
Abstract
Objective:
To investigate the role of extracellular-regulated protein kinase (ERK)1/2 and phosphorylated (p)-ERK1/2 in the pathogenesis of female stress urinary incontinence (SUI).
Methods:
Anterior vaginal wall tissue was collected from women with SUI and control subjects. Immunohistochemistry and Western blotting were performed for p-ERK1/2. Primary vaginal fibroblast cultures were incubated in the presence or absence of PD98059 (an inhibitor of ERK kinase) and levels of collagen I and III mRNA and protein were examined by quantitative reverse transcription–polymerase chain reaction and Western blot, respectively.
Results:
Levels of p-ERK1/2 were significantly lower in vaginal wall tissue from patients with SUI (n = 10) compared with controls (n = 10). PD98059 treatment significantly reduced levels of collagen I and III mRNA and protein.
Conclusions:
Female SUI is associated with reduced levels of p-ERK1/2 compared with controls, and inhibition of the ERK1/2 signalling pathway inhibits collagen type I and III synthesis in vaginal wall fibroblasts.
Keywords
Introduction
Stress urinary incontinence (SUI) is defined as the involuntary loss of urine that occurs when intra-abdominal pressure exceeds urethral pressure, for example during coughing, sneezing or physical exertion. It is a common condition in women, with an incidence of 18.9% in those aged >20 years in China, 1 and is a distressing problem with a profound impact on health-related quality of life.
Although the aetiology of SUI is unclear, urethral hypermobility and reduced periurethral connective tissue support (usually due to damage to functional extracellular matrix [ECM] in connective tissue) are considered important pathophysiological changes. 2 The functionality of ECM is maintained via the secretion of proteases and growth factors by connective tissue cells that modulate ECM remodelling (ECM metabolism). 3 Imbalances in ECM metabolism may therefore weaken the periurethral support structure, leading to a failure of pelvic support and increasing urethral hypermobility and involuntary urine leakage.
The ECM is a complex mixture of long chain proteins including collagen, elastin and proteoglycans. Collagen types I and III are the main structural components of ECM in periurethral support tissues, 3 which include the anterior vaginal wall, pelvic fascia and ligaments. Collagen levels are reduced in the pelvic fascia 4 and anterior vaginal wall 5 in women with SUI, implicating abnormal collagen metabolism in the pathogenesis of SUI. Collagen is mainly synthesized by fibroblasts, which also secrete matrix metalloproteinases and tissue inhibitors of matrix metalloproteinases that regulate collagen degradation. 6 There are limited data regarding the role of fibroblasts in collagen metabolism in SUI, however.
Mitogen-activated protein kinases (MAPKs) are common intracellular signalling molecules that are involved in a broad range of cellular activities including meiosis, mitosis and postmitotic functions, and are responsible for controlling cell proliferation, differentiation and survival in eukaryotes. 7 Extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK) and p38 are three major subfamilies of vertebrate MAPKs. Mammalian ERK has two main isoforms, ERK1 and ERK2, which play a pivotal role in a wide range of cellular activities in response to extracellular stimuli including mitogens, cytokines and growth factors. 7 In addition, the ERK1/2 signalling pathway is involved in the modulation of collagen metabolism in renal, 8 cardiac, 9 systemic sclerosis, 10 gingival, 11 dermal 12 and pulmonary fibroblasts. 13 However, the association between ERK1/2 in periurethral support tissue fibroblasts and SUI remains unclear.
The present study examined levels of phosphorylated (p)ERK1/2 (and therefore the activation of the ERK1/2 pathway) in the anterior vaginal wall of patients with SUI, and investigated the role of the ERK1/2 pathway in type I and III collagen synthesis by anterior vaginal fibroblasts in vitro, in order to elucidate the potential link between the ERK1/2 pathway and SUI pathogenesis.
