
Abstract
The ganglion of the trigeminal (V cranial) nerve is generally sampled at necropsy in nonrodent toxicology studies only when somatic or autonomic peripheral nervous system toxicity is suspected. The ganglion is far more difficult to locate in nonrodents than in rats and mice, and suitable methods to dissect it have been described only for swine. The trigeminal nerve caudal to the ganglion passes through a canal, roofed by bone in dogs and rabbits and by a tough layer of dura mater in swine and nonhuman primates. The ganglion is partly or wholly obscured by overlying dura mater. Of the 3 intracranial branches of the nerve, the ophthalmic is delicate and the maxillary and mandibular have extremely short courses within the cranial cavity. Methods that are practical in routine toxicologic pathology for the dissection of the ganglion in nonrodent laboratory species are illustrated and relevant species differences in the anatomy of the intracranial part of the trigeminal nerve are highlighted.
Introduction
It is the intention of this article to describe methods that are practical in routine toxicologic pathology for the dissection of the trigeminal ganglion in nonrodent laboratory species and to highlight relevant species differences in the anatomy of the intracranial part of the trigeminal (V cranial) nerve.
The trigeminal ganglion (also known as the Gasserian or semilunar ganglion) has been studied in anatomic and physiologic research. 1 –3 In experimental medicine, lesions are created in it when testing local therapies for trigeminal neuralgia of humans 4,5 and it is a site of naturally occurring latent herpes virus infection, including in humans 6 (herpes simplex and varicella zoster viruses), macaques 7 (macacine herpesvirus 1 or herpes B virus), dogs 8 (canine herpesvirus), and swine 9 (pseudorabies virus).
The trigeminal ganglion is also sampled in toxicology studies when somatic or autonomic peripheral nervous system (PNS) toxicity is suspected 10 and in developmental neurotoxicity testing (DNT), which is carried out on environmental chemicals in rats or mice from gestational day 6 to lactation day 20. 11 It is therefore important that toxicologic pathologists are able to sample it correctly.
The trigeminal ganglion lies lateral to the pituitary fossa, within the dura mater that is applied to the bone of the floor of the cranial cavity. Although in rodents the 3 intracranial branches of the trigeminal nerve are easily visible, 12,13 which facilitates locating the ganglion at necropsy, it is far more difficult to locate in the other species used commonly in toxicology (rabbits, mini-pigs, dogs, and cynomolgus macaques) because the dura mater is thicker and more opaque than in rodents, the distal part of the trigeminal nerve is partly surrounded by bone in some of these species, and the 3 branches are not easily visible from within the cranial cavity.
The dissection of the trigeminal ganglion in the postmortem skull of mini-pigs using an intracranial approach has been illustrated, 14 and an approach via the orbit in newborn pigs has also been described. 15 These publications describe methods that are feasible for harvesting the trigeminal ganglion in routine toxicology studies, where there are relatively large numbers—often 20 to 40—of animals and where no experimental manipulation of the ganglion or nerve in situ is required. There are also descriptions of the anatomy and intracranial dissection of the trigeminal ganglion in rabbits 5 and in monkeys. 1 –3 These publications describe experimental surgical manipulations of the ganglion or branches of the nerve, using dissection microscopy, but are less suitable as guides for toxicological pathology. The author is not aware of published descriptions of methods to harvest the trigeminal ganglion in dogs, although the position of the ganglion in dogs is well described. 16
The trigeminal ganglion is analogous to a dorsal root ganglion of a spinal nerve. In this article, the trigeminal nerve and its branches are considered as starting at the periphery and extending to the brain rather than vice versa, as it is predominantly sensory. The portion of the trigeminal nerve between the ganglion and brain is termed “trigeminal nerve” in this article. The 3 branches of the trigeminal nerve (ophthalmic, maxillary, and mandibular) enter the cranial cavity and converge on the trigeminal ganglion. The trigeminal nerve itself then runs as sensory and motor roots (analogous to the dorsal and ventral roots of a spinal nerve) in a common sheath and meets the ventrolateral brain stem at the junction of the pons and trapezoid body. As illustrated in Figure 1, the motor fibers are all within the mandibular branch, and although the cell bodies of the sensory nerves are within the trigeminal ganglion, the cell bodies of the motor nerves are within the brain. The trigeminal nuclei and tracts are also within the central nervous system.
