
Abstract
Objective
To investigate the role of semaphorin 6B in gastric cancer invasion and metastasis.
Methods
Immunohistochemistry for semaphorin 6B was performed on gastric cancer tumour tissue samples in this retrospective study. Levels of semaphorin 6B protein and mRNA were determined in gastric cancer cell lines by Western blotting and quantitative reverse transcription–polymerase chain reaction, respectively. The human gastric cancer cell line SGC-7901 was transfected with small interfering RNA targeting semaphorin 6B; effects on cell adhesion, migration and invasion were determined by cell adhesion assay, transwell chamber migration assay and wound healing assay, respectively.
Results
Tumour tissue samples from 220 patients were analysed. In vivo, semaphorin 6B immunopositivity correlated with tumour differentiation, lymph node metastasis and distant metastasis but not patient age, sex or tumour stage. Semaphorin 6B gene silencing significantly suppressed adhesion, migration and invasion of gastric cancer cells in vitro.
Conclusions
Semaphorin 6B is related to tumour differentiation and metastasis in vivo, and tumour cell migration, adhesion and invasion in vitro. Semaphorin 6B may represent a reliable biomarker for diagnosis, evaluation and gene-targeted therapy of gastric cancer.
Keywords
Introduction
Semaphorin 6B is a member of the semaphorin class of proteins; these are involved in axonal guidance and play vital roles in central nervous system development.1–4 Semaphorin expression is closely related to tumour invasion and metastasis in several human cancers,5–10 but the association between semaphorin 6B expression and pathological features of gastric cancer is still poorly understood. In addition, the function of semaphorin 6B in the tumorigenesis and development of gastric cancer remains unknown.
The objectives of this study were to determine the presence of semaphorin 6B in gastric cancer tissue and to investigate the associations of semaphorin 6B with tumour stage, differentiation and metastasis. In addition, a gastric tumour cell line with decreased semaphorin B levels was established by small interfering RNA (siRNA) technology, in order to explore the function of semaphorin 6B in gastric cancer.
Patients and methods
Study population
Tumour tissue specimens were retrospectively collected from patients with primary gastric cancer undergoing treatment at the Department of Surgery, Xiangya Hospital, Central South University, Changsha City, China between January 2005 and April 2010. Patients who had received previous radiotherapy or neoadjuvant therapy were excluded from the study; there were no other specific inclusion or exclusion criteria. Tumours were staged according to the stomach cancer staging guidelines of the American Joint Committee on Cancer. 11
All patients provided written informed consent for the use of tissue specimens, and the study was approved by the Institutional Review Board for Human Research, at the National Hepatobiliary and Enteric Surgery Research Centre of Central South University, Changsha, Hunan, China.
Immunohistochemistry
Tissue samples were fixed in 10% formalin, embedded in paraffin wax and cut into 4 -µm thick sections. Immunohistochemical staining was performed using the avidin–biotin–peroxidase (ABC) method.12,13 Briefly, after deparaffinizing in xylene and rehydrating in ethanol, sections were heated in citrate buffer (0.01 M, pH 6.5) at 120°C for 10 min, then incubated with mouse antihuman semaphorin 6B (1 : 500 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA) or mouse antihuman β-actin (1 : 200 dilution; Santa Cruz Biotechnology) overnight at 4–8°C. Sections were washed four times with 0.1 M phosphate buffered saline (PBS; pH 7.4), then incubated with biotinylated goat antimouse immunoglobulin G (1 : 200 dilution; Vector Laboratories, Burlingame, CA, USA) at room temperature for 90 min. Slides were washed three times with PBS and immunoreactive staining was visualized using 0.5 mg/ml 3,3′-diaminobenzidine tetrahydrochloride and 0.03% hydrogen peroxide. Bovine serum albumin was used in place of primary antibody, as a negative control.
