
Abstract
Toxic peripheral neuropathies are an important form of acquired polyneuropathy produced by a variety of xenobiotics and different exposure scenarios. Delineating the mechanisms of neurotoxicants and determining the degenerative biological pathways triggered by peripheral neurotoxicants will facilitate the development of sensitive and specific biochemical-based methods for identifying neurotoxicants, designing therapeutic interventions, and developing structure–activity relationships for predicting potential neurotoxicants. This review presents an overview of the general concepts of toxic peripheral neuropathies with the goal of providing insight into why certain agents target the peripheral nervous system and produce their associated lesions. Experimental data and the main hypotheses for the mechanisms of selected agents that produce neuronopathies, axonopathies, or myelinopathies including covalent or noncovalent modifications, compromised energy or protein biosynthesis, and oxidative injury and disruption of ionic gradients across membranes are presented. The relevance of signaling between the main components of peripheral nerve, that is, glia, neuronal perikaryon, and axon, as a target for neurotoxicants and the contribution of active programmed degenerative pathways to the lesions observed in toxic peripheral neuropathies is also discussed.
Keywords
Introduction
Peripheral neuropathy is an important clinical condition having almost 15% prevalence in the population over age 40 in the United States. 1,2 The etiologies of peripheral neuropathies are quite varied including metabolic and infectious diseases, hereditary conditions, and nutritional deficiencies. Toxic neuropathies that are produced by xenobiotics targeting components of the peripheral nervous system represent another important cause of acquired polyneuropathy. This review will present an overview of the general concepts of toxic peripheral neuropathies with the goal of providing insight into why certain agents target the peripheral nervous system and produce their associated lesions.
Toxic neuropathies can be environmental, occupational, recreational, or iatrogenic, and the prevalence of their cause is governed by geographical and economic factors. In developed countries, the most common cause of toxic neuropathy is drug toxicity, particularly that associated with chemotherapy treatments. 3 The prevalence of chemotherapy-induced peripheral neuropathy was reported to be as high as 68% within the first month of the end of therapy and can be expected to increase as cancer survivorship rates increase. 4 In developing countries, environmental and occupational exposures to various agents including arsenic, lead, mercury, and organophosphorous compounds are significant peripheral neurotoxicants. 5 Additionally, as many manufacturing processes have moved to less developed areas implementing less strict monitoring of occupational exposures, previously recognized peripheral neurotoxicants including hexane, carbon disulfide, and newer chlorofluorocarbon replacing agents like 1-bromopropane remain potential risks for peripheral neuropathy. 6 –8 Two of the most prevalent causes of polyneuropathy, diabetes mellitus and alcoholism, could arguably be considered toxic neuropathies due to the involvement of excess glucose and effects of ethanol. Those neuropathies are beyond the scope of this review and covered in detail elsewhere. 9 –14
Peripheral neurotoxicants can be categorized in numerous ways including their chemical structure, primary use, or the clinical signs they produce. A commonly used approach for classifying peripheral neuropathies is based upon the primary structural target involved, with those exhibiting initial effects in the neuronal perikaryon, axon, or the Schwann cell referred to as neuronopathies, axonopathies, or myelinopathies, respectively. 15 –18 The synapse is also important for the nervous system’s function of transmitting information across space and is the site of action for many pharmacologic and toxic agents. Most of the agents that affect the synapse do not produce structural lesions in the peripheral nervous system and the magnitude of compounds recognized and their mechanisms are beyond the scope of this review.
The specific clinical presentation of a toxic neuropathy will be dependent upon the severity and structural target affected, but there are some concepts that generally hold regarding peripheral neuropathies. 2,3,19 Typically, they are dose dependent, symmetrical, and reversible given adequate time following cessation of exposure. However, for some agents, a condition referred to as “coasting” in which clinical signs continue to progress or persist for a significant period of weeks to months following cessation of exposure has been observed. 20 –22 The most common presentation of toxic neuropathy involves the largest diameter longest axons associated with numbness, paraesthesia, or weakness in a stocking glove distribution. 19,23,24 This is a reflection of the fact that the large long axons are ultimately affected whether or not they are the primary critical targets. Accordingly, hind limb grip strength in rodent functional observation batteries has been used as a sensitive indicator for toxic neuropathies potentially signaling impairment prior to detectable morphological lesions. 25 Many peripheral neurotoxicants not only injure components within the peripheral nervous system but may also have central nervous system involvement, optic neuropathy, rhabdomyopathy (ie, skeletal muscle lesions), or other systemic involvement. Predisposing factors that can contribute to the risk of toxic neuropathy include diabetes mellitus, certain paraneoplastic conditions, genetics, and medical treatments having neurotoxic side effects.
General Concepts of Peripheral Nervous System Structure and Function Relevant to Mechanisms of Toxic Neuropathies
A principal function of the peripheral nervous system is the rapid transmission of signals over a substantial distance. To accomplish this function, it has developed unique structural and physiological characteristics. These characteristics present distinct vulnerabilities that are targeted by certain neurotoxicants and contribute to their target organ specificity. Additional factors that can contribute to peripheral nervous system target organ specificity including specific uptake transport systems that can concentrate toxic substances in some types of neurons and the compromised blood–nerve barrier at peripheral ganglia. 26 –29 When investigating mechanisms of peripheral neurotoxicants, the first step is often to examine morphological end points to identify and characterize the initial lesion. However, caution in interpreting changes must be exercised in that no component can be affected without accompanying changes in the other components. Although an axonopathy or myelinopathy may be observed, it is not immediately evident whether these are primary or secondary conditions. Thus, an understanding of the temporal and spatial changes, both primary and secondary, is essential to designing follow-up analyses to ascertain the initial site of action for a neurotoxicant. Additionally, as our knowledge of intercellular signaling between glia and axons and molecular responses to injury advance, there is promise for the development of sensitive and specific biochemical markers to aid in identifying initial targets of neurotoxicants.
That secondary changes occur in Schwann cells associated with axons experiencing pathological conditions is not surprising when the interdependence of the two is considered. Axons together with Schwann cells achieve their concerted goal through not only their unique structural composition and close physical proximity but also through dynamic communication that regulates the development, maintenance, and function of each other. 30 –34 A variety of molecules has been implicated in signaling between peripheral axons and Schwann cells including myelin-associated glycoprotein, 35 low-affinity nerve growth factor receptor (p75), 36 insulin-like growth factor 1, 37 and transforming growth factor beta. 38 The growth factor neuregulin 1 (NRG1) and the erbB receptors, tyrosine kinases structurally related to the epidermal growth factor receptor, are critical regulators in development and maintenance for all of the Schwann cell phenotypes. Schwann cells express erbB2 and erbB3 receptors, and multiple NRG1 isoforms (derived from the same gene) are expressed by neurons in the central and peripheral nervous systems that project their axons into the periphery. 39 –42 Studies support a pivotal role for NRG1 signaling through erbB receptors in promoting expansion and survival of Schwann cells during development and in determining which axons will be myelinated. Homozygous knockout of erbB2 or heterozygous knockout of NRG1 type III but not type I results in hypomyelination, while overexpression of NRG1 increases myelin thickness suggesting a role of the axon in regulating the thickness of its myelin sheath. 43,44 Interruption of axonal NRG1 growth factor expression through primary axonal injury would compromise this signaling and could contribute to the observed alterations in Schwann cell function and structure accompanying axonal injury.
