
Abstract
Ischaemic stroke results in the formation of a cerebral infarction bordered by an ischaemic penumbra. Characterising the proteins within the ischaemic penumbra may identify neuro-protective targets and novel circulating markers to improve patient care. This review assessed data from studies using proteomic platforms to compare ischaemic penumbra tissues to controls following experimental stroke in animal models. Proteins reported to differ significantly between penumbra and control tissues were analysed in silico to identify protein-protein interactions and over-represented pathways. Sixteen studies using rat (n = 12), mouse (n = 2) or primate (n = 2) models were included. Heterogeneity in the design of the studies and definition of the penumbra were observed. Analyses showed high abundance of p53 in the penumbra within 24 hours of permanent ischaemic stroke and was implicated in driving apoptosis, cell cycle progression, and ATM- MAPK- and p53- signalling. Between 1 and 7 days after stroke there were changes in the abundance of proteins involved in the complement and coagulation pathways. Favourable recovery 1 month after stroke was associated with an increase in the abundance of proteins involved in wound healing. Poor recovery was associated with increases in prostaglandin signalling. Findings suggest that p53 may be a target for novel therapeutics for ischaemic stroke.
Introduction
An estimated 7.63 million ischaemic strokes occur annually which result in a cerebral infarction bordered by a region of hypo-perfused tissue known as the ischaemic penumbra.1 –6 Clot retrieval and thrombolysis which work to restore the blood supply and salvage the ischaemic penumbra traditionally need to be performed within hours of stroke onset, but may provide benefit if administered to late-presenting patients in whom viable penumbra persists.7 –10 Elucidating the biology of the ischaemic penumbra may help to identify diagnostic markers useful for identifying people who would benefit from late revascularisation and highlight novel neuro-protective therapeutic targets.11 –13 Proteomic analysis technologies enable the simultaneous identification and quantification of hundreds to thousands of proteins within a biological sample and offer great potential to improve understanding of complex disease processes. 14 Proteomics is being increasingly applied to the stroke field, however, few studies have been conducted to specifically characterise the penumbra.15,16 The current review aimed to: i) systematically analyse evidence from studies applying protein screening to analyse the ischaemic penumbra in animal models, and ii) identify and characterise the proteins consistently reported to show significant differences in abundance between the penumbra and healthy regions of the brain.
Methods
This review adhered to PRISMA guidelines and the Systematic Review Centre for Laboratory animal Experimentation statement (PROSPERO database registration number CRD42023461673).17,18
Literature searches
The keyword terms (“prote−omics” OR “protein omics” OR proteinomics OR Proteomic) AND (“penumbra” OR “peri-infarct” OR periinfarct OR “peri infarct”) were used to search the Pubmed database. The Medline (Ovid), Scopus and Web of Science were searched using the terms (“prote−omics” OR “protein omics” OR proteinomics OR Proteomic*) AND (“penumbra” OR “peri-infarct*” OR periinfarct* OR “peri infarct*”). Subject heading searches in Medline and Cinahl provided no additional articles. Retrieved articles were screened by two authors (JVM and AFT) to identify relevant studies for inclusion in the current review. Eligible studies had to use a protein screening platform to compare ischaemic penumbra tissues recovered from animal models of acute ischaemic stroke to control tissues sourced from the same animal (e.g. biopsies from the contralateral brain hemisphere), or animals undergoing sham surgeries. No restrictions on date of publication were applied. Review articles, editorials and articles published in languages other than English were excluded. Studies assessing individual pre-selected proteins, or those not measuring proteins directly within the penumbra were excluded. Studies testing novel therapies or interventions were excluded unless data comparing penumbra and control tissues recovered from animals receiving a vehicle control or sham surgery were reported. Reference lists of related reviews were also manually searched to identify potentially eligible studies not identified by keyword searches. Study authors were contacted if additional information was needed. Literature searches commenced on the 17th May 2023 and were finalised on 1st December 2023.
Data extraction and risk of bias assessment
Data extraction was independently conducted by three authors (JVM, CP, AFT) focusing on: i) Animal species, strain, age and sex, ii) Methods of ischaemic stroke induction, iii) The time between stroke onset and sample collection, v) Methods used to identify the penumbra, and vi) The choice of protein screening platform. Risk of bias was assessed using a 3-domain tool addressing study design, and the reporting of proteomic outcomes (Supplement 1). Each domain was independently scored by 3 authors with relevant expertise (domain 1: JVM, CP, AFT; domains 2 and 3: JVM, MKH, PM). The CAMARADES checklist was modified to assess experimental design, including criteria specific to the current review. 19 The reporting of protein identification and quantification methodologies was assessed using applicable criteria of the Proteomics Standards Initiative checklist (Supplement 1). 20 All studies were assessed against domain 1, domains 2 and 3 were applied where appropriate. Domain scores, and an overall score were calculated (expressed as a percentage of applicable fields to normalise between studies), scores of <50%, 50–75% or >75% denote high, moderate or low risk of bias, respectively.
In silico analysis of protein expression data
Panels of proteins dysregulated within the penumbra were characterised in silico. To be eligible for inclusion, studies had to provide details of the relative abundance of named proteins which were reported to show significant differences in expression between penumbra and control samples. Wang et al. (2022) did not provide lists of differentially abundant proteins, but did upload raw proteomic data to the ProteomeXchange repository (Accession: PXD030788). Data were downloaded and reanalysed with the Spectronaut software package (v 10.0.230605.50606, Biognosys, Switzerland) in a library-free manner to compare protein abundance between samples collected from sham-operated controls, and animals undergoing cerebral ischaemia (vehicle control group, detailed methodology provided in Supplement 2). 21 Proteins showing a ≥2-fold difference in abundance between samples (false discovery rate corrected p-value <0.05) were considered to be significantly different, and were included in the in silico analyses.
