
Abstract
Activating mutations of the leucine-rich repeat kinase 2 (LRRK2) gene are associated with Parkinson disease (PD), prompting development of LRRK2 inhibitors as potential treatment for PD. However, kidney safety concerns have surfaced from LRRK2 knockout (KO) mice and rats and from repeat-dose studies in rodents administered LRRK2 inhibitors. To support drug development of this therapeutic target, we conducted a study of 26 weeks’ duration in 2-month-old wild-type and LRRK2 KO Long-Evans Hooded rats to systematically examine the performance of urinary safety biomarkers and to characterize the nature of the morphological changes in the kidneys by light microscopy and by ultrastructural evaluation. Our data reveal the time course of early-onset albuminuria at 3 and 4 months in LRRK2 KO female and male rats, respectively. The increases in urine albumin were not accompanied by concurrent increases in serum creatinine, blood urea nitrogen, or renal safety biomarkers such as kidney injury molecule 1 or clusterin, although morphological alterations in both glomerular and tubular structure were identified by light and transmission electron microscopy at 8 months of age. Diet optimization with controlled food intake attenuated the progression of albuminuria and associated renal changes.
Keywords
Introduction
Parkinson disease (PD) is a major neurodegenerative disease that poses a substantial burden on health care systems. PD is characterized by functional motor impairment and accumulation of proteinaceous inclusions (Lewy bodies) in the brain associated with progressive loss of dopamine-dependent neurons. 1 Since 2004, multiple activating mutations of the leucine-rich repeat kinase 2 (LRRK2) gene have been identified and shown to be associated with dominantly inherited PD and recently shown to be present in sporadic cases of PD.2 -4 The most common LRRK2-activating mutation is LRRK2 G2019S, which accounts for approximately 4% of familial PD and 1% of sporadic PD across all populations. This mutation exhibits a penetrance rate of 28% at 59 years, 51% at 69 years, and 74% at 79 years of age. 5
The ROCO family proteins are a group of proteins that contain the Roc and COR domains. LRRK2 is a 280-kDa, multidomain protein of the ROCO family that contains multiple protein scaffolds which include the N-terminus armadillo (ARM), the ankyrin repeat (ANK), the leucine repeat (LRR) domain, and the C-terminus WD40 domain. The enzymatic core of LRRK2 comprises the Ras of complex (Roc) GTPase domain and a kinase domain bridged by C-terminus of Roc (COR). The biological functions of LRRK2 are not entirely understood, but LRRK2 has been shown to have critical activity in vesicular function and cytoskeletal structures in a variety of cell types.2,4,6 The fact that dominant mutations that enhance LRRK2 kinase activity are associated with PD has sparked interest in developing LRRK2 inhibitors as a potential treatment for PD.1,7 -9
Homozygous LRRK2 knockout (KO) mice showed impairment of protein degradation pathways, accumulation of α-synuclein, apoptotic cell death,10,11 and an increase in the number and size of secondary lysosomes in the proximal tubular epithelium of the kidney or presence of enlarged lamellar bodies in type II cells of the lung. 12 Homozygous LRRK2 KO Long-Evans Hooded (LEH) rats displayed significant weight gain, histopathological alterations in the renal tubular epithelial cells such as hyaline droplets, cytoplasmic vacuolation, and brown pigment accumulation,13 -15 as well as abnormalities in the lung.14,16 Treatment of Sprague Dawley (SD) rats with the LRRK2 inhibitor PFE-360 induced darkened kidneys and progressive accumulation of hyaline droplets in the renal proximal tubular epithelium that were reversible, but with no effect on the lung. 7 Treatment with the LRRK2 inhibitor MLi-2 in the MitoPark mouse model was associated with very slight enlargement of randomly scattered alveolar epithelial cells that had features consistent with type II pneumocytes, but no adverse effects were noted in the kidney. 8 The findings in kidneys and lungs observed in LRRK2 KO rodents and other preclinical studies highlight the need for further investigation to determine human relevance and establish methods to monitor safety for the development of LRRK2 inhibitors.
For monitoring drug-induced effects on renal function, a set of urinary protein biomarkers, including albumin (ALB), kidney injury molecule 1 (KIM-1), and clusterin (CLU), have demonstrated better correlation with early histopathological findings in kidneys of preclinical species when compared with conventional serum biomarkers blood nitrogen urea (BUN) and serum creatinine (sCr).17 -20 However, no accessible biomarker(s) have been established or shown to be effective in monitoring renal function following administration of LRRK2 inhibitors in preclinical species.
In previous pilot studies, we retrospectively evaluated ad libitum–fed LEH LRRK2 KO rats at 5, 15, or 39 weeks of age to ascertain progression of morphologic changes in kidneys with advancing age. These evaluations identified renal changes in KO rats characterized by both glomerular and tubular alterations progressing to end-stage renal disease. These results were leveraged to design a pivotal prospective study comparing progression of light microscopic and ultrastructural renal changes in wild-type (WT) and KO rats, including analyses of urinary renal safety biomarkers ALB, KIM-1, and CLU and an assessment of the potential impact of dietary modifications. The kidney safety biomarkers ALB, KIM-1, and CLU were chosen for their combined overall coverage of both glomerular and tubular injuries in rats.17,18
It is known that spontaneous chronic nephropathy occurs in ad libitum–fed SD rats starting between 3 and 6 months of age, 21 and that dietary modifications attenuate the progression of kidney injury.21,22 As we intended to maintain the rats in this study until 8 months of age, a cohort with dietary optimization was included in the study design to minimize the impact of food intake on the progression of spontaneous renal changes in the LEH rat strain. Furthermore, the number of rats per group was set at 8 to facilitate differentiation of renal findings attributable to lack of LRRK2 gene expression compared with the occurrence of spontaneous and age-related renal changes in WT cohorts.
We report a detailed account of the outcomes on a set of urine biomarker (ALB, KIM-1, CLU) in aging LRRK2 KO rats for up to 8 months, along with morphological endpoints by light microscopic and ultrastructural evaluations conducted at 5 or 8 months of age, as well as the impact of dietary optimization on these outcomes.
Materials and Methods
The studies were approved by the Institutional Animal Care and Use Committee (IACUC) of Merck & Co., Inc., West Point, PA, USA, and were conducted in a facility accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC) in compliance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and the Animal Welfare Act. WT and LRRK2 KO (LEH- Lrrk2tm1sage, TGRL4620) LEH rats were purchased from SAGE Labs, Inc (Boyertown, Pennsylvania). The rats received PMI-certified rodent diet #5002, and tap water was provided ad libitum. The rats were acclimated for at least 7 days before sample collections. Vendor designated genotypes of all rats were further confirmed by conducting polymerase chain reactions (PCRs) and examining genotype-specific sizes of PCR amplicons (data not shown). For genotyping, DNA was extracted from blood using Qiagen DNeasy blood and tissue kits, and the sequences of the PCR primer set were CCGTGAGGAATTCTACAGCAC and GCTTCATGGCATCCACCT.
Study Design
In all, 40 control and 40 LRRK2 KO rats were included for this study, with 24 female and 16 male rats of each genotype (Table 1). These rats were born within an eleven-day window, and the median of this date range was considered as the birth date for each rat. These rats were approximately 7 weeks of age at the start of the study. Two different feeding regimens were used. An ad libitum group had no limit to the amount of food consumed, while the optimized diet group received 16 or 22 g of food/day for females or males, respectively. The quantity of food allowance of the optimized diet group was selected based on the effectiveness of dietary restriction on preventing chronic nephropathy in SD rats. 21 Sixteen female and eight male rats of each genotype were assigned to the ad libitum group, while eight of each gender were assigned to the optimized diet group (Table 1). ALB levels in urine were monitored monthly starting from 2 months of age until the end of the study when rats were eight months of age. When 75% of the female LRRK2 KO rats in the ad libitum group surpassed the urine ALB threshold of 300 µg/mg uCr, both WT and LRRK2 KO female rats on ad libitum feeding were ranked from high to low according to their urine ALB concentrations to guide selection of rats for termination. Half of these WT and LRRK2 KO female rats (8 rats ranked at positions 1, 4, 5, 8, 9, 12, 13, and 16) were terminated at approximately 22 weeks (5 months) of age (interim necropsy). The remainder of the rats in the study were euthanized at final necropsy when they reached approximately 33 weeks (8 months) of age (Table 2). A complete gross examination was conducted for each rat at termination.