Patients and methods
Study population
Women diagnosed with SUI according to the recommendations of International Continence Society (ICS) 14 who underwent tension-free vaginal tape surgery at the Department of Gynaecology, Women’s Hospital, Zhejiang University School of Medicine, Zhejiang, China between January 2007 and December 2010 were recruited to the study. Subjects without SUI or pelvic organ prolapse (POP) who were undergoing intravaginal cystectomy for vaginal wall cyst or radical hysterectomy for stage I cervical cancer were recruited as controls. Criteria for exclusion from the control group were: hormone replacement therapy within the previous 3 months; signs of urinary infection; oestrogen-related disease (endometriosis, myoma or functional ovarian tumour); clear clinical evidence of POP (≥grade 2); and urge incontinence. All participants were diagnosed via a combination of medical history, gynaecological examination, urinary stress test, ultrasonography and urodynamic examination, including POP-Q test. 15
Biopsy samples of the anterior vaginal wall were taken 1–2 cm from the uterine cervix, and included mucosa, submucosa, connective tissue and smooth muscle. A portion of tissue was fixed in neutral buffered formalin and embedded in paraffin for immunohistochemical analysis, and the remainder was frozen immediately at −80℃ for Western blot analysis.
The study was approved by the Institutional Review Committee of Zhejiang University School of Medicine, and all study participants provided written informed consent prior to biopsy.
Phospho-ERK1/2 immunohistochemistry
Paraffin wax-embedded vaginal wall tissue was cut into serial 4 -µm sections. Antigen retrieval was carried out by heating in 0.01 M citrate buffer (pH 6.0) at 95℃ for 40 min. Sections were then incubated with rabbit antihuman p-ERK1/2 (1 : 100 dilution; Cell Signaling Techology, Danvers. MA, USA) for 1 h at 25℃, then washed three times with 0.01 M phosphate buffered saline (PBS; pH 7.4) at 25℃ for 3 min each wash. Immunohistochemical staining was visualized using a ChemMate EnVision™ kit HRP/DAB (Dako, Glostrup, Denmark), according to the manufacturer’s instructions. Sections from known ERK1/2-positive breast cancers were used as a positive control, and either normal rabbit serum or the absence of primary antibody was used as negative control.
Phospho-ERK1/2 Western blotting
Total protein was extracted from vaginal tissue using T-PER Tissue Protein Extraction Reagent, and quantified using a BCA Protein Assay Kit (both Pierce, Rockford, IL, USA). Proteins (60 µg per sample) were separated by sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS–PAGE) and transferred to hybond-C pure nitrocellulose membranes (Amersham, Little Chalfont, UK). Membranes were blocked in 5% nonfat milk for 1 h at 25℃, then washed three times with 0.01 M Tris-buffered saline–Tween 20 (TBST; pH 7.5) at 25℃ (5 min each wash). After washing, membranes were incubated with rabbit antihuman ERK1/2 (1 : 1000 dilution; Cell Signaling Technology), rabbit antihuman p-ERK1/2 (1 : 1000 dilution; Cell Signaling Technology) or rabbit antihuman β-actin (1 : 2000 dilution; Cell Signaling Technology) for 12 h at 4℃, washed three times with TBST at 25℃ (5 min each wash), incubated with horseradish peroxidase (HRP)-conjugated goat antirabbit IgG (1 : 10000 dilution; Pierce) for 1 h at 25℃, then washed again three times with TBST at 25℃ (5 min each wash). Protein complexes were visualized using a chemiluminescence system (Amersham) and Kodak X-OMAT film (Eastman Kodak, Rochester, NY, USA). The relative band densities were analysed using NIH Image (version 1.62; available at http://rsb.info.nih.gov/nih-image/). ERK1/2 and p-ERK1/2 were quantified by reference to β-actin.