To permit data pooling, the naming of differentially expressed proteins was standardised according to gene symbol using the GeneCards database to identify aliases where needed. Some array-based papers reported data detailing the expression of post-translationally modified proteins, splice variants, or multiple isoforms of functionally related proteins which did not have a unique gene symbol and these could not be included in the analyses. Studies employing two-dimensional electrophoresis demonstrated that some differentially abundant spots contained multiple co-migrating proteins. Such data were not considered in the in silico analyses unless the protein(s) driving observed differences in abundance were identified using targeted approaches (e.g. western blotting or ELISA). Protein datasets were initially grouped according to whether they were generated from models of permanent or transient cerebral ischaemia and were subdivided into those providing data from samples collected during the early hyperacute, late hyperacute, acute, or sub-acute phases (0–≤6 hours, 6–24 hrs, 24 hrs-7 days and 7 days-3 months post stroke, respectively based on Stroke Roundtable Consortium definitions).22,23
Inter-study differences in the reporting of relative protein abundance and ambiguity regarding sample sizes for some investigations prevented formal meta-analysis. Thus proteins were reclassified according to whether they were reported to more or less abundant in the penumbra than control tissues. Those showing contradictory differences in the direction of over- or under-abundance between studies were removed. Protein panels were de-duplicated and imported into Cytoscape (v. 3.10.1) to identify protein-protein interaction networks (String database, confidence cut-off 0.8). Networks comprising at least 4 interacting proteins were analysed for functional enrichment to identify over-represented pathways. As 12 of the 14 included studies conducted in rodent models of stroke used experimental rats (the remainder used mice), network and pathway analyses were conducted using the Rattus norvegicus genome as a reference. Two studies used cynomolgus monkeys (Macaca fasicularis) for which there was no available reference genome within the Cytoscape package.24,25 Network and functional enrichment analysis of data from these studies was conducted using the human genome as a reference as the closest homologue. 24 Pathways and gene ontology categories showing a false discovery-rate corrected p-value <0.05 were considered to be significantly over-represented across the assessed proteins.
Results
Literature searches
Thirty-five potentially eligible articles were identified after excluding duplicates (Figure 1). Manual searches of reference lists of related articles identified an additional study which was considered to be likely to be eligible for inclusion. 24 The full texts of 31 articles were assessed, of which 15 were excluded, most commonly as they did not specifically assess tissues recovered from the ischaemic penumbra.26 –40 Sixteen studies fulfilled inclusion criteria.11,21,24,25,41 –52 Six included studies investigated the effect of novel interventions on penumbral protein expression, but provided datasets detailing penumbra protein expression in animals not receiving these treatments which were included.21,25,45,46,49,52
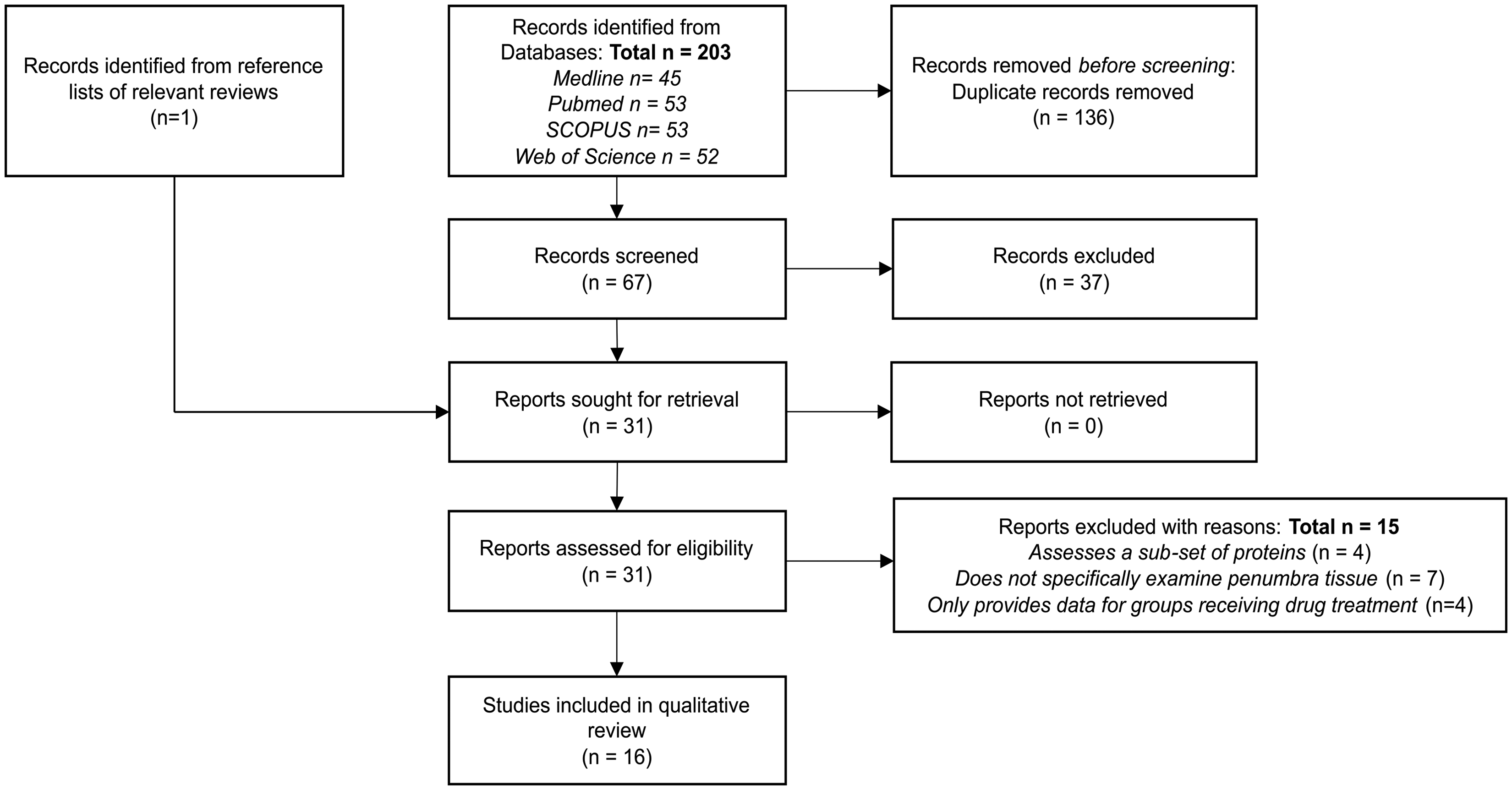
PRISMA flow diagram showing the selection of included studies.
