
Abstract
Long-tailed macaques are the predominant nonhuman primate species for the nonclinical safety testing of biopharmaceuticals. This species comprises 9 subspecies with Macaca fascicularis fascicularis naturally occurring in Southeast Asia. Since the 17th century, M. f. fascicularis also occurs on Mauritius. Cynomolgus macaques do not naturally occur in China, but are bred in many farms across the country. The current shortage in animal supply raises the question whether geographical animal origin matters and if animals from different geographical regions can be combined on a drug development program or even a single experiment. This article reviews geographical animal origin in relation to selected endpoints that are relevant in nonclinical drug safety testing. Animals from different countries within Asia mainland do not appear to show any meaningful difference. Very little data are available for animals from Asia island. Mauritian animals show consistent differences from Asian animals in several clinical and anatomical pathology parameters. For developmental parameters, animals from Mauritius and Asia are comparable with the exception that Mauritian animals mature faster. In the authors’ view, differences between the geographical clusters can be accounted for as long as baseline and reference data are available.
Introduction
Nonhuman primates (NHPs) are used increasingly for the safety testing of new targeted human medicines. The reason being that in those cases, NHPs frequently represent the only relevant animal species for safety testing, either because of the pharmacological effect or because of physiological and anatomical similarities to humans. This holds particularly true for the development of monoclonal antibodies for which it is reported that over 70% of compounds have NHPs as the relevant animal model.1-4 For the development of new gene therapies and therapeutic oligonucleotides, NHPs are also typically used. High demand for NHPs also comes from new vaccine development including vaccines against COVID-19. As a result, the demand for NHPs to be used in scientific procedures has increased substantially over the past years. While a report published in 2017 concluded that the global availability in F2/F2+ generation NHP exceeds the current and projected European Union (EU) demand, 5 this statement seems no longer valid. As of 2021, NHP availability seems lower than the current demand for animals, and the situation may even worsen in coming years. This NHP supply shortage has already gained public attention and it is of interest to note that a 2018 National Institutes of Health (NIH) primate report mentioned a “strategic monkey reserve” for the US National Primate Centers to provide “surge capability for unpredictable disease outbreaks.” The report recommended to expand the NIH-sponsored Indian rhesus monkey and marmoset monkey colonies, and to establish domestic colonies for long-tailed macaques. 6
Among all NHP species used for biomedical research (e.g., prosimians, marmosets and tamarins, rhesus monkeys, baboons, squirrel monkeys), cynomolgus monkeys are the most predominant (>90%) NHP species for the safety assessment of pharmaceuticals. The number of cynomolgus monkeys used in the EU under the category “Regulatory use and routine production” increased from 5371 in 2015 to 5433 in 2016, 6219 in 2017, and finally to 6870 animals in 2018. 7
The cynomolgus monkey is a macaque species that derives from South-East Asia and is known under additional names, i.e., Macaca cynomolgus, Macaca irus, crab-eating macaque, and Java macaque. The current scientific common name for Macaca fascicularis is long-tailed macaque, 8 since the tail in this species is particularly long relative to body length. Long-tailed macaques originate from equatorial latitudes without a distinct summer and winter. Therefore, the animals are fertile and sexually active throughout the entire year. Another advantage of the long-tailed macaque is that male and female animals are socially highly compatible even after having reached sexual maturity.
Geographical Origin of Long-Tailed Macaques
Long-tailed macaques have the third-largest habitat distribution among all NHPs. Natural habitats comprise entire South-East Asia (Philippines, Malaysia, Indonesia, Myanmar [Burma], India, Vietnam, Cambodia, Laos, and Thailand). In addition, long-tailed macaques were introduced to the island of Mauritius in the 17th century. Long-tailed macaques do not have natural habitats in China, but they are commonly bred in China as laboratory animals.
There are several ways how long-tailed macaques from different geographical regions have been categorized:
Based on fur coloration, 9 subspecies are being recognized 8 : M. f. fascicularis, M. f. aurea, M. f. umbrosa, M. f. atriceps, M. f. condorensis, M. f. fusca, M. f. lasiae, M. f. tua, and M. f. karimondjiwae. The former subspecies M. f. philippinensis is now also classified as M. fascicularis fascicularis.9,10 M. fascicularis fascicularis mainly occurs in Vietnam, Cambodia, Thailand, and Malaysia. The geographical distribution of subspecies is depicted in Figure 1. Many subspecies are located on small islands except for M. f. fascicularis and M. f. aurea, the latter being located in Myanmar and Thailand.