The case for a reciprocal dependence of axons on signaling and support from associated Schwann cells is also strengthening from experimental evidence. 45 –47 The neuronal soma has long been considered the most important if not the only source of protein biosynthesis for neurons that is then distributed via axonal transport. The presence of Nissl substance, a large accumulation of rough endoplasmic reticulum, in the soma is a testament to the protein synthetic capacity of the neuron. However, recent studies have supported the ability of protein synthesis to also occur within the axon. 31 The protein translated in the axon could contribute to multiple processes including growth, maintenance, and responses to injury and it has been shown that transcription factors are produced in developed axons and transported retrograde to the soma possibly to provide information on the status of the axon during injury. 48 Two possible sources for the messenger RNA (mRNA) used to translate proteins in axons have been recognized. One source is perikaryon-derived mRNA which has been detected in axons. 49,50 The other source is Schwann cell-derived RNA transferred to axons. This concept was introduced by Singer and Green 51 from experiments they performed using the distal ends of transected nerves, and subsequent studies have supported their observations. 52,53 The ability of Schwann cells to transfer ribosomes to axons has also been reported. 54,55 After injury, transfer of Schwann cell RNA appears concentrated in nodes of Ranvier and Schmidt-Lanterman incisures, the sites of initial myelin structural changes during Wallerian degeneration. That the transfer of RNA and organelles could occur through 2 plasma membranes is remarkable. Currently, it is not understood how this interchange proceeds but it has been suggested that exosomes play a role based upon observations of vesicles with exosomal characteristics in Schwann cells and axons under normal and regenerative conditions. 56 –58
The interdependence of Schwann cells and axons through signaling not only affords a potential target for neurotoxicants but also suggests that monitoring this interaction may provide a biochemical approach to detecting neuropathy. One marker that has shown promise for assessing the integrity of the Schwann cell axon relationship is p75, which is expressed on Schwann cell precursors and nonmyelinating Schwann cells, but not on myelinating Schwann cells. Accordingly, p75 levels are normally low in peripheral nerve but are upregulated in primary demyelinating conditions and as a secondary response to axon injury. This suggests that p75 expression could serve as a marker for neuropathy and upregulation of p75 has been detected in the sciatic nerves of rats administered isoniazid, acrylamide, or carbon disulfide at time points prior to the development of morphological or behavioral disturbances. 59,60 Additionally, increased levels of p75 protein in serum have been observed in animal models for diabetic neuropathy and nerve crush injury. 61 Therefore, analysis of p75 either at the protein or message level could provide a biochemical method for early detection of neuropathy. There is also the potential that analysis of the temporal relationship of p75 expression to other markers of injury could provide insight into the type of neuropathy. For example, if myelin-specific genes such as myelin protein zero (P0) or myelin basic protein (MBP) were downregulated early prior to changes in axonal markers, it would be consistent with a primary myelinopathy.
Examples are scarce for which a single mechanism is generally accepted to account for all the pathological events observed in a given toxic neuropathy. The major points of contention when considering mechanisms tend to center on the initial critical event and the most relevant molecular target. Typically, the more a neurotoxicant is investigated the greater the number of interpretations that exist, and for most neurotoxicants, there are several potential credible contributing events that are not necessarily mutually exclusive. Ultimately, however, there are a smaller number of general pathogenic processes through which toxic agents mediate their effects. Among these processes are covalent or noncovalent modifications, compromised energy or protein biosynthesis, oxidative injury, and disruption of ionic gradients across membranes. These general mechanisms are common to most toxicants and what bestows selective toxicity for the peripheral nervous system is an ability to target a vital process or structure unique to the peripheral nervous system or some characteristic that favors accumulation of the agent within the peripheral nervous system.
Vulnerabilities of Axons to Toxicants
The need to rapidly conduct impulses over relatively great distances presents the nervous system with considerable challenges. The typical cell shape is elongated with a narrow efferent axonal appendage extending from the neuronal perikaryon that in humans may be a meter in length and in larger species even longer. The cylindrical axon extension enables the cell to carry action potentials over distance while minimizing the cytoplasmic volume and cell surface area. However, despite the small axon diameter, the axoplasmic volume can be hundreds to thousands of times greater than the cytoplasmic volume within the neuronal cell body due to the axon length. Because the perikaryon is a major source of biosynthesis for components of the axon, there is an essential requirement to distribute these biomolecules that include neurotransmitters, proteins, and organelles along the length of the axon via axonal transport. 62,63
Transport of cargo synthesized within the neuron proceeds along microtubules mediated by the ATP-dependent microtubule motors kinesin and dynein (Figure 1). 63 These molecules provide mechanochemical force through physical interaction with microtubules and their ATPase activity. Depending on the cargo transported, the observed overall rates of movement vary. The fastest component moves about 400 mm/d and is primarily carrying vesicles. An intermediate component of transport moving 50 mm/d is associated with organelles. The slowest component transports cytoskeletal elements and is usually divided into slow component a (SCa) and slow component b (SCb). The SCa moves at a rate of 1 mm/d carrying microtubules and neurofilaments, with SCb proceeds at a rate of 2 to 4 mm/d carrying microfilaments and microfilament-associated proteins.