Ten high-power fields (×200 magnification) including the centre of the tumour and the invasive front, were examined by light microscopy; 100 tumour cells per field were evaluated to determine the presence of the distinct brown staining of the nucleus. Semaphorin 6B immunoreactivity was determined according to the mean proportion of positive cells: negative (<5% stained); or positive (≥5% stained).14–17
Cell lines
The human gastric cancer cell lines AGS, SNU1, SGC-7901 and HGEC were obtained from American Type Culture Collection (Manassas, VA, USA); MKN45 was obtained from Japanese Collection of Research Bioresources (Osaka, Japan). Cells were maintained under standard conditions in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum (FBS) and 100 µg/ml penicillin/streptomycin.
Western blotting
Total protein was extracted from cell lines by lysis in 0.1 M PBS (pH 7.4) containing 0.5% sodium deoxycholate, 1% Triton™ X-100 and 0.1% sodium dodecyl sulphate (SDS). Protein was quantified using bicinchoninic acid assay (Pierce, Rockford, IL, USA), and 100 µg protein per sample was dissolved in sample buffer (0.6 ml 1M Tris-HCl [pH 8.8], 2 ml 10% SDS, 5 ml 50% glycerol, 0.5 ml 2-mercaptoethanol, 1 ml 1% bromophenol blue and 40.9 ml sterilized water), separated via 10% SDS–polyacrylamide gel electrophoresis, and transferred to polyvinylidene difluoride membranes (Millipore, Bedford, IN, USA). Membranes were blocked with 5% nonfat dry milk in 1 × Tris buffered saline (pH 7.6) containing 0.1% Tween 20 (TBST) for 1 h at room temperature, then incubated with mouse antihuman semaphorin 6B (1 : 200 dilution; Santa Cruz Biotechnology) or mouse antihuman β-actin (1 : 500 dilution; Santa Cruz Biotechnology) at 4–8°C overnight. Membranes were washed five times in TBST (4 min each wash) then incubated in horseradish peroxidise-conjugated goat antimouse antibody (1 : 1000 dilution; Amersham Life Science, Buckinghamshire, UK) for 1 h at room temperature. Membranes were washed five times in TBST and immunoreactive proteins were visualized using an enhanced chemiluminescence kit (Amersham Life Science).
Real-time RT–PCR
Total RNA was extracted from cultured cells (5 × 105 cells/well) using the Trizol® isolation kit (Invitrogen, Austin, TX, USA) and reverse transcribed with AMV Reverse Transcriptase (HC) (Promega, Madison, WI, USA), according to the manufacturers’ instructions. Real-time quantitative polymerase chain reaction (PCR) was performed using SYBR® Select Master Mix and a 7900HT sequence detection system (both from Applied Biosystems, Foster City, CA, USA). Each reaction mixture contained 300 nM each primer (see below), 2.5 µl 2 × SYBR® Green master mix (Applied Biosystems), and 0.1 µg cDNA (in 1 µl), to a final volume of 5 µl. Primer sequences were: semaphorin 6B (SEMA6B), forward 5′-GCCATGCAGACCCCGCGAGC-3′ and reverse 5′-CCACGTGGCATGCATGTCAG-3′; glucose-6-phosphate dehydrogenase (G6PDH), forward 5′-TGGACCTGACCTACGGCAACAGATA-3′ and reverse 5′-GCCCTCATACTGGAAACCC-3′.3,18 The cycling programme involved preliminary denaturation at 50°C for 2 min then 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 15 s, annealing 60°C for 60 s, elongation at 60°C for 60 s, and a final dissociation step at 95°C for 15 s. All reactions were performed in triplicate wells of a 384-well microtitre plate. PCR products were quantitated relative to G6PDH levels.
Construction of siRNA plasmid expression vector
Double-stranded oligonucleotides with complementary sequences encoding short hairpin RNA (shRNA) were obtained from Genesil Biotechnology Company (Wuhan, Hubei, China). The complementary sequence comprised two reversed repeated sequences with nine inserted sequences (TTCAAGAGA, AAGAACTCT), with recognition sites for BamHI and HindIII on both sides to facilitate ligation to the pSilencer™ 2.1 plasmid (Invitrogen). The targeting site of the transcribed oligonucleotide siRNA was nucleotides 76–84 of SEMA6B (GenBank® no. NM-020241). The negative control was a transcribed siRNA sequence with no homology to any human gene sequence.