Fibroblast cell culture
Fibroblasts were prepared from fresh vaginal wall tissue of a single control subject as described, 16 and cultured in Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum, at 37℃ in a humidified atmosphere of 5% carbon dioxide in air. Fibroblasts were identified according to morphology and the presence of vimentin. For identification of vimentin, cultured cells were fixed in formaldehyde for 20 min then incubated with 3% hydrogen peroxide at 25℃ for 10 min, followed by mouse anti-human vimentin (1 : 1000 dilution; Boster Biological Engineering, Wuhan, China) overnight at 4℃. Cells were washed three times at 25℃ with PBS (0.01 M, pH 7.4), incubated with rabbit antimouse immunoglobulin G (1 : 1000 dilution; Boster Biological Engineering) at 37℃ for 30 min, then washed again three times at 25℃ with PBS. Staining was visualized by incubation with streptavidin fluorescein at 37℃ for 30 min. Primary cell cultures in which >95% of the cells were positively stained for vimentin were used in further experiments.
Real-time RT–PCR
Fibroblasts (1 × 104 in 0.5 ml medium) were seeded in 24-well culture plates and cultured in the presence or absence of 10 µmol/l PD98059 (an ERK kinase inhibitor; Cell Signaling Technology) for 48 h. 17
Total RNA was extracted with a Total RNA Isolation Kit (Haogene Biotech, Hangzhou, China) according to the manufacturer’s instructions and quantified using spectrophotometry (NANOdrop 2000; Thermo Fisher Scientific, Rockford, IL, USA). RNA (500 ng) was reverse transcribed in a 200 -μl volume containing 1 µl oligo-dT primer, 1 μl dNTP mixture, 25 U RNase inhibitor and 200 U SuperScript™ II reverse transcriptase (Invitrogen, Carslbad, CA, USA
Collagen I and III Western blotting
Fibroblasts (1 × 104 in 0.5 ml medium) were seeded in 24-well culture plates and cultured in the presence or absence of 10 µmol/l PD98059 (Cell Signaling Technology) for 48 h. 17 Total protein was extracted and quantified as described earlier. SDS–PAGE and Western blotting were carried out as described earlier, using rabbit antihuman collagen I (1 : 1000 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit antihuman collagen III (1 : 1000 dilution; Santa Cruz Biotechnology) or rabbit antihuman β-actin (1 : 1000 dilution; Santa Cruz Biotechnology) primary antibodies and HRP-conjugated goat antirabbit IgG secondary antibody (1 : 10000 dilution; Pierce).
Statistical analyses
Data were expressed as mean ± SD and analysed with Student’s t-test. Statistical analyses were performed using SPSS® version 13.0 (SPSS Inc., Chicago, IL, USA) for Windows®. P-values < 0.05 were considered statistically significant.
Results
The study included 10 patients with SUI (mean age 50.60 ± 5.87 years; parity 1.60 ± 0.70; body mass index [BMI] 21.71 ± 4.13 kg/m2) and 10 control subjects (mean age 45.70 ± 5.10 years; parity 1.80 ± 0.79; BMI 19.68 ± 5.71). There were no significant between-group differences in age, parity or BMI.
Representative light photomicrographs of phospho-ERK1/2 immunohistochemical staining are shown in Figure 1. Phospho-ERK1/2 was localized to the nuclei of vascular endothelia, smooth muscle cells and fibroblasts.
Representative light photomicrographs of immunohistochemical staining for phosphorylated extracellular signal-regulated kinase 1/2 in anterior vaginal wall tissue from: (A, C) women with stress urinary incontinence (SUI); and (B, D) women without SUI. Staining is localized to the nuclei of vascular endothelia, smooth muscle cells and fibroblasts. The colour version of this figure is available at: http://imr.sagepub.com.
Western blotting revealed significantly lower p-ERK1 and p-ERK2 levels in anterior vaginal wall tissue from women with SUI compared with those without SUI (P < 0.05 for each comparison, Figure 2). There were no significant between-group differences in ERK1/2 levels.
(A) Representative Western blots for extracellular signal-regulated kinase (ERK)1/2 and phosphorylated (p)-ERK1/2 in anterior vaginal wall tissue from women with stress urinary incontinence (SUI; n = 10, lanes 1–4); and women without SUI (n = 10, lanes 5–8). (B) Mean ± SD levels of p-ERK1 and p-ERK2, semi-quantified by reference to β-actin. *P < 0.05, Student’s t-test.