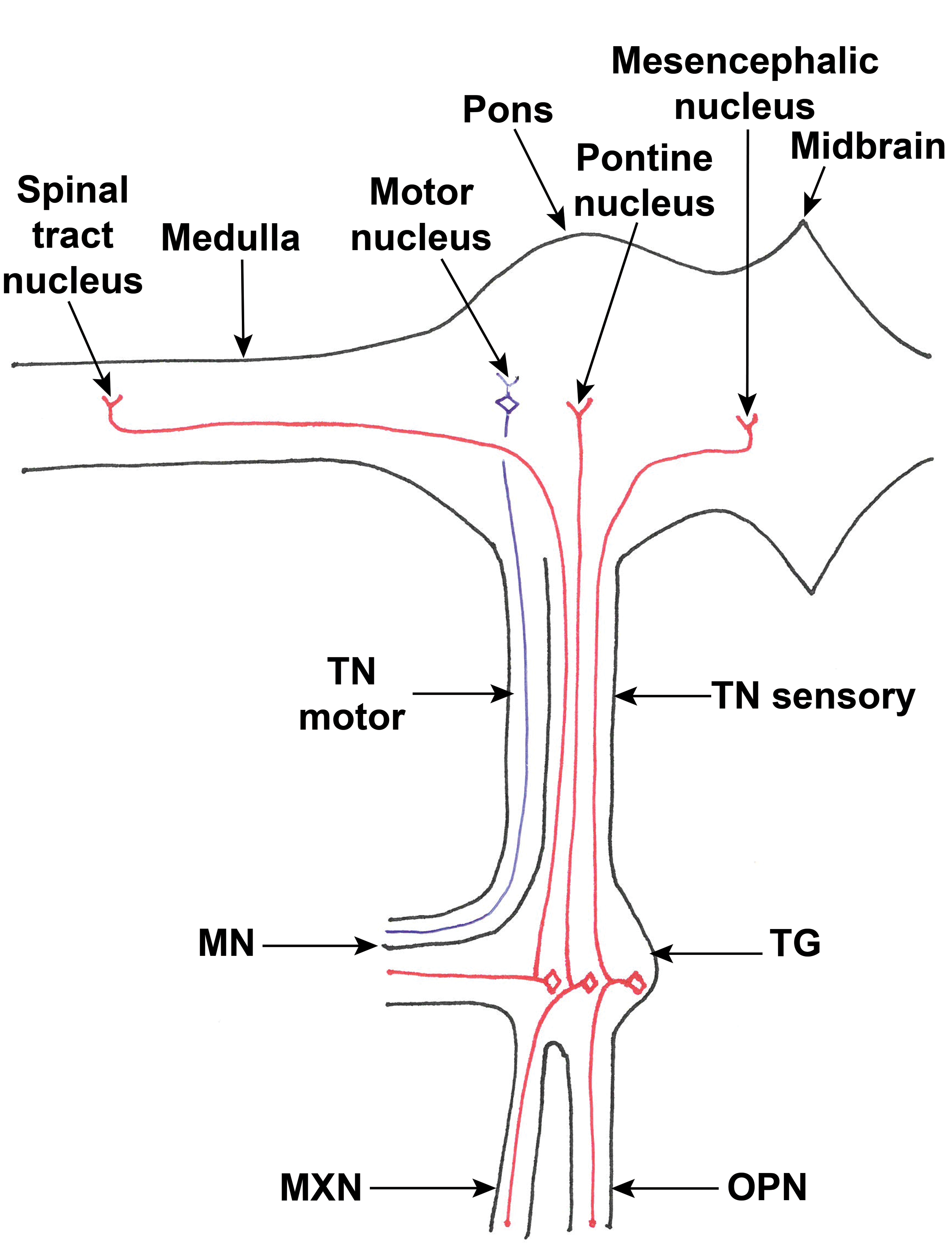
Diagram of the sensory (red) and motor (blue) nerve fibers and the nuclei of the left trigeminal nerve (ventral view). The positions of the terminals of the central axons of the sensory fibers in each of the 3 branches of the trigeminal nerve are shown for illustration only; it is not intended to imply that fibers from the ophthalmic branch all terminate in the mesencephalic nucleus, for example. Quadrilaterals indicate cell bodies; Y shapes indicate nerve or dendrite terminals. MN indicates mandibular nerve; MXN, maxillary nerve; OPN, ophthalmic nerve; TG, trigeminal ganglion, TN, trigeminal nerve (sensory and motor roots indicated).
Methods
Four skulls from each species (New Zealand white rabbits, Göttingen mini-pigs, Beagle dogs, and cynomolgus macaques) were retained for dissection after routine necropsies. The animals were young adult controls in toxicology studies, each given an innocuous vehicle by oral, intravenous, subcutaneous, or rectal routes. The toxicology studies were carried out in accordance with the UK Animals (Scientific Procedures) Act 1986, which conforms to EU Directive 2010/63/EU and achieves the standard of care required by the US Department of Health and Human Services’ Guide for the Care and Use of Laboratory Animals.
Animals of all 4 species were euthanized by intravenous injection of sodium pentobarbitone followed by exsanguination. As part of the routine necropsy, the head was separated from the vertebral column, the roof of the cranial cavity was removed, the falx cerebri and tentorium cerebelli removed, and the brain, pituitary, eyes, intraorbital optic nerves, and lacrimal glands removed for examination as part of the toxicology studies. The cranial nerves were generally cut or broken between their points of emergence from the brain and the points where they were attached to the lining of the cranial cavity. The remaining skull with nasal cavity, which would otherwise have been discarded, was placed in 10% neural-buffered formalin for 4 to 6 months before use for the work described here.
Trigeminal ganglia were dissected from all 4 species, using variations of the methods described for swine 14,15 until feasible methods were established to isolate the ganglia. The gross anatomy of the ganglion, the trigeminal nerve, and its branches were noted and photographs taken to illustrate the methods. The dissected tissues were examined histologically to confirm their identity as ganglia and associated nerves (results not presented). The skull from a CD1 mouse was also photographed at necropsy for comparison, but its ganglion was not dissected. In addition, a photograph taken at necropsy of the unfixed skull of a cynomolgus macaque (kept under Canadian legislation), kindly supplied by Charles River Laboratories, Senneville, Quebec, Canada, is included in this article.
Results
In all species, the dura mater was tightly applied over the trigeminal nerve near the ganglion and over the ganglion itself, although the dura mater could easily be peeled away from the inner aspect of the sphenoid and parietal bones of the side of the cranial cavity. The intracranial course of the trigeminal nerve in the cynomolgus macaque is shown in Figure 2.
Cynomolgus monkey skull, unfixed, craniodorsal view. The brain is reflected caudally, exposing the trigeminal nerve (TN) and the trigeminal ganglion (TG). Two of the trigeminal branches (MX and MN) are visible through the dura. MN indicates mandibular nerve; MX, maxillary nerve; VCN, vestibulocochlear nerve.
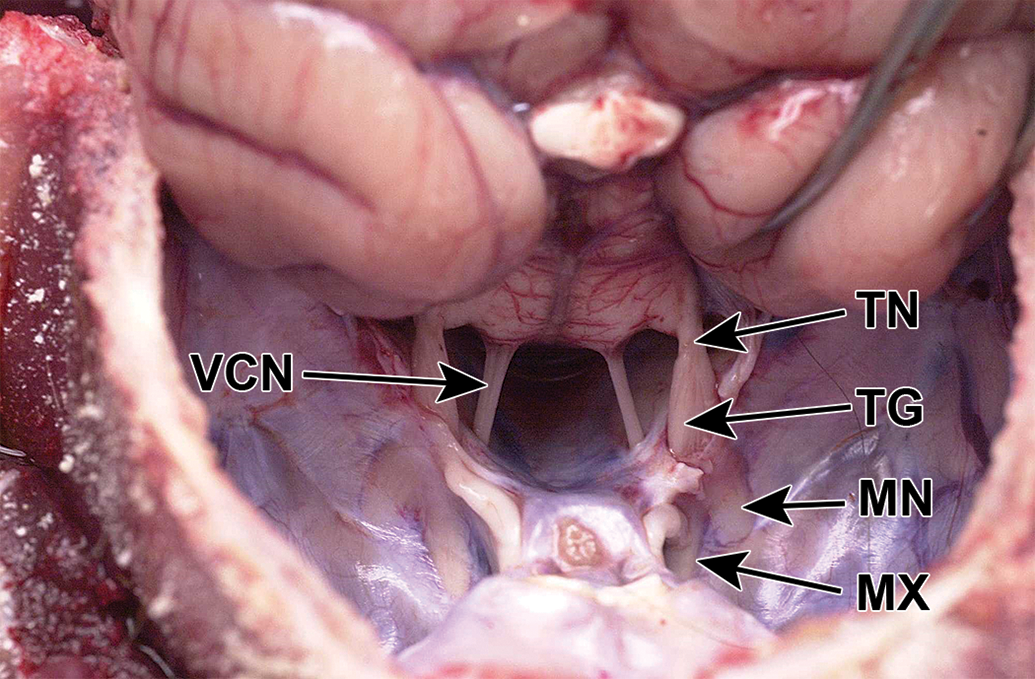
In contrast to the mouse (Figure 3), in the mini-pig and dog, the trigeminal nerve and ganglion and the intracranial parts of the 3 branches were not visible without dissection (Figures 4 and 5). In the cynomolgus monkey and rabbit, the ganglion and/or intracranial portions of the 3 branches could be discerned faintly through the semitransparent dura mater (Figure 2).
