
Abstract
Objective
To identify dysregulated miRNAs in testicular tissues from animal models and patients with cryptorchidism.
Methods
Databases were systematically searched for studies published before 10 May 2020 that had investigated miRNAs in cryptorchidism. Predicted targets of the identified miRNA biomarkers were obtained by searching TargetScan and Starbase. Gene ontology (GO) and Kyoto Encyclopaedia of Genes and Genomes (KEGG) pathway enrichment analyses were subsequently conducted.
Results
Five publications met the eligibility criteria for the review. 21 differentially expressed miRNAs were the most abundantly reported in 185 animal and human tissue samples. Three miRNAs (miR-210, miR-449a and miR-34c) were dysregulated in both animal and human testicular tissues. The top five relevant lncRNAs associated with the miRNAs were NEAT1, KCNQ1OT1, XIST, AC005154.1, and TUG1.
Conclusions
Further research is warranted to explore the potential of these dysregulated miRNAs as biomarkers or therapeutic targets for male infertility associated with cryptorchidism.
Introduction
Cryptorchidism or undescended testis (UDT) is the most common childhood congenital malformation in boys and occurs in approximately 1.0–4.6% of full-term and up to 45% of preterm neonates. 1 Following spontaneous descent within the first month of life up to 1.5% of all full-term male infants still have undescended testes at the age of one year. 2 The exact mechanism of testicular descent is not fully understood and UDT can occur anywhere along the normal developmental descent pathway, from the abdominal cavity to entrance of the scrotum. 3
While cryptorchidism can be treated by early surgery to improve cosmetic appearance and reduce the possibility of malignancy, some men exhibit low fertility as adults.4,5 In fact, 20%∼25% of boys with UDT may be at risk of infertility after treatment. 6 It has been suggested that blood hormone levels and testicular biopsies in these patients could assist in the assessment of adult reproductive function. 6 However, often due to ethical reasons, it is difficult to obtain testicular tissues from boys with UDT. Therefore, there is an urgent need to find a non-invasive marker that may improve the prevention and prognosis of cryptorchidism.
Micro (mi)RNAs are a class of short non-coding RNA molecules approximately 22 nucleotides in length, that negatively regulate gene expression either by mRNA degradation or translational repression at the post-transcriptional level. 7–11Several studies have investigated the potential role of miRNAs in spermatogenesis,12–16 and it has been suggested that miRNAs may regulate early embryonic development, spermatic function, and fertilization in humans and animals. 17 Indeed, the dysregulation of miRNAs has been shown to be a common event in spermatogenesis arrest and spermatocyte apoptosis.18–20Therefore, miRNAs may have potential as non-invasive diagnostic markers as well as therapeutic targets for male infertility. Accordingly, we conducted a systematic review according to PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) regulations 21 to identify dysregulated miRNAs associated with cryptorchidism.
Methods
PubMed, Web of Science and EMBASE databases were systematically searched for all studies published before 10 May 2020 that had investigated miRNAs in cryptorchidism. Keywords/terms included: (“Cryptorchidism” [Mesh]) “Cryptorchidism” OR “Cryptorchism” OR “Undescended Testes” OR “Undescended Testis” OR “UDT” AND (“MicroRNAs” [Mesh] OR “MicroRNAs” OR “Micro RNA” OR “MicroRNA” OR “miRNAs” OR “miRNA” OR “pri-miRNA” OR “pre-miRNA” OR “Small Temporal RNA” OR “stRNA”). All miRNA names were standardized by the miRbase. 22
Studies included in the review had miRNA expression data, from animal models or patients with cryptorchidism and controls. Studies were excluded if they used cell lines, had insufficient data, and/or were reviews, commentaries, editorials, or meeting abstracts. The following items were extracted: title; authors; date; country; species; miRNA expression profiling assay type; number of dysregulated miRNAs (downregulated and upregulated).
To identify the potential target of the miRNAs we used Starbase a database that provides comprehensive information on miRNA and long non-coding (lnc)RNA interactions 23,24The top 20 lncRNAs most closely related to the miRNAs were visualised using clusterProfiler (R) installed with Seaborn. In addition, to identify potential subcellular localizations of the lncRNAs, we used two databases (lncLocator 25 and lncATLAS 26 ). The lncLocator has the ability to predict five subcellular localizations of lncRNAs (i.e., cytoplasm, nucleus, cytosol, ribosome, and exosome 25 Results were analysed and presented by GraphPad Prism 7. LncRNAs considered the most relevant to cryptorchidism were uploaded into the lncATLAS database. 26
TargetScan was used to predict potential biological targets of the miRNAs. 27 Gene Ontology (GO) enrichment analysis and Kyoto Encyclopaedia of Genes and Genomes (KEGG) pathway analysis were subsequently conducted.