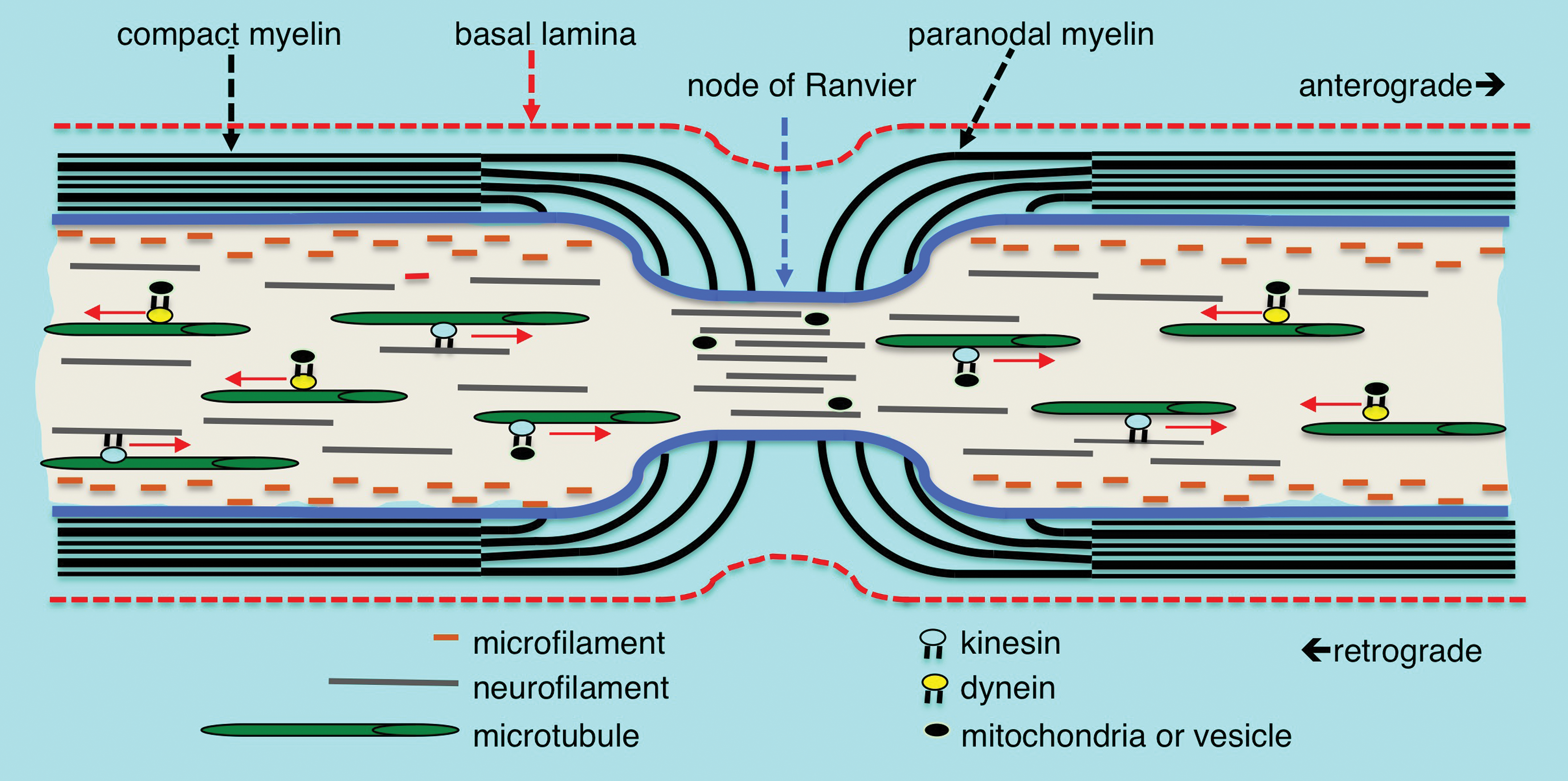
Myelin structure, cytoskeleton arrangement, and axonal transport at the node of Ranvier. Microfilaments of polymeric actin align along the axolemma, neurofilaments are distributed throughout the axoplasm and microtubules are arranged more centrally in the axoplasm. Cytoskeletal components, vesicles, and mitochondria are transported anterograde and retrograde along microtubule tracts by the mechano-chemical motors kinesin and dynein, respectively. Compact myelin opens into cytoplasmic loops in the paranodal region with microvilli projections contacting the axolemma. There is a rapid decrease in the axonal diameter at the node of Ranvier accompanied by an increased density of neurofilaments having a decreased state of phosphorylation. There is also an increased density of vesicles and other organelles within the axon at the node suggesting an impediment to transport at the nodal constriction.
Anterograde and retrograde transport are both essential, with the former providing a vital supply line of biosynthetic components and the latter thought to inform neurons on the status of the distal axon. As demonstrated by experiments, transecting or ligating axons stopping transport results in degeneration of the distal axon. Integral to maintaining normal structure and transport functions are the filamentous proteins of the axonal cytoskeleton. 62 These proteins comprise 3 families categorized based upon their size. The smallest microfilaments of assembled actin filaments are located along the periphery of the axoplasm juxtaposed to the axolemma. The intermediate filament network composed of neurofilament protein polymers extends throughout the axoplasm and plays a role in maintaining axon diameter modulated by associated Schwann cells. Microtubules composed of assembled α/β tubulin dimers arranged in polarized linear arrays more centrally located within the axoplasm are the largest cytoskeleton component. The microtubules play an essential role through providing structural tracts for anterograde and retrograde axonal transport. These cytoskeletal components are all dynamic structures that involve controlled assembly and disassembly to achieve their own transport along the axon and perform their structural functions. Considering that the slowest components migrate on the order of 1 mm/d and that axons can be over a meter in length provides a considerable period of time for cumulative events such as covalent modifications to accumulate on cytoskeletal proteins. Thus, collectively, the structural components and physiological processes required for axonal transport present unique targets that can account for the ability of some neurotoxicants to produce a primary axonopathy. Chemotherapeutic agents that compromise the dynamics of tubulin elongation and disassembly to disrupt mitotic spindle formation in cancer cells as well as certain environmental chemicals that covalently modify cytoskeletal proteins in a cumulative manner are examples of axonal toxicants that act through this mechanism. 62,64,65
Toxic Disorders of Axons
Axonal degeneration is the most commonly observed lesion in toxic peripheral neuropathies. This lesion can be classified as either primary or secondary depending on whether the critical effect is within the axon or another component of the peripheral nervous system, respectively. Using transected nerves in frogs, Augustus Waller described a sequence of structural changes in the degenerating axon segment distal to the transection. 66 Accordingly, the sequence of events he reported is referred to as Wallerian degeneration. Because the structural changes observed in degenerating axons of toxic neuropathies appear to occur through a similar sequence of events, it is often referred to as Wallerian-like axonal degeneration.