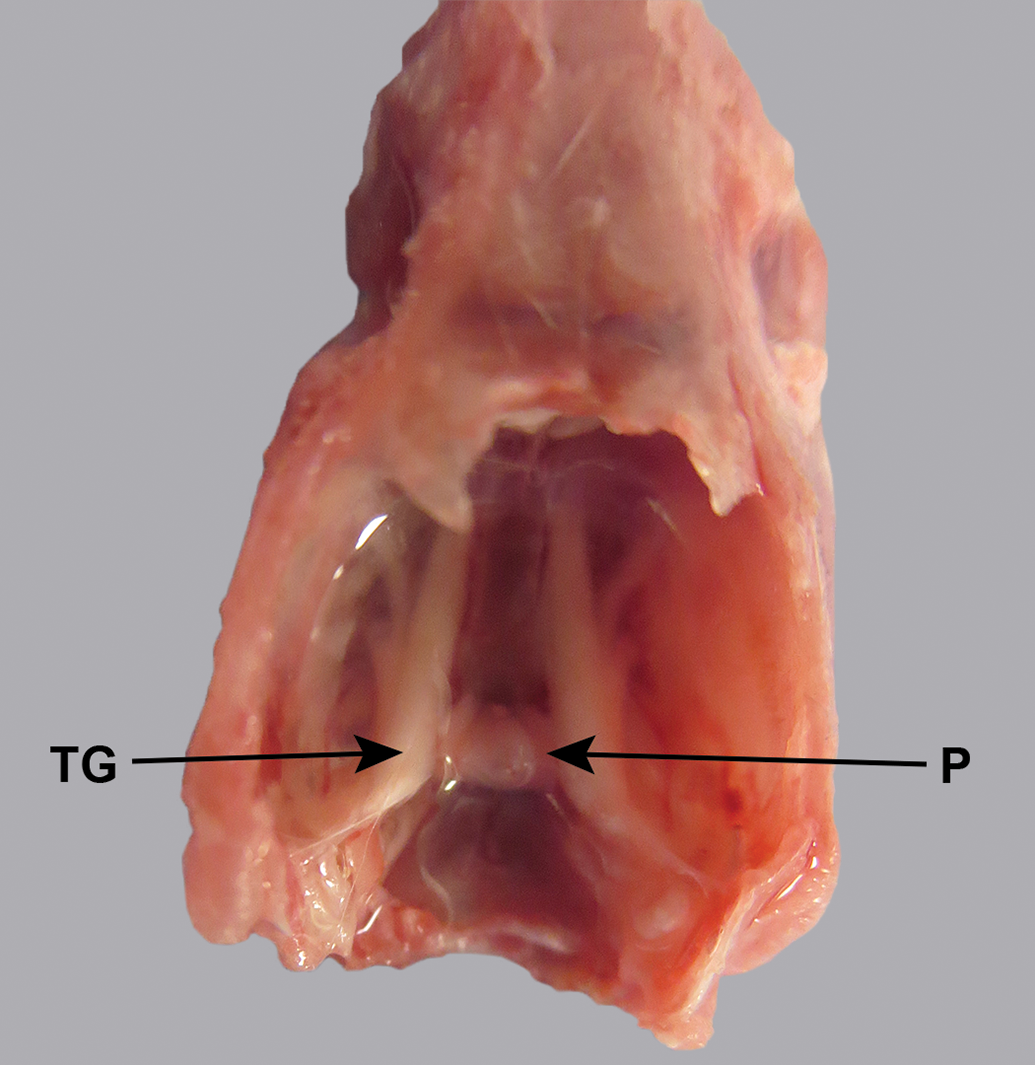
CD1 mouse skull, unfixed, dorsal view. The trigeminal nerve and its branches are visible through the dura mater. P indicates pituitary; TG, trigeminal ganglion.