From a genetic viewpoint, four geographical habitat clusters have been described for the long-tailed macaque: Indochinese, Indonesian-Malaysian, Philippine, and Mauritian.11-15 The distribution of these populations is depicted in Figure 1. The Indochinese cluster is also referred to as Asia mainland. The Indonesian-Malaysian cluster together with the Philippine cluster is also referred to as Asia island. Animals with island origin are more diverged than those with mainland origin due to genetic isolation over time. 16 The Mauritian and Philippine clusters appear most divergent from other populations. For the Philippine origin, this may be related to prolonged deep water separation.
Mitochondrial DNA analysis and analysis of Y-chromosome haplotypes revealed several genetically distinct lines: An Indochinese line and several Sundaic lines (the Sundaic region is a biogeographical region of Southeastern Asia corresponding to a larger landmass that was exposed throughout the last 2.6 million years during periods when sea levels were lower; it includes Java, Borneo, and Sumatra and their surrounding small islands, as well as the Malay Peninsula on the Asian mainland).17,18 A recent Y-chromosome haplotype analysis with focus on Malaysian populations confirmed the Indochinese line and several Sundaic lines (Figure 1). 18 It appears that Sumatra is the only region known to carry continental Y-chromosome DNA and insular mitochondrial DNA 17 and to carry two Sundaic haplotype variants. 18 Recently, a northern Thailand population of long-tailed macaques exhibiting a unique genetic structure was reported. 19 Indochinese long-tailed macaques are a genetically and immunologically distinct entity from Indonesian long-tailed macaques. 20 Malaysian long-tailed macaques appear rather diverse with variants between Malay Peninsula vs Borneo insular, east coast vs west coast of Malay Peninsular and mainland vs island Malay Peninsular. 21 The presence of continental and island Y haplotypes has been reported from the Indochinese island of Bali. 22
Mitochondrial whole genome sequencing revealed two main clades—Clade A includes mainland Southeast Asia, the Malay Peninsula, and North Sumatra and Clade B the islands of Bangka, Java, Borneo, Timor, and the Philippines. 23 A subsequent mitogenomic analysis confirmed a mainland/northern Sumatra clade and an additional clade that comprises all insular haplotypes along with lineages from southern Sumatra. 24
The island of Mauritius did not have a long-tailed macaque population until the 17th century, when presumably about 20 animals were transferred to Mauritius (anthropogenic translocation). This low number of transferred animals could impose a strong founder effect since it is assumed that no further animals had been transferred thereafter. Some researchers consider Mauritian long-tailed macaques as inbred and rather homogeneous. The precise geographical region from where the translocation initiated is still under debate. Since Mauritian long-tailed macaques carry a combination of island mitochondrial DNA and continental Y-chromosomal DNA, it is assumed that Mauritian long-tailed macaques originated from Sumatra. 17
Until 2020, China was a major supplier of purpose-bred long-tailed macaques, albeit this species is not endemic to China. In fiscal year 2014, China provided over 70% of the total cynomolgus macaques imported into the United States. 25 Analysis of DNA of 400 animals from 10 Chinese breeding farms revealed that most of these animals had an Indochinese origin. 26 Previous single-nucleotide polymorphism also indicated an Indochinese animal origin from Chinese breeding farms. 26 This observation is further supported by cytochrome P450 variant analysis, since the CYP2C9 and CYP2C19 variants—predominant in Cambodian and Vietnamese origin long-tailed macaques 27 —are also prevalent in long-tailed macaques bred in China. 28