Characteristics of included studies
Table 1 details the design of the sixteen included studies, 12 used experimental rats, two used mice, and two used cynomolgus macaques (Macaca fasicularis). Stroke was commonly induced using middle cerebral artery occlusion (MCAO),11,21,24,25,41,42,46,47,49 or photothrombosis,43 –45,48,50,51 and one study used endothelin-1 infusion. 52 Cerebral ischaemia was permanently induced in eight studies, 11,41 –45,48,50 and seven investigated temporary ischaemia (ischaemia-reperfusion).21,24,25,46,47,49,52 Yao et al. (2009) presented data from groups of animals exposed to either permanent, or transient cerebral ischemia. 51 The time between ischaemic stroke induction and brain sampling ranged from 1- hour to twenty eight days, and several studies used multiple groups to assess temporal changes in penumbral protein composition. Inter-study variations in the approaches used to identify and sample the ischaemic penumbra, choice of control tissues and protein screening platform were observed.
Characteristics of included studies.
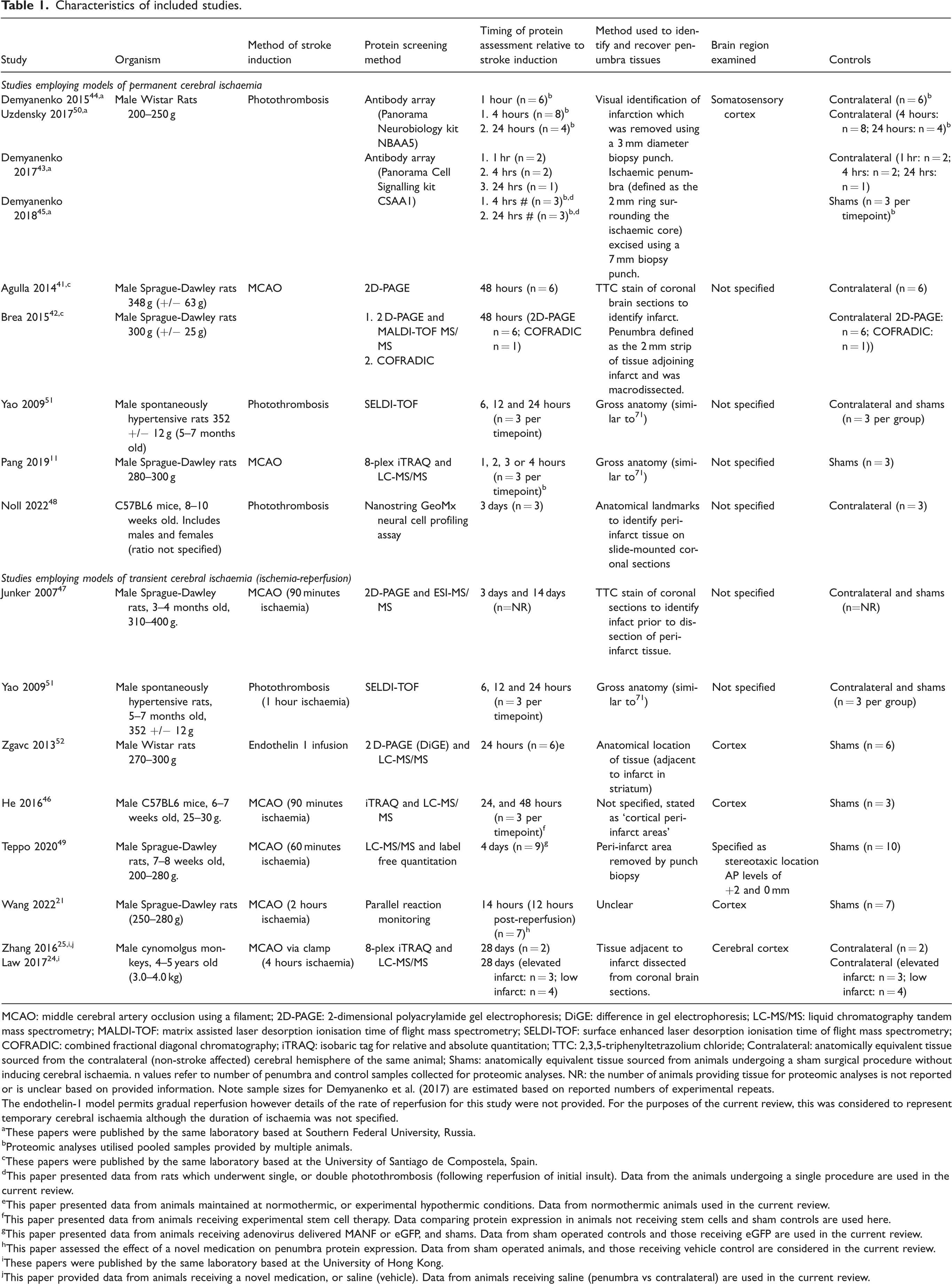
MCAO: middle cerebral artery occlusion using a filament; 2D-PAGE: 2-dimensional polyacrylamide gel electrophoresis; DiGE: difference in gel electrophoresis; LC-MS/MS: liquid chromatography tandem mass spectrometry; MALDI-TOF: matrix assisted laser desorption ionisation time of flight mass spectrometry; SELDI-TOF: surface enhanced laser desorption ionisation time of flight mass spectrometry; COFRADIC: combined fractional diagonal chromatography; iTRAQ: isobaric tag for relative and absolute quantitation; TTC: 2,3,5-triphenyltetrazolium chloride; Contralateral: anatomically equivalent tissue sourced from the contralateral (non-stroke affected) cerebral hemisphere of the same animal; Shams: anatomically equivalent tissue sourced from animals undergoing a sham surgical procedure without inducing cerebral ischaemia. n values refer to number of penumbra and control samples collected for proteomic analyses. NR: the number of animals providing tissue for proteomic analyses is not reported or is unclear based on provided information. Note sample sizes for Demyanenko et al. (2017) are estimated based on reported numbers of experimental repeats.
The endothelin-1 model permits gradual reperfusion however details of the rate of reperfusion for this study were not provided. For the purposes of the current review, this was considered to represent temporary cerebral ischaemia although the duration of ischaemia was not specified.
These papers were published by the same laboratory based at Southern Federal University, Russia.
Proteomic analyses utilised pooled samples provided by multiple animals.
These papers were published by the same laboratory based at the University of Santiago de Compostela, Spain.
This paper presented data from rats which underwent single, or double photothrombosis (following reperfusion of initial insult). Data from the animals undergoing a single procedure are used in the current review.
This paper presented data from animals maintained at normothermic, or experimental hypothermic conditions. Data from normothermic animals used in the current review.
This paper presented data from animals receiving experimental stem cell therapy. Data comparing protein expression in animals not receiving stem cells and sham controls are used here.