To confirm the sequence of the siRNA, ligation was performed according to the manufacturer’s instructions and the recombinant pSilencer™ plasmid was transformed into Escherichia coli DH5a cells. Ampicillin-resistant colonies were cultured on solid luria broth medium at 37°C overnight on a rocking bed, and the sequence of the inserted 76 bp siRNA was verified by electrophoresis of the recombinant semaphorin 6B siRNA plasmid. 19
Next, SGC-7901 cells were seeded in six-well plates (1 × 105/well in 100 µl) and transfected with 100 nM pSilencer™ plasmid, containing either semaphorin 6B siRNA or control siRNA when confluence reached 90%. Transfection was performed using X-tremeGene siRNA transfection reagent (Roche, Indianapolis, IN, USA), according to the manufacturer’s instructions. At 48 h after transfection, cells were passaged 1 : 10, then cultured for 14 days in DMEM containing 600 µg/ml G418. The resulting cell lines were designated SGC-7901/Silence+ and SGC-7901/Silence–.
Adhesion assay
Cells (SGC-7901/Silence+, SGC-7901/Silence– or SGC-7901) were seeded in 96-well plates (3 × 104 cells/well in 50 µl) that were either uncoated or coated with fibronectin or matrix agarose. Cells were allowed to settle for 5 min; nonadherent cells were removed by aspiration and adherent cells were fixed with 4% paraformaldehyde, for 10 min. Fixed cells were incubated with 0.04% (v/v) Coomassie brilliant blue G-250 (in 25% ethanol, 12% acetic acid in double-distilled water) at 22°C for 45 min, then washed three times with wash solution (10% ethanol, 5% acetic acid in double distilled water). Cell-bound stain was resolubilized by incubation with 1 M potassium acetate (100 µl/well) for 45 min at 22°C and detected by measuring optical density at 595 nm. The percentage of adherent cells was determined by comparison with wells from which cells were not aspirated before fixation (maximum total cells/well, 3 × 104).
Migration assay
A migration assay was performed using a transwell chamber system with 8-mm porous filters (24-well format; Millipore, Billerica, MA, USA), in which cellular components are grown without direct cell-to-cell contact. SGC-7901/Silence+, SGC-7901/Silence– and SGC-7901 cells were diluted to 1 × 106 cells/ml in DMEM containing 0.5% FBS, and 50 000 cells/well were seeded into the upper compartment. The lower compartment of the chamber was filled with 1 ml DMEM containing 50 mg/ml hyaluronic acid. The chambers were incubated at 37°C for 36 h then removed from the plate. Cells that had migrated to the upper surface of the membrane were removed with cotton swabs; filters were fixed in 95% ethanol for 30 min then washed twice in distilled water. Filters were stained with 0.2% crystal violet for 20 min, rinsed twice with distilled water and examined via light microscopy. Cells were counted in five low-power fields (×40 magnification) and the cell invasion score was expressed as a percentage of untreated control cells (parental SGC-7901) cells.
Scratch wound closure assay
Cell migration was determined using a scratch wound closure assay. 20 SGC-7901/Silence+, SGC-7901/Silence– and SGC-7901 cells (105 cells/well in 100 µl) were cultured in 48-well plates, and a portion of the cell monolayer was scraped away with a sterile disposable rubber policeman. The remaining cells were washed with DMEM and incubated for a further 24 h. Cell migration into the denuded area was quantified with a computer-assisted microscope (MIC00266; Zeiss, Aalen, Germany).
Statistical analyses
Data were expressed as mean ± SEM and compared using one-way analysis of variance. Statistical analyses were performed with SPSS® software, version 18.0 (SPSS Inc., Chicago, IL, USA) for Windows®. A P-value< 0.05 was considered statistically significant.
Results
Correlations between semaphorin 6B immunopositivity and demographic and clinicopathological characteristics of patients with primary gastric cancer (n = 220).

Data presented as n of patients.