Long-tailed macaques carry some relatedness to rhesus monkeys, who have the second-largest habitat distribution among NHPs. Rhesus monkey habitat overlaps with that of long-tailed macaques in Cambodia and southern regions of Thailand, Laos, and Vietnam (Figure 1). Both species mate successfully and interestingly the hybrid offspring show fertility when mature. Genetic analysis from feral long-tailed macaque samples revealed 5% to 48% rhesus ancestry. 26 Opposed to long-tailed macaques, rhesus monkeys are short-tailed and it is of interest to note that the relative tail length of rhesus/long-tailed hybrid animals correlates with the geographical origin of specimens such that relative tail length increases to some extent with more southern location. 29 Average relative tail length in rhesus monkeys from China, India, Myanmar, and Indochina is 35%, 42.5%, 44%, and 60%, respectively, as compared with 117% in Indochinese long-tailed macaques. 30 In long-tailed macaques from regions south of the isthmus of Kra, relative tail length exceeded 125%. 31 Overall, however, the species-specific characters of a shorter tail (<70%) and a bipartite pelage color pattern are still present in Indochinese rhesus macaques while the longer tail (>90%) and no bipartite pattern are still present in Indochinese long-tailed macaques. 32 The Indochinese rhesus monkeys have been considered a separate group in comparison with the Indian, Chinese, and Myanmar rhesus monkeys. Thailand appears to be a hotspot for rhesus and long-tailed macaque hybridization. 30 Overall, about 30% of the long-tailed macaque genome is believed to be of rhesus monkey origin. 33 The genetic migration rate from rhesus to long-tailed macaques appears slightly greater than in the opposite direction, 19 and the Indochinese animal population is more affected by rhesus introgression compared with more southern geographical regions. The reason likely being that in glacial times land barriers evolved, e.g., Isthmus of Kra, Surat Thani-Krabi depression, and Kangar-Pattani line 16 (Figure 1), that prevented further south-migration of rhesus monkeys. The Sundaic long-tailed macaques apparently lack rhesus Y-chromosomal DNA. 18 Gene flow occurred via male animals from rhesus to long-tailed macaque. Beyond that, however, a long-tailed to rhesus macaque gene flow boundary has been reported between northern Thailand and Myanmar. 35 It is also important to note that rhesus introgression continues to occur in regions of overlapping habitats.
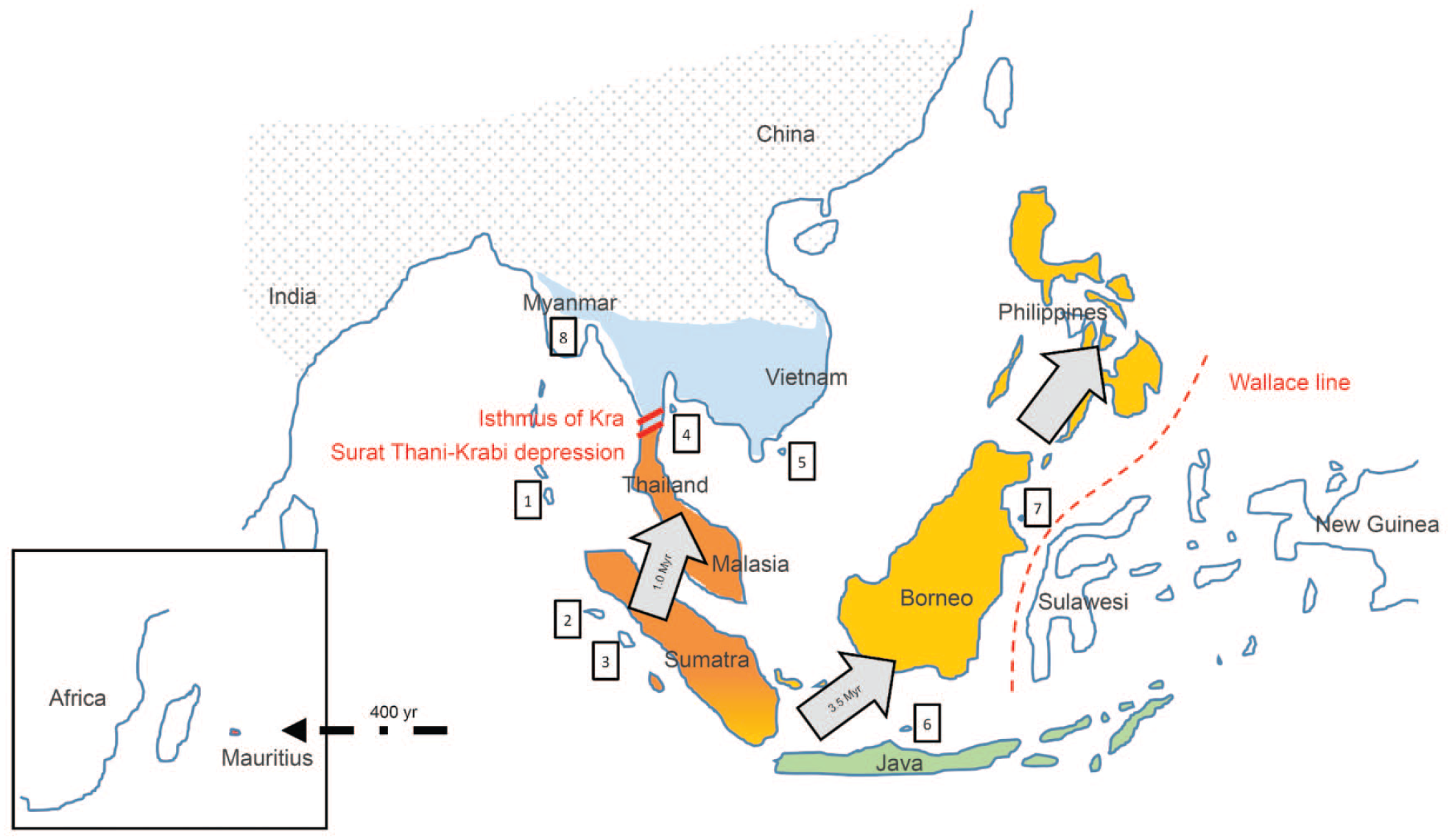
Map showing the distribution of subspecies of long-tailed macaques (Macaca fascicularis). Distribution of the subspecies M. f. fascicularis is shown in color, representing different Y-chromosome haplotypes (data taken from Rovie-Ryan et al. 18 ). Blue = Indochinese line (with influence from Rhesus macaques); yellow, orange, and green = 3 different Sundaic lines (without influence from Rhesus macaques). Distribution of Rhesus macaques is indicated by dots. Land barriers (Surat Thani-Krabi depression; isthmus of Kra) and the faunal boundary line separating Asian from Australian fauna (Wallace line) are indicated in red. Animals north of the Isthmus of Kra represent the Indochinese cluster, animals south of the isthmus of Kra together with animals on the islands of Sumatra, Java and Borneo represent the Indonesian-Malaysian cluster, animals on the Philippines represent the Philippine cluster, and animals on the island of Mauritius represent the Mauritian cluster (Haus et al. 33 ). Long-tailed macaques originate from the island of Sumatra and migrated to the island of Borneo approximately 3.5 million years ago from where they reached the Philippine islands (likely by rafting). They migrated to the South-East Asia peninsula approximately 1 million years ago and subsequently dispersed north. Long-tailed macaques were introduced onto the island of Mauritius from the Indonesian islands by sailors in the early 1600s (indicated by arrows). 