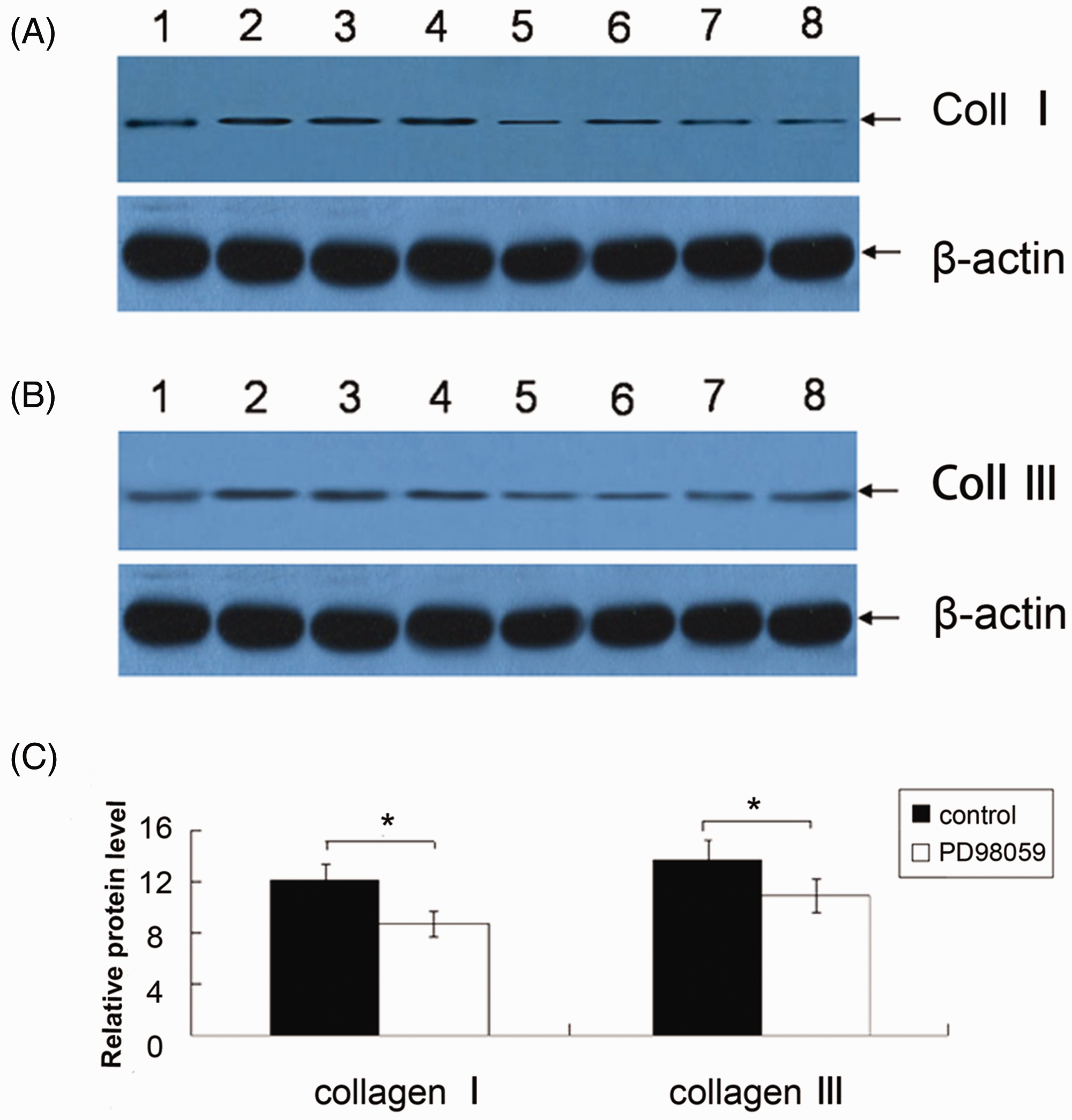
Incubation of cultured vaginal fibroblasts with PD98059 significantly reduced levels of collagen I and III mRNA and protein compared with control cultures (P < 0.05 for each comparison; Figures 3 and 4).