Degenerating axons can present in a variety of morphologic profiles depending on their stage of degeneration (Figure 2). In primary axonal degeneration, the initial injury occurs within the axon but is accompanied by changes in the associated glial cells and blood–nerve barrier that collectively provide a microenvironment supportive of axonal regeneration. Typically, the first observable structural change is proteolytic digestion of the axolemma and axoplasm leaving only a myelin sheath surrounding a swollen degenerate axon (Figure 2B). At the light microscopic level, this is identifiable as a loss of protein staining within the axoplasm. Axon degeneration proceeds through a programmed pathway of signaling leading to cytoskeletal degeneration. Studies stemming from the discovery of Wallerian Degeneration Slow (Wlds) mice that have a mutation in the enzyme nicotinamide mononucleotide adenylyltransferase 1 (NMNAT) have provided insight into some of the events triggering degeneration. Loss of function of NMNAT2, but not either of the other 2 isoforms of NMNAT, produces axonal degeneration. 67,68 A constant supply of NMNAT2 is required in the axon through axonal transport to prevent degeneration. Therefore, disruption of axonal NMNAT2 supply through blocked transport, increased proteosomal breakdown, or decreased translation caused by a toxicant is expected to initiate degeneration. Studies have also identified sterile α and TIR motif-containing 1 protein (SARM1) as a pro-regulator of degeneration that acts downstream of NMNAT2, and depleting SARM1 in animals that have loss of NMNAT2 function prevents degeneration. 68 –70 An important consequence of increased SARM1 activity or loss of NMNAT2 function is the activation of dual-leucine-zipper-bearing kinases/mitogen-activated protein kinases (MAPKs) signaling that leads to calpain-mediated proteolysis. 71,72 Maintenance of myelin-axon contact is essential to supporting intercellular signals across the periaxolemmal space, and if the axon is injured or destroyed, the Schwann cell ceases to support the myelin sheath. 73,74 The earliest changes in Schwann cells occur in noncompact myelin regions and are present prior to axonal changes observable at the light microscopic level (Figure 3). These morphological changes include paranodal retraction and dilatation of Schmidt-Lantern incisures. 75 –80 At the molecular level, increased expression of lipolytic enzymes such as phospholipase A2 and demyelinating pathways including the NRG1-erbB2 pathway are activated. 81 –84 The dilatation of Schmidt-Lanterman incisures results in elevated hydrostatic pressure that displaces compact myelin to form intracellular myelin ovoids (Figure 2C). Studies suggest that actin polymerization and recycling of E-cadherin contribute to these changes in Schmidt-Lanterman clefts and paranodal regions. 81 Accompanying the formation of myelin ovoids, Schwann cells become hypertrophic, filling with rough endoplasmic reticulum and various vesicular structures and disintegrating compact myelin (Figure 4A).
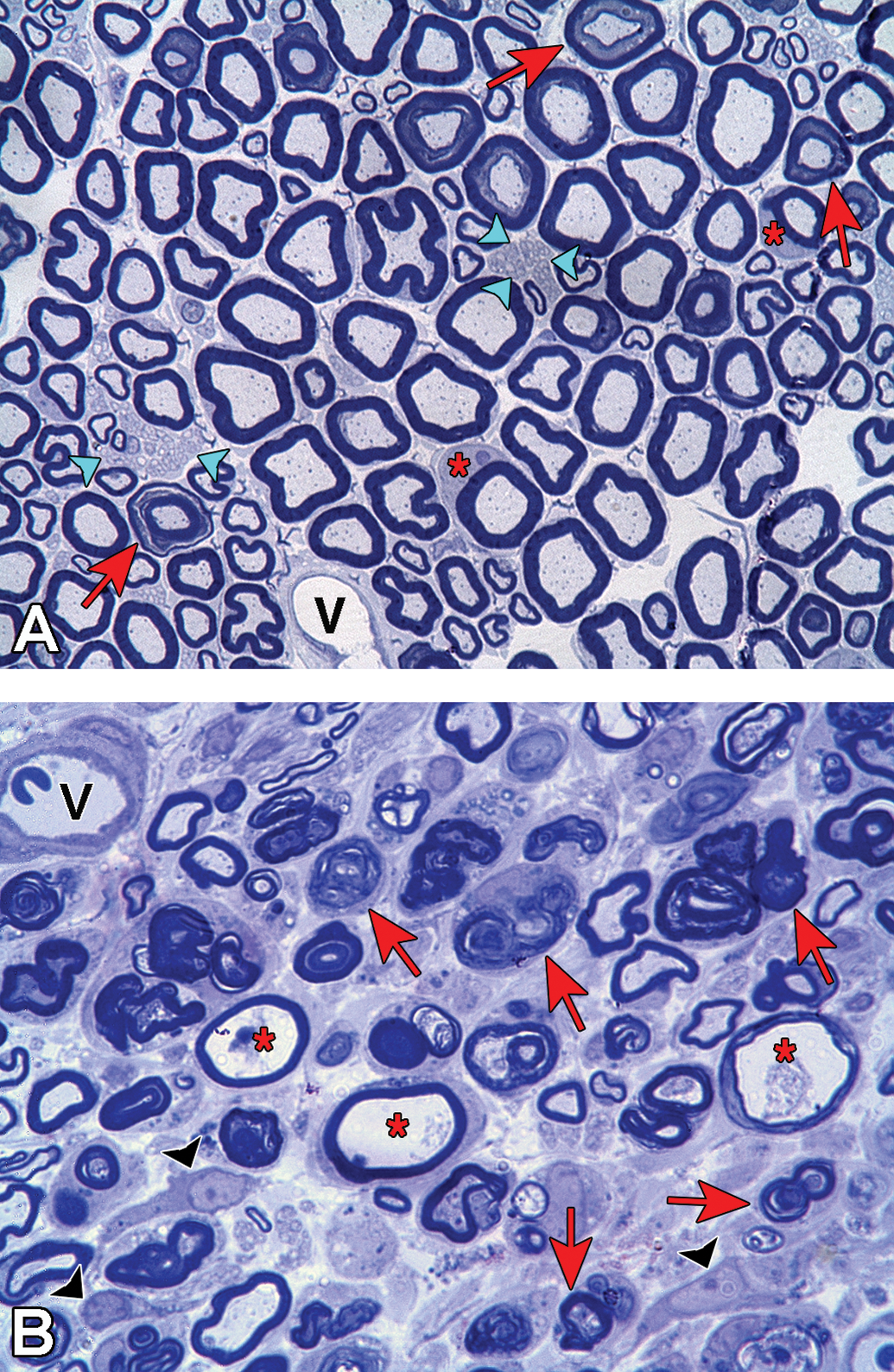
Toluidine blue stained plastic embedded sections of rat sciatic nerve showing stages of axonal degeneration after crush injury. A, Cross section from a control rat showing the normal axon density and bimodal distribution of large and small myelinated axons. The myelinated axons are surrounded by compact myelin and protein staining of the cytoskeletal and mitochondrial components in the axoplasm is evident. Two Schwann cells (*) have been sectioned at the middle of the internode through their cytoplasm and nucleus and axons sectioned through Schmidt-Lanterman incisures are present (arrows). Clusters of nonmyelinated axons (arrowheads) are present and a blood vessel (v) is located along the lower boarder of the section. B, Cross section obtained distal to crush injury. There is a decreased density of axons in the section with very few axons exhibiting normal axoplasm staining. Axons in early stages of axonal degeneration (*) are identifiable by the loss of axoplasm and axolemma staining with the myelin sheath still intact. Many axons are in later stages of axonal degeneration (arrows) demonstrating the progression of myelin collapse to form ovoids. Nucleated cells (arrowheads) present are consistent with dedifferentiated Schwann cells, macrophages, or fibroblasts. A blood vessel (v) containing a red blood cell is located in the upper left corner.