1 indicates M. f. umbrosa; 2, M. f. lasiae; 3, M. f. fusca; 4, M. f. atriceps; 5, M. f. condorensis; 6, M. f. karimondjiwae; 7, M. f. tua; 8, M. f. aureus (data taken from Roos and Zinner 34 ).
Geographical Origin of Long-Tailed Macaques Used in General Toxicology Studies
Long-tailed macaques are extensively used in general toxicity studies, particularly for biotechnology-derived pharmaceuticals. Most animals in these experiments originate from the Indochinese cluster (i.e., Asia mainland) or from Mauritius: In the year 2014, 60% of all long-tailed macaques used in the EU were supplied from Mauritius and 35% were supplied from Asia. 5 These numbers have not dramatically changed since 2014 (personal observation). There is no guidance what animal origin is preferred for the safety assessment of products, but it is recognized that variation between animals from different geographic origins can contribute to diverse drug pharmacologic and toxicologic responses. 36
The question if geographical origin of NHPs matters is typically answered by comparing baseline data between animal cohorts of different geographical origin. Unfortunately, investigations that explicitly evaluate such difference in the context of a regulatory general toxicity study are extremely scarce.
A notable example is the study by Kozlosky et al. 37 The study enrolled three different cohorts of long-tailed macaques: Animals of the Mauritian cluster, mixed animals of the Asia mainland cluster (originating from Cambodia, Vietnam, and Indonesia) and animals originating from Cambodia only. A total of 60 animals (10 females and 10 males per cohort) were enrolled in this general toxicity study exploring the safety of a test article. A large array of parameters was compared across the three cohorts including body weight gain, assessment of drug metabolizing enzyme activity, metabolomic analysis, immunologic assessments (lymphocyte subsets, T-cell Dependent Antigen Response [TDAR], serum IgG isotyping), clinical pathology, physical examinations (respiratory, neurologic, cardiovascular, and ophthalmologic examination), pathogen screening, organ weights, as well as gross and microscopic pathology. In this investigation, no notable difference was observed between animals of mixed Asia mainland origin and animals of Cambodia only. Yet, notable differences were observed when Asia mainland animals were compared with Mauritian animals. Mauritian animals had a lower inflammatory cell infiltrate in kidneys, stomach, heart, and lungs, a lower incidence of arteritis, and a lower incidence of lymphoid hyperplasia in spleen and lymph nodes. In addition, Mauritian animals had a lower incidence of morphological observations in the testis (such as tubular dilatation, degeneration, developmental anomaly), and testicular maturity was more advanced in age-matched Mauritian animals when compared with those from Asia mainland. Further differences among Mauritian and Asia animals were a lower absolute B-cell count in Mauritian animals, a higher TDAR antigen response in Mauritian animals, and a higher mean absolute spleen weight in Mauritian animals.
