
Abstract
Normal brain homeostasis depends on the integrity of the blood–brain barrier that controls the access of nutrients, humoral factors, and immune cells to the CNS. The blood–brain barrier is composed mainly of brain endothelial cells. Forming the interface between two compartments, they are highly polarized. Apical/luminal and basolateral/abluminal membranes differ in their lipid and (glyco-)protein composition, allowing brain endothelial cells to secrete or transport soluble factors in a polarized manner and to maintain blood flow. Here, we summarize the basic concepts of apicobasal cell polarity in brain endothelial cells. To address potential molecular mechanisms underlying apicobasal polarity in brain endothelial cells, we draw on investigations in epithelial cells and discuss how polarity may go awry in neurological diseases.
During vertebrate evolution, the blood–brain barrier changed from being a mainly glial barrier and became an endothelial one.
1
In mammals, astrocytes still play an essential part in the blood–brain barrier, for instance, by ensheathing capillaries with their endfeet (Figure 1), synthesizing a layer of the basement membrane, and promoting barrier properties in endothelial cells. The physical barrier, however, is composed of endothelial cells, which are firmly connected by tight junctions and for which the rate of transcytosis is lower than in other tissues. At the cellular level, tight junctions form the border between the apical/luminal and the basolateral/abluminal plasma membrane. Separation of the apical/luminal and the basolateral/abluminal side of endothelial cells is a prerequisite for apicobasal polarity and is at the core of the blood–brain barrier that prohibits polar compounds, pathogens, and immune cells from reaching the CNS.
Key components of the blood-brain barrier. Apicobasal and planar cell polarity of endothelial cells differ in their orientation to the vessel axis.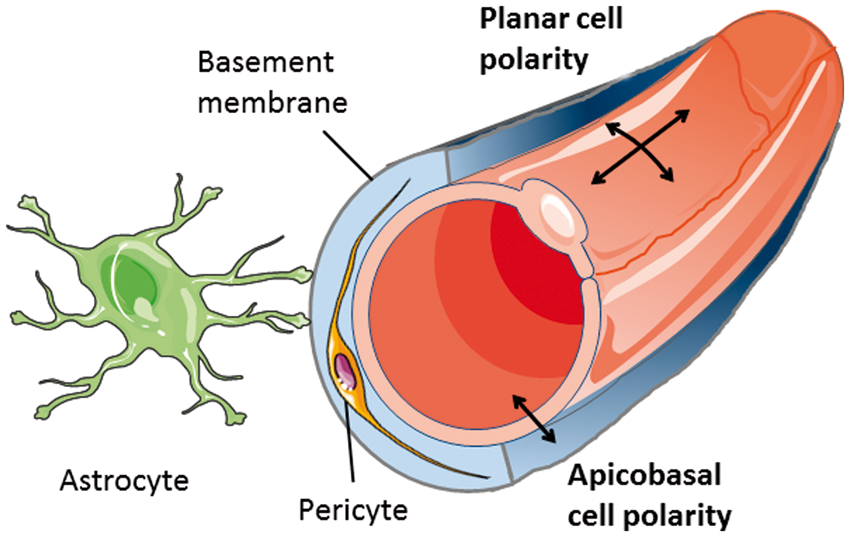
Forms of endothelial polarity
Apicobasal polarity is not the only form of endothelial cell polarity. Geometrically speaking, apicobasal polarity is set apart from planar cell polarity and the polarity of endothelial cells migrating during angiogenesis (Figure 1). At the molecular level the distinction is more blurred as some forms of polarity share certain pathways but have different orientation. WNT signaling, for example, is essential for the planar cell polarity program but is also involved in apicobasal polarity of brain endothelial cells. 2 The functional link between distinct polarity programs may facilitate the coordination of complex processes, such as angiogenesis, that involve both the polarity of migrating cells and apicobasal polarity. 3
With respect to endothelial polarity, many studies have focused on the polarity of migrating endothelial cells. Apicobasal polarity has been investigated in the context of lumen formation during development or angiogenesis4,5 but less is known about the molecular basis of apicobasal polarity in quiescent brain endothelial cells, despite the likely importance of this program for the blood–brain barrier. Here we summarize some key findings with the aim to demonstrate that much remains to be learned about apicobasal polarity programs in brain endothelial cells.
Why does apicobasal polarity matter?
By maintaining apicobasal cell polarity, brain endothelial cells accomplish an amazing task in very small dimensions. Understanding the underlying mechanisms is of general biological interest and, in addition, may help to elucidate the pathophysiology of neurological diseases and to improve their treatment.
As a physical border between the apical/luminal and the basolateral/abluminal cell compartments, intercellular junctions play an essential role in maintaining apicobasal cell polarity. 6 The complex relationship between intercellular junctions and apicobasal polarity involves two more aspects. Intercellular junctions provide docking sites for components of the apicobasal polarity program in the cell. Conversely, the polarity program regulates expression and function of intercellular junction proteins (see below). In many neurological diseases the continuous line of tight and adherens junctions is disrupted, leading to a loss of endothelial cell polarity and blood–brain barrier damage. 7 In MS lesions destabilized adherens junctions were linked to a disturbed apicobasal polarity of brain endothelial cells with increased apical/luminal localization of CXCL12 that may attract immune cells to invade the CNS. 8 Cerebral cavernous malformations seem to involve a primary defect in cell polarity. The genes that are mutated in familial cases, i.e. CCM1, CCM2, and CCM3, control cell polarity of brain endothelial cells9–11 (see below). Thus, correcting the polarity of brain endothelial cells may constitute a rational strategy for treating diverse neurological diseases.
On the other hand, the blood–brain barrier is an obstacle for treating brain diseases with macromolecules, such as recombinant proteins. Targeting brain endothelial cells with rAAV vectors or other tools may provide the means to express missing proteins in the CNS. Chen and colleagues 12 successfully used this approach in a mouse model of a lysosomal storage disease. After gene transfer lysosomal enzymes are released from endothelial cells and then taken up by neural cells. 13 To maximize the therapeutic potential of this approach, it is important to know how to direct transduced proteins to the correct side of brain endothelial cells and to understand polarized secretion.
Methods for studying apicobasal polarity of brain endothelial cells
In comparison to the situation in epithelium, endothelial polarity needs to occur in a miniaturized space. As brain endothelial cells are only about 200 nm thick,
14
membrane protein sorting must be very efficient. The close vicinity of the apical/luminal and basolateral/abluminal plasma membranes has posed problems when studying the localization of membrane proteins. In several cases contradictory results were obtained as to whether membrane transporters were localized in the apical/luminal or the basolateral/abluminal membrane (Table 1). Such discrepancies may be partially explained by the species or experimental details of the study but they are probably more often due to technical problems in localizing membrane proteins. The following methods have been used to localize proteins in subcellular compartments of brain endothelial cells.
