
Abstract
Adrenoceptor and calcium channel modulating medications are widely used in clinical practice for acute neurological and systemic conditions. It is generally assumed that the cerebrovascular effects of these drugs mirror that of their systemic effects – and this is reflected in how these medications are currently used in clinical practice. However, recent research suggests that there are distinct cerebrovascular-specific effects of these medications that are related to the unique characteristics of the cerebrovascular anatomy including the regional heterogeneity in density and distribution of adrenoceptor subtypes and calcium channels along the cerebrovasculature. In this review, we critically evaluate existing basic science and clinical research to discuss known and putative interactions between adrenoceptor and calcium channel modulating pharmacotherapies, the neurovascular unit, and cerebrovascular anatomy. In doing so, we provide a rationale for selecting vasoactive medications based on lesion location and lay a foundation for future investigations that will define neuroprotective paradigms of adrenoceptor and calcium channel modulating therapies to improve neurological outcomes in acute neurological and systemic disorders.
Introduction
The cerebrovascular tree is a complex physiological network comprised of a dynamic and regionally diverse architecture. These physiological systems have evolved to develop elaborate cerebral autoregulatory mechanisms that maintain adequacy of cerebral blood flow in response to changing physiological conditions, thereby optimizing neuronal function and survival.1,2 Several human and animal studies have shown that the mechanisms used to orchestrate cerebral autoregulation involve the intercommunication of specialized cells of the neurovascular unit and that adrenoceptor-mediated physiology regulates vasomotor tone and cerebrovascular blood flow.3–7
The ability to autoregulate blood flow is not unique to the brain – the myocardial vascular network is also subject to the effects of blood flow alteration to optimize myocardial perfusion. 8 These observations have led to the development of systemic vasoactive medications that act on adrenoceptors and calcium channels to improve myocardial and systemic perfusion in various disease states. While these therapies are ubiquitously applied to acute systemic and cardiovascular conditions, the cerebrovascular and neurological effects of these medications remain largely unknown. Adrenoceptor agonists, antagonists, and calcium channel antagonists are among the most common pharmacotherapies used in the acute care setting. As such, this review will focus on the cerebrovascular effects of these medications and their putative effects on neuronal function in acute neurological and systemic illnesses.
The neurovascular unit
Specialized cells
The cerebrovascular network is comprised of specialized cells with biological functions that are crucial to vascular structure and function. Collectively they are referred to as the neurovascular unit, a concept that emerged from the first Stroke Progress Review Group meeting of the National Institute of Neurological Disorders and Stroke of the National Institutes of Health (July 2001). The neurovascular unit is thought to play a central role in cerebral autoregulation, a process that is reliant on neurovascular coupling, or the linking of neuronal activation with blood flow. The key cellular components of the neurovascular unit that work in concert to mediate cerebral autoregulation are of neural and vascular origin (Figure 1).
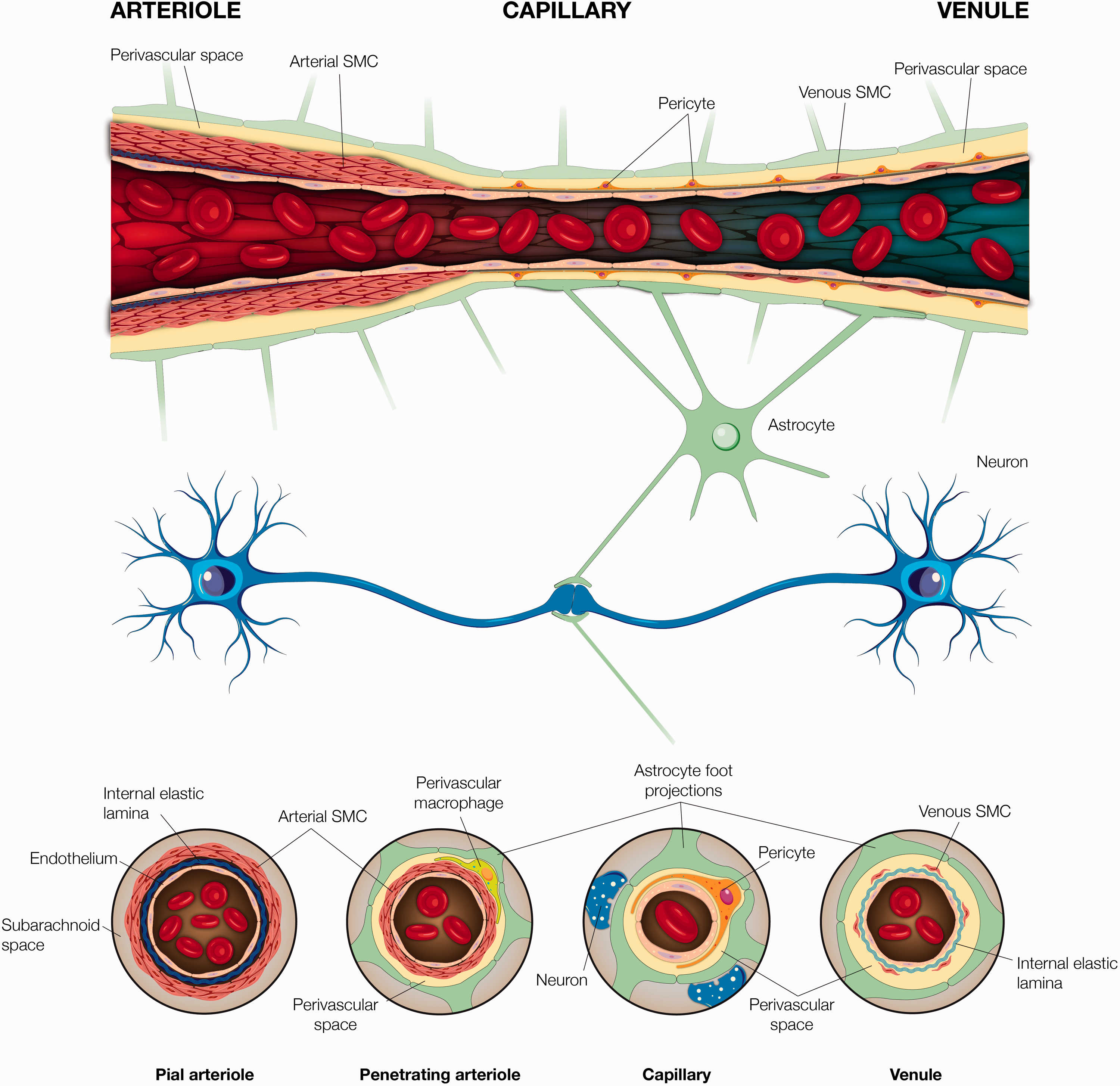
