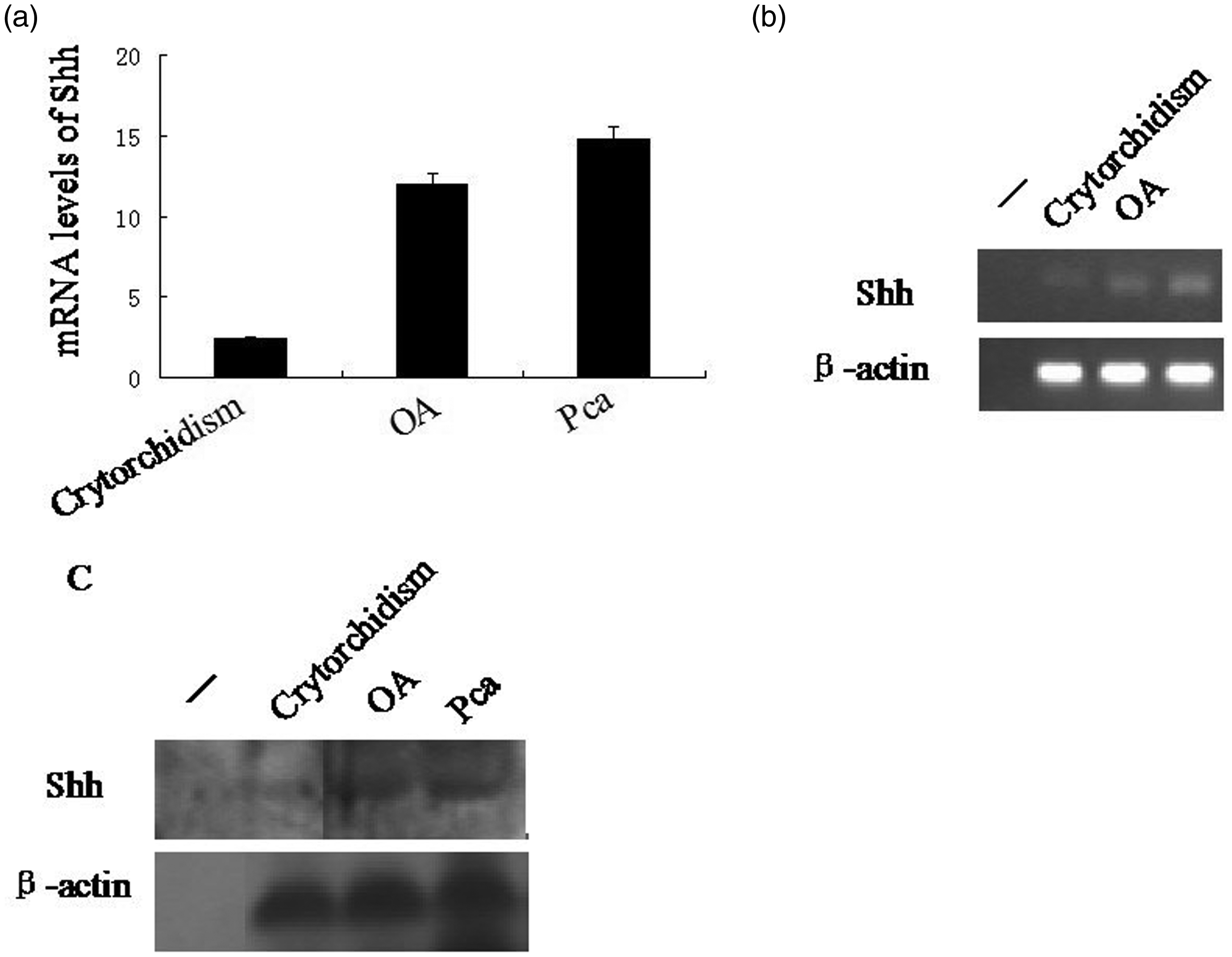
The levels of Shh protein in the three samples of human testis were also investigated using Western blot analysis. Testis samples were lysed with 5 × passive lysis buffer (Promega) and the protein content was quantified using a spectrophotometric assay (Chinese Academy of Sciences, Shanghai, China). Protein extracts (100 µg) were separated using 10% sodium dodecyl sulphate–polyacrylamide gel electrophoresis, then transferred to a polyvinylidene fluoride membrane (PVDF) membrane (Beyotime Institute, Shanghai, China) using Bio-Rad Trans-Blot® equipment (Bio-Rad, Hercules, CA, USA). The PVDF membrane was then blocked in 5% nonfat milk in Tris-buffered saline Tween-20 (TBST; pH 7.5; 20 mmol/l Tris–HCl, 150 mmol/l sodium chloride, 0.1% Tween-20; Beyotime Institute) at 37℃ for 1 h. The PVDF membrane was then incubated with rabbit antihuman Shh or rabbit antihuman β-actin primary antibodies (each diluted 1 : 10 000; Sigma-Aldrich) for 12 h at 4℃. Then it was washed three times (5 min per wash) with TBST (pH 7.5), followed by incubation with horseradish peroxidase-conjugated goat antirabbit IgG secondary antibody (1 : 8000 dilution; Sigma-Aldrich) for 2 h at 37℃. The membrane was washed three times (5 min per wash) with TBST (pH 7.5). Visualization of the protein bands was achieved using Thermo Scientific SuperSignal West Pico Chemiluminescent Substrate (Thermo Scientific, Rockford, IL, USA) according to the manufacturer’s instructions. The protein bands were photographed using an FR-200 A Bio-Electrophoresis Image Analysis System (Furi, Shanghai, China). There was a weaker Shh protein signal in the testis sample from the 30-year-old male patient with cryptorchidism compared with the patients with obstructive azoospermia and prostate cancer (Figure 3c).