Functional assays
When comparing the permeability of brain capillaries in vivo and in vitro, Betz and Goldstein 81 noted marked differences. After intravenous administration, hydrophilic compounds only have access to the apical/luminal side of the endothelium, whereas in vitro they come in contact with both the apical/luminal and basolateral/abluminal side of endothelial cells. The dependency of permeability on the side of administration led them to propose that brain endothelial cells are polarized. Asymmetry of brain endothelial plasma membranes has also been predicted from mathematical modeling of glucose and radioactive deoxy-glucose distribution under basal and stimulated conditions. 82
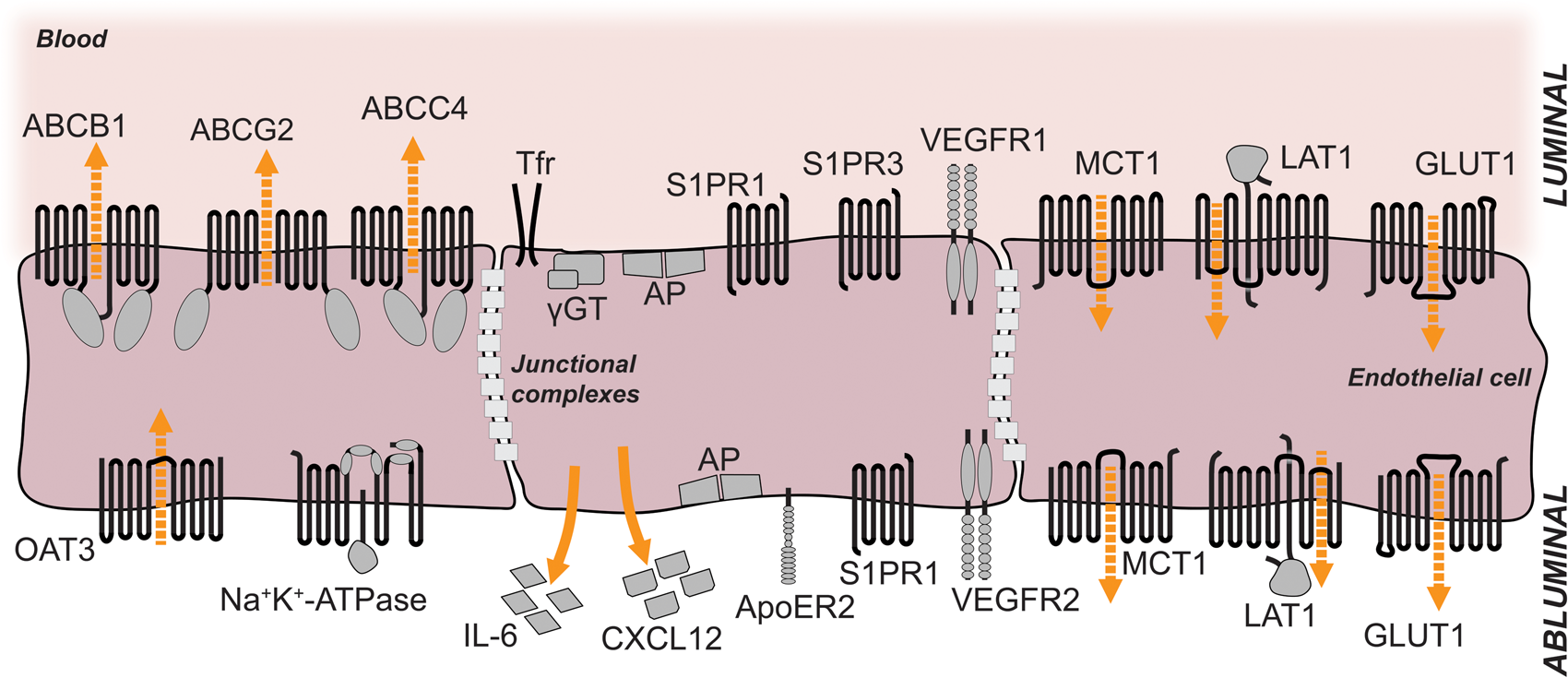
Importantly, the localization of transporters can be deduced from the accumulation of drugs in the brain of knockout mouse lines. The ABC transporter PGP (ABCB1, MDR1) pumps a wide range of compounds out of cells in an ATP-consuming manner (Figure 2). Thus, the direction of active transport depends on the localization in cell membranes. Localization only on the apical/luminal side of endothelial cells that links its activity to an outward transport of drugs across the blood–brain barrier explains accumulation of compounds in the brains of knockout mice.
83
According to the same line of evidence, MRP4 (ABCC4) and BCRP (ABCG2) are also mainly localized in the apical/luminal membrane of brain endothelial cells (Figure 2).35,84
Density-gradient centrifugation of membranes Localization of selected membrane proteins at the luminal and abluminal sides of brain microvessel endothelial cells. The luminal and abluminal membrane domains are separated by tight junctions. The sizes of the receptors and their respective domains are not drawn in scale. For details see tables 1 and 2. Orange arrows indicate direction of secretion; dashed orange arrows indicate direction of transport. ABCB1, ABCC4, ABCG2: ABC-binding cassette transporters; OAT3: organic anion transporter 3; Tfr: transferrin receptor; γGT: gamma-glutamyl-transferase; AP: alkaline phosphatase; S1PR1: sphingosine-1-phosphate receptor 1; S1PR3: sphingosine-1-phosphate receptor 3; VEGFR1: vascular endothelial growth factor receptor 1; VEGFR2: vascular endothelial growth factor receptor 2; IL-6: interleukin-6; CXCL12: C-X-C motif chemokine 12 = stromal cell-derived factor 1 (SDF-1); ApoER2: apolipoprotein E receptor 2 = low-density lipoprotein receptor-related protein 8 (LRP8); MCT1: monocarboxylate transporter 1; LAT1: large neutral amino acids transporter; GLUT1: facilitated glucose transporter member 1.
By using density-gradient centrifugation brain capillary membranes can be fractionated to obtain purified preparations of apical/luminal and basolateral/abluminal membranes.85,86 The strength of this technique lies in the fact that biochemical assays can be performed and detection does not rely exclusively on the specificity of antibodies. Using this approach, Lee and colleagues
87
found that the activity of GLUT1 was similar in the apical/luminal and basolateral/abluminal membranes of bovine brain endothelial cells although immunostains did detect it preferentially in the basolateral/abluminal membrane (Figure 2). Recently, a detergent-free technique was established.