The findings of Kozlosky et al. 37 are in line with reports from Drevon-Gaillot et al. 38 in 2006 and Caruso et al. 39 in 2020. Drevon-Gaillot et al. 38 report a lower grade of lymphoplasmacytic infiltration in the gastro-intestinal tract of Mauritian animals compared with animals from Vietnam or the Philippines. Caruso et al. 39 report a lower incidence of follicular lymphoid hyperplasia in the spleen and a lower incidence of gastric mononuclear cell infiltrates in Mauritian animals versus animals from Asia mainland (Cambodia, China, and Vietnam). However, they report a higher incidence of mononuclear cell infiltrates in the brain and eyes, a higher incidence of mineralization in the brain, and a higher incidence of cell infiltrates in the liver.
Recently, a retrospective in-house analysis was undertaken to compare baseline body weights, organ weights, and clinical pathology parameters in 1310 long-tailed macaques from four different geographical origins (Cambodia, China, Vietnam, Mauritius). 40 Data were segregated for age (2-3 year, 3-4 year, 4-5 year, 5-6 year) and values were compared across cohorts of different geographical origin using analysis of variance and t-tests for pairwise comparisons. Not surprisingly, no statistically significant differences were found between animals from Cambodia, China, and Vietnam, since all three are included in the Indochinese habitat cluster, also referred to as Asia mainland. Between 3 and 5 years of age, a statistically significant (P < .05) higher body weight was found in Mauritian animals compared with Asia mainland animals, with absolute differences of up to 1200 grams for males and up to 500 grams for females (Figure 2). Since body weights were comparable across all cohorts after 5 years of age, this indicates a faster growth rate of Mauritian animals. The higher body weight in Mauritian animals correlated to higher absolute organ weights in this cohort as