Neurovascular anatomy and its associated components. A visual representation of the neurovascular anatomy provided here in a longitudinal-and-cross sectional fashion along pial and penetrating arterioles, capillary, and venule. Arterioles comprise of a thick coat of smooth muscle cells, which become thinner and are replaced by pericytes as the vessel transitions into a capillary. A thin layer of smooth muscle cells reappear as capillaries transition into venules. A continuous endothelial cell layer, as well as a neural innervation is present along the vascular transition from penetrating arteriole to venule.
The neural components involved with neurovascular coupling include neurons and astrocytes. Under normal physiological conditions, one mechanism by which neurons regulate cerebral blood flow is via neurohormonal action on adrenergic receptors. Neurohormones, such as norepinephrine, arise from subcortical nucleus projections to generate molecular and electrical signals that modulate cerebral perfusion 9 by increasing vasoconstrictor tone typically in small cerebral vessels along the cortex. 3 However, it is important to recognize that there are multiple, and perhaps redundant pathways to regulate cerebral blood flow including ones that are independent of adrenoceptor modulation, such as through metabolic factors and somatosensory stimulation.10–13 Modulation of vascular tone mediated by neuronal metabolic demands leads to processes that promote vasodilation, while increasing vasoconstrictor tone to focus oxygen delivery to activated areas. 3 Astrocytes are diverse cell types that vary substantially in distribution, morphology, and function. Astrocytic end-feet surround over 90% of cerebral capillaries and alter their vascular tone in response to neuronal activation. 14 In addition to serving as a cellular link between neurons and endothelial cells, astrocytes directly affect endothelial function and adjacent microvessel diameter via calcium channels. 15
The vascular component of the neurovascular unit consists of myocytes, pericytes, and endothelia. Myocytes, found primarily in proximal cerebral arteries, pial vessels, and penetrating arterioles, are considered the main modulators of vasomotor tone. This tone is generated either in response to fluctuations in intravascular pressure or via neural activity. 16 Myocyte-generated vascular tone, predominantly in the penetrating arterioles, is essential for cerebral autoregulation and is thought to underlie neurovascular coupling. 17
Further distally along the cerebrovascular tree, as early as the pre-capillary arterioles, pericytes replace myocytes. Pericytes appear to contribute to vessel stability, permeability, remodeling, and angiogenesis. Like myocytes, pericytes are hypothesized to serve contractile functions, especially in capillaries proximal to arterioles, 18 however given their morphological heterogeneity, they also serve non-contractile functions, such as maintaining the blood brain barrier and vascular structure.19,20 Both pericytes and myocytes are intricately linked by a single layer of cells that structurally comprise the inner walls of cerebral vessels, known as the endothelium. The endothelium plays a significant role in modulating vascular tone and cerebral blood flow via calcium channels, 21 electrical signaling, 22 and longitudinal signaling via endothelial gap junctions. 23
Recent investigations have increasingly focused on cellular mechanisms that underlie the organization and function of the neurovascular unit; however, several gaps remain in our understanding of specialized cell roles, and the essential mechanisms and factors necessary to affect cerebral blood flow. Better characterization of mechanisms relevant to producing physiologic responses in the regulation of cerebral blood flow is warranted. In general, current paradigms of treatment with vasoactive pharmacotherapies do not account for the nuances and complexity of cerebrovascular anatomy, and future research is needed to develop current therapeutic paradigms that account for these factors.