88
Protein levels can be quantitatively measured in various species independent of available antibodies.89,90 However, identifying fractions depends on the enrichment of marker proteins; therefore, allocating individual fractions to cellular compartments may be ambiguous.
91
Often, the apical/luminal and basolateral/abluminal membranes can only be partially purified and glial or pericytic contamination is a concern.
86
Microscopic techniques
The resolution of conventional light microscopy (about 200 nm) is limiting when attempting to reliably distinguish between apical/luminal and basolateral/abluminal plasma membranes of brain endothelial cells. In the perinuclear space, the endothelial plasma membranes are so far apart that they can be identified.2,15 With the help of high-resolution confocal microscopy and quantitative colocalization studies, membrane proteins could be localized in subcellular compartments of brain endothelial cells.15,23,29,46 In the future, super-resolution microscopy may bring critical progress. So far, spectral precision distance microscopy/spectral position determination microscopy (SPDM), a super-resolution technique, has been used to localize PGP within the apical/luminal membrane of brain endothelial cells.
92
Up to now, subcellular localization of membrane proteins has largely relied on immunogold techniques in electron microscopy.
91
Despite the high potential and the relative ease in quantifying results, practical limitations result from the fact that antibody binding is often impaired by tissue fixation and processing. Indeed, electron microscopical techniques have produced conflicting data about the localization of proteins, e.g. PGP and GLUT1.16,30,76,91
In vivo labeling of apical/luminal proteins
Proteins in the apical/luminal membrane of brain endothelial cells can be labeled independently of antibodies by perfusing animals with a biotinylation reagent.
23
Biotinylated proteins in microvessel extracts are subsequently purified with the help of neutravidin-coated agarose beads. While this in vivo approach labels apical/luminal proteins, it does not detect proteins in the basolateral/abluminal membrane. Both basolateral/abluminal and intracellular proteins are included in the unlabeled fraction, which complicates the direct comparison of apical/luminal and basolateral/abluminal membranes.
In vitro studies of cell polarity
Multiple in vitro models of the blood–brain barrier have been developed that differ in how endothelial cells are cultured. 93 All of these models have in common a lack of some of the decisive external factors that induce polarity in the endothelial cells, including blood flow, the basement membrane, astrocytic endfeet, or pericytes (see below). These shortcomings lower the barrier quality of in vitro models compared to the in vivo situation. 93 Nevertheless, endothelial polarity is at least partially preserved in vitro, thus enabling analytical techniques that would be out of reach for in vivo studies. For example, culturing porcine brain capillary endothelial cells revealed that the concentration of phosphatidylcholines is higher and the concentration of sphingomyelins and glucosylceramides lower in the apical/luminal plasma membrane than in the basolateral/abluminal membrane. 94 In vitro studies also provide information about human cells. The immortalized human brain endothelial cell line hCMEC/D3 retains some degree of cell polarity, such as polarized localization of PGP predominantly in the apical/luminal membrane. 95 Importantly, in vitro models offer the opportunity to investigate polarized secretion13,96,97 (Table 2). To study polarized secretion, brain endothelial cells are plated on a permeable, porous membrane in a cell culture insert that has been coated with collagen IV or other basement membrane components. Once the endothelial layer has reached confluency, as shown by a high transendothelial resistance or a low permeability of tracers, e.g. luciferin yellow, concentrations of secreted factors in the two compartments are compared. The upper compartment corresponds to blood while the lower represents the brain parenchyma.
The apicobasal polarity of brain endothelial cells
As interface between blood and the vessel wall, endothelial cells are polarized in an apicobasal orientation. In the CNS, apicobasal polarity seems to be more pronounced than in other vascular territories in order to form the blood–brain barrier and to meet the metabolic demands of this highly active tissue. Apicobasal polarity of brain endothelial cells presents in several ways.
Differences in lipid and (glyco-)protein composition of the apical/luminal and basolateral/abluminal plasma membranes
Localization of membrane proteins in brain endothelial cells.

Note: The above table shows predominant localization of transmembrane or membrane-associated proteins in brain microvessel endothelial cells.
Both luminal and abluminal localization of PGP have been reported 76 in rat and human brain tissue sections using immunogold staining (electron microscopy).
Peripheral inflammatory hyperalgesia promotes translocation of PGP to the luminal membrane. 28
The apical localization of PGP depends on PAR-32.
An apical localization of MRP1 has been reported in primary cultured bovine brain microvessel endothelial cells using immunostaining and Western Blot analysis of plasma membrane fractions. 33
Both luminal and abluminal localization of MRP2 have been reported 16 in mouse brain tissue sections using immunofluorescence staining.
Both luminal and abluminal localization of MRP4 have been reported 34 in isolated rat brain microvessels using immunofluorescence staining. Both apical and basolateral localization of MRP4 have been reported in primary cultured bovine brain microvessel endothelial cells using immunostaining and Western Blot analysis of plasma membrane fractions. 33
In Bronger et al., 36 sections of brain gliomas were analyzed.
An abluminal localization of MRP5 has been reported 23 in isolated rat brain microvessels using immunofluorescence staining.
Both luminal and abluminal localization of Na+-K+-ATPase have been reported77,78 using cytochemical analysis (electron microscopy) of rat brain tissue sections.
ASCT1 expression is only detectable at embryonic and neonatal stages, not at adult stages. 45
GLUT1 is predominantly expressed at the abluminal membrane; however, significant levels of GLUT1 can also be found at the luminal membrane.
MCT1 expression shifts to the abluminal membrane during ketogenic diet. 62
An abluminal localization of OATP1A4 has been reported 23 in isolated rat brain microvessels using immunofluorescence staining.
Polar localization of PGT at the apical membrane is lost after treatment with LPS. 64
A luminal localization of alkaline phosphatase has been reported in mouse brain tissue sections using a cytochemical analysis (electron microscopy).67,79 Abluminal expression of alkaline phosphatase increases during scrapie infection. 67
Abluminal expression of thiamine pyrophosphatase increases during scrapie infection as shown by a cytochemical analysis (electron microscopy) of mouse brain tissue sections. 67
Abluminal expression of 5′-nucleotidase increases during scrapie infection as shown by a cytochemical analysis (electron microscopy) of mouse brain tissue sections. 67
Both luminal and abluminal localization of γGT have been reported 43 using measurement of enzyme activity in plasma membrane fractions of isolated cow brain microvessels.
Both luminal and abluminal localization of the transferrin receptor have been reported 80 by measuring the uptake of fluorescent immunoliposomes into isolated rat brain microvessels.