This paper presented data from animals receiving adenovirus delivered MANF or eGFP, and shams. Data from sham operated controls and those receiving eGFP are used in the current review.
This paper assessed the effect of a novel medication on penumbra protein expression. Data from sham operated animals, and those receiving vehicle control are considered in the current review.
These papers were published by the same laboratory based at the University of Hong Kong.
This paper provided data from animals receiving a novel medication, or saline (vehicle). Data from animals receiving saline (penumbra vs contralateral) are used in the current review.
Risk of bias assessment
Risk of bias outcomes are shown in Supplement 3. All included papers were assessed against Domain 1 criteria (scores ranging from 30.8%–61.5%). All except Noll et al. (2022) exclusively used male animals, 48 and experiments by Yao et al. (2009) were the only ones to incorporate common risk factors of ischaemic stroke within models. 51 The STAIR guidelines were referenced by one study, 41 and methods for stroke induction were variably reported. Sample sizes for all investigations were relatively small and were not informed by power calculations. None of the studies used cerebral imaging to visualise the ischaemic penumbra and instead relied on post-mortem anatomy to estimate penumbra location, resulting in inter-study variation in the origin of the analysed tissues. Only one paper fulfilled all criteria relating to protein identification and quantification via mass spectrometry. 24 Total scores for the included papers ranged from 22.2%–61.5% denoting moderate - high risk of bias.
Changes to the penumbra proteome during permanent cerebral ischaemia
Six studies assessed the penumbra proteome within 24 hours of permanent stroke induction.11,21,43 –45,50 Of these, Yao et al. (2009) and Pang et al. (2019) did not report the identity or relative abundance of proteins detected during the proteomic assessment, preventing reanalysis of their mass spectrometry-derived data.11,51 Western blots presented by Pang et al. (2019) demonstrate an increased abundance of transforming protein RhoA (RHOA), and isoform 1 of cell division control protein 42 homolog (CDC42) in the penumbra within 4 hours of stroke induction. 11 The remaining four studies used commercially available cell signalling- and neurobiology antibody arrays to measure panels of proteins in brain samples collected during the early (≤6 hours post-stroke induction) and/or late (6–24 hours post-stroke) hyper-acute phase (Table 1) and provided quantitative datasets which were included in the in silico analyses.43 –45,50
Characterising the penumbra proteome during the early hyper-acute phase (≤6 hours post stroke)
Lists of differentially proteins showing differences in abundance between penumbra and control samples within 6 hours of stroke onset were compiled (Supplementary file 2). Inter-study contradictions of the association of MAP1A (microtubule associated protein 1A), MYO5A (myosin 5a), TPH1 (tryptophan hydroxylase 1) and PLCG1 (phospholipase C gamma 1) with the penumbra were observed (more abundant in some studies, less abundant in others). These proteins were removed leaving a non-redundant list of 131 proteins for analysis (75 more abundant, 56 less abundant in the penumbra compared to controls). These proteins generated a highly interactive network, in which the over-abundant over-abundant protein p53 (gene name TP53) was the most connected hub (Figure 2(a)). Pathway analysis revealed significant over-representation of mediators of apoptotic, cell cycle and signalling pathways (ATM-, IL-3-, p53-, MAPK-, and Wnt signalling) within the network, with p53 implicated in seven of the ten most highly ranked pathways (Figure 2(a)).
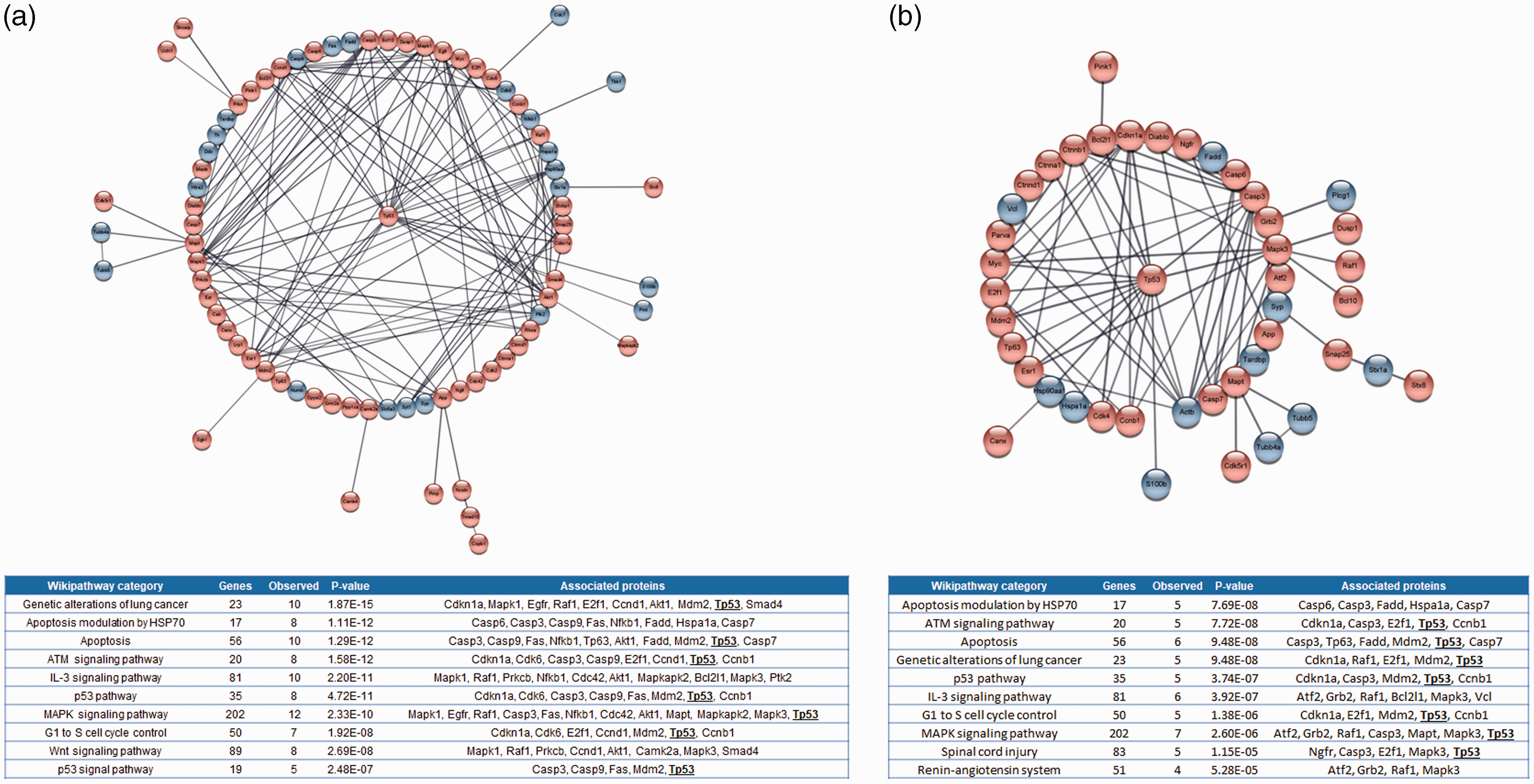
Protein-protein networks and functional enrichment analysis of proteins reported to show significant differences in abundance between penumbra samples and controls collected during the: a) early- (≤6 hours post-stroke) and b) late (6–24 hours post-stroke) hyperacute phases after ischaemic stroke induction. Red and blue shaded bubbles denote proteins which are more, or less abundant in the penumbra compared to control respectively. p53 is positioned centrally. Singletons and interactions involving only 2 proteins are not shown (expanded views are shown in Supplements 4 and 5). Inset tables detail the 10 most highly ranked wikipathway categories represented by proteins within the networks using the Rattus norvegicus genome as a reference. Genes in category refers to the number of proteins belonging to the Wikipathway category, observed genes refers to the number of proteins present in the network for each category. P-values refer to the degree of over-representation (corrected for multiple testing). The inclusion of p53 in multiple pathways is denoted in bold and underlined text.