Architecture
The organization of vascular endothelium and neuronal innervation varies significantly along the cerebrovascular tree. Branching off from the circle of Willis anteriorly and the basilar artery posteriorly, cerebral arteries progressively give rise to the pial arteries. This network of interconnecting arteries courses along the surface of the brain, which then infiltrates the surface to produce the penetrating arteries and subsequently become the parenchymal arterioles. 24 Vessel organization and anatomy vary at different segments along the cerebrovascular tree (Figure 1)– as pial arteries penetrate deep into the brain to become parenchymal arterioles, the myocyte layer becomes thinner until a single layer remains, 25 perivascular nerves become sparse 26 and eventually, the perivascular space is obliterated. 27 At these levels, a single or discontinuous layer of myocytes and astrocytic end-feet encase arterioles, 26 displaying features that are unique to the neurovascular unit.
As arterioles transition into the extensive capillary network of the brain, pericytes replace myocytes to surround capillary endothelium along with the basal lamina. Along the pre-capillary vessel, pericytes are tightly wrapped around the endothelium in a perpendicular axis. Here, pericytes are thought to provide contractile support given their orientation to the endothelium and for expressing a similar marker to myocytes– smooth muscle α-actin. 25 At the level of capillaries, the function of pericytes remains controversial since they either express less or lack smooth muscle α-actin18,28 with shifting orientation18,29 along the endothelium suggesting that they may not have contractile properties.
Given their large surface area and ability to produce large changes in flow with minimal changes to individual diameters, capillaries are thought to be well-suited to rapidly regulate cerebral blood flow to address the metabolic demands of neurons, which are positioned in close proximity. 30 The role of capillaries in neurovascular coupling was studied in mice by Cai et al who identified that first- and second-order capillaries initiate dilation in response to neurovascular coupling signals before penetrating arterioles and higher order capillaries. 31 Similar findings were reported during cerebral ischemia. 32 These findings suggest that capillaries proximal to penetrating arterioles may play a role in regulating local blood flow, however, identifying the exact role of capillaries in neurovascular coupling and the cell types that first sense and then generate the vascular response remain unclear.
The cerebral venous system comprises of valveless veins that transition from venules to superficial cortical veins that empty into the dural venous sinuses and finally into the jugular veins. As cerebral capillaries transition into small venules, endothelium remains lined with pericytes with stellate cell bodies and many branch-like projections. Contrary to arterioles, these projections do not encircle endothelium and randomly orient with respect to vessel axis, forming a dense meshwork. Given this organization in post-capillary venules, pericytes are thought to have minimal veno-constrictive properties, 19 however there is some evidence that they may produce contractile forces to help regulate blood flow. 33 As small venules become larger, they are surrounded by a small layer of myocytes. The membrane potentials of myocytes and pericytes are modulated via voltage dependent calcium channels that can be inhibited by calcium channel antagonists. 34 In bovine retinal pericytes grown in tissue culture, norepinephrine has also been shown to induce action potentials, which are dependent on extracellular calcium. 34 Given the similar types of specialized cells in both arterioles and venules including myocytes and pericytes, (Figure 1), it is possible that adrenoceptor and calcium channel modulation may influence venular tone and blood flow; however, this remains an active area of research at this time with unknown clinical implications.