CCL2 induces the translocation of JAM-1 from tight junctions to the apical membrane in a RHOA- and ROCK-dependent manner. 74
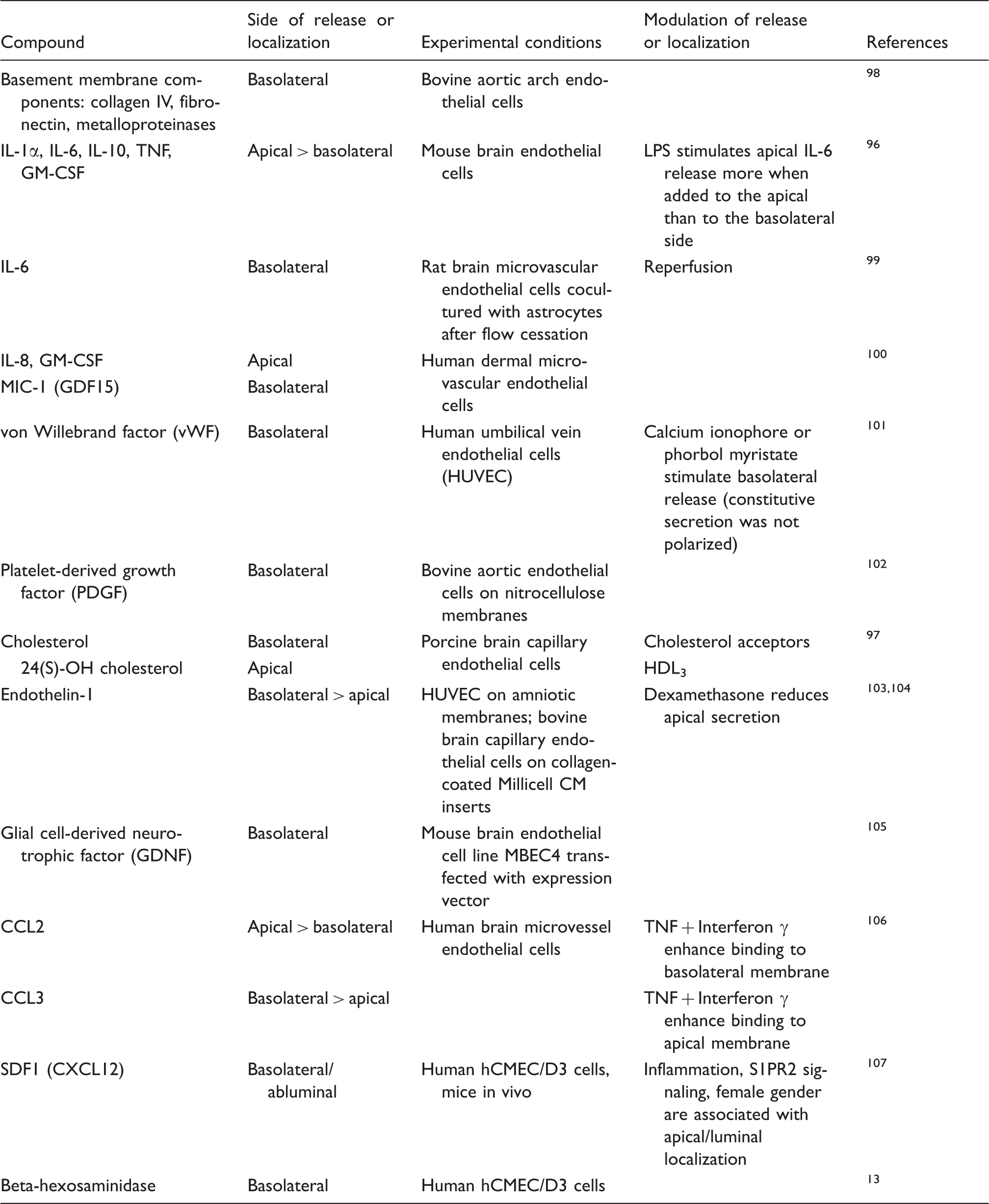
Factors released from endothelial cells in a polarized manner.
Directed transcytosis
Transcytosis involves different mechanisms of endocytosis. Best understood is the clathrin-mediated pathway that is responsible for transferrin transport through the blood–brain barrier.110,111 Among the clathrin-independent pathways, caveolae mediate the transport of albumin and other factors. Disruption of caveolae-dependent transcytosis seems to contribute to juvenile neuronal ceroid lipofuscinosis caused by mutations in ceroid lipofuscinosis neuronal-3 (CLN3).
112
Another only recently identified form of clathrin-independent endocytosis relies on endophilin, a protein that is expressed in brain endothelial cells.113,114 Target selectivity of transcytosis results from the involvement of membrane receptors that bind targets, such as insulin, leptin, cytokines, transferrin, and lipoproteins, and that are included in the endocytotic vescicle. The distribution of target receptors between the apical/luminal and basolateral/abluminal membranes represents an important determinant of the direction of transcytosis. In the case of transferrin, its receptor is found on the apical/luminal side of brain endothelial cells, largely limiting endocytosis to the blood side and transport in a blood-to-brain direction.
111
For bidirectional transport two receptors may be needed, as in the case of Aβ, which is transported out of the brain by LRP1 and into the brain by RAGE.115,116
Polarized release
For the differential interaction with blood and the parenchyma, endothelial cells secrete proteins and other factors in a polarized manner either to the apical/luminal or the basolateral/abluminal side (Table 2). In some cases, external stimuli were reported to control the side of secretion. As in epithelial cells, some factors, including von Willebrand factor (vWF), are packaged in clathrin-coated vesicles and released at the basolateral/abluminal side
117
(Table 2). However, the signals that direct secretion to the apical/luminal side are less well understood. Apparently, N-linked glycosylation is not involved, in contrast to what has been reported for epithelial cells.
100
Polarized response to stimuli
Cell polarity also influences the response of brain endothelial cells to external stimuli. The cytokines IL-6 and GM-CSF increase neuroinvasion of HIV-1 when applied to the apical/luminal, but not to the basolateral/abluminal side. 118 Vascular endothelial growth factor (VEGF), histamine, and insulin-like growth factor-binding protein 3 (IGFBP3) increase the transendothelial permeability of the blood–brain barrier when administered to the basolateral/abluminal, but not to the apical/luminal side. 72 In contrast, thrombin and bradykinin are effective on both sides and lysophosphatidic acid (LPA) mostly on the apical/luminal side and less on the basolateral/abluminal side. Further investigations demonstrated that VEGF binds to VEGFR2 on the basolateral/abluminal membrane and increases the permeability through p38 kinase, whereas on the apical/luminal side VEGFR1 stimulates Akt and facilitates cytoprotection. Intriguingly, the polarized response to VEGF is found exclusively in brain and retinal, but not in peripheral endothelial cells, highlighting the special status of endothelial cells in the CNS.