Quantitative real-time reverse transcription–polymerase chain reaction analysis of collagen I and collagen III mRNA in primary cultures of human vaginal fibroblasts incubated with or without 10 µmol/l PD98059 (an extracellular-regulated kinase kinase inhibitor) for 48 h. Data are mean ± SD of six experiments; *P < 0.05, Student’s t-test. Representative Western blots for (A) collagen I and (B) collagen III in primary cultures of human vaginal fibroblasts incubated without (lanes 1–4) or with (lanes 5–8) 10 µmol/l PD98059 (an inhibitor of extracellular-regulated kinase kinase) for 48 h. (C) Levels of collagen I and collagen III, semi-quantified by reference to β-actin. Data are mean ± SD of six experiments; *P < 0.05, Student’s t-test.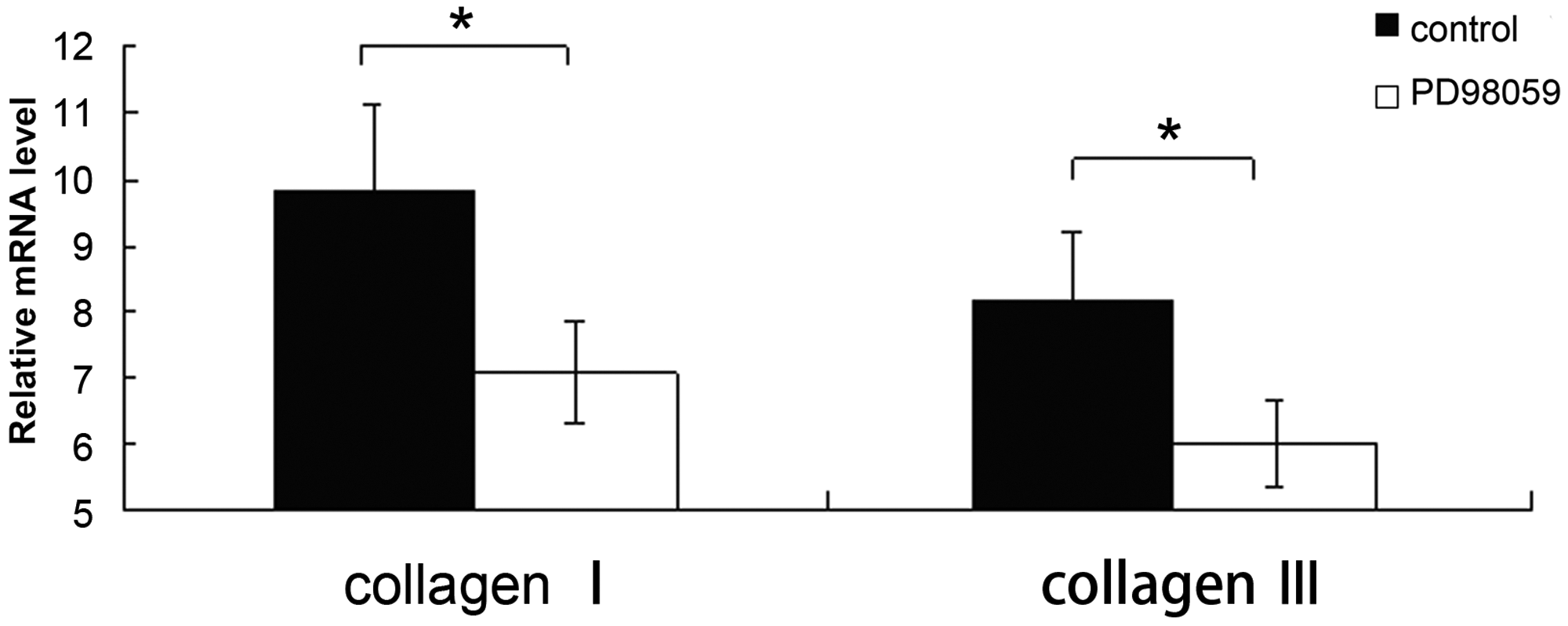
Discussion
The present study found significantly lower levels of p-ERK1/2 in the anterior vaginal wall of women with SUI compared with control women. In addition, inhibition of ERK kinase using PD98059 significantly reduced collagen I and III mRNA and protein levels in primary human vaginal fibroblast cultures. These findings suggest that the ERK signalling pathway may play a role in the pathogenesis of SUI.