One-way analysis of variance.
NS, not statistically significant (P ≥ 0.05).
Levels of semaphorin 6B mRNA were significantly higher in SGC-7901 cells (4.76 ± 0.25) than in MKN45 (2.23 ± 0.21; P = 0.031), SNU1 (2.12 ± 0.12; P = 0.026), AGS (1.98 ± 0.09; P = 0.014) or HGEC cells (0.95 ± 0.14; P = 0.008). In addition, semaphorin 6B protein levels were visibly higher in SGC-7901 cells than in other cell lines studied (Figure 1). As a result, SGC-7901 cells were selected for subsequent assays.
Detection of semaphorin 6B protein in five human gastric carcinoma cell lines (HGEC, AGS, SNU1, MKN45 and SG-7901) by Western blotting. β-actin used as the protein loading control.
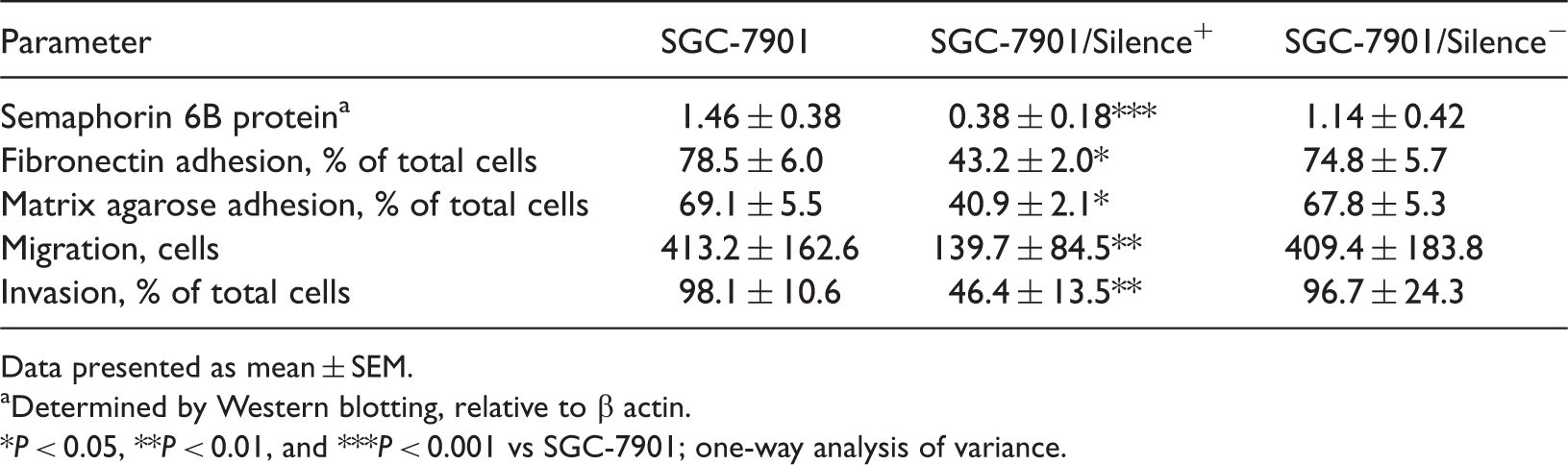
Transfection of SGC-7901 cells with semaphorin 6B siRNA significantly reduced semaphorin 6B protein levels, compared with those observed in parental SGC-7901 cells (P < 0.001; Table 2, Figure 2), and significantly reduced adherence to fibronectin and matrix agarose compared with parental cells (P = 0.04 and P = 0.01, respectively; Table 2). In addition, transwell cell migration and scratch closure migration were also significantly reduced by semaphorin 6B siRNA transfection (P < 0.01 for both comparisons; Table 2).