Epithelial polarity program
While the molecular mechanisms governing endothelial apicobasal polarity are just beginning to emerge, a wealth of literature has reported on epithelial apicobasal polarity regulation. Epithelial apicobasal polarity is established and maintained by a molecular network of polarity protein complexes, adhesion complexes, small GTPases, and lipids. Fundamental concepts of epithelial polarity were initially discovered in flies and worms; albeit more complex, many of these concepts also apply to vertebrate cells. In the following paragraph, we will introduce important molecular determinants of epithelial polarity and their interplay. For more detailed information on this topic, we refer to several excellent recent reviews.119–124
Polarity complexes in epithelial cells
Par complex
The Par proteins comprise a family of structurally and functionally distinct cytoplasmic proteins, which were identified as genes essential for the polarity of the early C. elegans embryo.
125
They are evolutionarily conserved in other animals and widely expressed. Whereas all Par proteins are encoded by single genes in Drosophila and C. elegans, some proteins have several isoforms in vertebrates, which might carry out specific functions in particular cell types. Par proteins have been shown to function as central regulators not only of epithelial polarity, but also of endothelial polarity, particularly in angiogenesis.126,127 They comprise the following members: the serine/threonine protein kinases, PAR-1 ( = microtubule affinity-regulating kinase 2, MARK2, in vertebrates), and PAR-4 ( = Liver kinase B1, LKB1 or Serine/threonine-protein kinase 11, STK11, in vertebrates); the scaffold and adapter proteins PAR-3 (two isoforms, PAR-3 and PAR-3 L, also known as PAR-3B, in vertebrates) and PAR-6 (three isoforms, PAR-6 A = PAR-6 C, PAR-6B, and PAR-6D = PAR-6 G in vertebrates); the adapter protein PAR-5 (a 14-3-3 isoform). The serine/threonine protein kinase atypical PKC (two isoforms, aPKCζ and aPKCι/λ in vertebrates) and the small GTPase CDC42 (see also below) are closely linked to Par proteins both physically and functionally; together with PAR-3 and PAR-6, they are therefore referred to as the “Par complex”. The membrane localization of Par proteins is established and maintained by a system of reciprocal exclusion119,120,128 (Figure 3(a)): aPKC, which forms a complex with PAR-6 and CDC42 at the apical membrane, phosphorylates PAR-1 and other basolateral proteins to exclude them from the apical membrane, while at the basolateral membrane PAR-1 phosphorylates PAR-3 and apical proteins to exclude them from the basolateral membrane. The adapter protein PAR-5 binds proteins phosphorylated by aPKC and PAR-1 to detach them from the plasma membrane into the cytosol, where they are dephosphorylated, and thereby allows shuttling to the correct membrane domain. Par proteins form a highly interconnected network with other polarity proteins. For example, aPKC also phosphorylates LGL2, a member of the Scribble family of polarity proteins (see below), to restrict it to the basolateral membrane.
129
PAR-6 binds to CRB3 and PALS1, members of the Crumbs family of polarity proteins (see below), which confer the anchoring of PAR-6 to the apical membrane.
130
Moreover, PAR-4 phosphorylates and activates AMPK to control cell metabolism and growth.
131
Epithelial and endothelial apicobasal polarity. (a) Molecular mechanisms of epithelial apicobasal polarity. For details see text. The straight arrow indicates activation, straight dashed arrows indicate phosphorylation, and curved arrows indicate enzymatic reactions. (b) Molecular mechanisms that establish endothelial apicobasal polarity during vascular lumen formation. Only molecules for which functional roles have been demonstrated in vivo are depicted. For further details see text. Similar to epithelial cells, β1 integrin-matrix interactions provide an initial cue to establish endothelial apicobasal polarity.60 β1 integrin is required for proper PAR-3 expression levels and for the correct localization of VE-cadherin at lateral cell-cell contacts.60 VE-cadherin, which can bind directly to PAR-3,202 forms a complex with CCM1.9 CCM1 stabilizes VE-cadherin at adherens junctions to further establish and maintain endothelial apicobasal polarity.9,203 Rasip1 and its binding partner Arhgap29 suppress the activity of the small GTPase RHOA to promote integrin-mediated adhesion, and to regulate the correct localization of PAR-3, as well as of intercellular junctions.204 In contrast to epithelial cells, the organization of tight and adherens junctions in endothelial cells is less clearly defined, with tight and adherens junctions frequently being intermingled. CCM1: cerebral cavernous malformations 1; Rasip1: Ras interacting protein 1.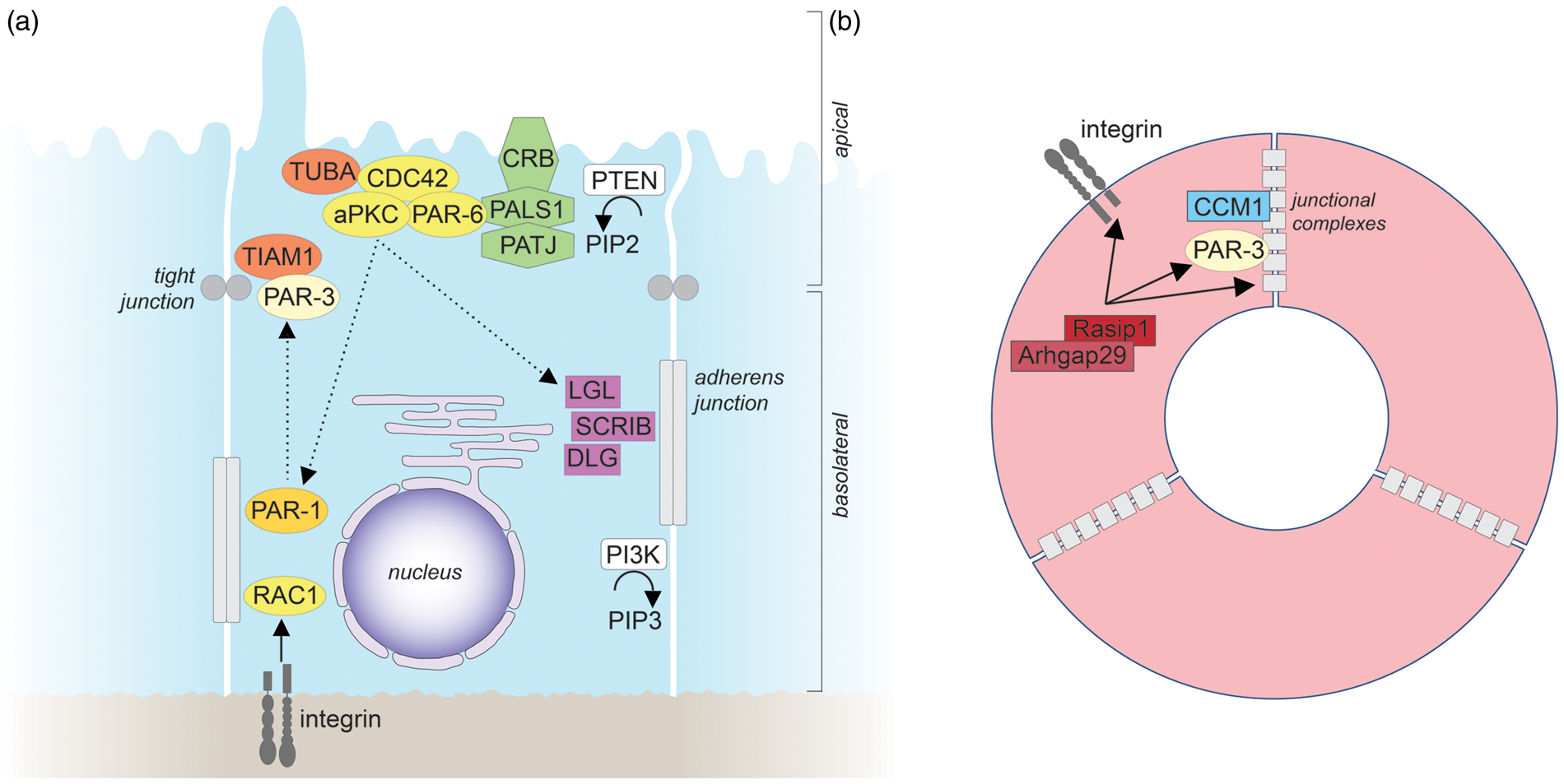
Crumbs complex