Both ERK1 and ERK2 are widely expressed in human tissue such as lung, 13 kidney, 8 heart 9 and brain, 18 and the ERK1/2 signalling pathway is involved in the development of human diseases, including cancer. 19 The role of ERK1/2 in pelvic support tissues and the pathology of female SUI remain unknown, however. The present study used anterior vaginal wall tissue since this is an important part of the periurethral support structure that has been examined by others.20,21 Immunohistochemical staining of vaginal wall fibroblasts in the current study revealed high levels of ERK1/2. Taken together with the reduction in p-ERK1/2 protein in women with SUI compared with controls, this indicates that abnormalities in the ERK1/2 signalling pathway may play a role in the pathogenesis of SUI. The Ras/Raf/MEK/ERK signalling pathway is known to play key roles in the transmission of proliferative signals from membrane-bound receptors, 22 and phosphorylation of ERK1/2 regulates the activities of several transcription factors (including c-myc and cyclic AMP response element-binding protein [CREB]). This leads to alterations in the transcription of genes that are important for fibroblast proliferation and differentiation. 23 The ERK1/2 present in vaginal wall fibroblasts may therefore be important for fibroblast activation.
Abnormal collagen metabolism in SUI is well documented. 5 Others have explored the role of ERK signalling in fibroblast collagen metabolism,8–13,17 but there are limited data regarding the involvement of ERK in collagen metabolism in pelvic support tissues. Collagen types I and III are important in maintaining tissue tensile strength and the mechanical stability of pelvic support tissues. 24 The ERK kinase inhibitor PD98059 binds to inactive ERK1/2 and prevents its activation by c-Raf, thereby inhibiting phosphorylation. 25 Levels of collagen I and III mRNA and protein in primary human vaginal fibroblasts were significantly reduced by PD98059 treatment in the present study, suggesting that ERK1/2 signalling is crucial for collagen I and III synthesis in the pelvic floor. The activity of the ERK1/2 pathway is regulated by many different stimuli, including growth factors, cytokines, viral infection, transforming agents and carcinogens, which can then influence collagen synthesis.26–28
In conclusion, female SUI is associated with reduced levels of p-ERK1/2 compared with controls, and inhibition of the ERK1/2 signalling pathway inhibits collagen types I and III synthesis in vaginal wall fibroblasts. It is possible that abnormalities in ERK1/2 activation may be involved in the pathogenesis of female SUI. Further, larger scale investigations are required to fully elucidate the influence of ERK1/2 on collagen metabolism in SUI.
Footnotes
Declaration of conflicting interest
The authors declare that there are no conflicts of interest.
Funding
This study was supported by grants from the National Natural Science Foundation of China (81200429), the Zhejiang Provincial Natural Science Foundation of China (LY13H090005), the Qianjiang Talents Project of Science and Technology Department of Zhejiang Province (2010R10069), Zhejiang Provincial Chinese Medicine Research Foundation (2010ZA066, 2012ZA091) and Zhejiang Provincial Education Committee Research Foundation (Y200909135, Y201019012). The content is solely the responsibility of the authors and does not necessarily represent the official views of the grant-awarding bodies.