Study design and progression of albuminuria.
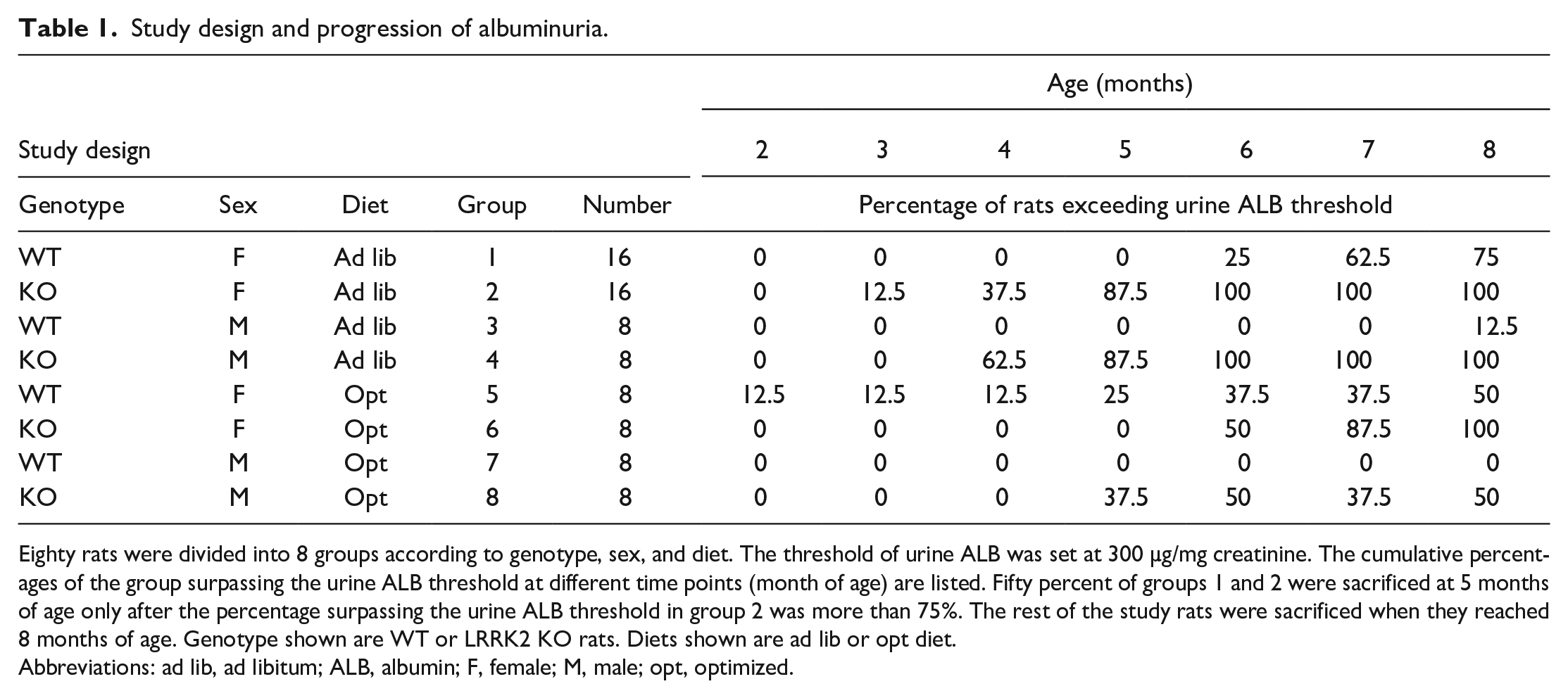
Eighty rats were divided into 8 groups according to genotype, sex, and diet. The threshold of urine ALB was set at 300 µg/mg creatinine. The cumulative percentages of the group surpassing the urine ALB threshold at different time points (month of age) are listed. Fifty percent of groups 1 and 2 were sacrificed at 5 months of age only after the percentage surpassing the urine ALB threshold in group 2 was more than 75%. The rest of the study rats were sacrificed when they reached 8 months of age. Genotype shown are WT or LRRK2 KO rats. Diets shown are ad lib or opt diet.
Abbreviations: ad lib, ad libitum; ALB, albumin; F, female; M, male; opt, optimized.
Kidney histomorphological findings.
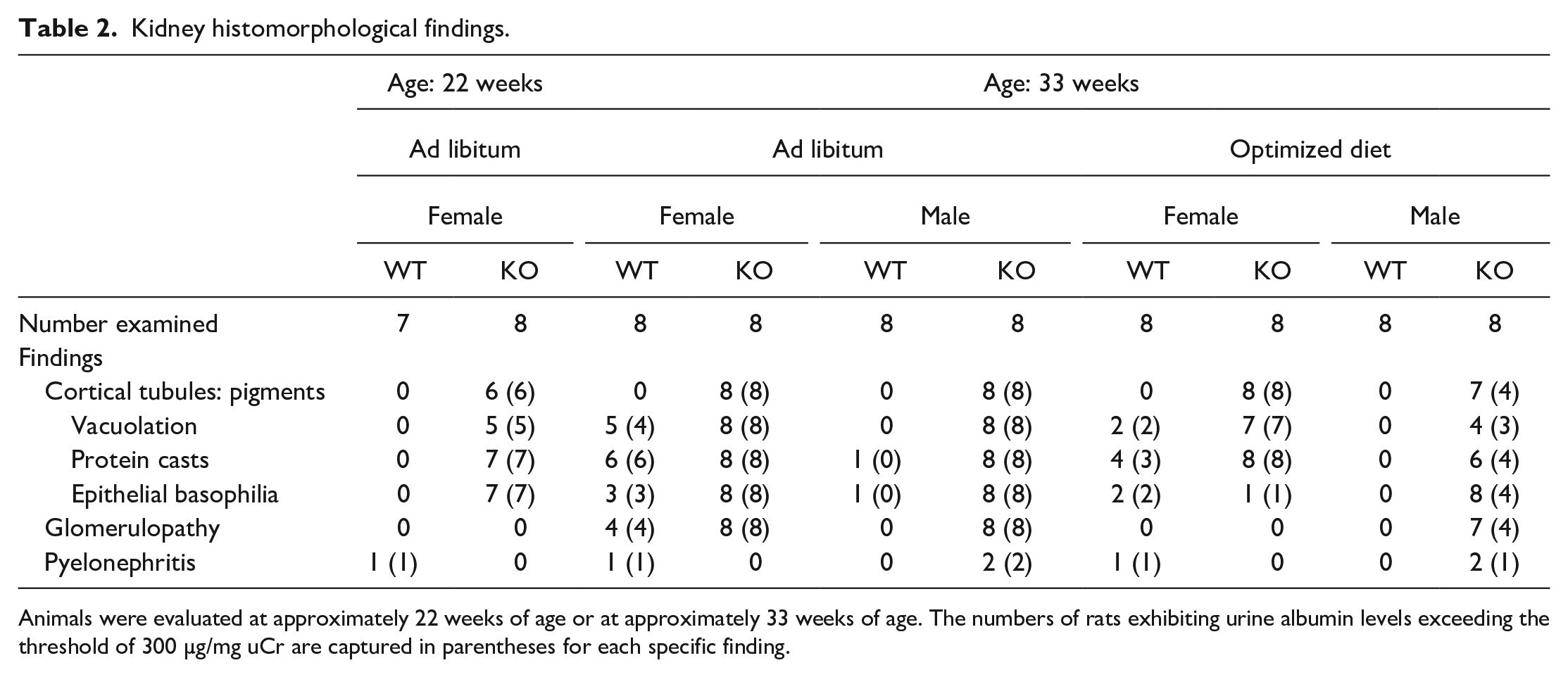
Animals were evaluated at approximately 22 weeks of age or at approximately 33 weeks of age. The numbers of rats exhibiting urine albumin levels exceeding the threshold of 300 µg/mg uCr are captured in parentheses for each specific finding.