Characterising the penumbra proteome during the late hyper-acute phase (6–24 hours post stroke)
Demyanenko et al. (2017, 2018) and Uzdensky et al. (2017) presented array data comparing protein abundance in penumbra and control samples collected 24 hours after ischaemic stroke induction.43,45,50 PRKCB was removed due to conflicting inter-study reports of over- or under-abundance in the penumbra leaving 85 unique proteins (32 more abundant, 53 less abundant) for in silico analysis. Over representation of p53 related proteins in the ischaemic penumbra was suggested by both network and pathway enrichment analyses (Figure 2(b)).
Characterising the penumbra proteome during the acute phase (24 hours – 7 days post-stroke)
Acute phase changes to the penumbra proteome were assessed in three studies.41,42,48 Agulla et al. (2014) and Brea et al. (2015) employed comparable methods to sequentially extract proteins from the soluble, insoluble and membrane fractions of penumbra and contralateral controls, 48 hours after the induction of permanent cerebral ischaemia.41,42 Protein screening data were not provided by Agulla et al. (2014), although presented western blots and IHC images indicated a significantly higher abundance of heat shock protein HSP72 in the penumbra. 41 In contrast, Brea et al. (2015) reported significant differences in the abundance of 60 proteins (including HSP72) detected using 2-dimensional electrophoresis and/or gel-free combined fractional diagonal chromatography (COFRADIC, employed on a single biological sample). 42 Of note, HSP-70, serotransferrin and serum albumin were the only proteins suggested to differ in abundance between penumbra and control tissues by both 2DE and COFRADIC. Noll and colleagues (2022) used an array-based method to measure proteins in situ using tissue sections from mice experiencing 3 days of permanent cerebral ischaemia, and reported significant differences in the abundance of 3 proteins (BAG family molecular chaperone regulator 3 (BAG3), Glial fibrillary acidic protein (GFAP) and Phospho-Tau S199 (tau-S199)) between the penumbra (defined as the peri-infarct region by the authors) and contralateral equivalent. 48 Proteins showing significant differences between penumbra and control samples reported by the three studies were compiled. After de-duplication and exclusion of proteins without a unique symbol a non-redundant list of 59 proteins (four at lower-, 55 at higher-abundance in the penumbra) was analysed in silico. Two small networks of interacting proteins were observed, in which mediators of the complement and coagulation cascades, but no other pathways, were over-represented (Supplement 3).
Changes to the penumbra proteome in response to ischaemia-reperfusion
Late hyper-acute phase changes
Four studies investigated the penumbra proteome within the hyper-acute phase following transient cerebral ischaemia.21,46,51,52 One did not report the differentially abundant proteins and was excluded from in silico analyses. 51 Zgavc et al. (2013) reported significance differences in the relative abundance of 14 protein spots following 2DE analysis, from which 26 constituent proteins were identified via mass spectrometry. 52 He et al. (2016), and Wang et al. (2022) both used quantitative mass spectrometry to characterise the penumbra proteome, 24, and 12 hours post-stroke induction.21,46 He and colleagues reported significant differences in the abundance of 9 proteins between penumbra and controls, whereas Wang et al. (2022) did not report the identity of differentially abundant proteins. Reanalysis of mass spectrometry data deposited by Wang et al. (2022), resulted in the identification of 25,000 peptides corresponding to approximately 2,300 quantifiable proteins. Comparing protein abundance between vehicle controls and sham-operated rats revealed significant differences in the abundance of 33 proteins (15 more, and 18 less abundant in the penumbra samples compared to controls, Table 2, Supplement 5). Collectively, the studies provided a non-redundant list of 49 proteins reported to vary significantly between penumbra and control samples. An interaction network of eight proteins was observed (Supplement 6), in which mediators of the complement and coagulation cascades were over-represented (FDR corrected p-value: 0.023).
Proteins showing significant differences in abundance between penumbra samples and controls following reanalysis of data from Wang et al. (2022).