was demonstrated for spleen, testis, adrenal, kidney, liver, pituitary, and thymus. Differences were also identified in clinical pathology parameters, e.g., lower reticulocyte and white blood cell counts in Mauritian animals, higher red blood cell counts in Mauritian animals and increased gamma-glutamyl transferase activity in Mauritian animals.37,38,41,42 For further reading on clinical pathology parameters in reference to NHP geographical origin, the reader is referred to Arndt et al. 43 in this journal volume.
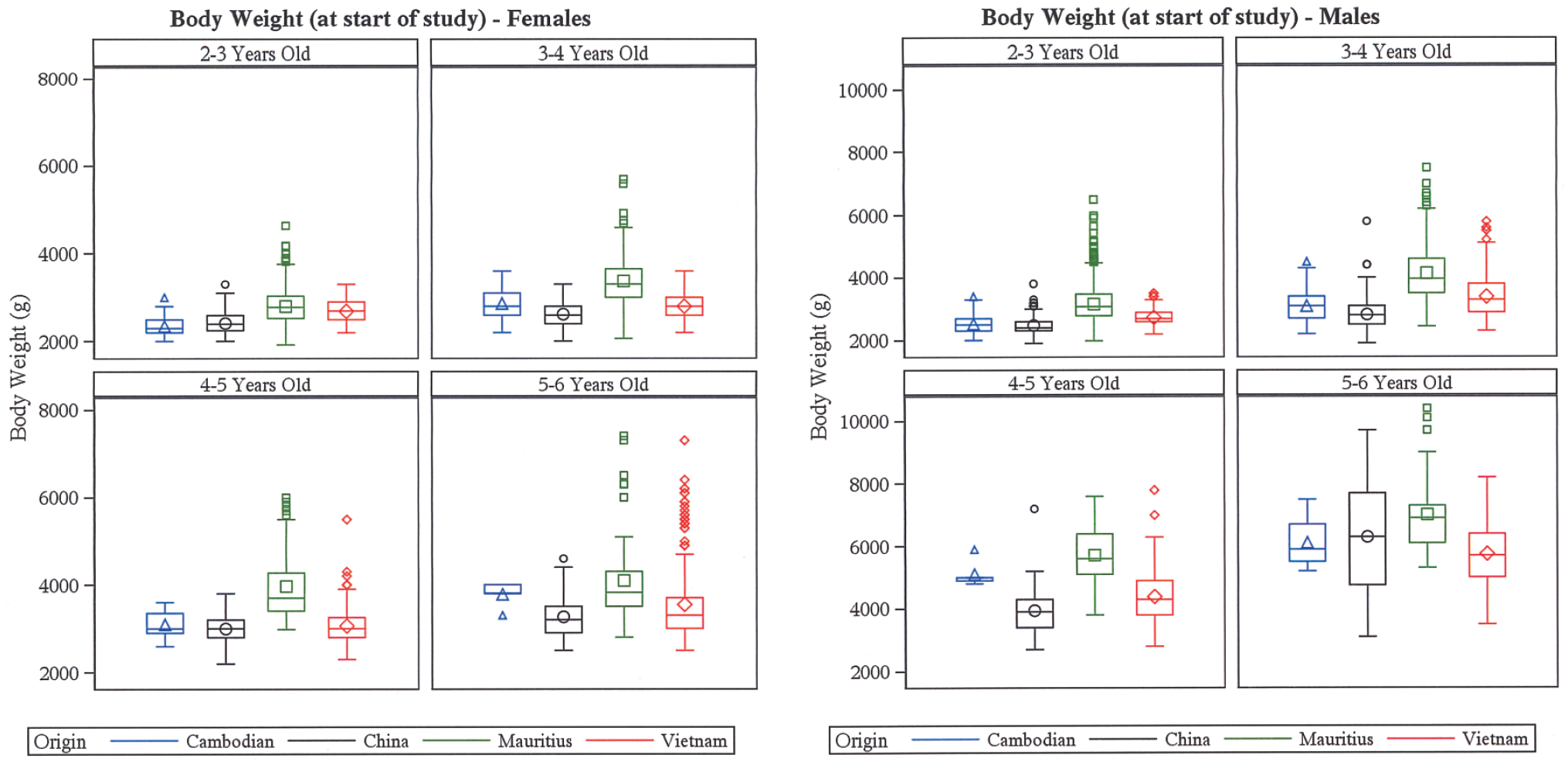
Absolute body weight of female and male long-tailed macaques from 4 different geographical regions—Cambodia (blue), China (black), Vietnam (red), and Mauritius (green)—segregated into age ranges (2-3 years, 3-4 years, 4-5 years, 5-6 years). Box blots show minimum, first quartile, median, third quartile, and maximum. From Ashcroft-Hawley et al. 40
Geographical Origin of Long-Tailed Macaques Used in Developmental and Reproductive Toxicity Studies
Examination of reproductive toxicity in the long-tailed macaque is limited to an indirect assessment of fertility, based upon clinical and morphological endpoints in sexually mature animals. 44
A study published by Luetjens and Weinbauer in 2012 compared sexual maturity in male long-tailed macaques of Mauritian origin and of Asia mainland origin. 45 Sexual maturity was reached earlier in Mauritian animals when compared with Asia mainland animals. Mauritian animals grew at 1.7 kg/year compared with 0.9 kg/year in Asia mainland animals, and average body weight at confirmation of sexual maturity was 5.7 kg vs 4.4 kg (P < .05) in Mauritian vs Asian animals. These observations are supported by the notion that testicular maturity is achieved earlier in Mauritian animals compared with Asia mainland animals 35 and that age-adjusted testes weights are higher in Mauritian animals compared with animals from Asia mainland.