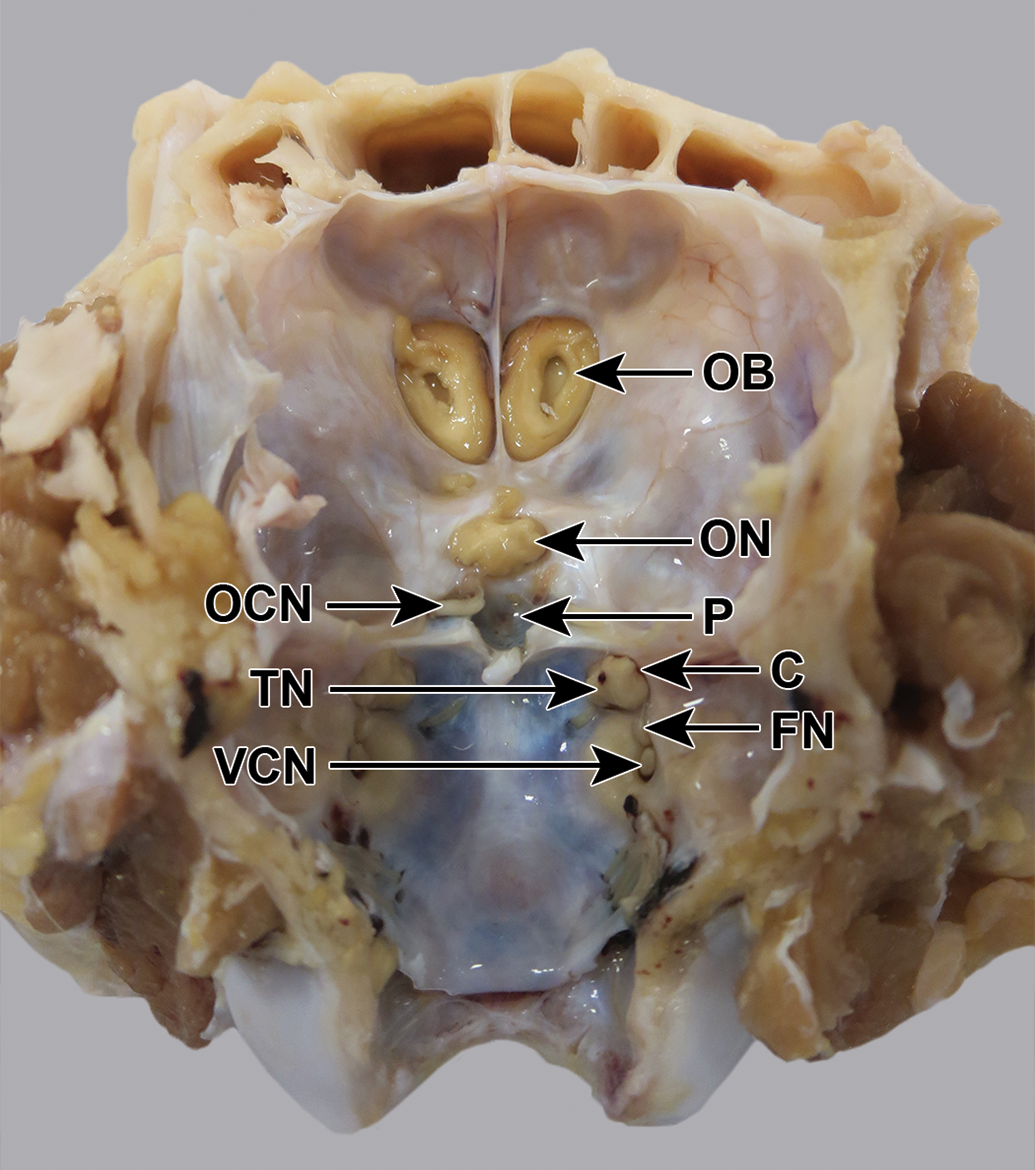
Göttingen mini-pig skull, fixed, undissected, dorsocaudal view. The trigeminal nerve (TN) is visible emerging from a canal (C), but the ganglion and the 3 branches are not visible. FN indicates facial nerve; OB, olfactory bulb; OCN, oculomotor nerve; ON, optic nerve; P, pituitary fossa; VCN, vestibulocochlear nerve.
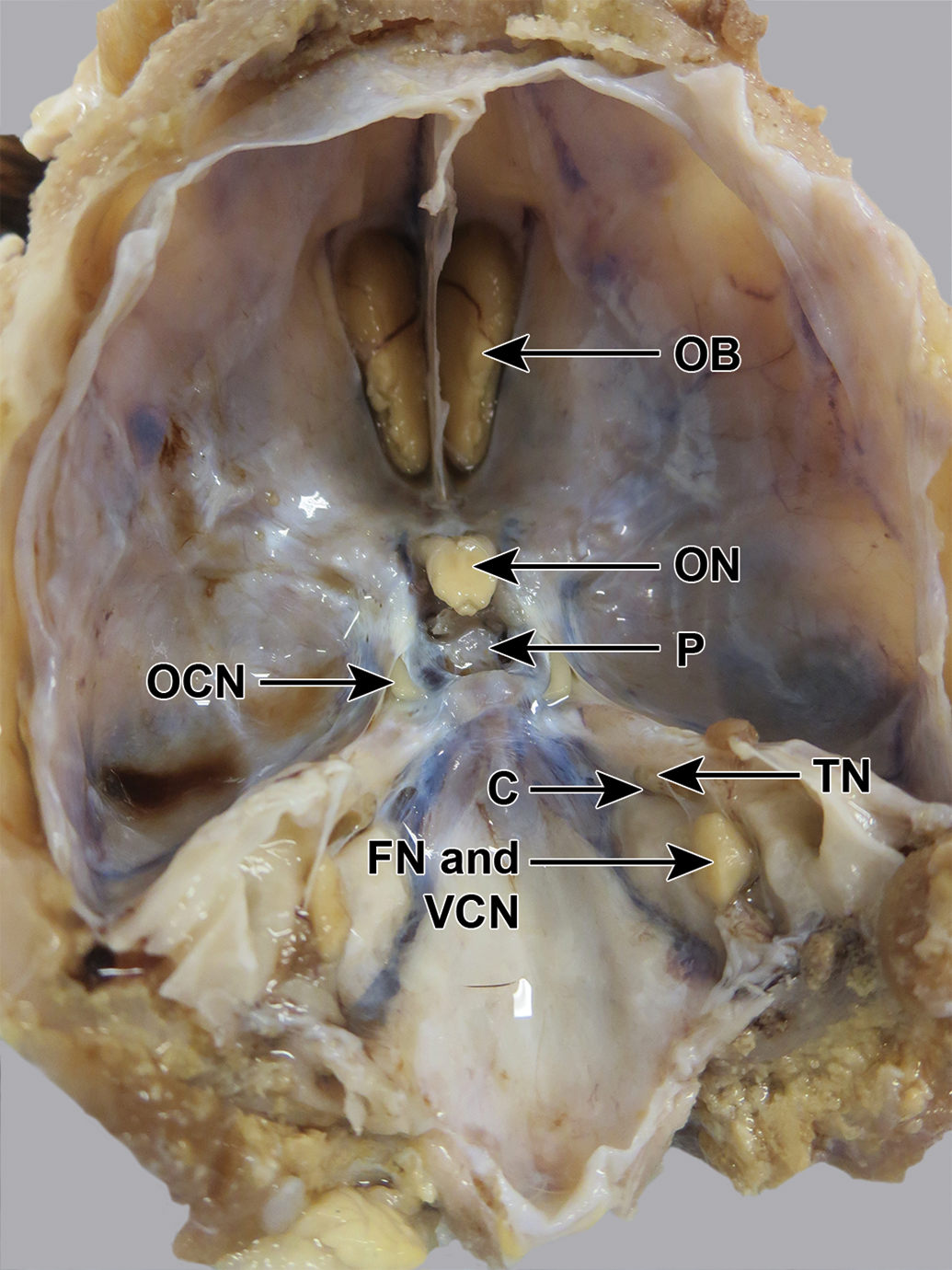
Beagle dog skull, fixed, undissected, dorsocaudal view. The trigeminal nerve (TN) is visible emerging from a canal (C), but the ganglion and the 3 branches are not visible. FN indicates facial nerve; OB, olfactory bulb; OCN, oculomotor nerve; ON, optic nerve; P, pituitary fossa; VCN, vestibulocochlear nerve.