Crumbs proteins are apically localized, single-pass transmembrane proteins that were initially identified in Drosophila. 132 In vertebrates, three different Crumbs isoforms (CRB1, CRB2, and CRB3) with largely non-overlapping expression patterns are found, CRB3 being the major isoform in epithelial cells. 133 Via their C-terminus, Crumbs proteins bind PALS1 (protein associated with Lin-7 1), an adapter protein that forms a complex with the multi-PDZ domain protein PATJ (PALS1-associated tight junction protein) (Figure 3(a)). Crumbs proteins are important determinants of apical membrane identity and play a central role in tight junction assembly and maintenance.119,133 Very little is known about the role of Crumbs proteins in endothelial cell polarity. Recent evidence in primary endothelial cells in vitro suggests that Crumbs proteins might be involved in regulating endothelial cell junctions. 134
Scribble complex
The Scribble family of polarity proteins was first identified in Drosophila as important regulator of apicobasal cell polarity and tissue growth. Loss of any of these genes in Drosophila results in the mistargeting of apical proteins to the basolateral membrane, the disorganization of adherens junctions, and tumor formation. 135 The Scribble protein complex consists of Scribble (SCRIB), Discs Large (DLG; several isoforms in vertebrates), and Lethal Giant Larvae (LGL; several isoforms in vertebrates), which all localize to the basolateral membrane (Figure 3(a)). The basolateral localization of LGL depends on its phosphorylation and exclusion from the apical membrane by aPKC. 136 In vertebrate epithelial cells, SCRIB is required for E-cadherin–mediated cell–cell adhesion. 137 While the role of the Scribble complex in endothelial cell polarity is largely unexplored, one recent study shows that SCRIB is expressed in primary endothelial cells and involved in endothelial cell migration. 138
RhoGTPases in epithelial polarity
Small GTPases of the Rho family have been extensively studied for their role as master regulators of the actin cytoskeleton. A large body of evidence indicates that these GTPases, particularly CDC42 (see also above) and RAC1, are centrally involved in epithelial apicobasal polarity. 139 CDC42, a master regulator of epithelial cell polarity, is activated by TUBA, an apically localized guanine nucleotide exchange factor (GEF) for CDC42, and positively regulates the activity of aPKC to promote apical membrane identity and the formation of adherens junctions, and to control vesicle trafficking6,119,129,140,141 (Figure 3(a)). RAC1, which often cooperates functionally with CDC42, is enriched basolaterally, is required for the assembly of the basement membrane, and is crucially involved in the formation and maintenance of E-cadherin-based adherens junctions. 119 Activation of RAC1 is triggered by β1 integrin, E-cadherin, or the RAC GEF TIAM1 that interacts directly with PAR-3.119,142,143 Moreover, CDC42 and RAC1 activity can be regulated by SCRIB, which binds and activates the RAC/CDC42 GEF βPix; 144 however, the significance of the SCRIB/βPix interaction for epithelial polarity is still unclear. RHOA, which frequently acts antagonistically to RAC, controls actomyosin contractility through its downstream effector kinase ROCK. 139 Furthermore, ROCK can phosphorylate PAR-3 to disrupt its association with aPKC and PAR-6. 145 Conversely, aPKC phosphorylates ROCK to exclude it from apical junctional complexes in order to prevent excessive apical constriction. 146 CDC42, RAC1, and RHOA are also crucial for endothelial apicobasal polarity and formation of endothelial junctions (see below).
Lipids in epithelial polarity
Phosphoinositides have been shown to play a central role in epithelial cell polarity by determining the identities of apical and basolateral plasma membranes. 147 Whereas phosphatidylinositol 4,5-bisphosphate (PIP2) localizes to the apical membrane, phosphatidylinositol 3,4,5-trisphosphate (PIP3) is enriched at the basolateral membrane6,148 (Figure 3(a)). Insertion of exogenous PIP2 into the basolateral membrane of epithelial cells causes the mistargeting of apical proteins to the basolateral membrane, while, conversely, insertion of exogenous PIP3 into the apical membrane results in the formation of membrane protrusions, which contain basolateral proteins.6,148 Via an adapter protein, PIP2 binds to CDC42 and recruits it to the apical membrane. 6 The polar distribution of PIP2 and PIP3 is established and maintained by two enzymes, phosphatidylinositol-3,4,5-trisphosphate 3-phosphatase (PTEN) and phosphatidylinositol 3-kinase (PI3K), which localize to the apical and basolateral membrane, respectively.6,149 At the apical membrane, PTEN catalyzes the formation of PIP2 and the removal of PIP3. 6 At the basolateral membrane, the activity of PI3K results in the conversion of PIP2 into PIP3. 149 Like epithelial cells, endothelial cells – including cultured brain capillary endothelial cells – show an asymmetric distribution of lipids between the luminal and the abluminal membrane. 94 However, whether lipids play a role in establishing endothelial polarity is so far unknown.