Sample Collection and End-Point Measurements
Blood and urine were collected on the same day from fasted animals once per month starting at 2 months of age. BUN and sCr were measured for rats at 2 months of age, while the full routine battery of serum parameters was measured starting at 3 months of age. Urine was collected overnight (16-18 hours) on wet ice for measurements of routine urinalysis parameters and concentration of urinary creatinine. The urine was centrifuged, the supernatant frozen, and stored at −70°C for subsequent analyses of urinary biomarkers.
At termination at week 22 (5 months of age) or at week 33 (8 months of age), the rats were euthanatized and portions of the kidneys were collected and preserved in 10% neutral-buffered formalin. Routine methods were used to prepare paraffin-embedded tissue blocks that were sectioned at 5 µm; sections were subsequently stained with hematoxylin and eosin for light microscopic evaluation. Histomorphologic findings were recorded and graded using a 0-5 scale: 0 (no observable change), 1 (minimal), 2 (mild), 3 (moderate), 4 (marked), and 5 (severe).
Additional specimens of kidneys were collected from rats at scheduled termination for ultrastructural evaluation by transmission electron microscopy (TEM) for all animals. Briefly, the kidney specimens were preserved in 4% paraformaldehyde:1% glutaraldehyde in 0.1 M phosphate buffer, embedded in LX-112 epoxy (Ladd Research, Williston, Vermont), and sectioned at 90 nm. The tissue was stained with 2% uranyl acetate and Reynold’s lead citrate, mounted on a copper mesh grid, and evaluated with a Tecnai Spirit BioTWIN electron microscope (FEI, Hillsboro, Oregon). Ultrastructural images were obtained with a Gatan Orius Digital CCD photography system (Gatan, Inc., Pleasanton, California).
A panel of routine serum chemistry parameters including BUN, sCr, and urinary creatinine were determined using a Beckman AU5800 Chemistry Analyzer according to the manufacturer’s instructions. Urine kidney safety biomarkers ALB, KIM-1, and CLU were measured using a 3-plex custom assay from Meso Scale Discovery (Rockville, Maryland) according to the manufacturer’s instructions. Urine creatinine concentration was used to normalize values of each urinary biomarker. For data analysis and interpretation, the threshold for meaningful changes of urine ALB was set at 300 µg/mg uCr, a threshold used in clinical settings for macroalbuminuria. 23 The threshold for meaningful changes of urine KIM-1 and CLU in urine was set at 3 times the averages of respective baseline values determined from urine collected at 2 months of age of each group. 17
Results
Antemortem Results
Body weight was obtained weekly over the course of the study, with group mean values presented in Figure 1. The mean body weight at approximately 2 months of age (at study start) for LRRK2 KO rats was 27% greater in males and 15% greater in females than for respective WT rats in the ad libitum diet group. These differences were generally evident over the course of the study, with mean body weight of LRRK2 KO rats at 8 months being 25% greater in males and 13% greater in females compared with the respective WT rats in the ad libitum–fed groups. As expected, weight differences between the male LRRK2 KO group and WT groups receiving the optimized diet were less pronounced, with LRRK2 KO rats weighing 6% greater compared with WT at 8 months of age. There were no discernible differences in mean body weights between female KO and WT rats at 8 months of age (Figure 1).
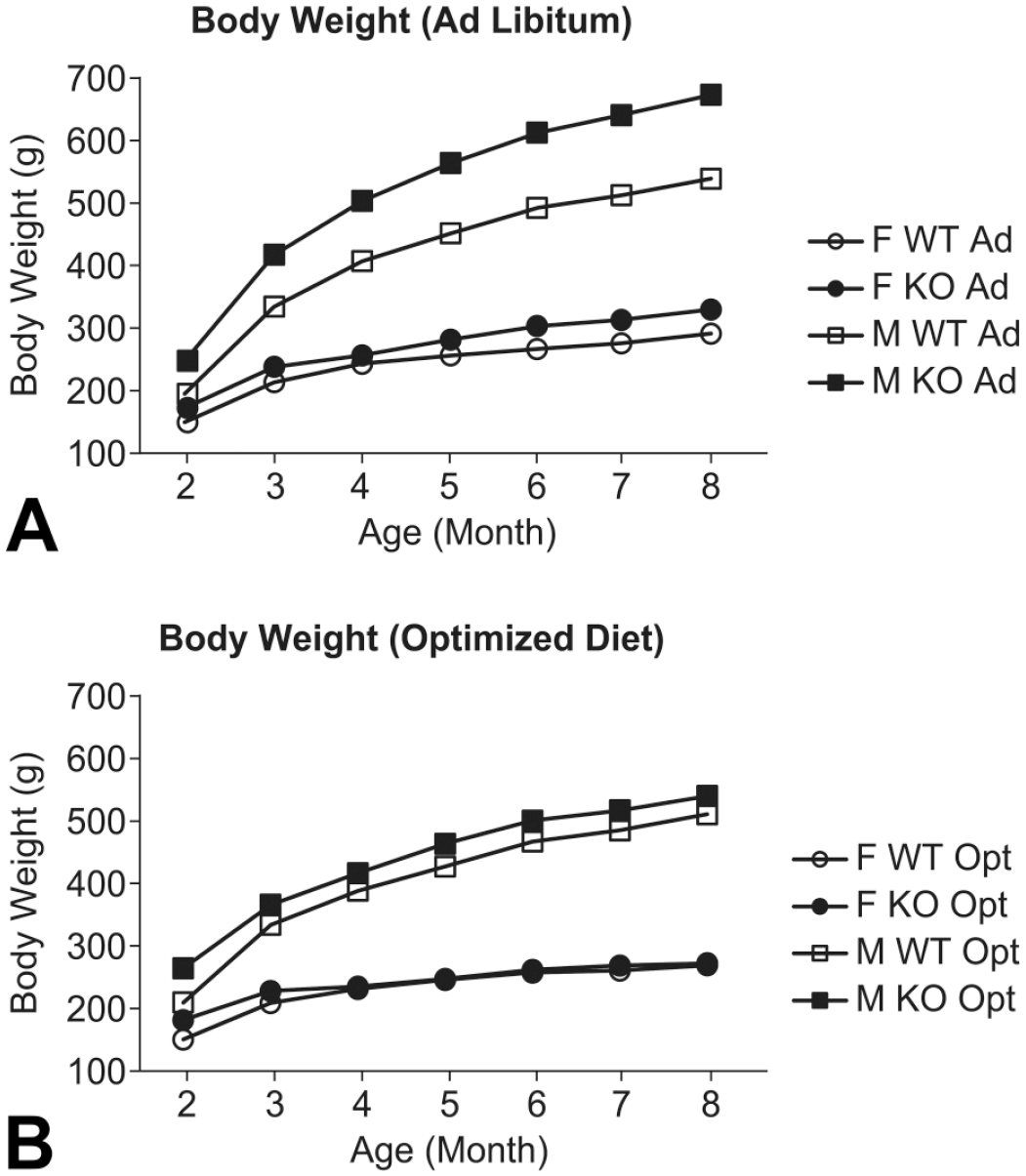
Body weight increase in WT and LRRK2 knockout rats. Average body weight (g) are shown over age (in months) of both WT and LRRK2 KO rats in either ad libitum (Ad) or optimized diet (Opt). Square and circle represent M and F rats, respectively. Solid and no fill represent KO and WT rats, respectively. F indicates female; M, male.