Results
The literature search yielded an initial pool of 55 articles from which five reports ultimately met the eligibility criteria (Figure 1).12–16 Four studies were performed in China and one in Japan (Table 1); three studies used human testicular tissues, two murine, one horse and one rat.
Characteristics of miRNA expression studies included in the systematic review.
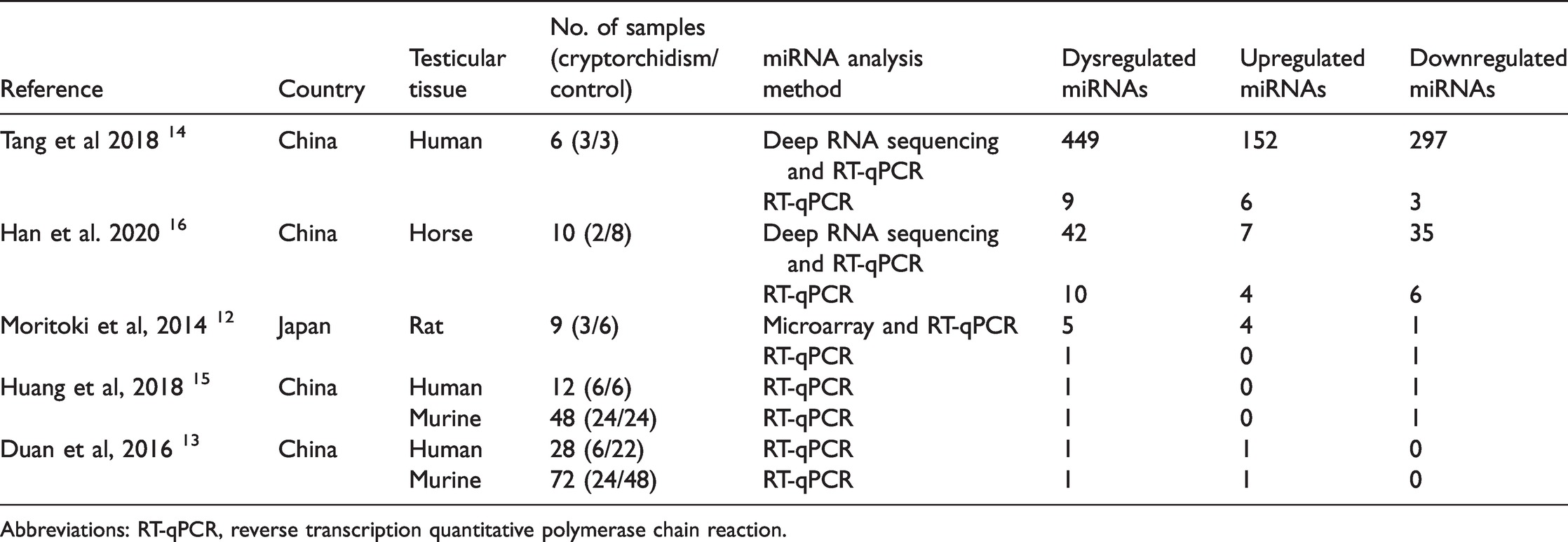
Abbreviations: RT-qPCR, reverse transcription quantitative polymerase chain reaction.