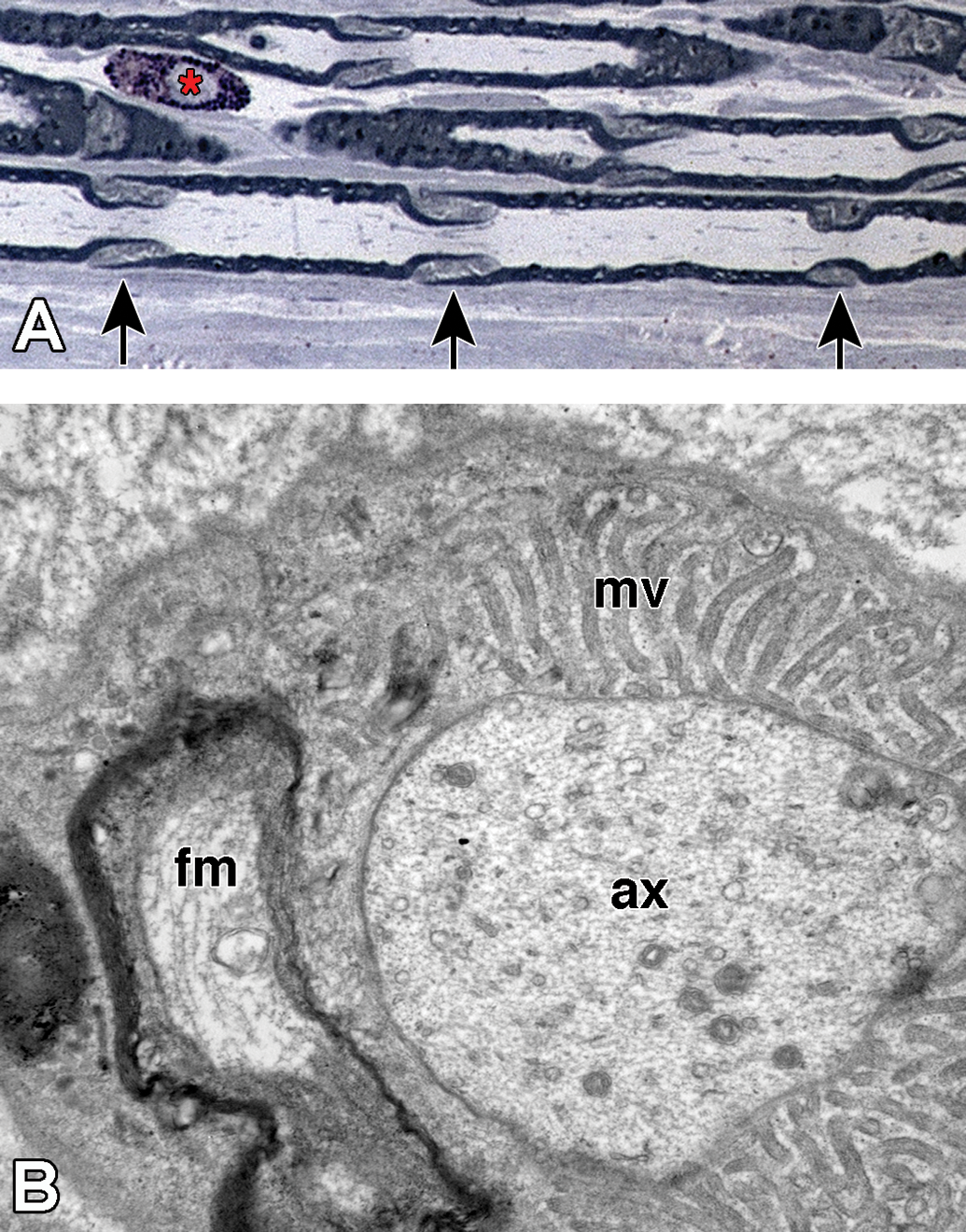
Regions of noncompact myelin that are affected earliest during axonal degeneration. A, Longitudinal section of plastic-embedded toluidine blue-stained rat sciatic nerve showing Schmidt-Lanterman incisures (arrows) distributed along the internode. These structures consist of noncompact myelin that provide a cytoplasmic pathway connecting the Schwann cell cytoplasm adjacent to the axon to that on the external myelin sheath. A mast cell (*) is present in the endoneurium. B, Transverse section through the paranodal region of a large myelinated fiber. The axon (ax) is constricted and contains a high density of neuroflaments and organelles. The compact myelin has opened to form folded myelin (fm) and microvilli (mv) that contact the axolemma.