Postmortem Observations
The majority of female LRRK2 KO rats at 5 months of age and receiving an ad libitum feeding regimen exhibited minimal cytoplasmic vacuolation of the proximal tubular epithelium within the renal cortices often associated with intracellular accumulation of punctate to granular-appearing golden-brown pigment. Minimal basophilia of proximal tubular epithelium, variably evident among scattered individual tubules or as small clusters of tubules, exhibited more prominent accumulation of the golden-brown pigment. Small numbers of scattered tubules within the cortices, some of which exhibited basophilia, contained luminal protein casts for a majority of these females. Tubular basophilia was characterized by variable enlargement of individual epithelial cells or enlargement of the entire lining circumference of epithelium, and usually with more prominent nuclei. These features were not observed in kidneys of ad libitum–fed WT females at 5 months of age or confined to only one WT male at termination of the study. No appreciable glomerular changes were evident by light microscopy in ad libitum–fed LRRK2 KO or ad libitum–fed WT rats at 5 months of age.
Ultrastructural differences were observed, however, in glomeruli and tubular epithelial cells in kidneys of female LRRK2 KO rats at 5 months of age, but only in those receiving ad libitum regimen, when compared with age-matched WT (control) rats. Glomerular features in LRRK2 KO females were characterized by deposits of amorphous, punctuate, or granular electron-dense material within foot processes of podocytes and within the cell body of mesangial cells (Figure 2A and B). The primary and secondary foot processes of the podocytes were expanded in LRRK2 KO female rats when compared with foot processes of ad libitum–fed WT rats. Furthermore, the proximal tubular epithelial cells of ad libitum–fed LRRK2 KO rats exhibited an increased number of lysosomes containing variably electron-dense material compared with ad libitum–fed WT rats (Figure 2C and D). No such ultrastructural differences from age-matched WT rats were noted in kidneys of male LRRK2 KO rats at 5 months of age regardless of the feeding regimen.
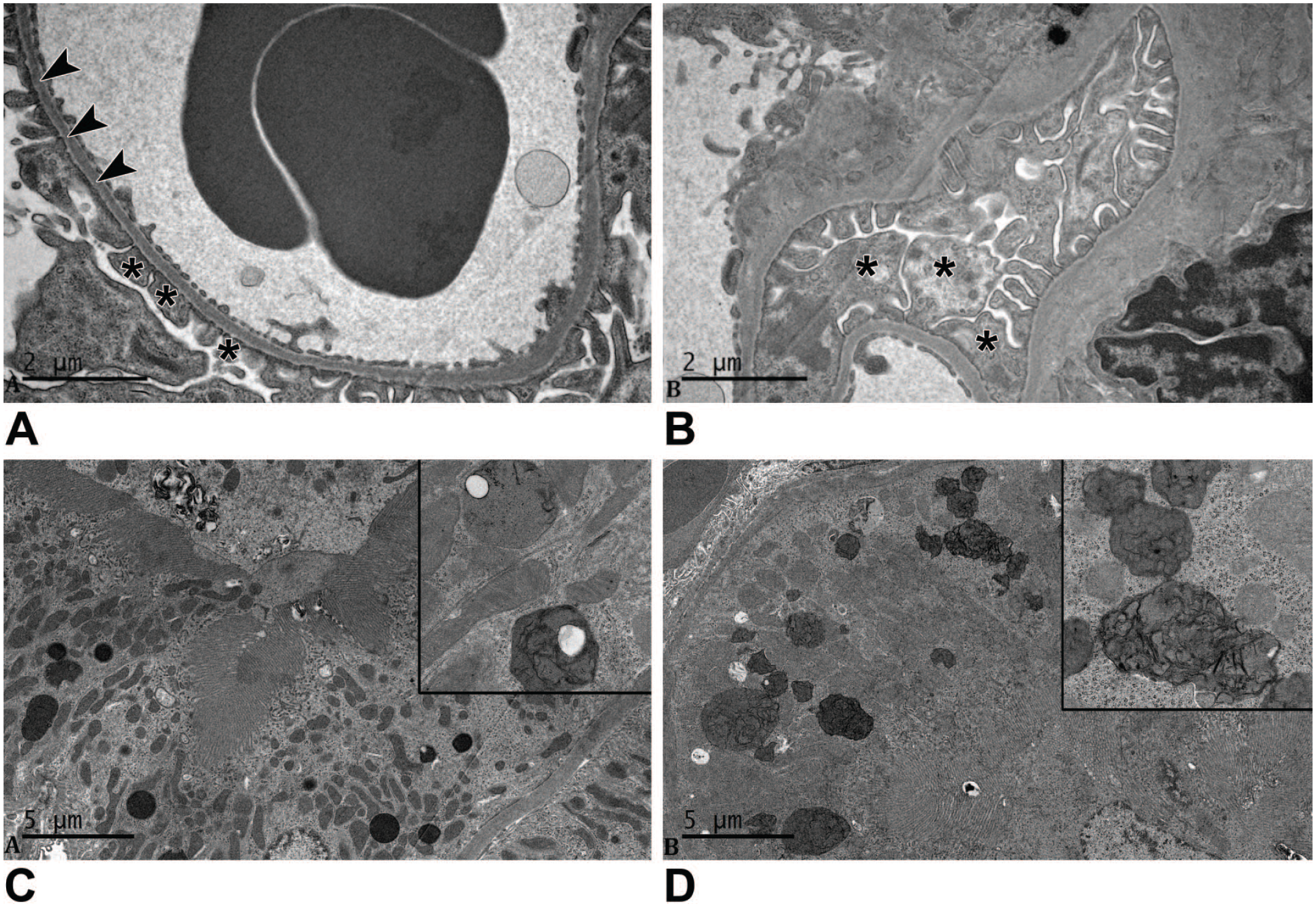
Ultrastructure of WT and LRRK2 KO rats at 5 months. (A) The glomerular basement membrane (arrowheads) and the podocyte foot processes (asterisks) of ad libitum–fed WT rat are not remarkable. (B) The glomerulus of an ad libitum–fed LRRK2 knockout rat differed from the WT rat due to the presence of enlarged podocyte foot processes containing variably electron-dense deposits (asterisks). Scale bars, 2 µm. (C) The renal proximal tubular epithelial cells from an ad libitum–fed WT rat were not remarkable. (D) The proximal renal tubular epithelial cells from an ad libitum–fed LRRK2 knockout rat exhibit increased numbers of lysosomes containing variably electron-dense material. The deposits had a punctate to lamellar form (insert). Scale bars, 5 µm.
Among rats at 8 months of age, glomerular and tubular alterations were more prominent in LRRK2 KO females and KO males that received ad libitum feeding (Table 2 and Figure 3). Alterations of glomerular structure, termed glomerulopathy and generally scored as minimal or occasionally as mild, were readily evident and more advanced, characterized by adhesion of the tuft to parietal epithelium of Bowman’s capsule (synechia formation), thickening of mesangium within the tuft, and variable thickening of Bowman’s capsule, sometimes culminating in glomerulosclerosis. Tubular alterations, minimal or often mild but rarely moderate, were more extensive, characterized by more prominent accumulation of golden-brown material within the tubular epithelium of the outer cortices, more extensive basophilia of the tubular epithelium, and more numerous tubular profiles containing protein casts. Although the nature of tubular features observed in rats at 8 months of age exhibited similar character to those described for female LRRK2 KO rats at 5 months of age, both older female WT and older female LRRK2 KO rats receiving ad libitum regimen had more extensive glomerular and tubular changes within the kidneys, compared with the respective cohorts at 5 months of age.