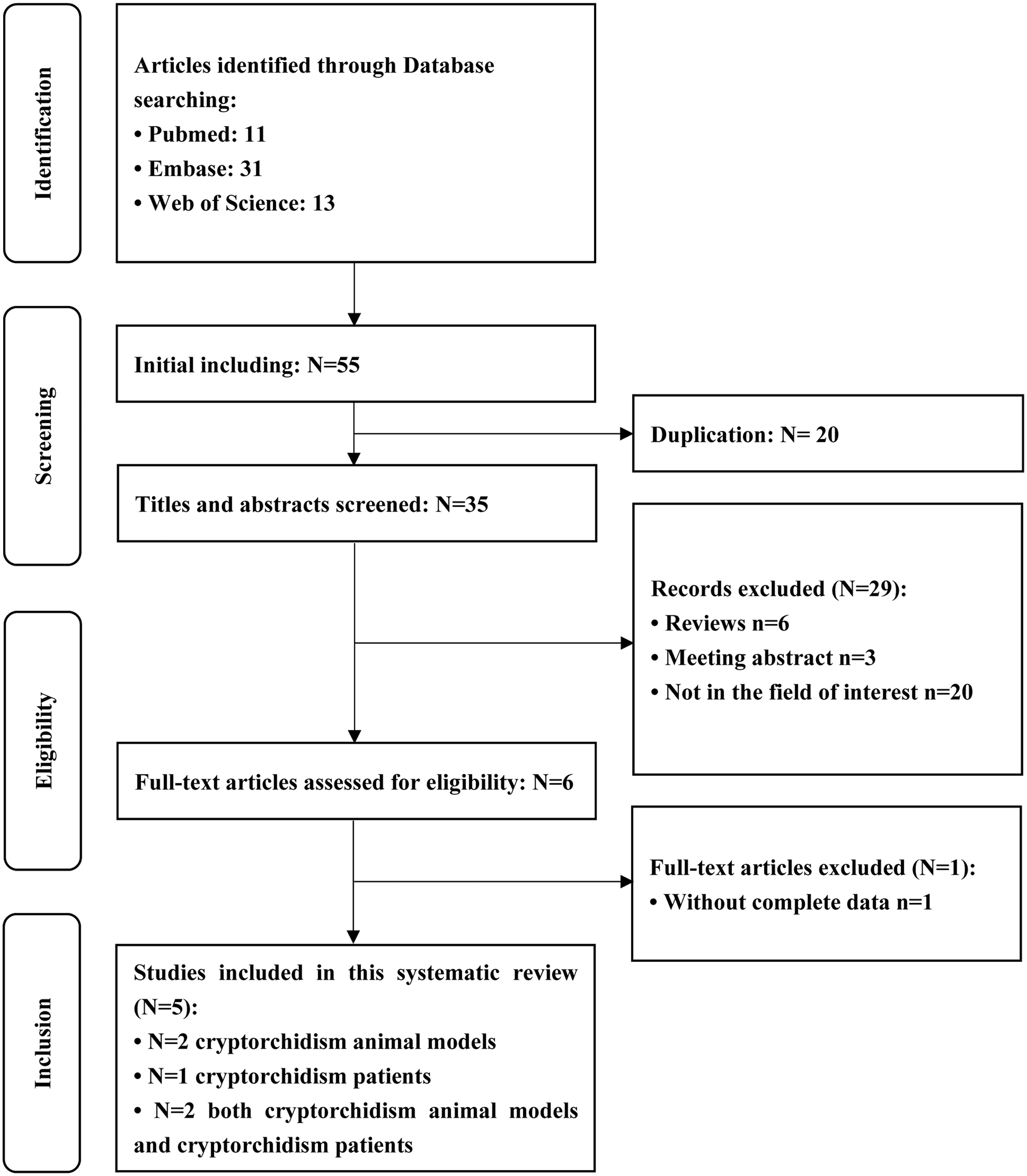
Flow diagram of study selection.
Analysis showed that 21 differentially expressed miRNAs were the most abundant in testicular tissue samples obtained from animal models and patients with cryptorchidism. In tissues from animal models, five miRNAs were differentially upregulated and eight downregulated. (Table 2). In human tissues, seven miRNAs were differentially upregulated and four downregulated. (Table 3). Three miRNAs (mi-210, miR-449a, miR-34c) were dysregulated in both types of tissues, miR-210 was upregulated, 13 whereas, miR-449a and miR-34c were downregulated.14–16 The overlap of most abundant miRNAs among animal and human tissues is represented as a Venn diagram in Figure 2.
The most abundant differentially expressed miRNAs in testicular tissue from animal models of cryptorchidism (validated by qRT-PCR).