Generation and maintenance of epithelial polarity
The initial cues for the generation of epithelial polarity are provided by epithelial cell-cell contacts and extracellular matrix proteins, which directly activate receptor-mediated signal transduction pathways and, in addition, act through mechanical forces and cell confinement. 122 Early cell-cell contacts induce the formation of intercellular junctions, which recruit and activate polarity proteins. In turn, in a complex interplay, polarity proteins regulate the further maturation of adherens and tight junctions. 120 The formation and maintenance of junctions is tightly coupled to the reorganization of the cytoskeleton and depends in particular on actin dynamics. 119 At the same time, activation of β1 integrin by collagen results in RAC1 activation, which leads to the secretion of laminin and assembly of the basement membrane. These processes promote the establishment of apical and basal membrane domains. 120 The identities of the apical and basolateral membrane further critically rely on a polarized trafficking machinery that drives the targeted transport of vesicles carrying distinct lipids and proteins. 122 The delivery of these lipids and proteins to the correct plasma membrane is ensured by diverse apical and basolateral sorting signals and mechanisms. 122 Tight junctions that prevent diffusion of proteins and lipids from one membrane domain to the other are required to maintain membrane identity. 120 Finally, the polarization of individual cells is translated into the arrangement of higher-order cell structures to build the proper epithelial tissue architecture.
Mechanisms of endothelial polarity
Studies in epithelial cells have largely formed our current concepts of apicobasal cell polarity. However, how polarity is achieved may differ among cell types. MRP4, for example, is localized in the basolateral membrane of epithelial cells in the choroid plexus, but in the apical/luminal membrane of brain capillaries. 35 Endothelial polarity has often been studied in the context of lumen formation of blood vessels, some key findings of which are depicted in Figure 3(b). In the following we will summarize what is known about the mechanisms that control polarity in endothelial cells, with a special focus on the brain.
Par complex in endothelial polarity
The Par complex (PAR-3, PAR-6, aPKC, and CDC42) is involved in endothelial polarity during lumen formation 126 (Figure 3(b)). Components of this complex are expressed in brain endothelial cells.2,150,151 The Par complex is localized to tight junctions of endothelial cells by binding PAR-3 to JAM proteins.2,152 In addition, PAR-3 binds to VE-cadherin, explaining why PAR-3, phosphorylated aPKCζ, and TIAM localize to intercellular junctions, depending on the presence of VE-cadherin. 9 When meningococci adhere to brain endothelial cells, the Par complex moves away from intercellular junctions to the site of meningococcal adhesion, disturbing tight junction function and increasing paracellular permeability. 150 Knockdown of PAR-3 interferes with the morphology and function of tight junctions as well as with apicobasal polarity and lumen formation.2,9 Interestingly, WNT5a, a growth factor expressed by astrocytes, seems to enhance PAR-3 levels in intercellular junctions and to increase apicobasal polarity,2,153 suggesting that astrocytes promote endothelial polarity via this pathway.
Crumbs complex in endothelial polarity
Components of the Crumbs complex (PALS1, PATJ) are expressed in endothelial cells. 134 The complex is localized to tight junctions of endothelial cells. By binding SYX, a RHOA-specific GEF that activates RHOA, members of the Crumbs complex stabilize intercellular junctions and decrease paracellular permeability. 134 SYX deficiency impairs normal vascular development. 154 The Crumbs complex also interacts with angiomotin (Amot) and Angiomotin-like Protein 1 (AmotL1). 155 Both proteins are localized in tight junctions. Knockdown of Amot and AmotL1 increases paracellular permeability, suggesting that Amot and AmotL1 are required for tight junctions to function properly. They are also essential for the polarity of migrating endothelial cells. 155 Amot knockout mice have a high embryonic lethality and develop dilated vessels in the brain. 156 Less is known about the role of Amot or AmotL1 in apicobasal polarity beyond formation of tight junctions.
Scribble complex in endothelial polarity
Members of the Scribble complex are found in brain endothelial cells. DLG1 is expressed in endothelial cells during mouse development and in the human brain endothelial cell line hCMEC.157,158 In brain endothelial cells, DLG1 is not colocalized with SCRIB, suggesting that the two factors do not form a complex as in Drosophila epithelial cells. 117 By binding to GPR124 as a cofactor, DLG1 enhances canonical WNT signaling in response to WNT7a. 157 The latter is released by neuroepithelial cells and has a profound effect on the development of the blood–brain barrier.159,160 Other interaction partners of DLG1 include clathrin, AP-1, and von Willebrand factor. 117 Knockdown of DLG1 inhibited the formation of Weibel-Palade bodies and induced disassembly of the trans-Golgi network. SCRIB mediates planar cell polarity and migration of endothelial cells. 138 Mice with a frameshift mutation in SCRIB (circle tail mice) or zebrafish treated with SCRIB morpholinos developed vascular malformations in the brain and hemorrhages 138 that are reminiscent of the phenotype of mice deficient in LGL1. 161 However, it is still unclear whether endothelial SCRIB or LGL1 affects CNS development. Thus, despite evidence that mammalian homologues of SCRIB, DLG, and LGL play a role in endothelial cells, how they contribute to apicobasal polarity is still largely unclear.