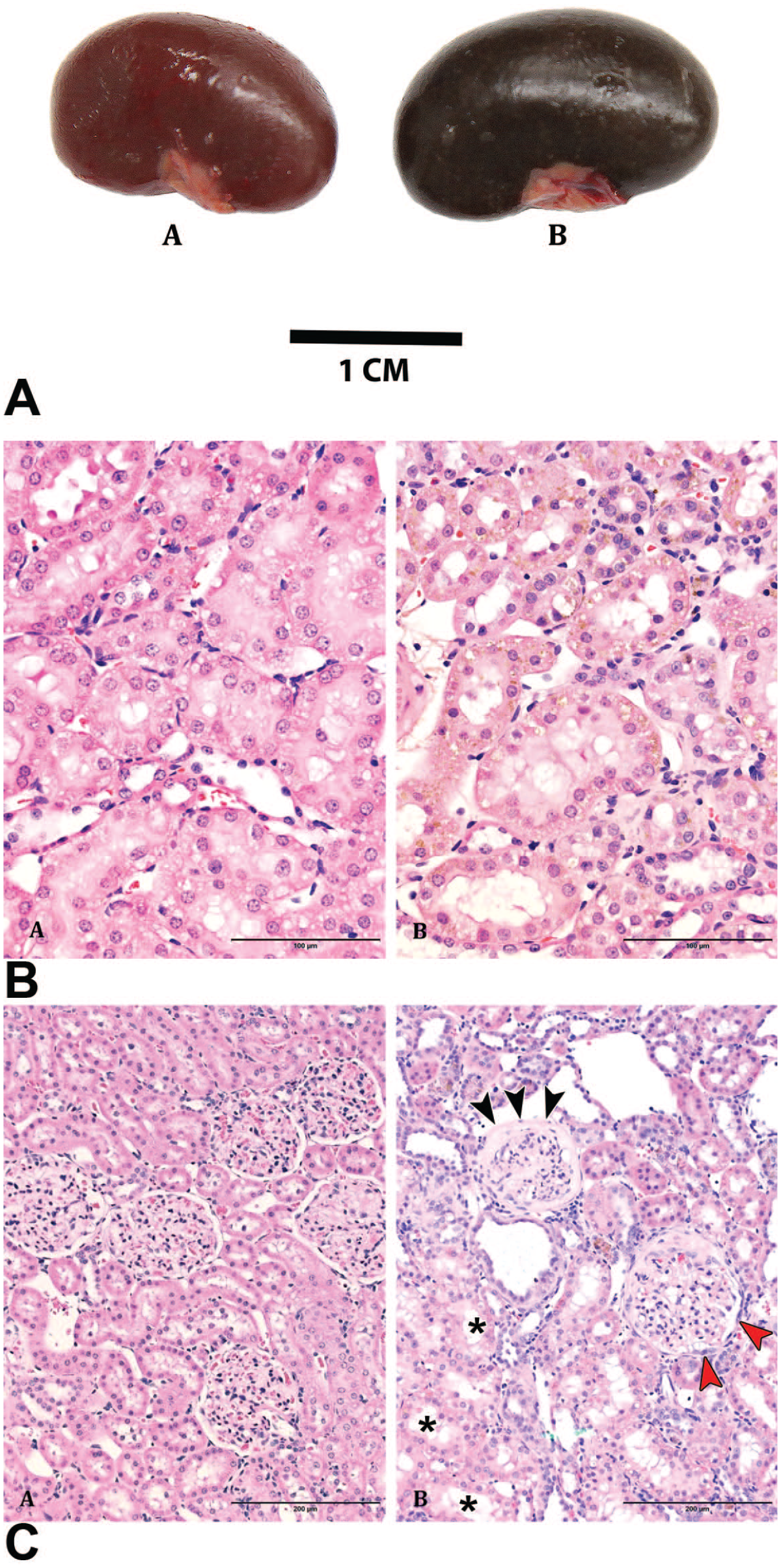
Features of kidneys in WT and LRRK2 KO rats at 8 months. (A). The kidney of LRRK2 KO rats exhibits brown-black discoloration of capsular and cut surfaces (not shown in image) compared with age-matched WT rats. An accumulation of a golden-brown pigment was observed in the cytoplasm of proximal cortical tubules in LRRK2 KO rats B(B) when compared with age-matched WT rats B(A). Scale bar 100 µm. Hematoxylin and eosin staining. Glomerulopathy, observed at greater incidence in LRRK2 KO rats at 8 months C(B), included mesangial thickening of the glomerular tuft, synechia formation, tuft collapse, and thickening of Bowman’s capsule (black arrowhead) with hypertrophy of the parietal epithelium (red arrowhead) compared with age-matched WT rats C(A). Asterisks indicate sites of vacuolation in the tubular epithelium. Scale bars, 200µm. Hematoxylin and eosin staining.
Although KO males at 8 months of age demonstrated minor differences regarding glomerular and tubular features when comparing the ad libitum cohort with the diet optimization cohort, the KO males had lower numbers with overt albuminuria on the optimized diet. In contrast, LRRK2 KO females exhibited lower incidence of glomerular changes and lower incidence of tubular basophilia with diet optimization compared with KO females receiving an ad libitum regimen, while the incidence of albuminuria was similar for KO females regardless of the feeding regimen.
Irrespective of the feeding regimen, similar incidence and minimal to mild pigment accumulation within the tubular epithelium in kidneys of male LRRK2 KO and female LRRK2 KO rats were manifest as brown-black discoloration of the kidneys described by gross examination (Figure 3A). Minimal to mild accumulations of the golden-brown pigment were observed in the renal tubular epithelium of most LRRK2 KO rats, while no pigment accumulation was present in the kidneys of WT rats (Figure 3B and C). Most LRRK2 KO rats exhibited cytoplasmic vacuolation of the proximal tubular epithelium in renal cortices with increased incidence of cytoplasmic vacuolation in ad libitum–fed male and female rats when compared with cohorts fed an optimized diet (Table 2). In addition, more prominent cytoplasmic vacuolation (mild) was noted among ad libitum–fed female LRRK2 KO rats when compared with other LRRK2 KO rats.
Basophilia of proximal tubular epithelium was observed for rats from each cohort with the exception of a WT male on the optimized regimen. Greater incidence and/or severity of tubular basophilia in ad libitum–fed female and male LRRK2 KO rats was evident in comparison with female and male LRRK2 KO rats fed an optimized diet. The male LRRK2 KO rats fed an optimized diet exhibited minimal to mild tubular basophilia; this feature was not observed in male WT rats on a similar diet. Female LRRK2 KO rats fed an optimized diet had a low incidence (1 animal) of tubular basophilia, similar to the female WT rats fed an optimized diet. Randomly in the cortex of some LRRK2 KO and WT female rats were small numbers of dilated cortical tubules containing a luminal eosinophilic material (protein cast). The incidence of protein casts in LRRK2 KO rats fed ad libitum was greater when compared with LRRK2 KO rats fed an optimized diet (Table 2).
Ultrastructural evaluation of proximal tubular epithelium revealed increased numbers of lysosomes containing variably electron-dense material in the proximal renal tubular epithelium of LRRK2 KO rats at 8 months of age when compared with LRRK2 KO rats at 5 months of age or in comparison with concurrent WT rats at 8 months of age (Figure 4A and B). This electron-dense material, having similar ultrastructural appearance to that observed in LRRK2 KO rats 5 months of age, likely corresponds to the pigment described by light microscopy. At 8 months of age, the cytoplasm of the mesangial cell body and foot processes of podocytes in both LRRK2 KO rats and WT rats exhibiting albuminuria contain deposits of an amorphous, punctuate, or granular electron-dense material (Figure 4C and D). The primary and secondary foot processes of podocytes were expanded when compared with the foot processes of non-albuminuric WT rats. The presence of amorphous electron-dense material may represent uptake of ALB from the ultrafiltrate by mesangial cells and podocytes in LRRK2 KO rats.