The organization of vascular components is matched in complexity by their organization and innervation by astrocytes and neurons along the cerebrovascular tree. Pial arteries and arterioles receive sympathetic, parasympathetic and sensory innervation from peripheral autonomic ganglia, specifically the superior cervical, trigeminal, and sphenopalatine ganglia respectively.2,35 Peripheral innervation is progressively lost as arteries enter parenchymal tissue. Parenchymal arterioles and cortical microvessels are thought to receive input by adrenergic nerve projections originating from locus coeruleus, raphe nucleus, basal forebrain, or cortical interneurons.35,36 Nearly half of the cerebrovascular resistance is thought to be mediated via parenchymal arteriolar contribution. 37 At the capillary level, there is continuous astrocytic end-feet coverage of basal lamina, with only sparse support by neurons. Additionally, perivascular innervation for both serotonin and norepinephrine is thought to decrease when moving from rostral to caudal vasculature, such that the internal carotid arteries are more densely innervated than the vertebrobasilar system. 38 Neuronal input to the neurovascular tree presents in the form of neurohormones, such as norepinephrine, which target adrenoceptors, specifically alpha (α)- and beta (β)-receptors on endothelium, pericytes and myocytes (Figure 2). These adrenoceptors serve as targets that are thought to modulate cerebrovascular tone in response to vasoactive medications that exert agonistic and antagonistic effects (Table 1).13,39–46
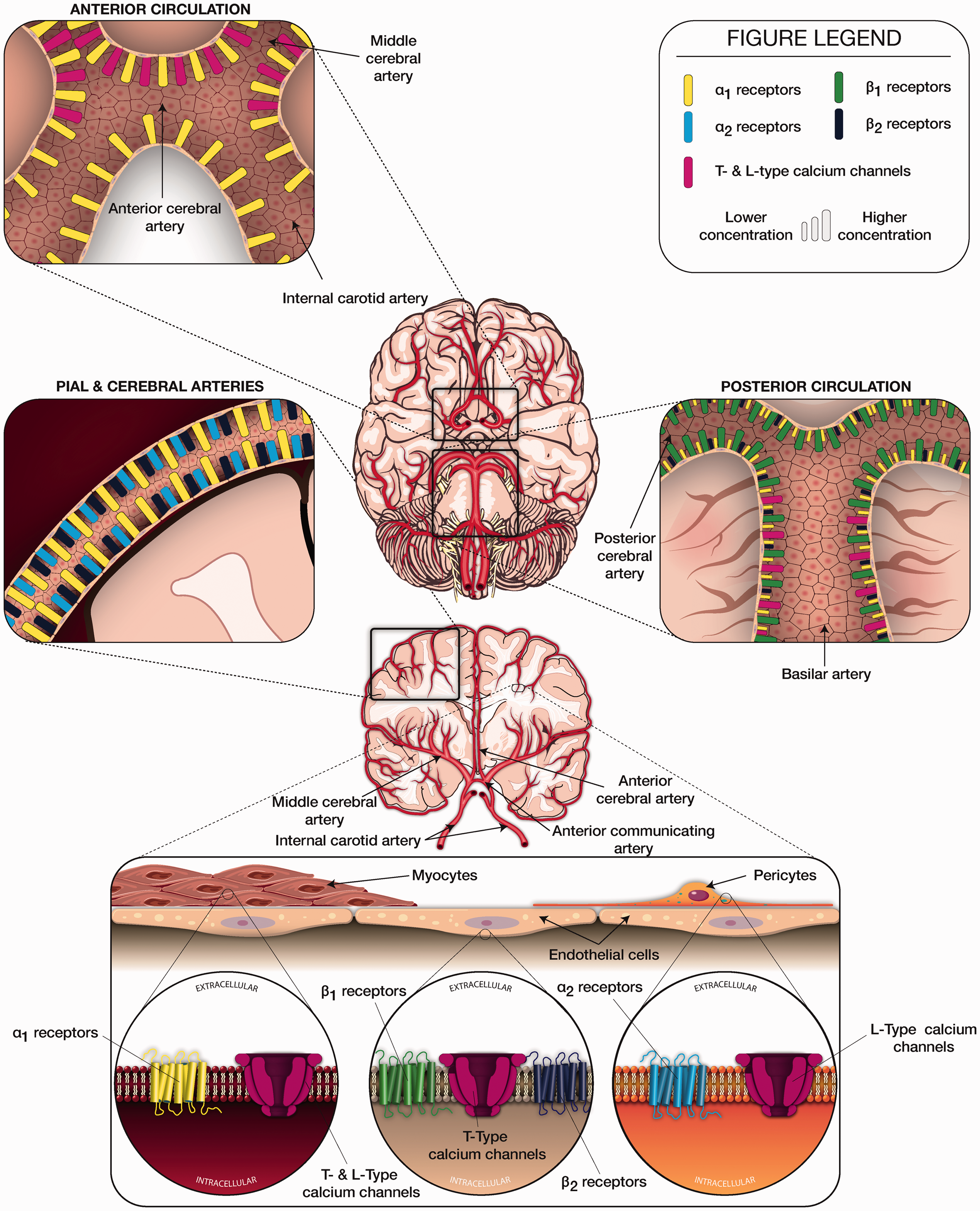
Schematic description of subtypes, distribution, and density of adrenoceptors and calcium channels along the cerebrovascular tree.
Summary of adrenoceptor subtypes and pharmacology. Asterix denotes degree of agonist effect (*= mild effect, **= moderate effect, and ***= marked effect).

Adrenoceptors and calcium channel subtypes
Neurohormonal receptors, in coordination with the neurovascular unit, are thought to play a central role in modulating cerebrovascular autoregulation. 47 Although the neurovascular unit has unique components that help modulate cerebral blood flow, the presence of adrenoceptors and calcium channels is a feature shared with the systemic vasculature. Clinically, it is widely assumed that the therapeutic cerebrovascular effects of medications that act on adrenoceptors and calcium channels mirror the systemic effects; however, careful consideration of the components of the neurovascular unit and cerebrovascular anatomy suggest this assumption to be less plausible given the substantial heterogeneity in receptor distribution along the cerebrovascular tree, among other factors.