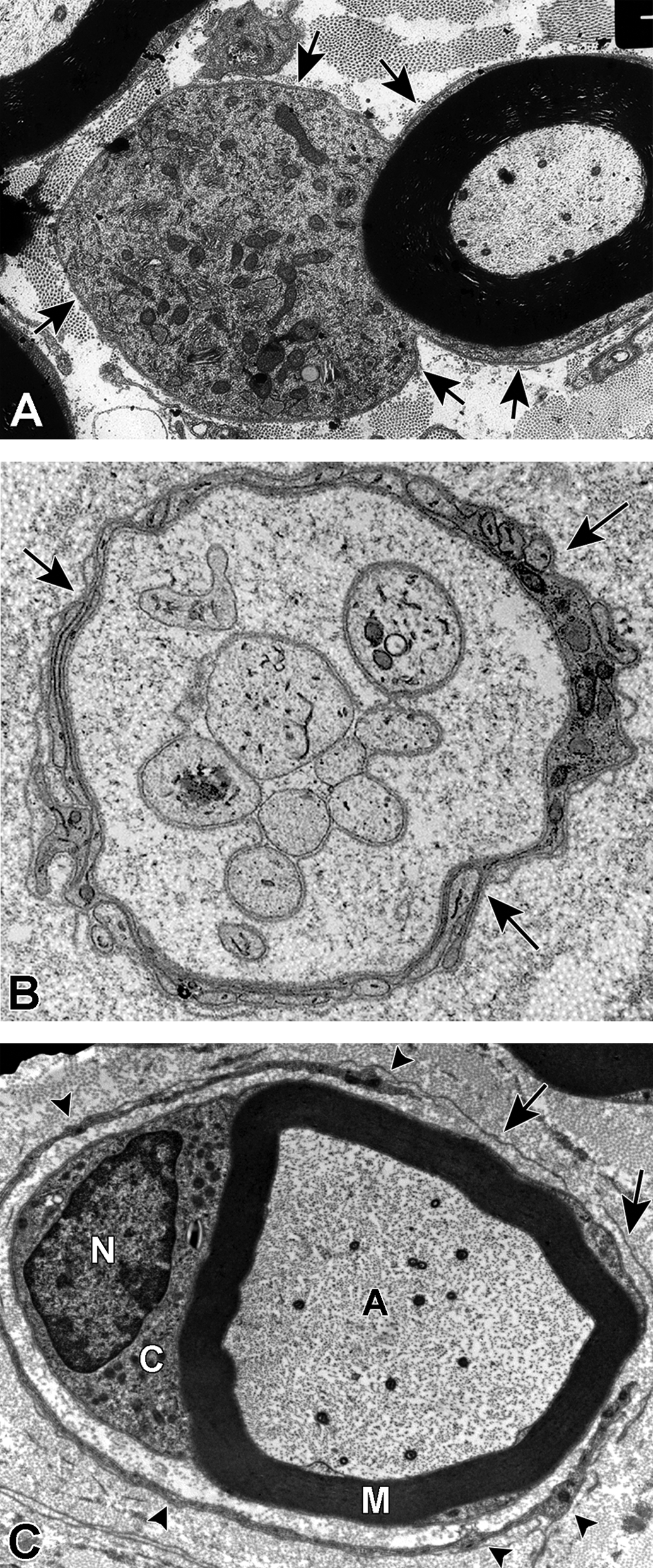
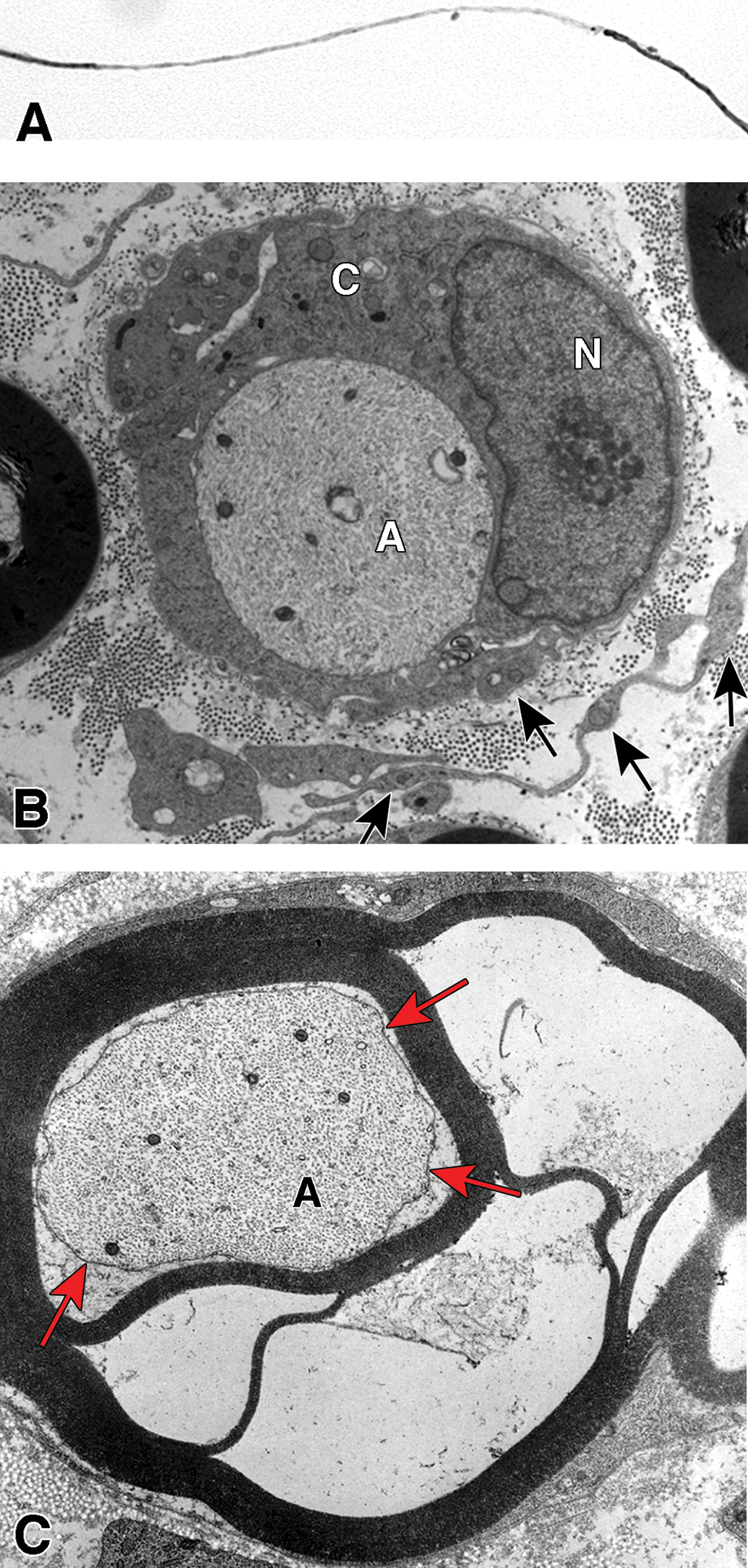
Examples of degenerative and regenerative responses of Schwann cells and axons. A, Schwann cell hyperplasia exhibiting an increased cytoplasmic volume, enclosed by the extension of basal lamina (arrows), that is filled with rough endoplasmic reticulum and various vesicular structures from disrupted membranous organelles. B, A cluster of regenerating axonal sprouts that have not yet been myelinated surrounded by persistent basal lamina from previous Schwann cells and the processes of supernumerary Schwann cells (arrows). C, A regenerated axon (A) ensheathed with thin myelin (M) from its current Schwann cell sectioned through the cell body showing the nucleus (N) and cytoplasm (C) that is surrounded by persistent basal lamina (arrows) of previous Schwann cells and the processes of supernumerary Schwann cells (arrowheads).