Mini-Pig
The trigeminal nerve caudal to the ganglion passed medially to the petrous temporal bone and immediately lateral to the cavernous sinus, which, when the dura mater overlying it was removed, could be seen to contain the well-developed arterial rete. Immediately cranial to the tip of the petrous temporal bone, there was a roof of tough dura mater dorsal to the nerve, which was cut away using fine scissors. The ganglion was then visible cranial to this as a structure of a slightly greater diameter than the nerve, but with a similar color (Figure 6). The ophthalmic nerve was visible ventral to the optic nerve and was transected. The cranial end of the ganglion extended cranioventrally to be covered by bone and the ganglion lay in a bony groove, so to isolate the ganglion fully, the bone dorsal, cranial, and lateral to the ganglion was chipped away using small bone forceps, and the part of the skull and wall of the orbit cranial to the ganglion were also removed. This enabled the ganglion and a good length of the ophthalmic, maxillary, and mandibular branches cranial and ventral to the ganglion to be seen.
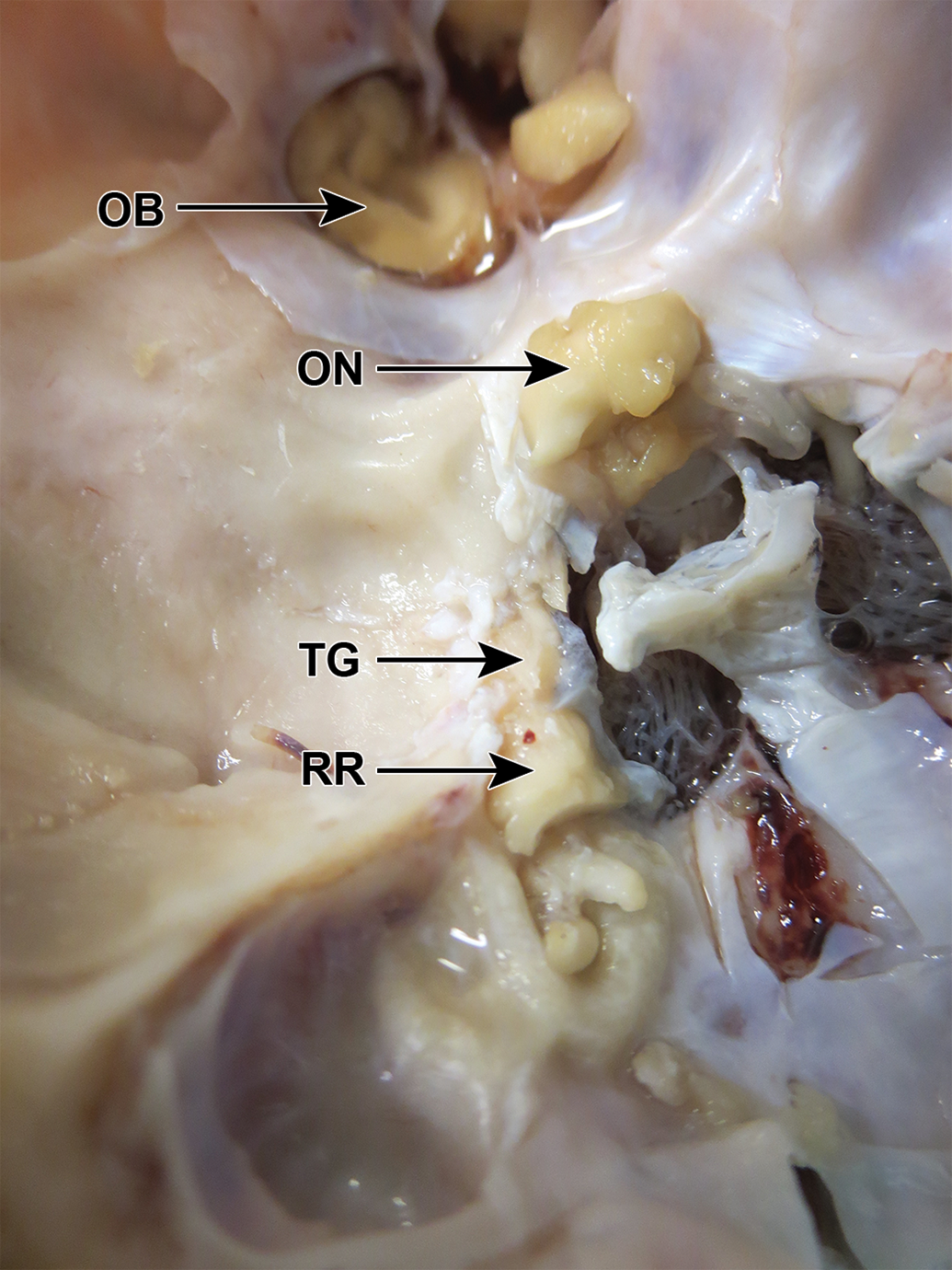
Göttingen mini-pig skull, fixed, dorsocaudal view, after removal of the roof of the canal (RR) on the left side, to expose the trigeminal ganglion (TG), which was the same color as the nerve. OB indicates olfactory bulb; ON, optic nerve.
Dog
The trigeminal nerve caudal to the ganglion passed through a canal in the petrous temporal bone (Figure 5). The roof of this canal was chipped away using small bone forceps, and the dura mater over the ganglion removed with fine scissors. This revealed the ganglion, which was in some cases brown, in contrast to the cream color of the nerve, but in other cases was the same color as the nerve. Figure 7 shows the further dissection of the ganglion in a fixed canine skull, where removal of bone lateral to the petrous crest has revealed the mandibular branch extending dorsally from the round foramen to meet the ganglion. The ophthalmic nerve was visible as the narrowest and most dorsal of the 3 branches, running from the caudal wall of the orbit to meet the ganglion. By further removal of bone cranial to the ganglion, the maxillary branch could be followed cranially, and the ganglion could be excised with a good portion of maxillary nerve.