Adrenoceptors
Adrenoceptors consist of two major classes, α- and β-receptors, which mediate the actions of the neurotransmitter hormones norepinephrine and epinephrine. The direct binding of norepinephrine to adrenoreceptors is believed to trigger G-protein receptor coupled pathways that activate a cascade of biochemical events that affect a multitude of physiological functions.30,48 Examples of such adrenoceptor-mediated functions include: α1- and β2-receptor modulation of vascular, genitourinary, and gastrointestinal smooth muscle tone, α1- and β1-receptor-mediated control of cardiac inotropy and chronotropy, α1- and β2-receptor-mediated regulation of glycogen and glucose metabolism in the liver, α2- and β2-effect on insulin secretion from pancreatic cells, α2- receptor- mediated effects on platelet aggregation, vascular smooth muscle contraction, and feedback inhibition of the sympathetic transmission in neurons. 49 Adrenoceptor dysfunction is implicated in several neurological and systemic pathophysiological states. For instance, α1-receptors are implicated in neurodegenerative diseases, such as Alzheimer’s disease, 50 while β1- and β2- receptor dysfunction contribute to cardiac pathology, 51 and α2-receptors regulate sympathetic tone, neurotransmitter release, and blood pressure.52,53
Although it is widely assumed that the effects of sympathetic innervation of cerebrovasculature mirror that of the systemic vasculature, the high variation in type, density and distribution of adrenergic receptors in the cerebrovasculature suggest that medications that act on these receptors may have differential effects based on location and pathology.54–56 Further, a large body of basic and clinical evidence has recently come to demonstrate cerebrovascular effects of extrinsic neurohormonal modulation via pharmacotherapies, which are commonly prescribed in the acute care setting.
Adrenoceptor agonists and antagonists are widely used in the care of neurologically ill patients, most often to modulate systemic and cerebral blood pressure with the intent to optimize cerebral blood flow. Several animal and human studies have attempted to clarify the identity and nature of adrenoceptor subtypes in different regions of the cerebrovascular tree by using adrenoceptor modulating agents such as epinephrine (α1-, α2- and β2- receptor agonist), norepinephrine (α1- and α2-receptor agonist), phenylephrine (α1- receptor agonist), prazosin (α1-receptor antagonist) and yohimbine (α2-receptor antagonist). A summary of adrenoceptor subtypes and relevant medications is provided in Table 1.39–46,57–61 Prior studies have identified α1-receptors in regulating cerebrovascular tone and cerebral blood flow via action on myocytes. 62 In addition to myocytes housing adrenoceptors, Elfont and colleagues studied pericytes and endothelial cells of bovine cerebral microvessels to find β1- and β2-receptors predominantly on endothelial cells and α2-receptors on pericytes 63 (Figure 2). Alpha2-receptors have also been identified in neurons and are involved with analgesic and sedating effects. 64
The anatomical distribution of adrenoceptor subtypes and concentrations along the cerebrovascular tree is believed to be variable, as demonstrated in various human and animal studies (Figure 2). In rat and bovine cerebrovasculature, a relative abundance of α1-receptors on pial and superficial cerebral arteries, α2-receptors on intraparenchymal cerebral microvessels, and relatively high concentrations of β2-receptors on cerebral microvessels have been described. 44 Ayajiki and Toda also studied bovine cerebrovasculature and found α1-receptor mediated vasoconstriction of rostral arteries (internal carotid, anterior cerebral, and middle cerebral arteries) and β1-receptor mediated vasodilation of caudal arteries (posterior communicating, posterior cerebral, and basilar arteries).39,40 Although the precise mechanism of β1-receptor mediated vasodilation remains unclear, studies suggest nitric oxide release through activation of β1-receptors on endothelial cells. 65 Gaw and Wadsworth investigated post-synaptic α -receptors in sheep by applying norepinephrine to basilar, middle cerebral, and small pial arteries. 43 The effects on pial and middle cerebral arteries were larger compared to the basilar artery (78%, 92%, and 28% of maximum contraction respectively), which is in line with previously noted regional differences in adrenoceptor distribution. Norepinephrine induced contraction with α1-receptor activation in middle cerebral arteries has been demonstrated, in variable degrees, in humans and numerous other species of animals.41,42 Vasoconstriction via norepinephrine use was also reported in postmortem human basilar artery 46 while vasodilatory properties, or no response, were reported along the posterior circulation in various other animal species. 41 Stimulation of α2-receptors did not result in rat cerebral vasoconstriction. 45 These anatomical patterns suggest variability in distribution of adrenoceptor concentrations and types along the cerebrovascular tree (Figure 2) and that differential activation of these adrenoceptors may affect regional cerebral perfusion patterns.