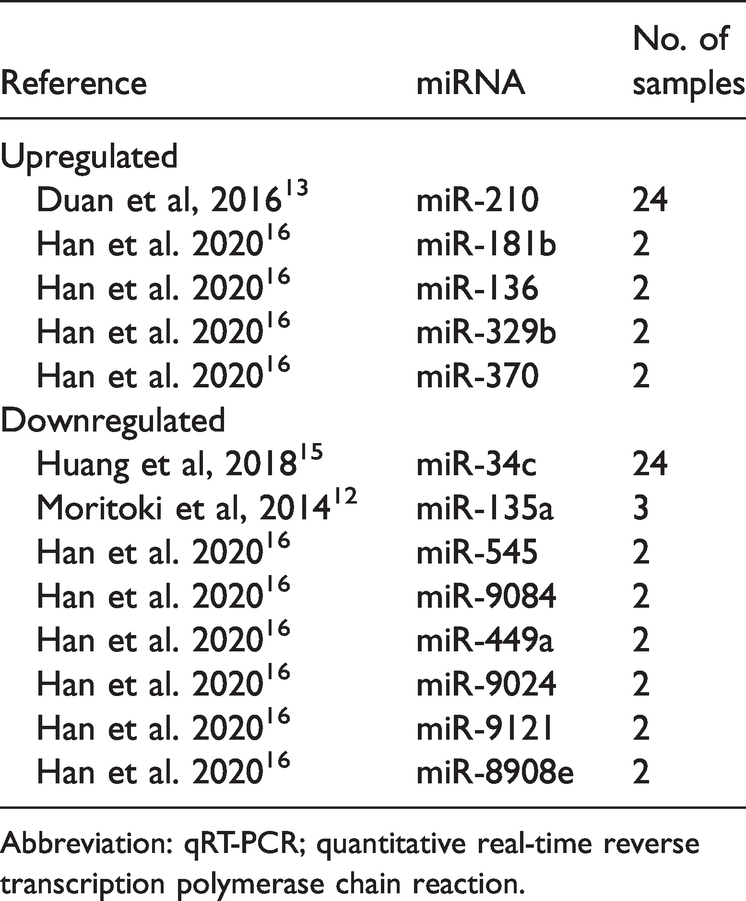
Abbreviation: qRT-PCR; quantitative real-time reverse transcription polymerase chain reaction.
The most abundant differentially expressed miRNAs in testicular tissue from patients with cryptorchidism (validated by qRT-PCR).

Abbreviation: qRT-PCR; quantitative real-time reverse transcription polymerase chain reaction.
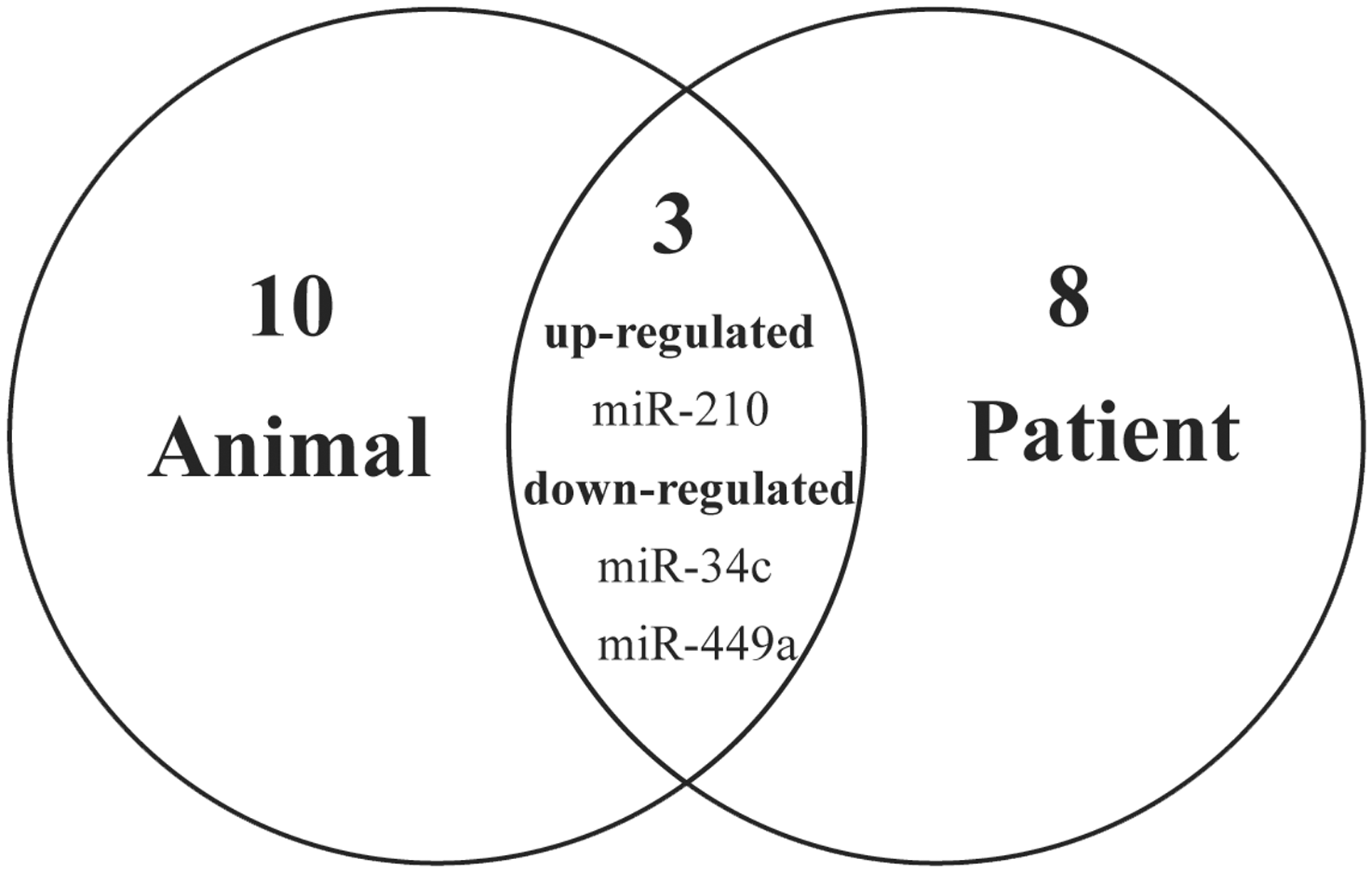
Venn diagram showing the most abundant miRNAs detected in testicular tissue samples obtained from animal models and patients with cryptorchidism. Of the 21 dysregulated miRNAs detected, 10 were in human tissues, eight in animal tissues and three (miR-210 [upregulated], miR-449a and miR-34c [downregulated]) were in both animal and human tissues.
The top 20 lncRNAs most closely related to the top miRNAs are shown in Figure 3. The top five lncRNAs were, NEAT1 (nuclear paraspeckle assembly transcript 1), KCNQ1OT1, XIST (X inactive specific transcript), AC005154.1 and TUG1(taurine up-regulated 1). Among the top 20 lncRNAs, 12 were identified by their sequences uploaded in a fasta file on lncLocator. The current lncLocator predicted five subcellular localizations of these validated lncRNAs and the