Secondary demyelination accompanying Wallerian-like degeneration is not a result of Schwann cell death by necrosis or apoptosis but an active process mediated through reprogramming of myelinating Schwann cells to a demyelinating phenotype. At the molecular level, this reprogramming likely recapitulates the events of primary demyelination once the triggering signal from the axon has been transduced. Schwann cells downregulate lipid and myelin-related proteins and reexpress genes that were active in pro-myelinating or previous immature states. However, the demyelinating Schwann cell is not just a copy of an immature state and expresses some different genes including sonic hedgehog (SHH) and oligocyte transcription factor 1. 85,86 A recognized event in the reprogramming is activation of c-Jun; downregulating c-Jun delays demyelination after injury and prolongs regeneration. 85,87,88 Upregulation of c-Jun appears to be a key factor in reprogramming as it upregulates genes related to neuronal growth and regeneration including N-cadherin, p75, and neuronal cell adhesion molecule and the signaling molecules glial cell-derived neurotrophic factor (GDNF), artemin, SHH, and brain-derived neurotrophic factor (BDNF), while it downregulates myelin-related genes including early growth response protein 2 (Egr2), P0, MBP, periaxin, and E-cadherin. 85,87,89
There are a number of possible ways Schwann cells can be signaled into the demyelination program, and in severed nerves, the process starts rapidly within the first hour. However, the exact molecular events responsible for initiating the process are not known. One hypothesis is that erbB1/2 receptors that have been observed to become phosphorylated within minutes of injury contribute to the triggering process. 82 Phosphorylation of erbB receptors occurs earliest at Schwann cell microvilli located at the nodes of Ranvier and is then propagated along the plasma membrane. This modification activates Ras-related C3 botulinum substrate GTPase regulation of actin polymerization, an early event in Schmidt-Lanterman incisure collapse and paranodal myelin retraction. Interestingly, Mycobacterium leprae that binds to the α-dystroglycan receptor of laminin 2 of Schwann cells produces demyelination and has been observed to activate erbB2. 90 This would suggest that the erbB receptors that are activated by NRG1 for proliferation, survival, and myelination could also play a role in demyelination.
Removal of myelin debris is more efficient in the peripheral nervous system than the central nervous system and is a concerted effort between demyelinating Schwann cells and macrophages. 91,92 Schwann cells appear capable of digesting some of their own myelin sheath, but a substantial fraction of the degrading myelin along with parts of the degrading axon is also ejected by the Schwann cell. The ejected myelin is then cleared by macrophages in part by resident macrophages present within the endoneurium and by recruited circulating hematogenous macrophages. Both Schwann cells and macrophages are able to phagocytose extracellular myelin, but in addition, Schwann cells appear to clear their own intracellular myelin debris through autophagy based upon the presence of autophagosome-like structures and high levels of lysozyme in hypertrophic Schwann cells. 93
Additionally, Schwann cells dedifferentiate to a premyelinating mitotic phenotype that align along the basal lamina that continuously encircles the outer Schwann cell plasma membrane over the entire length of each axon including nodes of Ranvier in the peripheral nervous system to form tubular structures referred to as bands of Büngner (Figure 4B). 94,95 Through providing structural guidance and trophic support by expressing several growth factors such as artemin, BDNF and GDNF for the neurites sprouting from the proximal stump these bands enhance the regenerating potential of axons in peripheral nerve. 89,96 –99 Specialized terminal nonmyelinating Schwann cells also guide axon terminals to the neuromuscular junction. 100 Once axon regeneration is complete, the repair phase Schwann cells stop dividing and differentiate into either the myelinating or nonmyelinating phenotype Schwann cells. The telling signs of remyelination are shorter internodes and a thinner myelin sheath (Figure 4C). 101 Typically, the ratio of internode length (L) to myelin outer diameter (D) is ∼100 and the ratio of the inner myelin diameter (d) measured between nodes of Ranvier to the outer myelin diameter (D) referred to as the g-ratio is 0.7 to 0.8. 102 –104 Thus, when L/D is less than 100 and the g-ratio greater than 0.8, it is consistent with remyelination. In contrast, axons within the central nervous system do not have an associated basal lamina, proliferating astrocytes form glial scars, and the presence of persistent myelin debris all inhibit neurite outgrowth diminishing the potential of axon regeneration. However, even in peripheral nerve, if axon/Schwann cell contact is not reestablished, eventually the Schwann cells and bands of Büngner will diminish and be replaced with collagen decreasing the probability of successful regeneration.
Secondary axonal degeneration is either a consequence of neuronal perikaryon death or persistent primary demyelination. In the case of neuronal cell death, the associated dendrites and axonal cellular processes are lost without the potential of regeneration. In the case of primary demyelination, the axonal response varies depending on the type and duration of the myelin injury. During incomplete demyelinating conditions such as intramyelinic edema or early stages of complete demyelinating conditions, axonal atrophy is a common finding whereas ongoing complete demyelination can progress to secondary axonal degeneration.
Another type of toxic axonopathy are the neurofilamentous axonopathies that involve accumulations of neurofilaments at various distances from the cell body and in some cases within the perikaryon (Figure 5). 64 Neurofilaments are type IV intermediate filaments that are heteropolymers of noncovalently associated neurofilament protein subunits. 62 The subunits include 3 neurofilament proteins termed low-, medium-, and high-molecular-weight neurofilament protein based on their rates of migration in gel electrophoresis and lengths of their carboxy terminus. Additionally, the filamentous structures also incorporate α internexin and peripherin as additional subunits. The neurofilament component of the cytoskeleton is thought to regulate axon diameter and thus plays a role in conduction speed. Schwann cells locally modulate levels of neurofilament phosphorylation providing a potential link between loss of Schwann cell contact and axonal atrophy. 105 –107 A number of chemicals have been identified through human exposures or experimental animals that produce significant alterations in neurofilament distribution that are accompanied by varying levels of axonal degeneration. These chemicals include carbon disulfide, hexane, methyl-n-butyl ketone, acrylamide, and 3,3′-iminodiproprionitrile, among others.