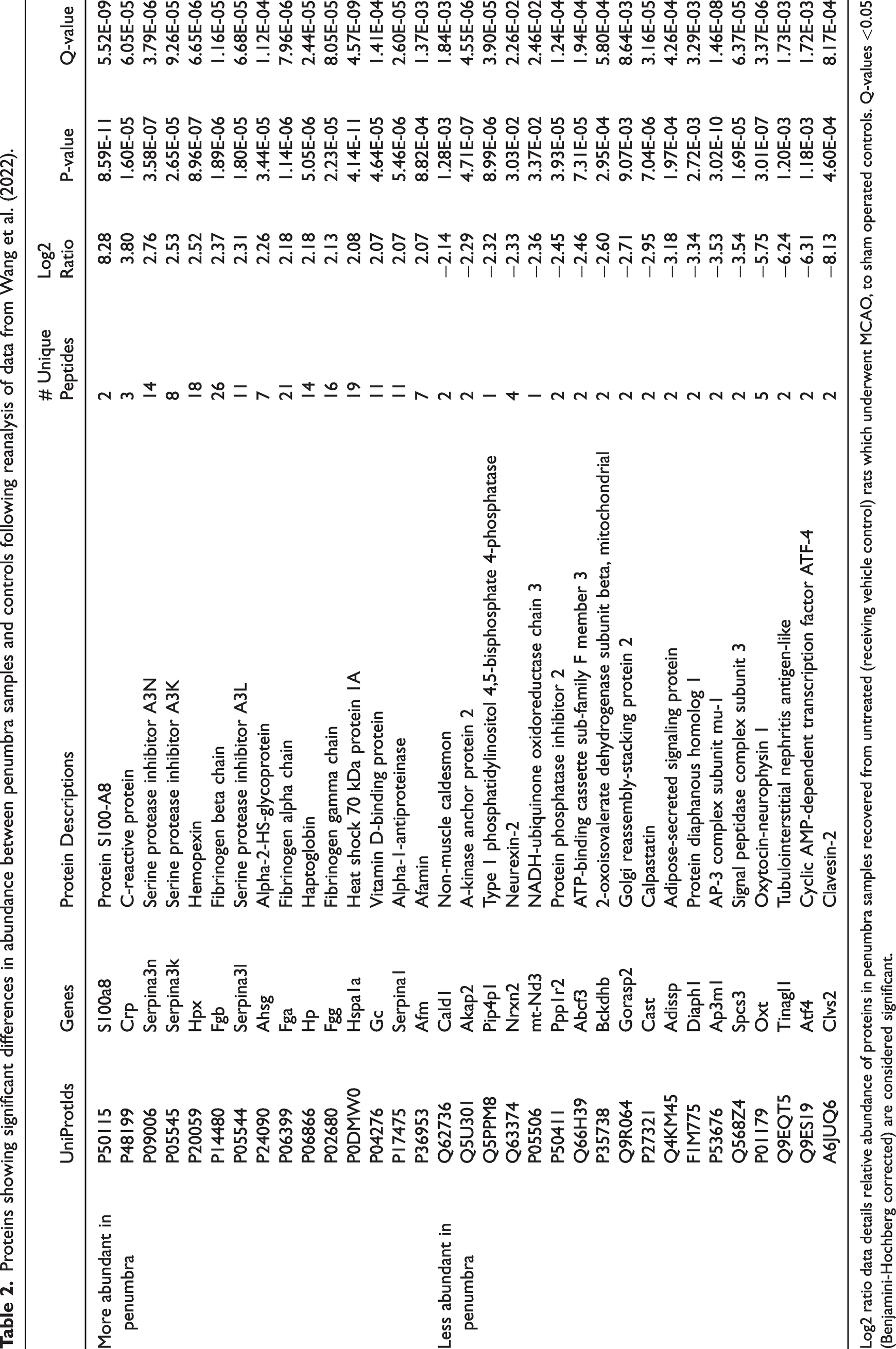
Log2 ratio data details relative abundance of proteins in penumbra samples recovered from untreated (receiving vehicle control) rats which underwent MCAO, to sham operated controls. Q-values <0.05 (Benjamini-Hochberg corrected) are considered significant.
Acute phase changes
Junker et al. (2007), He et al. (2016) and Teppo et al. (2020) detailed protein abundance in penumbra and control samples recovered from mice, 46 and rats,47,49 within 2–4 days of stroke onset. Junker and colleagues conducted a gel-based protein screen, but only presented data to support a significantly higher abundance of annexin-3 and -5 within the penumbra compared to sham controls (corroborated by Teppo et al. (2020)). He et al. (2016) and Teppo et al. (2020) reported differences in the abundance of 4, and 308 proteins, respectively. Of note, both studies reported significant differences in the relative abundance of acidic leucine-rich nuclear phosphoprotein 32 family member B (ANP32B) within the analysed tissues but provided contradictory information regarding the direction of difference and this protein was therefore excluded from further analyses. Compiling data from all three studies provided a non-redundant list of 308 proteins for analysis (202 more, and 106 less abundant in the penumbra than controls). A complex interaction network, most significantly enriched with mediators of protein translation was observed (Wikipathways term WP149, FDR corrected p-value 9.69 × 10−7, Supplement 7). Other pathways associated with IL-3 and G-protein signalling, recovery from spinal cord injury, and cytoskeleton regulation were weakly over-represented (all p-values: 0.043, Supplement 7).
Early Sub-acute changes
Two studies published by the same laboratory employed non-human primate models to investigate the ischaemic penumbra proteome, one month after experimental stroke induction (Table 1).24,25 Zhang et al. (2016) conducted an analysis to identify proteins modulated by tetramethylpyrazine nitrone; proteomic data reported from animals receiving vehicle only were considered in this review (published data contrasts penumbra and paired contra-lateral samples). 25 Law et al. (2017) reported findings of experiments to identify proteins associated with favourable, or poor recovery evidenced by low, or elevated cerebral infarctions, respectively. 24 Both studies used comparable quantitative mass spectrometry (isobaric tag for relative and absolute quantitation). For the purposes of the current review, data provided by Zhang et al (2016) and Law et al. (2017) were divided into those associated with good outcome (differentially regulated proteins identified by Law et al. (2017), and those in the low-infarct group reported by Zhang et al. (2016), total number of proteins: 48); and those associated with poor outcome (differentially regulated proteins identified by Law et al. (2017), and those in the high-infarct group reported by Zhang et al. (2016), total number of proteins: 49). Protein-protein interaction networks generated from these data are shown in Figure 3, with functional enrichment analyses suggesting that favourable outcome was associated with an over-abundance of proteins associated with wound healing (3 A). In contrast proteins associated with prostaglandin synthesis and signalling were highly over-represented in animals experiencing poor outcomes (3B).