CCM proteins in endothelial polarity
An important regulatory pathway for endothelial polarity was uncovered through the study of cerebral cavernous malformations. These enlarged, thin-walled, and leaky vascular structures consist mainly of endothelial cells with only few glia or pericytes. Three structurally unrelated genes are mutated in familiar cases of the disease, CCM1 (Krit1), CCM2 (OSM), and CCM3 (PDCD10). CCM1 is a binding partner and effector of the small G protein RAP1 that is involved in epithelial polarity programs and tightens endothelial barriers.162–164 Knockdown of RAP1 impairs apicobasal polarization and lumen formation of endothelial cells. 9 Upon activation RAP1 mediates the translocation of CCM1 from microtubule to adherens junctions, where CCM1 associates with β-catenin and VE-cadherin.164,165 Loss of CCM1 causes β-catenin to dissociate from VE-cadherin.9,166 Consequently, the released β-catenin translocates to the nucleus and stimulates gene transcription, whereas adherens junctions are disrupted. 166 In CCM1-deficient endothelial cells, the lack of junctional VE-cadherin that normally recruits the Par complex (see above) results in a defect in apicobasal polarity and lumen formation. 9
CCM1 forms a complex with CCM2 and CCM3. CCM2 deletion in endothelial cells causes a defect in lumen formation of branchial arch arteries during development but does not affect apicobasal polarity in endothelial cells.11,167 In addition, deletion of CCM3 interrupts lumen formation but not the development of branchial arch arteries.168,169 At the biochemical level, CCM2 is required for maintaining the cytoskeleton and limiting RHOA activation. 10 CCM2 deficiency leads to low levels of CDC42, a component of the Par complex. 11 The effect of CCM3 on RHOA activation is the subject of controversy.169,170 Instead, CCM3 is located on the Golgi apparatus and stabilizes the germinal center kinase III (GCKIII). 171 In support of a pathogenic role of GCKIII in CCMs, overexpression of the GCKIII kinase STK25b rescued the vascular defects in zebrafish deficient in CCM3. 168 Furthermore, deletion of STK25b mimicked the effect of CCM3 deletion. The presence of CCM3 in the Golgi apparatus seems to determine Golgi orientation, 171 which itself has implications for cell polarity and polarized secretion, at least in the epithelial HeLa cell line. 172 So far, very little is known about the mechanisms underlying polarized secretion in endothelial cells. As epithelial and endothelial cells secrete some proteins in different directions,111,173,174 concepts that have been established to explain the mechanisms underlying polarized secretions in epithelial cells may not be true for endothelial cells.
RhoGTPases and shear stress in endothelial polarity
The small GTPases RHOA, RAC, and CDC42 control brain endothelial polarity in a complex manner. A physiological activator of all three is shear stress, as reviewed previously. 175 The spatial distribution of activity in response to flow is polarized in the cell plane with CDC42 and RAC1 activation at the downstream edge of the cell. RHOA is first inhibited and then activated by flow.176,177 Other stimuli of RHOA include inflammatory mediators that increase the permeability of the barrier, such as LPS, 178 thrombin, 179 and CCL2. 180 In contrast, inhibitors of RHOA, such as ANXA1 which acts via G protein-coupled FPR2 receptors178,181 and CCM1 or CCM2, stabilize barrier function.10,11 During shear stress RHOA is involved in cell alignment and stress fiber formation in endothelial cells. 177 The RHOA-mediated increase in blood–brain barrier permeability has been attributed to increased phosphorylation of the myosin light chain and contraction of actomyosin, pulling intercellular adhesions apart. 182 Additionally, RHOA enhances the phosphorylation of tight junction proteins, which leads to their disintegration. 183 RHOA modulates the apicobasal polarity of brain endothelial cells by redirecting JAM-1 from tight junctions to the apical/luminal surface, where it functions as a leukocyte adhesion molecule. 74 Through this mechanism RHOA stimulates the transendothelial migration of neutrophils and monocytes. RHOA is inhibited by CCM1, CCM2, and CCM3 (see above). 184 Thus, a deficiency in any of the CCM proteins leads to RHOA overactivation. Interestingly, the increased endothelial permeability in CCM1- or CCM2-deficient mice could be ameliorated by fasudil, a pharmacological blocker of the RHOA effector ROCK. 10
In contrast to the destabilizing effects of RHOA on the barrier, CDC42 enhances cell polarity and barrier properties. 185 As part of the Par complex, it is normally located at intercellular junctions. Adhesion of meningococci to the apical membrane of brain endothelial cells relocates the Par complex to the site of adhesion and CDC42 plays an active part in this process. 150 CDC42 also promotes endothelial lumen formation, a process that depends on other members of the Par complex. 186
External stimuli of endothelial polarity in the blood–brain barrier
External cues provide spatial orientation for brain endothelial cells to maintain their apicobasal polarity. On the basolateral/abluminal side, astrocytes and pericytes are found in close proximity. Astrocytes induce barrier properties and promote the apicobasal polarity of brain endothelial cells.187,188 Sonic hedgehog is one of the factors released by astrocytes; it activates the receptor Patched-1 on brain endothelial cells and stimulates the expression of tight junction components, such as claudin-5 and occludin. 189 Other factors that are produced by astrocytes and stabilize the barrier include angiotensin-II, GDNF, angiopoietin 1,188,190 and possibly WNT5a.2,153
More recently, an important influence of pericytes on the basolateral/abluminal side of brain endothelial cells was recognized.191–193 Pericytes are not only essential for establishing the blood–brain barrier, but also for maintaining it in adulthood. They induce the expression of the tight junction proteins ZO-1 and occludin in brain endothelial cells and thus reduce paracellular permeability. 192 Moreover, pericytes control the expression of MFSD2a in neighboring endothelial cells. 194 MFSD2a is a transmembrane protein localized in the apical/luminal membrane of brain endothelial cells. It functions as a transporter for the ω3 fatty acid docosahexaenoic acid that is esterified with lysophosphatidylcholines 195 and at the same time reduces transcytosis across the blood–brain barrier. 194 Pericytes and endothelial cells are in direct cell-cell contact at so-called peg-and-socket junctions that contain N-cadherin. 196 These specialized junctions may be the site where membrane receptors interact directly, e.g., Notch3 on pericytes and DLL4 on endothelial cells. 197
Outside of peg-and-socket junctions, pericytes and endothelial cells are separated by the endothelial sheath of the basement membrane, consisting of collagen type IV, perlecan, the laminins 411 and 511, and other proteins. Collagen IV binds to β1 integrins (α1β1, α2β1), which play an essential role in endothelial polarity and arterial lumen formation and in the development of intercellular junctions (Figure 3(b)).126,198 β1 integrin seems to be located upstream of PAR-3 because PAR-3 levels were lower in endothelial cells deficient in β1 integrin and their phenotype could be partially rescued by replacing PAR-3. 126
On the apical/luminal side brain endothelial cells are continuously exposed to a shear stress of 1 – 10 N/cm2 in capillaries. 199 Applying this level of shear stress to brain endothelial cells in vitro increases their barrier properties and the expression of tight junction components.93,200,201
Conclusion
Apicobasal polarity seems to be a cellular hallmark of the blood–brain barrier. So far the integrity of intercellular junctions as an isolated aspect of cell polarity has received the most attention. Other important aspects, including the polar distribution of membrane proteins, were less frequently investigated in a functional or pathophysiological context. Largely unclear are the molecular mechanisms that determine cell polarity in adult brain endothelial cells. However, important clues come from the study of endothelial lumen formation during development or the analogy of epithelial cells, where apicobasal polarity has been extensively studied. An in-depth investigation of apicobasal polarity could open new windows to understanding the blood–brain barrier. Progress will likely depend on improved genetic tools to manipulate brain endothelial cells in vivo and on high resolution fluorescence microscopy.
Footnotes
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.