Alpha and β-receptor antagonists are typically used in clinical practice to treat hypertension-related emergencies. For these indications, β-receptors are more frequently targeted via the use of β1-receptor selective agents, such as esmolol, atenolol or metoprolol, and β-receptor non-selective agents, such as propranolol (Table 1). Data regarding the effects of β-receptor antagonists on cerebrovasculature are limited, however studies have suggested that propranolol may have vasodilatory properties in rat and porcine basilar arteries. 66 Early propranolol administration in patients with moderate to severe traumatic brain injury may provide neuroprotective effects and improved mortality. 66 These observed clinical benefits of therapy may address hyper-adrenergic states related to norepinephrine and epinephrine surges as seen in traumatic brain injury and subarachnoid hemorrhage. 67 In mice with traumatic brain injury, propranolol was associated with improved cerebral perfusion and neurologic recovery. 68
It is unclear whether the benefits from β-receptor antagonism in traumatic or stress induced brain injury may be related to systemic or central effects. Potential central benefits of β-receptor antagonism are speculative and may be mediated through decreased cerebral metabolism and blood flow. Similar findings have been demonstrated with esmolol use during recovery from neurosurgical anesthesia where cerebral hyperemia from sympathetic overactivity was blunted with esmolol administration, resulting in decreased cerebral blood flow velocity, heart rate and cardiac output. 69 Beta 1- and β2-receptors have been identified in various cerebral structures in animal models. Rainbow et al used 125I-labeled pindolol and identified high levels of β1-receptors in the axons or cell membranes of neurons in the hippocampus, layers I and II of cerebral cortex, thalamic mediodorsal and ventral nuclei, and cingulate gyrus and high β2-receptor concentrations in the cerebellum, pia mater, thalamic central, paraventricular and caudal lateral posterior nuclei, and paraventricular structures. 70 These findings suggest central effects of certain adrenoceptors may be related to blood flow and cellular metabolism.
Alpha and β – receptor modulating pharmacotherapies hold important clinical relevance as these receptors are seen in multiple organ systems, including myocardial vasculature (Table 1). Such therapies are utilized in critical care and emergency settings to treat acute shock and hypotensive states. Relatively more is known about the systemic and coronary effects of adrenoceptor modulating therapies; thus, they are commonly used with limited understanding of their short- and long-term effects on cerebrovasculature and neuronal health. In one study, Gedeborg et al investigated the effects of high-dose epinephrine on cerebral blood flow during experimental cardiopulmonary resuscitation in cows to find that high-dose epinephrine (200ug/kg) used during resuscitation resulted in attenuated blood flow in superficial cerebral cortex, as well as higher coronary perfusion pressures. 71 As a consequence, high doses of epinephrine resulted in the redistribution and physiologic shunting of blood flow from the superficial cortex, making this region vulnerable to ischemia and possibly reperfusion injuries. 72 An even earlier experiment by Greenfield and Tindall from 1968 investigated the effects of norepinephrine and epinephrine on internal carotid artery blood flow in humans and reported differential effects of arterial versus intravenous administration of norepinephrine and epinephrine on cerebral blood flow. 73 Taken together, these studies suggest that vasopressors may alter cerebrovascular hemodynamics through multiple adrenoceptor-mediated mechanisms that are not clearly defined at this time.