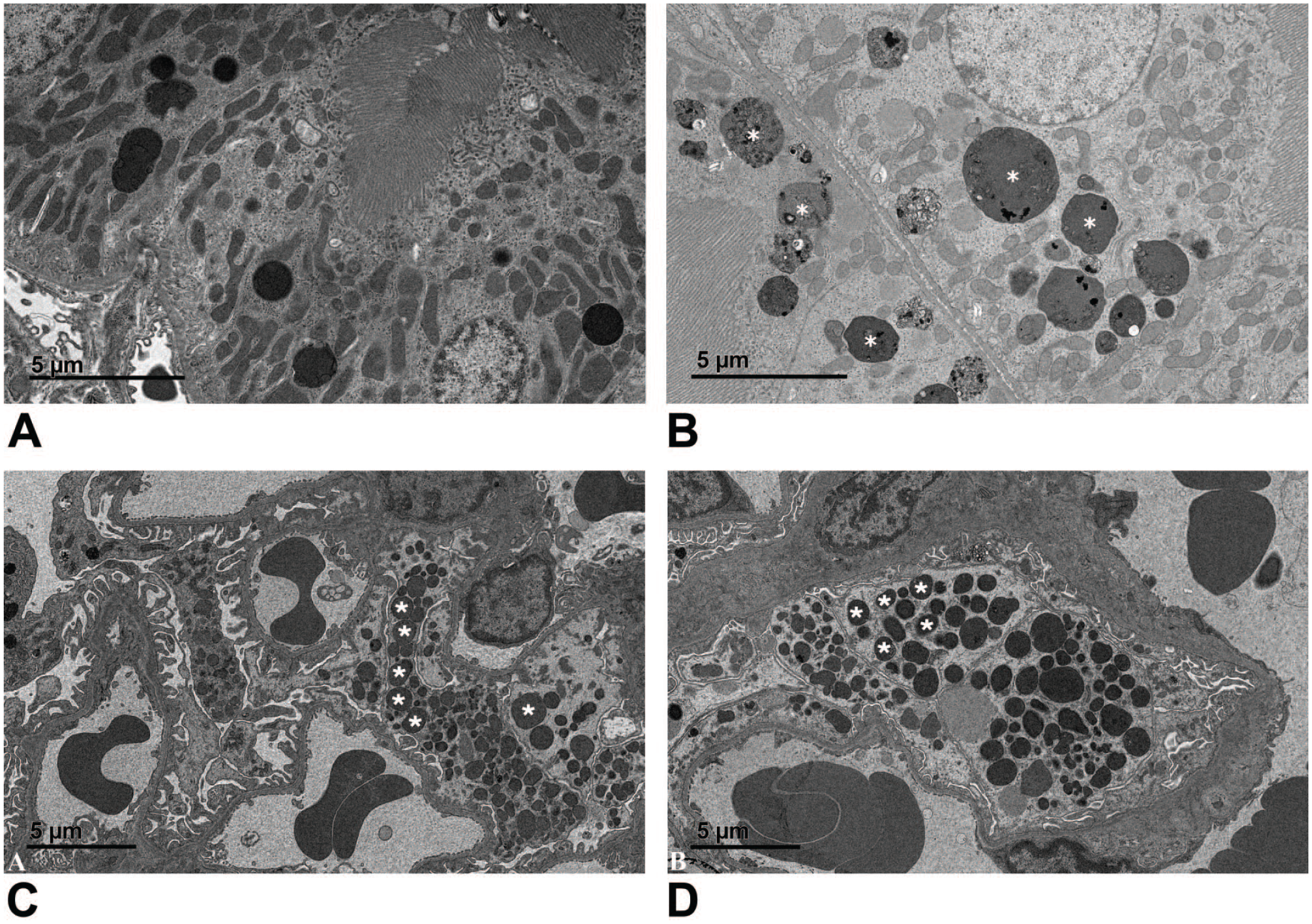
Ultrastructure of 8-month-old WT and LRRK2 KO rats. (A) Proximal renal tubular epithelial cells from an ad libitum–fed WT rat containing lysosomes typical of Long Evans Hooded rats at this age. (B) Proximal renal tubular epithelial cells from an ad libitum–fed LRRK2 knockout rat have increased numbers of lysosomes containing variably electron-dense material (white asterisks). At 8 months of age, independent of the feeding regimen, LRRK2 knockout rats showed increased numbers of lysosomes containing variably electron-dense material when compared with rats at 5 months of age or compared with concurrent, age-matched WT rats. Scale bar, 5 µm. Podocyte foot processes were frequently enlarged and contained variably electron-dense deposits (white asterisks) in WT rats (C) and LRRK2 KO rats (D) with albuminuria. This feature was rarely observed in WT rats without albuminuria (not shown). Glomerular basement membranes were not remarkable. Scale bars, 5 µm.
Serum and Urinary Kidney Biomarkers
Serum chemistry parameters were measured starting from 2 months of age for BUN and sCr only, and a routine battery of serum chemistry parameters including BUN and sCr were measured starting at 3 months of age. There were no significant changes in BUN and sCr over the course of the study regardless of the sex, genotype, or diet (Table 3). Considering the spectrum of morphologic changes described for kidneys at study termination, the results of this study provide another example regarding the limited sensitivity of these conventional serum biomarkers in detecting early or low-grade renal changes in rodents.
Serum chemistry parameters over the study period.
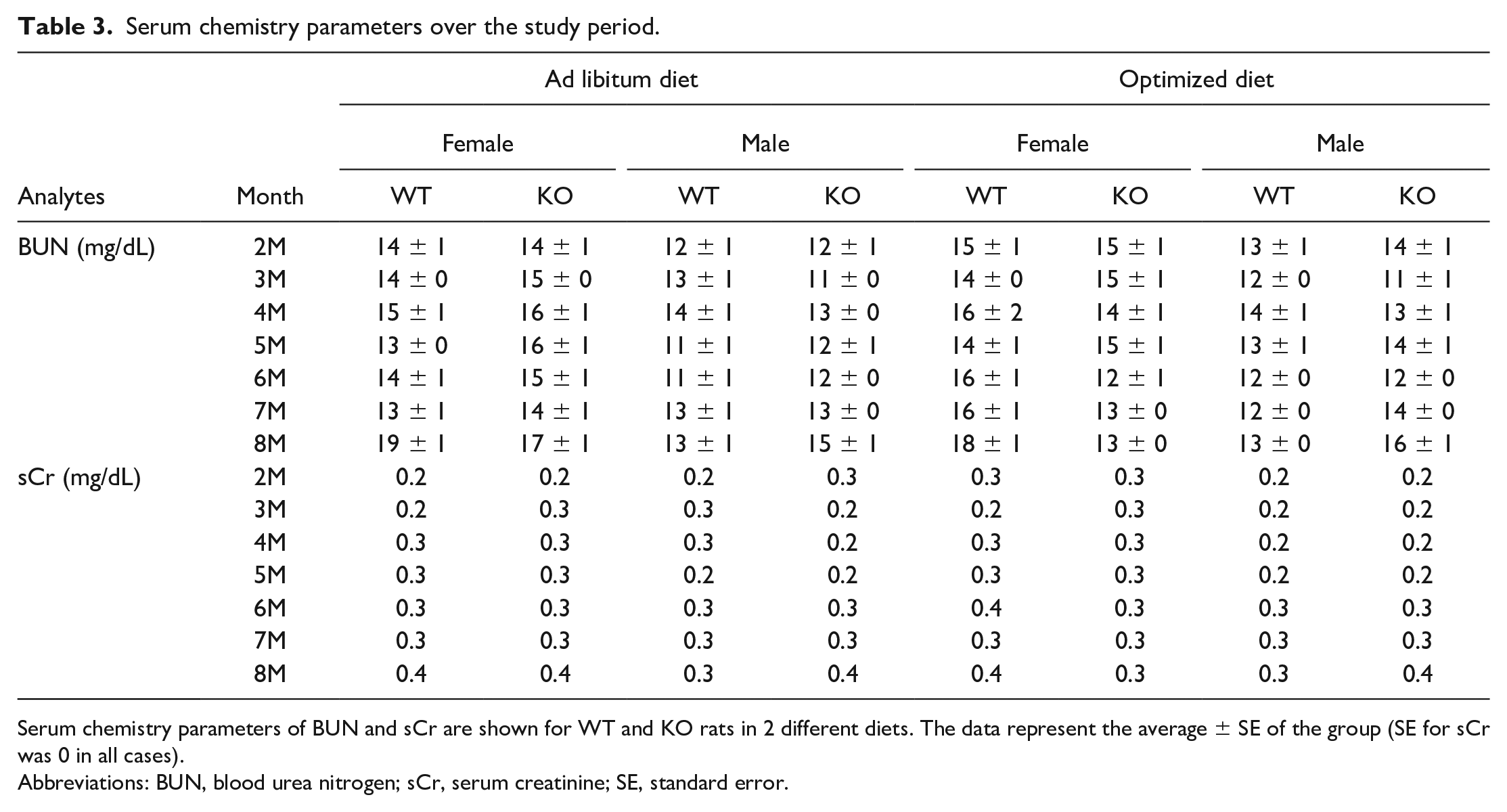
Serum chemistry parameters of BUN and sCr are shown for WT and KO rats in 2 different diets. The data represent the average ± SE of the group (SE for sCr was 0 in all cases).