results were sorted by possible proportions (Figure 4A). Four lncRNAs (NEAT1, KCNQ1OT1, XIST and TUG1) tended to be located in the nucleus. Results from lncATLAS confirmed that both NEAT1 and TUG1 primarily existed in the nucleus (Figure 4B).

The top 20 lncRNAs most closely related to the dysregulated miRNAs.
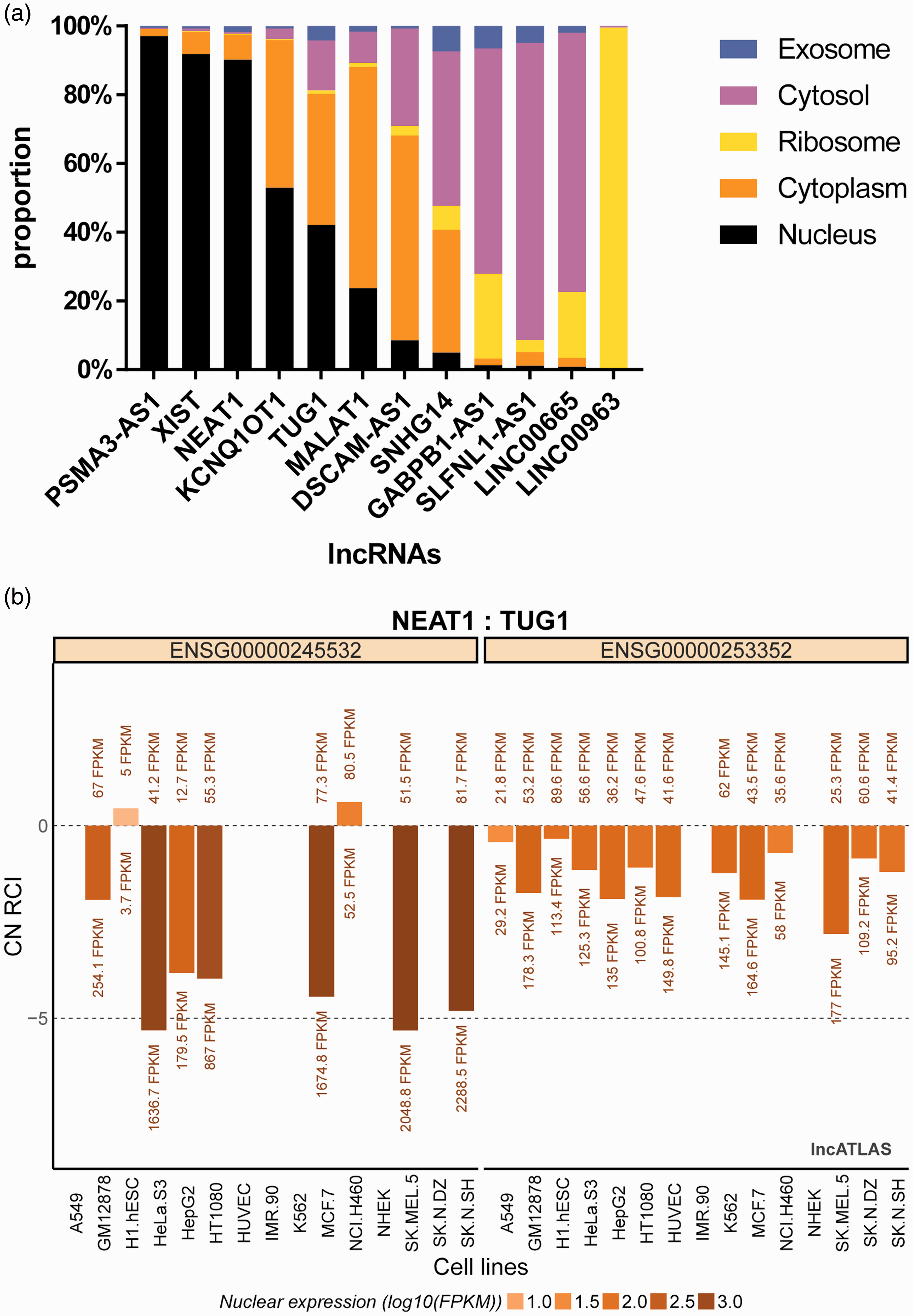
(a) The lncLocator predicted five subcellular localizations of the 12 validated lncRNAs and the results were sorted by possible proportions. Among the top five lncRNAs, NEAT1, KCNQ1OT1, XIST, TUG1 tended to be located in the nucleus. (b) Subcellular localization plot displayed by lncATLAS for NEAT1 and TUG1 genes. Bars represent CN-RCI (relative concentration index calculated for the cytoplasm and nucleus) values for the genes across the cell lines. Nuclear expression values (FPKMs) for the genes are shown for both compartments (cytoplasm >zero, nucleus <zero). Bars are coloured by their absolute nuclear expression.

(a) Functional enrichment analysis of differentially expressed miRNAs by Kyoto Encyclopaedia of Genes and Genomes (KEGG) pathway analysis. The bubble size was directly proportional to the number of miRNAs. Among the most enriched KEGG pathways, the top five terms were prostate cancer, miRNAs in cancer, autophagy, cellular senescence, and bacterial invasion of epithelial cells. (b) GO enrichment analysis of the differentially expressed miRNAs (Top 10 GO enrichment are presented). The most significant GO term under the biological process category was ‘cotranslational protein targeting to membrane’. ‘Endosome’ and ‘synapse’ were the most enriched GO terms under cellular component category and ‘glutamate binding’ was the most enriched term under molecular function category.
KEGG pathway analysis indicated that the differentially expressed genes tended to participate in pathways involving prostate cancer, microRNAs in cancer, autophagy, cellular senescence, and bacterial invasion of epithelial cells (Figure 5A).
The top 10 enriched GO terms, included in ‘biological process’, ‘cellular component’ and ‘molecular function’ are displayed in Figure 5B. ‘Cotranslational protein targeting to membrane’ was the most significant GO term under ‘biological process’ ‘endosome’ and ‘synapse’ were the most enriched terms under ‘cellular component’ and ‘glutamate binding’ was the most enriched term under ‘molecular function’.