Calcium channel receptors
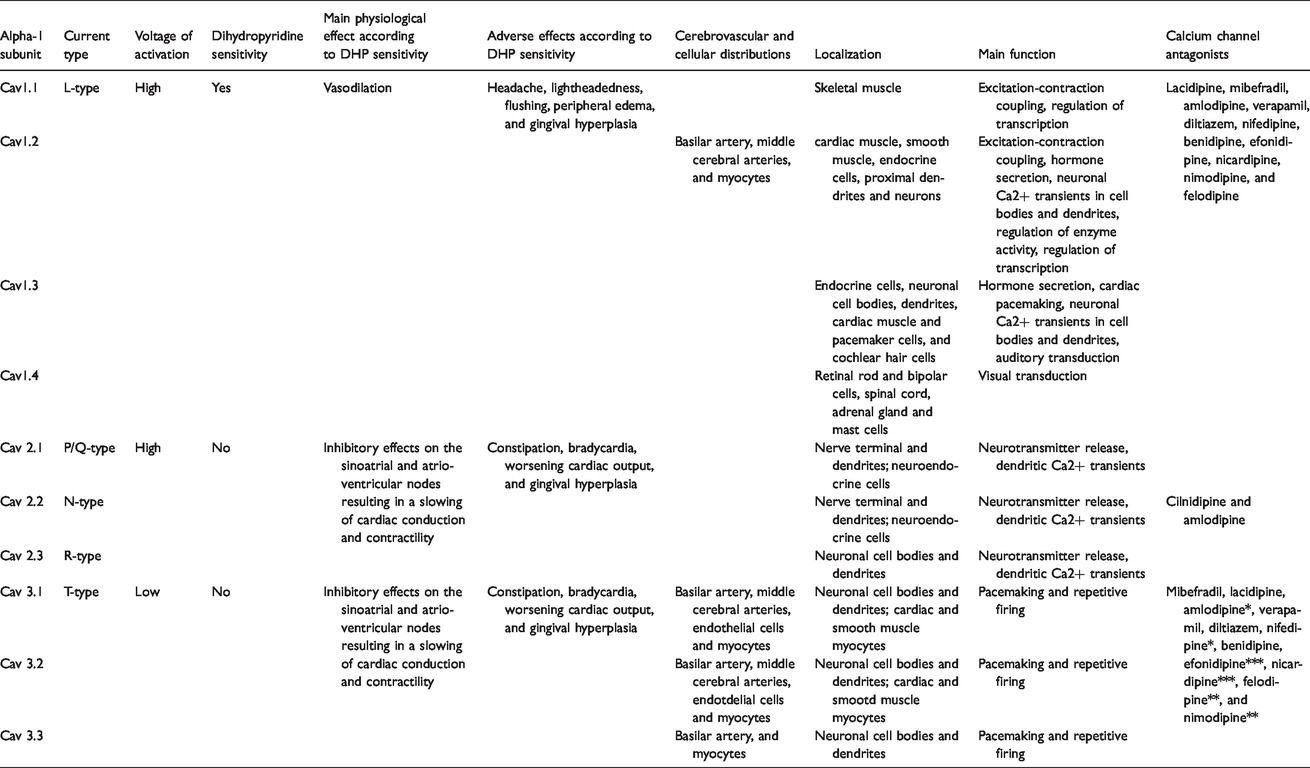
Voltage gated calcium channels perform important physiological roles that help facilitate cerebral blood flow through various mechanisms, such as coupling of excitation-contraction functions in myocytes 74 or via transmitter or hormone release in neuronal synapses. 75 Electrophysiological studies on vertebrates have shown that the α1-subunit of voltage gated calcium channels determines both the pharmacological and biophysical properties of calcium currents. 76 The CaV1 family of calcium channels, which are characterized by long-lasting inward currents during depolarization or “L-type” channels, are mainly involved with skeletal muscle contraction, however other subtypes of channels within CaV1 family are located in neurons, 77 adrenal chromaffin cells, 78 and cardiomyocytes and sinoatrial nodes 79 to induce cellular excitation. The CaV2 family of calcium channels are primarily located in synapses and are thought to drive evoked synaptic transmission 80 of neurotransmitters 81 or hormones 82 via synaptic vesicle formation and release and assist with conductance via calcium-activated potassium channels. 83 Finally, the CaV3 family of calcium channels are characterized by rapidly decaying calcium currents, termed transient or “T-type” channels, that allow for regulation of rapid changes in neuronal, cerebrovascular, and cardiac functions.84–86 A summary of calcium channel physiology and relevant medications is provided in Table 2.87–93
Summary of calcium channel physiology and pharmacology. Medications listed in parenthesis represent established T- and L-type calcium channel antagonists. Level of affinity of a medication to the T-type Cav 3.2 calcium channel represented with an asterix (* = low, ** = medium, and *** = high). Pericytes also expresses L-type calcium channels, however their molecular makeup is currently unknown.
The types, density, and distributions calcium channels in cerebrovasculature have been examined in select animal studies. Relatively high concentration of CaV1.1 were reported in human basal ganglia by Takahashi et al. 94 Schlick et al reported similar CaV subtypes expressed in the cortex and hippocampus of adult mice brain, which were strikingly different in the cerebellum. 95 Similar findings highlighting a heterogeneous population of calcium channels were mirrored in a study by Kuo et al, who reported dihydropyridine –insensitive and sensitive channels in the basilar and middle cerebral arteries and their branching small vessel myocytes of rats 92 and Nikitina et al, who reported low-voltage-activated T-type channels in basilar artery myocytes in dogs. 93 However, although findings in both studies suggest species variation, insensitive channels were more prominent in smaller vessels while sensitive channels, but of both L- and T-types, were present in basilar and middle cerebral arteries. 92 In addition to myocytes, which express both L- and T-type channels, pericytes express L-type voltage gated calcium channels with excitation-contraction coupling that is much less in comparison to myocytes,92,96,97 and endothelial cells express T-type channels whose function is yet to be determined. 92 The regional variations in calcium channel subtypes along with differing responsiveness to dihydropyridine based therapies suggest that opportunities may exist to select calcium channel antagonist therapy that is tailored to the characteristics of the underlying neurological pathology and intended neurological outcome.