For female animals, baseline menstrual cycle data (based upon daily vaginal swabs using four bleeding intensity categories) 46 was compared between 1174 Asia mainland and 660 Mauritian animals (menstrual cycles with durations shorter than 20 days and longer than 50 days were excluded from analysis). Mauritian animals showed a menstrual cycle duration of 31.5 ± 4.3 days and Asia mainland animals showed a cycle duration of 31.6 ± 5.3 days. Although this minimal difference was statistically significant, it is not considered to represent a biologically meaningful difference. However, the duration of menstruation was significantly longer in Mauritian animals compared with Asia mainland animals (median 2.9 vs 2.6 days) and the menstrual bleeding intensity was significantly higher (Mauritian animals: median 3.0; Asian animals: median 2.5). At the onset of sexual maturity, Mauritian animals were significantly (P < .05) younger and heavier compared with Asia mainland animals (4.0 ± 0.6 years vs 4.7 ± 1.2 years, and 3.75 ± 0.6 kg vs 3.25 ± 0.6 kg).
For developmental toxicity evaluation, time-mated animals are not available from commercial breeders but need to be generated at the test site through a timed mating program.46,47 At the authors’ test site, animals of Asia mainland origin were compared with animals of Mauritian origin (100 control animals from two embryo-fetal development [EFD] and five pre- and postnatal development [PPND] studies) concerning key developmental parameters. Gestation range was 144 to 174 days in Mauritian animals compared with 134 to 184 days in Asia mainland animals. 48 Pregnancy losses were comparable between Mauritian and Asia mainland animals. Pregnancy duration, prenatal loss, stillbirth incidence, and postnatal loss were also comparable. In addition, fetal and infant morphology parameters are comparable (data not shown) as are birth weights, and body weight growth for the first 90 postnatal days. After those 90 days, however, Mauritian infants (male and female) grow at a faster rate compared with Asia mainland animals (Figure 3).
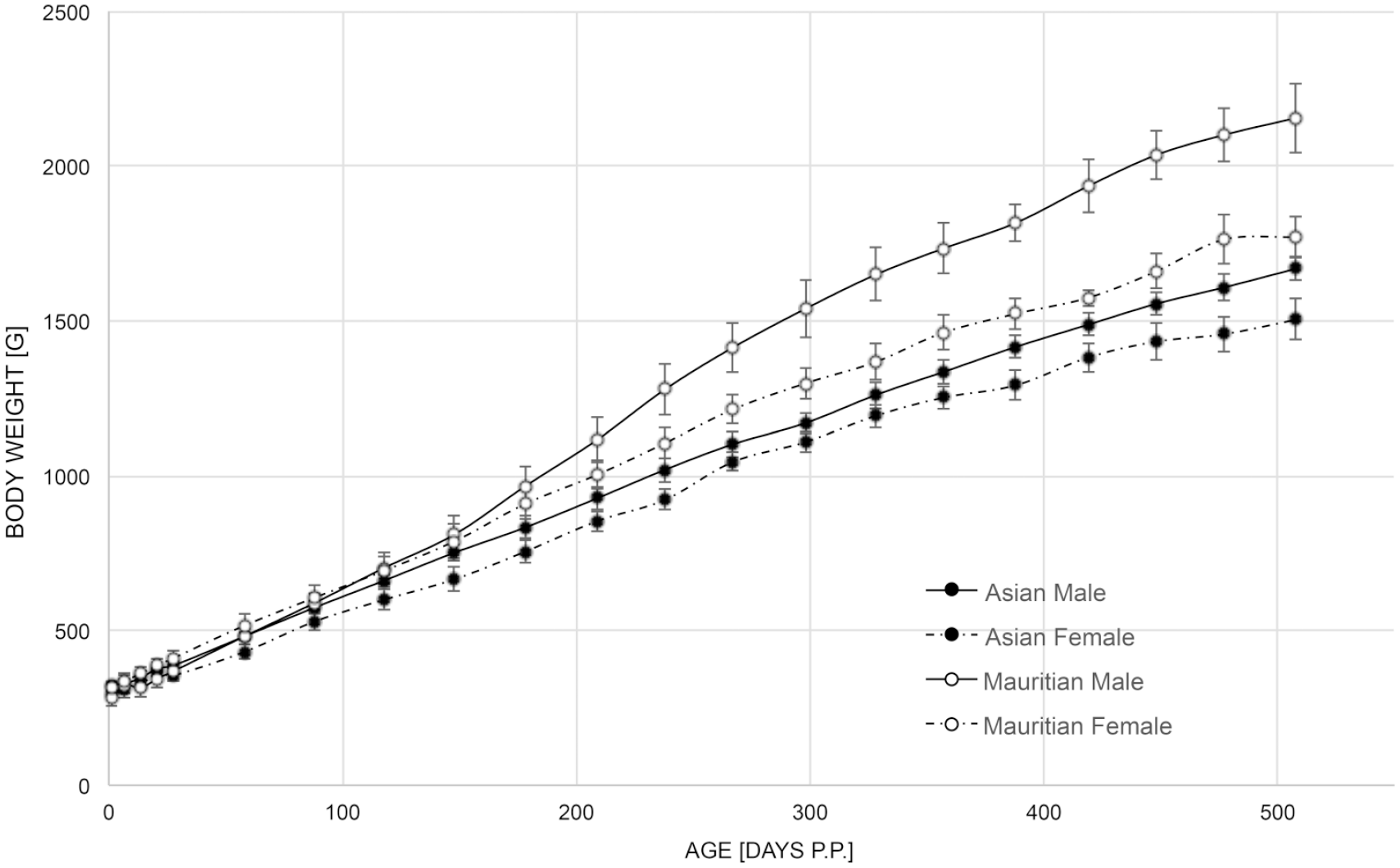
Body weight development in long-tailed macaques of Mauritian (5 males, 5 females) or Asian mainland (13 males, 18 females) origin, starting at the day of birth. g indicates gram; p.p., postpartum.