Discussion
We performed a systematic review to identify studies of miRNAs in animal models or patients with cryptorchidism. Levels of miRNAs in testicular tissue or seminal fluid may have potential as non-invasive diagnostic markers as well as be therapeutic targets for male infertility. 28 Five studies met our eligibility criteria and had investigated dysregulated miRNA in cryptorchidism.12–16 Of the 21 miRNAs that were most abundant, three (miR-210, miR-449a and miR-34c) were dysregulated in both animal and human testicular tissues.13–16 In one study in a rat model of cryptorchidism, miR-135a was expressed at a low level and had a downregulation effect on Forehead box protein O1 (FoxO1) which is essential for spermatogonial stem cell maintenance. 12 It has been reported that both miRNAs and mRNAs contribute to the formation and differentiation of spermatogonia stem cells.29–31 Therefore, it is important to determine the exact function of each miRNA in testicular tissue and determine the molecular pathways involved.
Using Starbase, we identified the top five lncRNAs targets of the miRNAs and four of these (i.e., NEAT1, KCNQ1OT1, XIST, TUG1) tended to be located in the nucleus. Subsequent data from lncATLAS confirmed that NEAT1 and TUG1 primarily existed in the nucleus. NEAT1 is thought to act as a miRNA sponge suppressing the interactions between miRNAs and target mRNAs. 32 In addition to its role in several cancers, 32 NEAT 1 has been reported to exert modulated action in several other diseases, such as non-alcoholic fatty liver disease, 33 Parkinson's disease 34 and congenital heart disease. 35 NEAT1 has been reported to alleviate hypoxia-provoked H9c2 cell apoptosis and autophagy via miRNA-181b. 35 One of the studies included in this review reported that compared with controls, miRNA-181b was up-regulated in the cryptorchidism group. 16 We suggest that dysregulation of miRNA–181b may be associated with spermatogonial apoptosis.
A mutual regulation between miR-142-3p and TUG1 has been reported in some diseases, such as bladder cancer, 36 ulcerative colitis, 37 and septic acute kidney injury. 38 Interestingly, miR-142-3p was found to be significantly upregulated in human cryptorchidism tissues compared with controls. 14 Therefore, we hypothesise that in cryptorchidism, downregulated TUG1 results in the downregulation of the miR-142-3p targeted gene.
The most enriched GO terms under ‘cellular components’ were ‘endosome’ and ‘synapse’, and indicated a cell-cell contact role mediated by the miRNAs. A previous study found that the endosome marker, RAB11A, was co-distributed with nectin 2 (located in the Sertoli cell plasma membrane) at junctions, and indicated that some of the internalized junction proteins might be recycled to form junctions with the next generation of spermatids. 39 This research is consistent with our results and confirms a possible cell-cell contact role mediated by these miRNAs through exosomes.
A KEGG pathway analysis indicated that the dysregulated miRNAs were associated with several pathways including autophagy. It has been reported that abnormal autophagy can cause a variety of diseases, including testicular dysfunction. 40 Moreover, Sertoli cell function is vital in spermatogenesis and autophagy has been shown to affect Sertoli cells by regulating the production of inflammatory factors and apoptosis levels. 41 In addition, Notch, AMPK, Ras, and p53 signalling pathways have been found to be associated with spermatogonial apoptosis and dysfunction in spermatogenesis, which is consistent with our results. 19,20,33,42–44
This review had several limitations. For example, we chose the phenotype ‘cryptorchidism’ rather than ‘spermatogenesis’ which limited our literature search. In addition, we analysed a limited number of miRNAs validated by qRT-PCR, which may have underestimated the effect of other miRNAs involved in cryptorchidism.
Although the aetiology of cryptorchidism is unclear and may be affected by many factors, this systematic review identified 21 significantly dysregulated miRNAs in testicular tissues from animal models or patients with UDT. Further research is warranted to explore the potential of these dysregulated miRNAs as biomarkers or therapeutic targets.
Supplemental Material
sj-pdf-1-imr-10.1177_0300060521999950 - Supplemental material for Exploring dysregulated miRNAs in cryptorchidism: a systematic review
Supplemental material, sj-pdf-1-imr-10.1177_0300060521999950 for Exploring dysregulated miRNAs in cryptorchidism: a systematic review by Hongshuai Jia and Chunsheng Hao in Journal of International Medical Research
Footnotes
Declaration of conflicting interests
The authors declare that there are no conflicts of interest.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the Key Medical Specialty Development Plan of Beijing Hospital Administration (Grant No. ZYLX201825), and the Special Fund of the Paediatric Medical Coordinated Development Centre of Beijing Hospitals Authority (Grant No. XTYB201823).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.