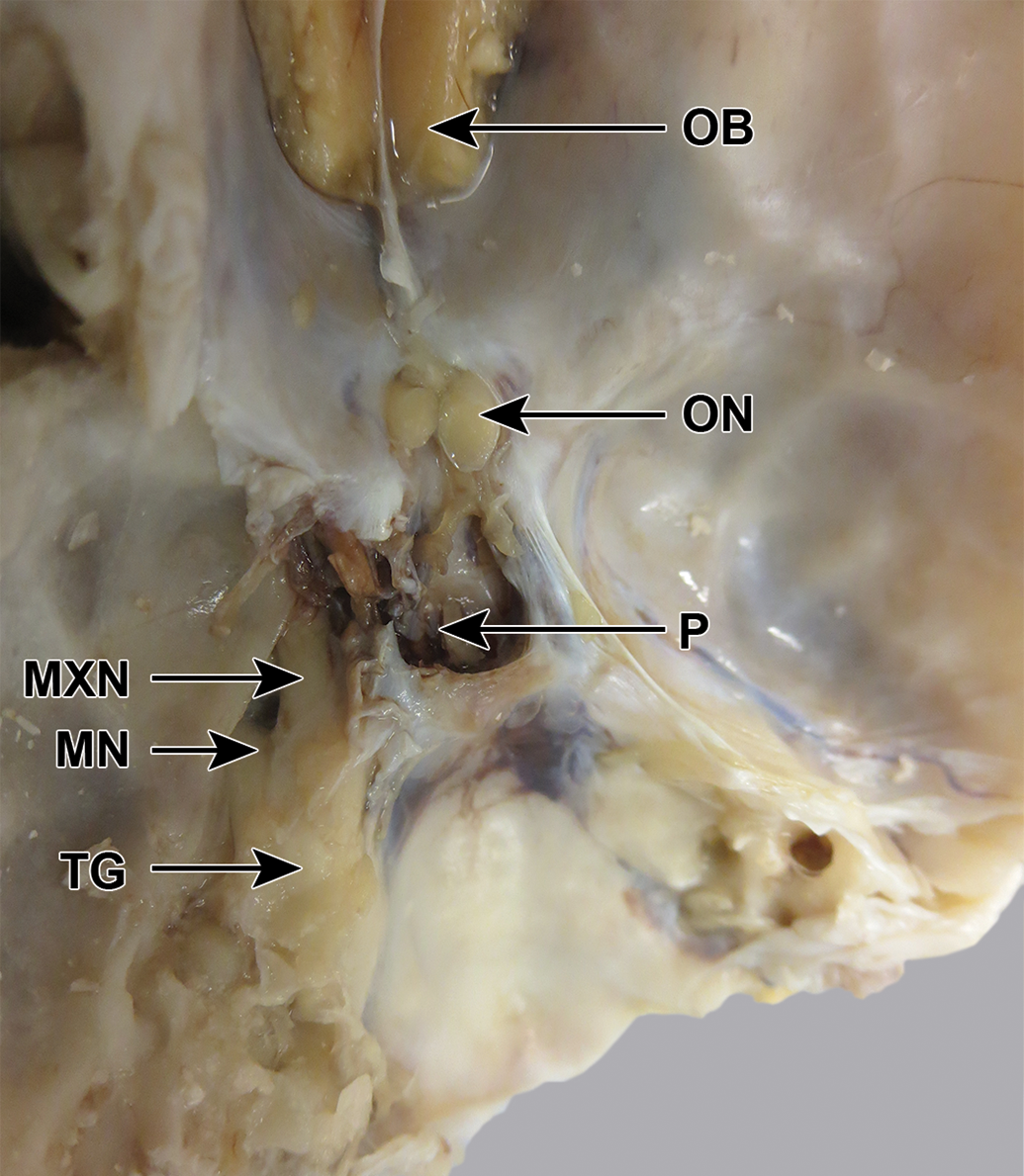
Beagle dog skull, fixed, dorsocaudal view, after removal of some of the bone lateral to the trigeminal ganglion (TG), to reveal the mandibular (MN) and maxillary (MXN) branches. The ophthalmic branch has been removed. OB indicates olfactory bulb; ON, optic nerve; P, pituitary fossa.
Cynomolgus Monkey
Before dissection (Figure 8), the trigeminal nerve could be seen emerging from a canal, roofed by dura mater. The ganglion was flattened in the lateral to medial direction and was a similar color to that of the nerve caudal to it. After removal of the dura mater over the ganglion, the 3 branches were visible dorsally, cranially, and ventrally to the ganglion. The ophthalmic branch was thin, had tightly applied dura mater, and was visible ventral to the optic and oculomotor nerves (Figure 9). The ganglion and branches were dissected free (Figure 10).
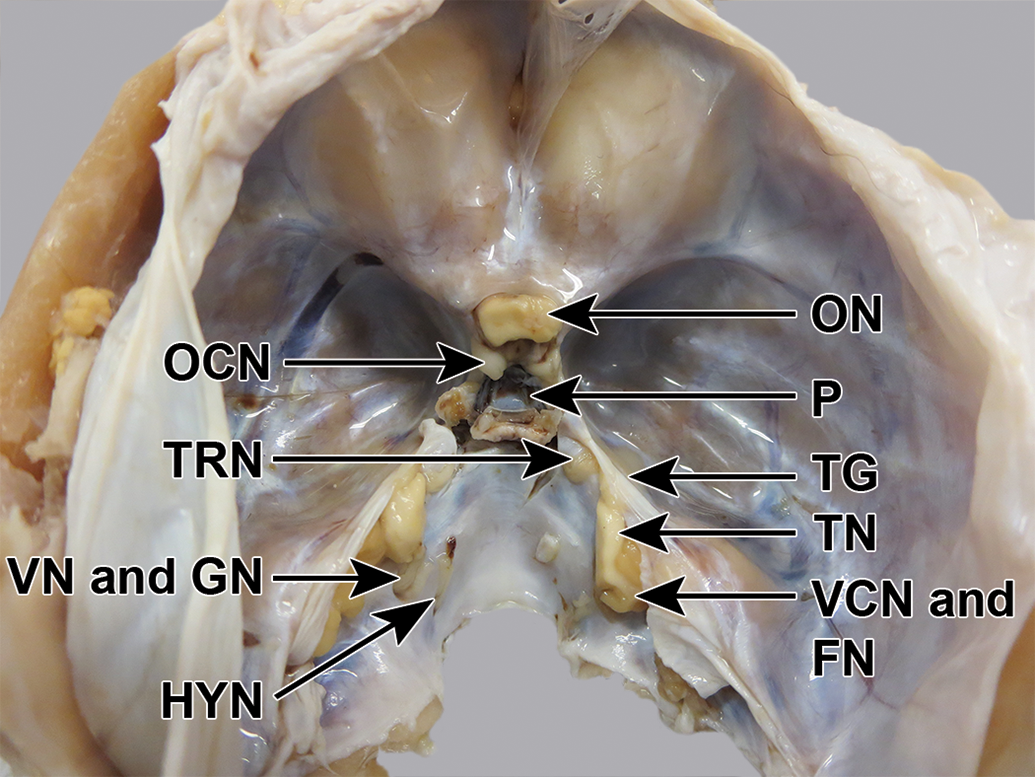
Cynomolgus monkey skull, fixed, undissected, dorsocaudal view. The trigeminal ganglion (TG) is faintly visible through the dura mater. A small part of the brain, including the origins of the trigeminal nerve (TN), facial nerve (FN), and vestibulocochlear nerve (VCN), has accidentally been left behind in the skull when the brain was removed. GN indicates glossopharyngeal nerve; HYN, hypoglossal nerve; OCN, oculomotor nerve; ON, optic nerve; P, pituitary fossa; TRN, trochlear nerve; VN, vagus nerve.