Further Differences Between Long-Tailed Macaques of Different Geographical Regions
Long-tailed macaques of Mauritian origin have never been exposed to simian herpes B virus, since the virus does not exist on this island. Therefore, these animals have been preferred over animals from Asia who carry a higher risk of transmitting simian B virus to humans and hence need to be tested serologically before they can be exported.
Mauritian animals have been demonstrated to be more susceptible to an experimental infection with Plasmodium coatneyi, 49 but were resistant to an experimental infection with Shigella spp. when compared with animals from Asia. 50 Finally, Mauritian animals demonstrated a stronger graft rejection (kidney allograft survival under cyclosporine immunosuppression) compared with animals from Asia island origin (median graft survival was over 150 days in Philippine animals compared with 26 days in Mauritian animals). 51
Those differences in immune response might be related to known differences in the major histocompatibility complex (MHC). While the MHC of long-tailed macaques is similarly structured to that of humans, it differs by a high degree of polymorphisms and gene duplications, potentially causing variability in the immune response. Mauritian animals and animals from the Philippines have the most restricted MHC polymorphisms, compared with animals from Indonesia or Vietnam. 16
Conclusion
Commercial breeders of long-tailed macaques to be used for scientific purposes are located in Asia mainland (e.g., Cambodia, China, Vietnam), in Asia island (Malaysia, Indonesia and Philippines), and in Mauritius. For the past 10 years, China has exported about 20,000 long-tailed macaques per year, but with start of the COVID-19 pandemic, this export was stopped. 52 Simultaneously, the demand for long-tailed macaques has continuously grown, resulting in a global shortage of animal availability. 53 This shortage has triggered questions on whether cynomolgus macaques of different geographical origin could be used in the same drug development program or even within a single experimental study.
This question is not new, and some experience already exists from a number of medicinal products. For example, Mauritian and Asian origin animals were used within the same drug development program for Rituximab 54 and Gazyva (obinutuzumab). 55 Both programs used Mauritian animals for the assessment of general toxicity and animals from Asia for the assessment of reproductive and developmental toxicity.
Should the shortage of long-tailed macaques continue or even aggravate, the need for combining animals from different geographical origins within the assessment of general toxicity could arise. This is a concern to drug developers, since differences between geographical habitat clusters have been described as detailed in this article. In the authors’ view, differences between the geographical clusters can be accounted for as long as baseline and reference data are available that characterize animals from a particular cluster. Therefore, it is proposed that cynomolgus macaques from the 4 different geographical habitat clusters (Asia mainland, Indonesian-Malaysian, Philippines, Mauritius) can be used within a drug development program, if separate reference databases are maintained for each geographical habitat cluster. It does not appear to be necessary to split reference data within the Indochinese habitat cluster, because differences are very minor. Reference data within the Asia island cluster (Indonesian-Malaysian and Philippines) should be kept separate, as this cluster is very divergent and because relatively little data are available concerning their comparability. Reference data from Mauritian animals should always be separated from the other clusters, since Mauritian animals all originate from the anthropogenic translocation of few animals in the 17th century.
Footnotes
Authors’ Note
Some data were presented as a poster at the 53rd Annual Meeting of the Teratology Society (2013), and at the 52nd Annual Meeting (2013) and 54th Annual Meeting (2015) of the Society of Toxicology.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.