Abbreviations: BUN, blood urea nitrogen; sCr, serum creatinine; SE, standard error.
Urinary levels of kidney safety biomarkers ALB, KIM-1, and CLU were monitored, and the fold changes over threshold values of urinary ALB (at 300 µg/mg uCr) and KIM-1/CLU (3 times the average of the biomarker baseline levels at 2 months of age) over the study time course are shown in Figure 5. Urinary ALB levels of female and male LRRK2 KO rats on ad libitum diet started to cross the 300 µg/mg uCr threshold at 3 and 4 months of age, respectively. The female and male WT rats with the same diet did not cross the threshold until 6 and 8 months, respectively (Figure 5A and Table 1). One hundred percent of female and male LRRK2 KO rats on ad libitum diet crossed the ALB threshold by 6 months, while at the same time point only 25 and 0% of female and male WT rats with the same diet did. These data demonstrated the early-onset albuminuria in LRRK2 KO rats on the ad libitum diet compared with WT rats.
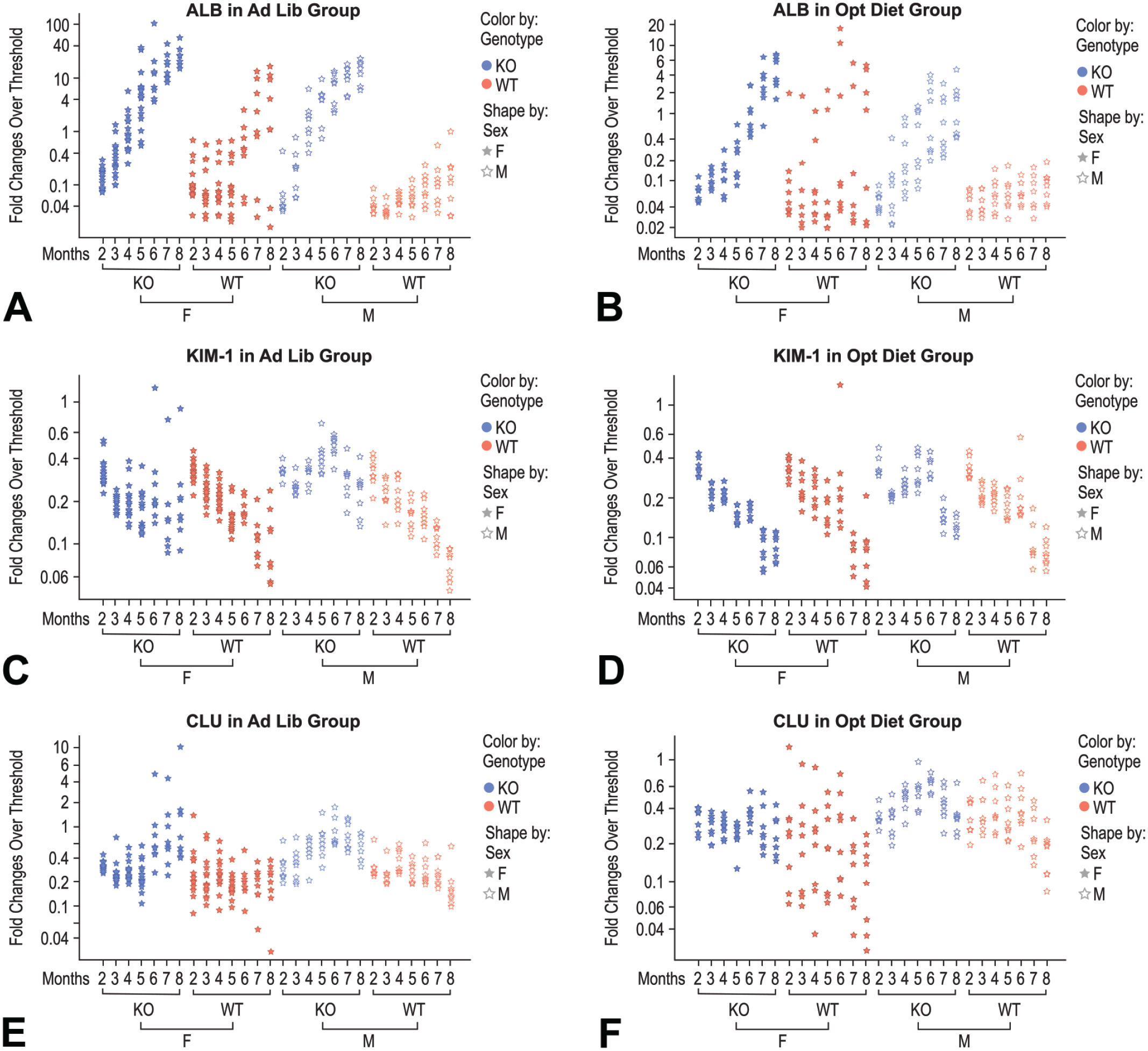
Fold change over threshold values of urinary ALB, KIM-1, and CLU over the course of the study. Fold change of urine ALB, KIM-1, and CLU over threshold was plotted in log scale against age (month), genotype, and sex of all rats of the study. Female and male rats are represented with solid and open stars, respectively. LRRK2 KO and WT rats are represented with blue and orange colors, respectively. Thresholds were set at an absolute value of 300 µg/mg uCr for ALB and at 3-fold over the group mean of baseline urine creatinine normalized values for KIM-1 and CLU measured at 2 months of age. ALB indicates albumin; CLU, clusterin; KIM-1, kidney injury molecule 1.
Adjusting food intake delayed onset of albuminuria in WT females and LRRK2 KO female and male rats. It was not until 7 months of age that the majority (87.5%) of female LRRK2 KO rats in the optimized diet groups crossed the ALB threshold, compared with 5 months of age for the majority of female LRRK2 KO rats on ad libitum diet to cross the ALB threshold (group 2 vs group 6, Table 1). The male LRRK2 KO rats in the optimized diet group took 6 months to reach the 50% rate of albuminuria, while the same LRRK KO male rats with ad libitum diet had a 62.5% albuminuria rate at 4 months of age (group 4 vs group 8, Table 1). Furthermore, WT female rats developed albuminuria more rapidly than WT males in both diet regimens (groups 1 vs group 3, and groups 5 vs group 7, Table 1), while the differences in the onset of albuminuria between male and female rats were minimal among LRRK2 KO animals (group 2 vs group 4, and group 6 vs group 8, Table 1). In addition, the application of the optimized diet successfully attenuated spontaneous renal changes in WT female rats (Table 2).
The magnitude of urine ALB fold change over the 300 µg/mg Cr threshold over time was substantial, making ALB stand out as a strong performing kidney injury biomarker. By 8 months of age, the range of ALB changes over threshold was between 15- and 56-fold in female LRRK2 KO rats on ad libitum diet compared with up to 17-fold in female WT rats on the same diet. The range of ALB changes was between 6- and 23-fold over the 300 µg/mg threshold in male LRRK2 KO rats on ad libitum diet compared with only 1 male WT rat with an ALB level at the threshold on the same diet (Figure 5A). The limitation of food intake attenuated the rapid progression to albuminuria and decreased the magnitude of urine ALB fold changes as well. The range of urine ALB fold changes over threshold was between 1.5 and 6.7 in female LRRK2 KO rats on the optimized diet, with similar ALB fold change reductions observed in other groups on the optimized diet as well (Figure 5B).
Urinary KIM-1 levels showed a different profile compared with that of ALB. No KIM-1 increase was observed in any rat of any group at the time of termination. There was only a transient increase in KIM-1, barely surpassing this urine biomarker’s threshold in 1 female LRRK2 KO rat on the ad libitum diet and 1 female WT rat on the optimized diet (Figure 5C and D). There was a trend of KIM-1 levels decreasing over the study time course in both the ad libitum and optimized diet groups.