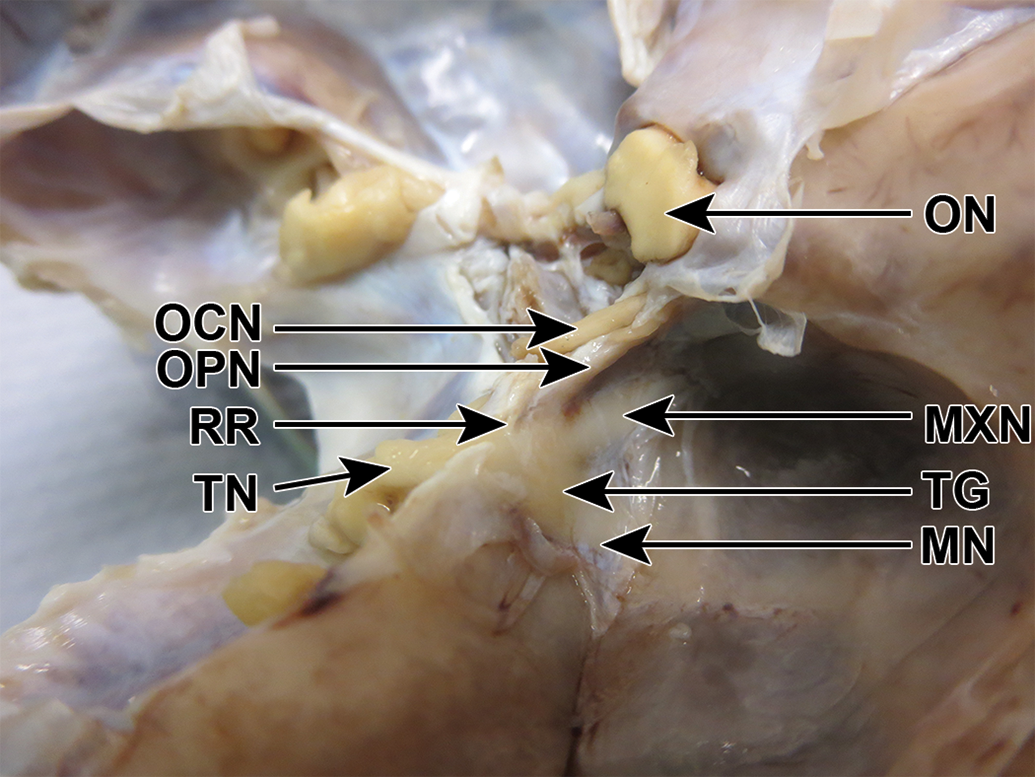
Cynomolgus monkey skull, fixed, dorsolateral view. On the right side, the roof (RR) of dura mater over the trigeminal nerve (TN) has been removed. The trigeminal ganglion (TG) and its 3 branches are visible. MN indicates mandibular nerve; MXN, maxillary nerve; OCN, oculomotor nerve; ON, optic nerve; OPN, ophthalmic nerve.
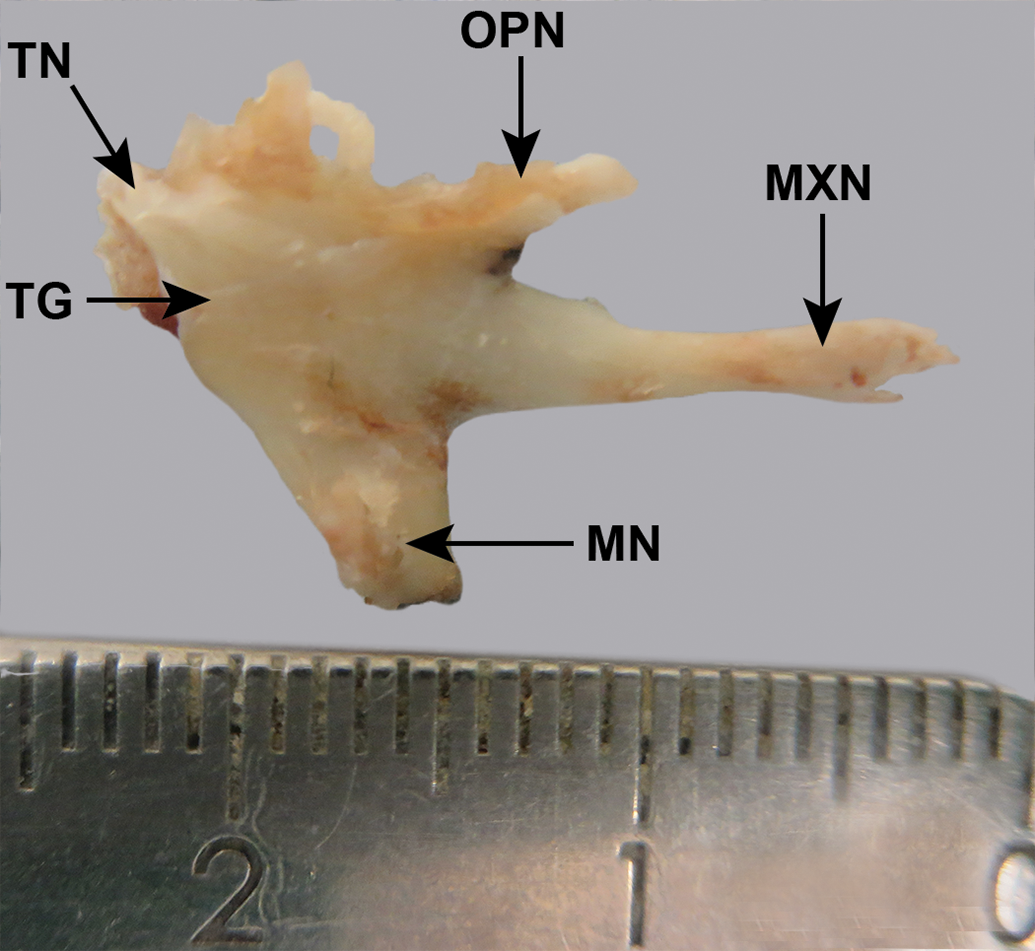
Cynomolgus monkey, excised, fixed, trigeminal ganglion (TG) and 3 branches of the trigeminal nerve. The trigeminal nerve itself (TN) has been largely removed. Scale: mm. MN indicates mandibular nerve; MXN, maxillary nerve; OPN, ophthalmic nerve.