Discussion
Immunostaining of Patched (PTCH) 2 in spermatocytes has been reported by Carpenter et al., 9 and this preliminary study further reports immunohistochemical detection of Shh proteins in the spermatocytes of both juvenile and adult mouse testis. This present study also demonstrated the same pattern of Shh immunostaining in normal adult human testis, suggesting its importance in spermatogenesis. A broad pattern of Shh protein secretion has been reported in a range of tissues during normal development and in some cancers during tumorigenesis. 10 For example, epithelial tissues, limb buds, adrenal gland, central nervous system, pancreatic cancer and gastric adenocarcinomas have all been demonstrated to have Shh protein secretion, 10 but not the testis. Whether or not the Shh protein is a marker for and regulator of spermatocytes needs to be confirmed by further research, and the details of the role played by Shh signalling during sperm formation remains unknown.
Although little is known about Shh signalling in the testis, the crucial role that it plays in the adrenal developmental processes has been verified by considerable data, which have demonstrated that Shh mainly influences the cortical progenitor cells by interacting with the NR5A1 gene, and thereby affecting the synthesis of glucocorticoids. 11 Moreover, in the testis, studies have demonstrated that Sf-1 is also produced by the Sertoli and Leydig cells. 8 Sf-1 has been shown to regulate the synthesis and secretion of steroid hormone (androgen) through directing the expression of steroidogenic enzymes, such as steroidogenic acute regulatory cytochrome P450, to produce testosterone. 8 Therefore, the similarity between the developmental processes and the synthesis of steroid hormone in the testis and adrenal gland suggest that Shh signalling might be related to the regulation of androgen production and it might also regulate the process of spermatogenesis via an adrenal-like mechanism. However, detailed studies of the mechanism of Shh signalling revealed that Shh signalling was required for the nonsteroidogenic cells, but not the steroidogenic cells, during adrenal development. 11 In the adrenal cortex, Shh protein is located in the steroidogenic cells, which mainly synthesize glucocorticoids 11 Using Shh−/− mice, research has demonstrated that Shh promoted the proliferation of adrenal capsule cells. 12 These data indicated that the Shh signalling pathway functioned as a junction between Sf-positive cells and Sf-negative cells, reflecting the crucial role that it plays in the development and maintenance of the adrenal capsule and cortex. 12 Moreover, targeted disruption of the Shh gene in mice showed the phenotype of adrenal hypoplasia with the defects in the adrenal cortex. 13 The Shh gene is expressed in Sf-positive cells, while the direct target is the Sf-negative cells, controlling the normal development of the adrenal capsule and cortex. 12
The present study has provided evidence that Shh protein is located in the spermatocytes of healthy juvenile and adult mice and in adult humans with normal testicular development. The receptors for Shh protein, Patched2 and Smoothened, are also expressed in spermatogonia during testis development, indicating that Shh is an essential morphogen in testis development,5,7 especially for spermatogenesis. Because the regulatory mechanism between Shh signalling and the NR5A1 gene has not been elucidated, it is not certain whether Shh signalling controls the synthesis of testosterone and whether the ectopic expression of Shh signalling in the Leydig cells in this present case contributed to the development of an undescended testis. Testosterone and INSL-3 peptide, produced by the Leydig cells, are both regulated by Sf-1 and if there is a disruption in their synthesis this can lead to cryptorchidism. 3
The present case report demonstrated ectopic secretion of Shh protein in the Leydig cells of the left undescended testis of a 30-year-old male patient with cryptorchidism. This present finding appears to be consistent with a previous report that demonstrated that ectopic Shh signalling was observed in the expanded Leydig cells of mice in which Sf-1/NR5A1 sumoylation had been eliminated, which resulted in disruption of testicular development. 14 Since the interaction between Shh and the NR5A1 gene in adrenal development has been clarified above, ectopic Shh signalling might also interact with the NR5A1 gene in the testis, leading to an increase in the number of Leydig cells and testicular dysfunction.
In conclusion, this present case report has demonstrated novel ectopic Shh protein immunostaining in the Leydig cells in the undescended left testis of a 30-year-old male patient with cryptorchidism, which suggests that Shh might be involved in the expansion of the Leydig cell population in the testis. This is in direct contrast to the pattern of immunostaining that was observed in normal juvenile and adult mice and adult human testis, which demonstrated that Shh protein is usually localized to the spermatocytes lining the seminiferous tubules. These preliminary data on the appearance of Shh protein during normal spermatogenesis might provide the basis for further investigations to clarify the role of Shh signalling in spermatogenesis during normal and pathogenic testis development.
Footnotes
Declaration of conflicting interest
The authors declare that there are no conflicts of interest.
Funding
This study received funding from The National Natural Science Foundation of China (Grant no. 30973069, 30672146) and the Foundation of Pujiang (Grant no. 08PJ14100).