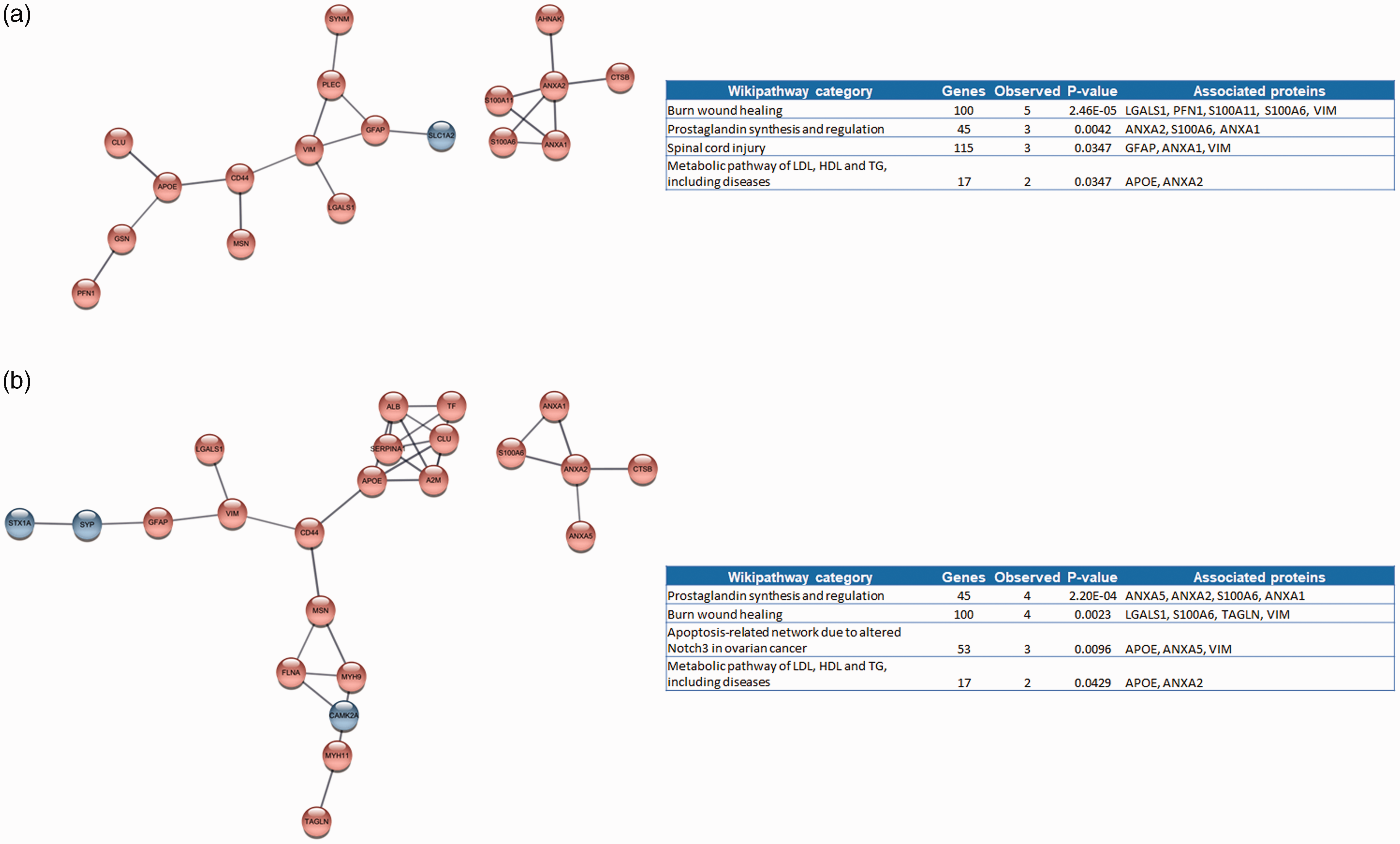
Protein-protein networks and functional enrichment analysis of proteins reported to show significant differences in abundance between penumbra samples and controls associated with a) favourable, and b) poor outcome one month following induction of transient cerebral ischaemia in cynomolgus monkeys. Inset table details wikipathway categories over-represented by proteins within the networks using the Homo sapiens genome as a reference. Genes in category refers to the number of proteins belonging to the wikipathway category, observed genes refers to the number of proteins present in the network for each category. P-values refer to the degree of over-representation (corrected for multiple testing).
Discussion
The current review aimed to assess all available proteomic data provided by studies specifically designed to characterise the ischaemic penumbra following experimental ischaemic stroke. Sixteen eligible studies were included which collectively appraised penumbral biology in the context of permanent cerebral ischaemia or following reperfusion. Marked inter-study differences in experimental design, and the sensitivity and specificity of the employed protein screening tools must be considered when interpreting findings, however available data provide some insight into the processes occurring within the ischaemic penumbra as a foundation for discovery of novel drug targets, and diagnostic and prognostic markers.
Homogenous design and reporting made it possible to analyse data from four included studies.43 –45,50 All four studies reported a higher abundance of the tumour suppressor protein p53 within the penumbra, a finding corroborated by independent investigations (reviewed in 53 ). p53 is a transcription factor that regulates multiple processes including apoptosis, angiogenesis, neuro-inflammation and cell proliferation,54 –56 as reflected in the current network analyses. Although the pathophysiological consequences of an increase in p53 abundance cannot be inferred from proteomic data alone, independent reports suggest that this protein may contribute to stroke pathology by influencing several key processes. 57 For example, suppressing p53 activity significantly reduced the inflammatory response in both in vitro, and in vivo models of ischaemic stroke (discussed in detail by Gao et al. (2023)). 56 Similarly, rats receiving the synthetic p53 inhibitor pifithrin-α before, or within one hour of stroke induction developed fewer apoptotic neurons (evidenced by fewer caspase-3 and TUNEL positive cells on IHC), had smaller cerebral infarctions and experienced better functional outcomes than those receiving vehicle alone.58,59 Teertam et al. (2020), observed similar reductions in stroke-induced apoptosis in rats receiving resveratrol, compared to vehicle controls, and attributed this to increases in sirtuin-1 and mir-149-5p expression which suppressed p53 activity. 55 Other reported benefits of p53 inhibition following stroke, include increased proliferation of neural progenitor cells, 57 preservation of the blood-brain barrier, 60 and enhanced angiogenesis. 61 In contrast Guzenko and colleagues (2023) reported the accumulation of p53 isoforms acetylated at lysines 320 and 373 (p53k320 and 373, respectively) within the cytoplasm of neurons within the penumbra following experimental ischaemic stroke and observed that the p53k320 isoform promoted cell survival. 62 Collectively, available evidence suggests that p53 inhibition during the hyper-acute phase following ischaemic stroke may be beneficial however, the involvement of p53 in multiple homeostatic processes, and potential cross-reactivity of available inhibitors complicates the clinical assessment of this hypothesis. 53 Clarifying which of the multiple pathways mediated by p53 confer the greatest damage following ischaemic stroke onset may help to uncover more specific therapeutic targets.