The vascular component of the neurovascular unit utilizes an intrinsic response to intravascular transmural pressures, referred to as the myogenic response, to maintain adequate cerebral blood flow. 98 This response was previously thought to be exclusively in arteries and arterioles, 99 however, more recently it has been hypothesized that a myogenic response may also occur in the venous circulation due to the presence of myocytes in venules. 29 Myocytes are recognized as key components of the myogenic response, due to their ability to serve as the main effectors of vascular diameter and tone. The myogenic response consists of a cascade of events, initiated by increased transmural pressures that depolarize the myocyte membrane and activates multiple ion channels, including calcium, potassium and chloride channels. 100 The calcium influx into the depolarized myocytes via opened voltage-gated calcium channels increases myosin light-chain phosphorylation, producing vasoconstriction, while the binding of drugs to the pore-forming α1-subunit inhibits inward flow of calcium ions and halts membrane depolarization, leading to vasodilatation. 101 CaV1.2 is the main calcium channel in the cardiac and systemic vascular systems and is also thought to be the prominent channel involved in the cerebrovascular myogenic response, although other CaV channels are present in certain segments of small parenchymal arterioles. 102
Clinical manifestations of pathophysiological conditions related to calcium channels are often associated with congenital genetic disorders, such as hypokalemic periodic paralysis, 103 Timothy Syndrome, 104 and Brugada syndrome. 105 Calcium channelopathies are also associated with neurological conditions, such as episodic ataxia 106 and various epilepsy syndromes. 106 Impaired vascular relaxation has also been reported to occur in calcium channelopathies, 107 which suggests that calcium channels may play a role in cerebrovascular neuropathology. Neuropathological conditions, such as cerebral ischemia, may affect the myogenic response, due to endothelial damage or myocyte-actin cytoskeletal depolymerization. 37 It is unclear whether various neuropathological conditions may alter calcium channel physiology and function. One study demonstrated differential effects of non-specific calcium channel antagonism based on calcium channel subtypes in the systemic circulation. Specifically, Liao and Soong demonstrated how alternative splicing of CaV 1.2 channels raises the possibility of expressing unique splice variants of calcium channels in different organ systems, under normal and non-physiologic conditions and even at different time points during a disease course. 108 Greater understanding as to how alternative splicing influences calcium channel pharmacology in different cerebrovascular disease processes may help to develop individualized therapies that minimize unintended consequences of less discriminate calcium channel antagonism given that the effects of non-specific calcium channel antagonism on specific calcium channel subtypes in the cerebrovasculature are unknown.
Inconsistent and unexpected findings from clinical studies of calcium channel antagonism further highlight the importance of gaining a more complete understanding of the cerebrovascular and neurological effects of these medications. For example, a recent clinical study assessing cerebrovascular hemodynamics using transcranial doppler imaging demonstrated a paradoxical distal cerebral vasoconstrictive effect of a dihydropyridine calcium channel antagonist, nicardipine, which may be modulated by the medication’s effects on cerebral venous circulation. 109 Similarly, studies using nimodipine, also a dihydropyridine calcium channel antagonist used in patients with aneurysmal subarachnoid hemorrhage, demonstrate modestly improved overall outcomes despite negligible effects on angiographic vasospasm. 110 These findings suggest that the neuroprotective effects of nimodipine may not be related to reducing vasospasm as was previously widely believed, but instead be due to neuroinflammatory benefits arising from its pleiotropic effects including decreased intracellular influx of calcium. 111
Conclusion
This review draws upon basic research and clinical studies to provide an in-depth analysis of the cerebrovascular effects of adrenoceptor and calcium channel modulating medications. The reviewed findings demonstrate a strong justification to consider cerebrovascular-specific effects of these vasoactive medications that may differ from their systemic effects.
There are important potential clinical implications of tailoring vasoactive medications to neurological endpoints. For example, several neurological and systemic conditions that are treated with vasoactive medications are highly comorbid with neurological impairments, including stroke, anoxic brain injury, after cardiopulmonary resuscitation, septic shock, and acute respiratory distress syndrome. Despite improvements in survival112,113 and biomarkers 114 of cardiac and neurological function, 115 the use of vasopressors in these conditions is associated with considerably high rates of neurocognitive impairment and it remains unknown to what extent this morbidity can be mitigated through neuroprotective paradigms of vasopressor selection. For example, vasopressors that prioritize perfusion of hippocampal and frontal cortical structures may result in improved neurocognitive clinical outcomes, such as delirium, which is exceedingly common in the acute care setting.
This review further provides a rationale to select vasoactive pharmacotherapies based on lesion location and consideration of the regional variability in distribution and density of adrenoceptor subtypes and calcium channels along the cerebrovasculature. Future investigations are needed to determine if pathology-directed or neurocentric selection of vasoactive drugs that modulate adrenoceptors and calcium channels can improve neurological outcomes in acute neurological and systemic conditions.
Footnotes
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Acknowledgement
We acknowledge Sam Gilchrist for his contribution to graphic design for Figures 1 and ![]() .
.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.