Urinary CLU also did not show a general increase. At 8 months of age, only 3 female LRRK2 KO rats on the ad libitum diet showed CLU levels above threshold with 2 of the 3 just surpassing the threshold. There were also 3 male LRRK2 KO rats on the ad libitum diet showing transient increases in CLU just crossing the threshold, but their levels fell below threshold at the time of termination (Figure 5E and F). Urinary KIM-1 or CLU levels were concluded to not have a positive correlation with the histopathologic findings observed for the kidneys in this study.
Discussion
We document the time course of early-onset albuminuria without concurrent increases in the urinary tubular biomarkers KIM-1 and CLU in aging LRRK2 KO rats. The presence of albuminuria is consistent with previously described proteinuria in LRRK2 KO mouse. 12 The onset of albuminuria observed in the LRRK2 KO rats starts approximately at 3 and 4 months of age for female and male rats, respectively, consistent with the prior report that no albuminuria was observed in male LRRK2 KO rats of the same strain at 3 months of age. 13 sCr and BUN also did not show significant changes throughout the duration of the study, indicating that these conventional serum kidney safety parameters were not useful indicators for monitoring subtle effects on kidneys caused by the lack of LRRK2 expression or associated with pharmacologic inhibition of LRRK2, which is consistent with the existing literature.13,14
We describe greater body weight increases in male LRRK2 KO rats than in male WT rats up to 8 months of age. These data are consistent with prior published data in the same rat strain reaching 4 months of age. 13 We also report greater body weight increases in female LRRK2 KO LEH rats than in female WT rats, but the percentage of difference between the KO and WT female rats was much less than the corresponding male rats. While the molecular mechanisms behind the body weight gain in LRRK2 KO rats are unknown, there were increases of weekly average food consumption of 26% and 8% for male and female LRRK2 KO rats, respectively, compared with their WT controls. This is consistent with more body weight gain observed in male versus female LRRK2 KO rats. Adjusting food intake greatly reduced the rate of body weight gain in KO animals, as expected. However, lower body weight gain in female LRRK2 KO rats did not impact albuminuria in this group of animals, suggesting that body weight gain per se is not the dominant factor for developing albuminuria in LRRK2 KO rats.
It is interesting to note that WT females exhibit a range of spontaneous renal changes and eventually develop albuminuria without an impact on urinary biomarkers. Furthermore, it is relevant to clarify that the spectrum of morphologic findings identified in kidneys of WT LEH and LRRK2 knock-out animals does not mimic or resemble those described as chronic progressive nephropathy (CPN) in SD rats and do not resemble spontaneous changes noted in kidneys of Han-Wistar rats.24,25 Furthermore, the kidney findings in LRRK2 KO rats do not resemble the characteristics of minimal change disease.
ALB is a 69-kDa negatively charged globular protein produced in the liver and secreted into the plasma. Although ALB is the most abundant plasma protein, it is relatively impermeable through the glomerular capillary wall due to its molecular weight and its negative charge.26,27 Limited but significant amounts of ALB are filtered and then almost entirely reabsorbed by proximal tubular cells through binding to a megalin/cubulin receptor complex, followed by receptor-mediated endocytosis.26,28 Large quantities of ALB would be present in urine either when there is a glomerular injury that allows large amounts of ALB in the glomerular filtrate through an altered glomerular basement membrane or when the amount of filtered ALB overwhelms the reabsorption capacity of compromised tubules. 29 Therefore, the increase in urine ALB concentration can be due to either glomerular or tubular injury or both, although primary functional effects on the kidney have not been excluded. The temporal sequence for an initial alteration of the structural change in the kidney during this study is uncertain because tubular alterations were present at the earliest time point of histomorphologic examination (interim termination at 5 months of age).
On the contrary, urinary KIM-1 and CLU are regarded as tubular injury-response biomarkers. KIM-1 expression is confined to proximal tubular epithelium, while CLU expression is more widely distributed within the renal tubules.30 -32 The presence of both glomerular and tubular changes with concurrent urine ALB increase is expected. However, the observed tubular alterations noted in the LRRK2 KO rats without an increase in urinary KIM-1 and CLU levels in this study are intriguing, and the underlying mechanisms are unclear. The observed decrease in basal KIM-1 urine protein levels over time up to 8 months is different from the previously reported relatively stable KIM-1 baseline kidney tissue expression at both messenger RNA and protein levels over the same age span in Fisher 344 rats. 33 Although albuminuria in WT LEH and LRRK2 KO rats is identified in this study, it is unclear what role LRRK2 plays in proximal tubular epithelium and in the uptake of ALB. LRRK2 is demonstrated to interact with vesicular proteins, including the clathrin adaptor protein complex 2 (AP2), while adaptor protein complex 3 (AP-3) acts downstream of LRRK2, as described in kidneys of mice.34,35 Altered lysosomal or cytoskeletal proteins associated with defective vesicular function in renal epithelium are described subsequent to knockdown on LRRK2.11,34,36,37 These observations are consistent with the notion that LRRK2 protein may play an important role in ALB uptake by proximal tubular epithelium.
The presence of albuminuria is associated with morphological alterations described for both glomeruli and proximal tubular epithelium, as demonstrated by an overall concordance of albuminuria with tubular casts, glomerulopathy, and tubular basophilia by 8 months of age in LRRK2 KO rats. This is not surprising because ALB is a highly abundant urine protein filtered through glomerular basement membrane (GBM). Furthermore, urinary ALB started to rise at 5 months of age with increases over threshold reaching as high as 74-fold (Figure 5) without evidence of glomerulopathy by light microscopic evaluation, suggesting that ALB is an early indicator of renal function. This conclusion is supported by patterns of persistent urinary ALB levels surpassing the threshold over longer periods (more than 2 months) prior to detection of glomerulopathy associated with tubular changes based on light microscopic examination at 8 months of age. This conclusion is further supported by the ultrastructural examination of 5-month-old rats that demonstrated accumulation of amorphous electron-dense material within mesangial cells and podocytes of the glomeruli, judged to reflect functional uptake of protein.28,29 Over time, the light microscopic findings of tubular epithelial vacuolation, the presence of tubular pigment, proteinaceous casts in cortical tubules, and tubular epithelial basophilia correlated with albuminuria in LRRK2 KO rats at the 5-month time point (Figure 2), indicating that urine ALB serves as a biomarker for subsequent glomerular and tubular effects. The association between urine ALB increase and eventual morphologic changes in the kidneys establishes urine ALB as a useful biomarker for monitoring renal function in LRRK2 KO rats. However, the temporal sequence between the onset of albuminuria and functional changes or morphological changes in tubular and glomerular structure is not fully elucidated, given the limited time points of morphological examinations performed during this study.
It was recently shown that human heterozygous loss of function (LOF) variants in LRRK2 coding gene with concomitant reduced LRRK2 protein levels were not associated with any specific phenotype or disease state (including kidney disease), and that there was similar urine ALB levels in the LOF variants compared with controls. 38 Given the gain of function for LRRK2 in familial PD and sporadic forms of PD, administration of an efficacious LRRK2 inhibitor would likely have limited risk of renal insult. Our study data indicate that a measurement of urine albumin to creatinine ratio (ACR) could potentially support kidney safety monitoring in preclinical and clinical development of LRRK2 inhibitors, either alone or in combination with other kidney toxicity parameters.
In summary, we report that early-onset albuminuria is a characteristic of LRRK2 KO rats, with urinary ALB serving as a sensitive biomarker for monitoring potential effects on renal function and related morphologic changes in kidneys of LRRK2 KO rats. These results provide further insight regarding LRRK2 function in renal physiology of rodents, particularly regarding proximal tubular function in the kidney. Urinary albumin could potentially be used as a sensitive kidney safety biomarker in the development of LRRK2 inhibitors.
Footnotes
Acknowledgements
The authors wish to acknowledge the consultations and perspectives provided by Dr George Parker regarding light microscopic and ultrastructural features of WT and LRRK2 KO animals.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.