Western blotting detection of semaphorin 6B protein in (A) the human gastric carcinoma cell line SGC-7901, SGC-7901 cells transfected with pSilencer™ 2.1 plasmid (Invitrogen, Austin, TX, USA) encoding (B) control small interfering RNA (siRNA) (SGC-7901/Silence–) and (C) siRNA targeting semaphorin 6B expression (SGC-7901/Silence+). β-actin used as the protein loading control. Semaphorin 6B protein levels, adhesion, migration and invasion in cells of the human gastric carcinoma cell line SGC-7901 transfected with pSilencer™ 2.1 plasmid (Invitrogen, Austin, TX, USA) encoding control small interfering RNA targeting semaphorin 6B expression (SGC-7901/Silence+) or control siRNA (SGC-7901/Silence–), and parental SGC-7901 cells. Data presented as mean ± SEM. Determined by Western blotting, relative to β actin. P < 0.05, **P < 0.01, and ***P < 0.001 vs SGC-7901; one-way analysis of variance.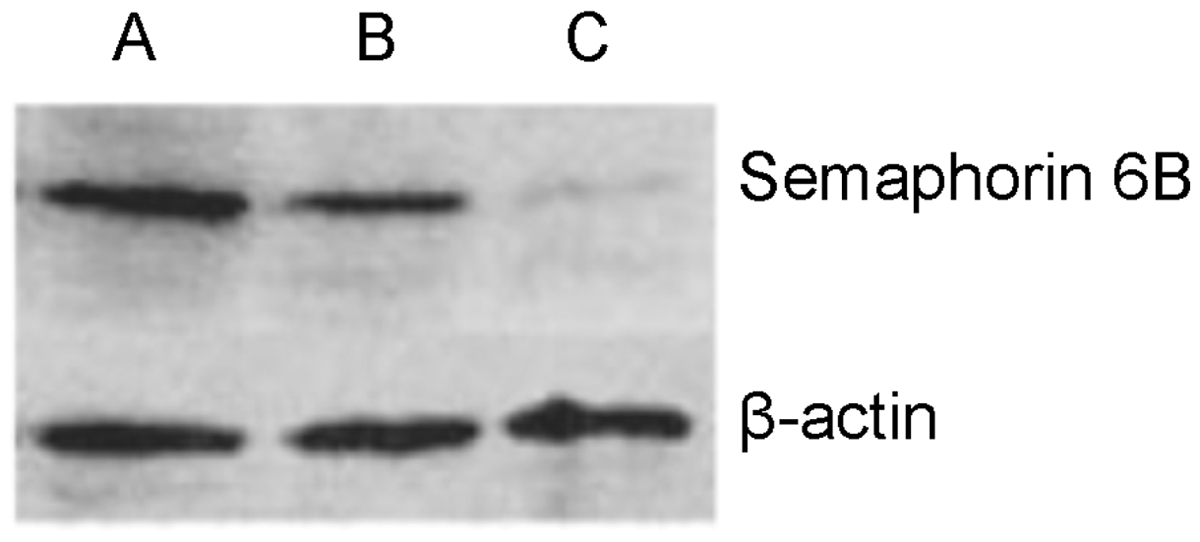
Discussion
Semaphorins are a group of axonal growth cone guidance proteins, with over 30 members identified in eight major subfamilies.21–23 The semaphorin proteins share a similar structure, defined by a highly conserved cysteine-rich sema domain at the
Cancer invasion and metastasis are closely related to adhesion, migration and invasion of cancer cells. 37 In the present study, inhibition of semaphorin 6B expression via RNA interference significantly inhibited the migration, adhesion and invasion abilities of SGC-7901 gastric cancer cells in vitro. These findings indicate that semaphorin 6B may promote gastric cancer invasion and metastasis by upregulation of extracellular matrix adhesion, migration and invasion, but the mechanism remains to be elucidated.
In conclusion, the presence of semaphorin 6B is closely related to tumour differentiation and metastasis in vivo, and to tumour cell migration, adhesion and invasion in vitro. Semaphorin 6B may represent a reliable biomarker for the clinical diagnosis, evaluation and gene-targeted therapy for gastric cancer.
Footnotes
Declaration of conflicting interest
The authors had no conflicts of interest to declare in relation to this article.
Funding
This study was supported by the Fundamental Research Fund for the Central Universities of Central South University, Changsha City, China.