The processes driving penumbral biology during later stages of permanent cerebral ischaemia, or in response to reperfusion are less clear. Relatively few proteins were reported to differ between penumbra and control tissues recovered from animals experiencing >24 hours of cerebral ischaemia, or those undergoing less than <24 hours of reperfusion. Interestingly, network analyses suggested that coagulation and complement activation were occurring in both groups. The greatest proteomic differences were observed in samples collected between one and seven days after the induction of transient cerebral ischemia, however only one Wikipathway category, ‘translation factors’, showed clear over-representation. This suggests that the penumbra undergoes widespread protein synthesis in the acute phase following reperfusion, however other key processes occurring at this time cannot be inferred from current data.
Only two investigations assessing the sub-acute proteomic response to ischaemic stroke were identified. 24 , 25 Proteins associated with wound healing were associated with favourable outcome 1 month following stroke induction. In contrast molecules involved in prostaglandin synthesis and signalling (e.g. annexins A 1, 2 and 5) were at high abundance in the ischaemic penumbra of animals experiencing poor outcome. Annexins A1 and 2 have been reported to act to reduce cerebral infarct volume, inflammation and thrombosis in animals models of ischaemic stroke (see Ansari et al. (2018) and Mendez-Barbero et al. (2022) for comprehensive reviews).63,64 It is therefore possible that increased expression of these proteins following stroke may be a compensatory response and they may be useful prognostic biomarkers to predict stroke recovery. High serum annexin A7 concentration has been positively correlated to NIHSS score and was associated with early neurological deficit and severe disability or death within 3 months for patients experiencing intra-cerebral haemorrhage, and high plasma annexin 5 concentration was reported to predict high risk of death following myocardial infarction. 65 , 66 Clinical cohort studies investigating the association of annexins with outcome following ischaemic stroke are needed.
Determining the relevance of the penumbra proteome of animal models to patients is complicated by a lack of comparable studies conducted on human penumbra biopsies. 16 Two studies have conducted proteomic analyses on samples recovered from donors, and have reported significant differences in the proteins present within the penumbra (defined as the peri-infarct tissue), the infarction core and biopsies from non-stroke controls.67,68 Samples in both studies were provided after donor death (within 6 hours), and it is unclear whether tissues collected post-mortem reflect the in vivo proteome. In contrast, Dayon et al. (2010) demonstrated significant differences in the protein content of cerebral fluid aspirates collected from the site of the ischaemic infarct, penumbra and contralateral brain hemisphere of patients undergoing decompressive craniotomy following middle cerebral artery territory stroke. 69 Whilst this novel approach bypasses limitations associated with the analysis of post-mortem tissues, generated data do not represent the penumbra and there remains little direct evidence of the protein-level changes occurring within the penumbra of patients experiencing ischaemic stroke. 69
The current review highlights significant inter-study heterogeneity in the design and reporting of outcomes from investigations of the ischaemic penumbra proteome. Importantly, no consistent method to identify and sample the penumbra was employed and tissues were obtained from different regions of the brain. RCTs demonstrating benefits of late reperfusion have detected ischaemic penumbra in patients by identifying a mis-match in areas highlighted by diffusion- and perfusion-weighted magnetic resonance imaging (MRI).8 –10 Imaging studies have demonstrated that the same mis-match method can be applied to animal models of ischaemic stroke,6,70 however this was not used by any of the included investigations and it is therefore unclear whether the analysed tissues reflect the penumbra identified in patients. Some studies mitigated this risk by presenting histological evidence demonstrating hypoxic stress in the assessed tissues (e.g. perineuronal oedema, hyperchromic nuclei or expression of neurological stress markers).24,25,43,44,48,50 Others11,51 adopted methods to isolate regions of the brain reported to functionally recover post-stroke described by a prior investigation, 71 or did not provide evidence to confirm the phenotype of the assessed tissue, and the relevance of the sample is therefore unclear.21,24,41,42,45 –47 It is also noteworthy that most of the included studies used young, healthy animals which does not represent the human population at risk of stroke. 4 Only one study included female animals, 48 and all proteomic investigations had group sizes of 10 animals or fewer. Moreover, all studies contributing data into the hyper-acute response to cerebral ischaemia used antibody arrays to assess the penumbral proteome. This is a potential source of bias since the arrays assess a relatively small panel of proteins known to drive a range of pre-selected cellular pathways, and the possibility of overlooking other differentially abundant proteins must be considered. Finally, the current analysis was conducted using simplified data in which proteins were classified as either over- or under-abundant within the penumbra. Whilst this highlighted over-arching cellular pathways, an analysis using quantitative proteomic data would provide greater sensitivity to identify pathways showing the greatest dysregulation within the penumbra.
In conclusion, the current study identified consistent evidence of over-representation of p53 related proteins in the ischaemic penumbra following experimental stroke. Well-powered, studies which combine clinically accepted approaches to identify the ischaemic penumbra with high-throughput modern proteomics to detect changes in protein abundance and post-translational modification are needed to further current understanding. Dedicated studies in females who have been excluded from many of the past studies are also needed.
Supplemental Material
sj-pdf-1-jcb-10.1177_0271678X241248502 - Supplemental material for A systematic review and in silico analysis of studies investigating the ischaemic penumbra proteome in animal models of experimental stroke
Supplemental material, sj-pdf-1-jcb-10.1177_0271678X241248502 for A systematic review and in silico analysis of studies investigating the ischaemic penumbra proteome in animal models of experimental stroke by Joseph V Moxon, Cornea Pretorius, Alexandra F Trollope, Parul Mittal, Manuela Klingler-Hoffmann, Peter Hoffmann and Jonathan Golledge in Journal of Cerebral Blood Flow & Metabolism
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: JVM is supported by a donation from the Donald and Joan Wilson Foundation and a Leading Edge grant (EMLE-204R36-2021) from the Emergency Medicine Foundation (Australasia) Queensland Program. PH gratefully acknowledges the support of the National Collaborative Research Infrastructure Strategy (NCRIS) and the South Australian Government. Research by Jonathan Golledge is supported by grants from the National Health and Medical Research Council, Heart Foundation, Medical Research Futures Fund, Queensland Government, Townsville and Hospital Health Services and Tropical Australian Academic Health Centre. Funders had no role in data collection and analysis, manuscript preparation and publication.
Acknowledgements
The authors thank Mr Stephen Anderson (James Cook University Library) for expert assistance in developing the literature search strategy.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplementary material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.