Rabbit
The trigeminal nerve caudal to the ganglion passed through a canal in the bone (Figure 11, right side). The roof and lateral wall of this canal were chipped away using fine scissors. The dura mater was peeled back, revealing the ganglion, which was a pink color, in contrast to the cream color of the nerve (Figure 11). The ganglion (approximately 1 cm long) was dissected free.
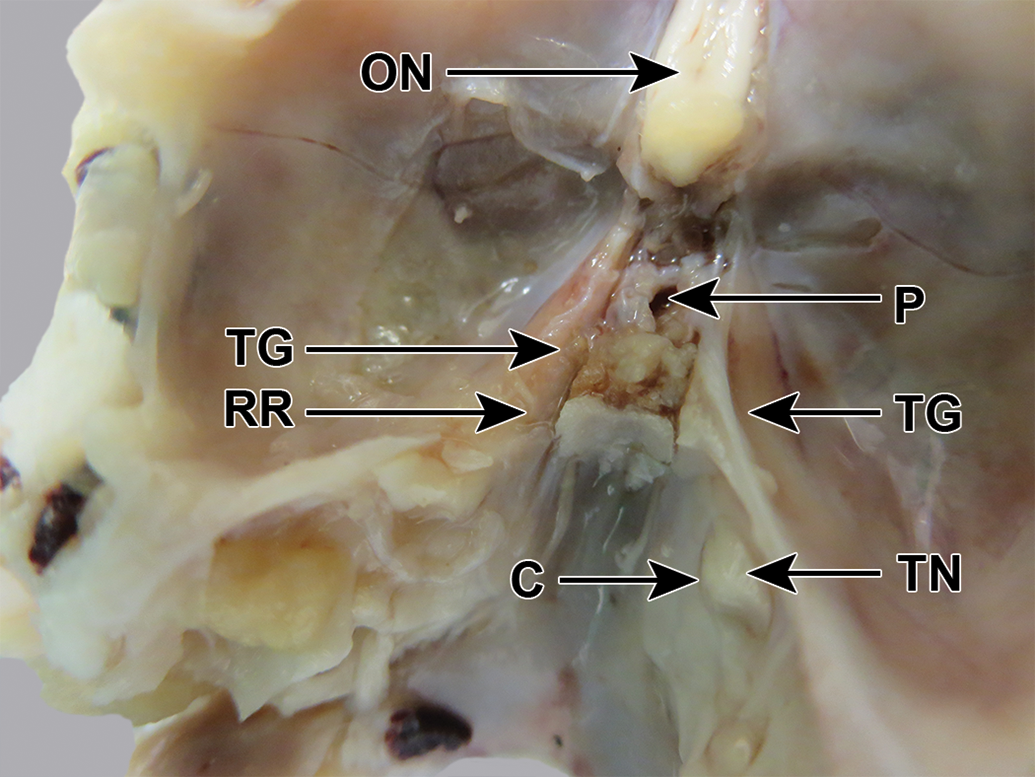
New Zealand white rabbit skull, fixed, dorsocaudal view. The right side is undissected; the trigeminal ganglion (TG) is visible through the dura mater and the trigeminal nerve (TN) can be seen emerging from a canal (C). The left side has been partly dissected by removing the roof (RR) of the canal, to expose the pink trigeminal ganglion (TG). ON indicates optic nerve; P, pituitary fossa.
Discussion
Sampling the trigeminal ganglia at necropsy in nonrodent studies may be challenging for a laboratory that does not specialize in neuropathology, because suitable methods are described only for swine. The trigeminal ganglion is sampled in toxicology studies, including in nonrodents, when somatic or autonomic PNS toxicity is suspected. In addition, pathology evaluation according to recommendations for DNT is occasionally included in juvenile toxicology studies on pharmaceuticals for human use, even if nonrodents are used and even if there is no indication that the nervous system will be targeted. The Draft ICH S11 guideline, under public consultation, 17 proposes that the selection of any additional central nervous system assessments in juvenile studies should be based only on particular concerns identified in a weight-of-evidence evaluation, so once this guideline is finalized, this situation may become even less frequent.
The ganglion is not visible without dissection in the dog and swine, and even in cynomolgus monkeys and rabbits, it is not as distinct as in rodents. Knowledge of its location is, therefore, essential to its correct sampling. In humans, the ganglion lies in a depression on the dorsal surface of the petrous part of the temporal bone, covered by dura mater (forming the trigeminal, or Meckel’s, cave). In dogs, the trigeminal ganglion lies between the apex of the petrosal crest dorsally and the basisphenoid bone (which forms the base of the skull in the area of the pituitary fossa) ventrally. 16 The ganglion in the mini-pig was in a similar position and orientation to that in the dog, although it was not necessary to remove bone to expose it. The present work illustrates that in rabbits and cynomolgus monkeys, the ganglion is situated on the lateral face of the crest formed by the petrous temporal bone and the dura mater rostral to this bone.
In dogs and rabbits, the nerve just caudal to the ganglion passes through a bony canal. 16,18 In dogs, this canal is in the base of the petrosal crest of the petrous temporal bone. In rabbits, the canal is an arch formed by the cranial tip of the petrosal crest meeting the caudal extension of the caudal clinoid process 18 ; in dogs, these 2 bones do not meet. No bony canal is described in humans, and the present work did not detect one in pigs or cynomolgus monkeys, although there was a roof of tough dura mater dorsal to the nerve in these species.
Methods for the dissection of the trigeminal ganglion in nonrodent laboratory species are described, which may be useful in toxicologic pathology, where this ganglion is required to be examined if PNS toxicity is suspected.
Footnotes
Acknowledgments
Steve Côté of Charles River Laboratories, Canada, who supplied the photograph of the unfixed macaque specimen and Drs F. Goulet and A. Bradley who commented on the text.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: The author is an employee of Charles River Laboratories, which conducts studies of the types discussed in this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.