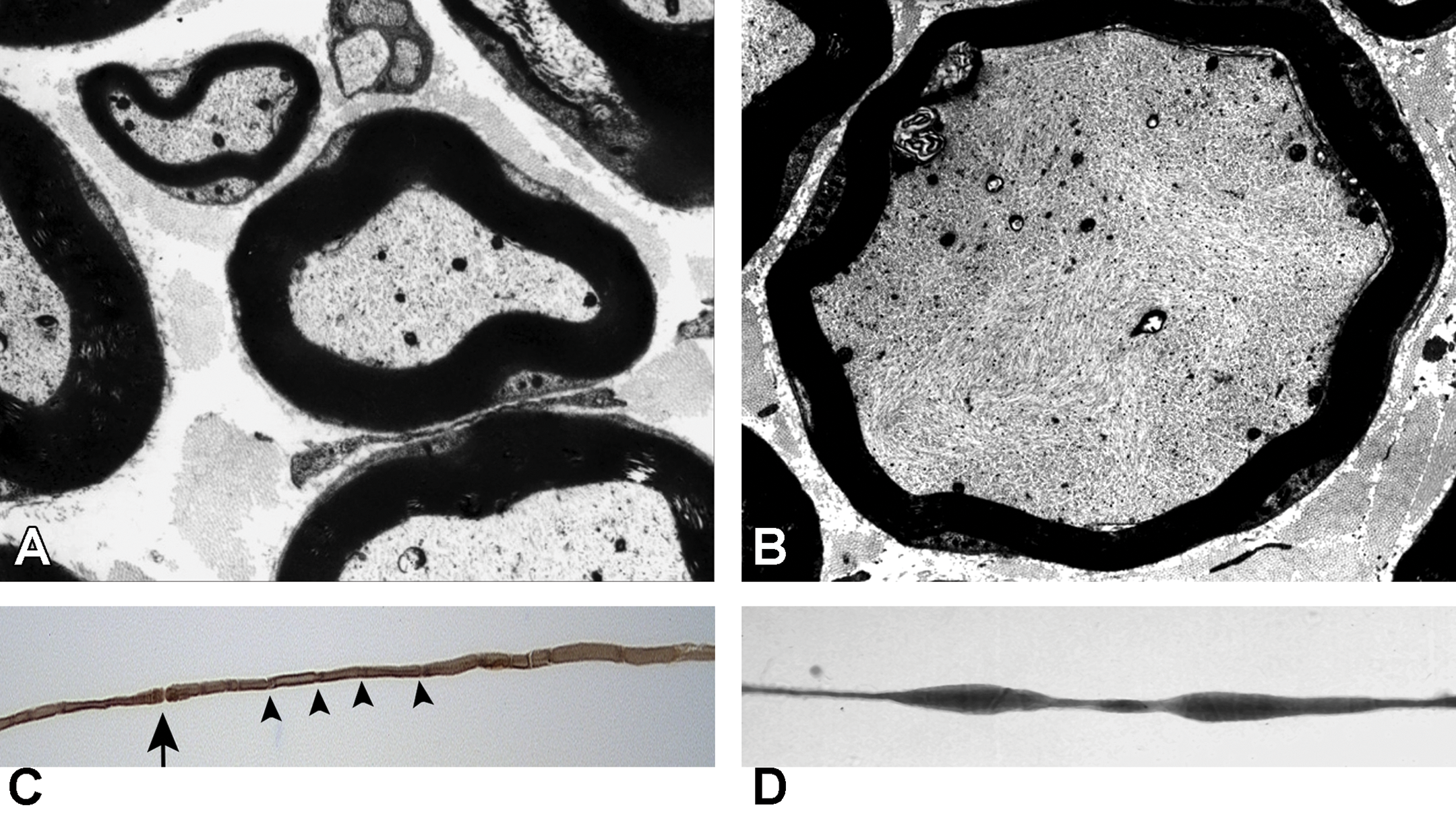
Neurofilamentous axonopathy produced by carbon disulfide released from N,N-diethyldithiocarbamate following oral administration in the muscular branch of the posterior tibial nerve of a rat. A, Transverse section of large and small myelinated axons obtained from a control animal showing the normal thickness of compact myelin relative to axon diameter and the density and distribution of cytoskeletal components. B, Transverse section of a myelinated axon present in a treated animal. There is a high density of disorganized neurofilaments in the swollen axon compared to the orderly distribution and alignment in the normal axon. Additionally the myelin is thin and there is sequestering of organelles to the periphery. C, Teased fiber from a control animal showing a constant diameter and even myelin staining along the internode. A node of Ranvier (arrow) is present and Schmidt-Lanterman incisures (arrowheads) are distributed along the axon. D, Teased fiber from a treated animal showing the presence of fusiform swellings.
Examples of Agents Producing Peripheral Axonopathies
Taxanes
Paclitaxel is an extract from the Pacific yew tree (Taxus brevifolia) and docetaxel is a semisynthetic analog of paclitaxel. Both are used as chemotherapeutic agents for several cancers including breast, ovarian, and lung cancers. Paclitaxel is more neurotoxic, but both agents can produce dose-dependent neuropathies. 4,22,65 The sensory system is more sensitive, but motor involvement can also occur. Clinical signs include numbness, neuropathic pain, and cold and mechanical allodynia. The neuropathy is characterized by axonal degeneration with secondary demyelination. An early change is disrupted growth at the distal tips of sensory axons that require ongoing growth to maintain innervation of the epidermis as it turns over.
These compounds cross plasma membranes and bind to the N-terminal region of β-tubulin. This leads to enhanced polymerization and decreased depolymerization. 108,109 Integrity of microtubule dynamics is required for axonal transport, and disturbance of microtubule function is thought to interrupt transport of essential cellular components. 110 More recently, the observation that paclitaxel decreased levels of some axonal mRNAs but fast transport of organelles was not disrupted suggests this compound may have selective effects on components of transport. 111,112 Additional contributing mechanisms that have been proposed include interacting with mitochondria to open the mitochondrial permeability transition pore and activation of calpains. 113,114
Vinca Alkaloids
Vincristine and the related vinca alkaloids vinblastine, vinflunine, and vinorelbine are important chemotherapeutics used primarily for hematological cancers. Vincristine is the most neurotoxic and has a high prevalence of peripheral neuropathy associated with treatment. 22,65 Neurotoxicity is a common reason to stop treatment; clinical signs may worsen for a period after treatment is stopped. Clinically, sensory signs develop earliest but are often followed by motor and autonomic involvement.
These compounds bind to free tubulin causing the free tubulin to aggregate into nonfunctional polymers, which decreases the free tubulin available to be incorporated into microtubules. 115 Similar to taxanes, the disturbance of microtubule function is thought to impair normal axonal transport function. 23,110 Vincristine has also been reported to interact with mitochondrial membrane permeability to decrease the uptake and efflux of Ca2+, potentially disturbing intracellular signaling. 114
Hexane and Methyl-n-Butyl Ketone (2-Hexanone)
The neurotoxicity of these compounds was recognized by the development of a sensorimotor distal axonopathy in furniture and leather workers and from intentional repeated inhalation of hexane-containing glues. 116 –119 The ability to reproduce the neuropathy in experimental animals has provided considerable insight into the lesions and molecular mechanisms contributing to the neuropathy. 120 Morphologically, the neuropathy is characterized by a central peripheral distal neurofilamentous axonopathy. The distal parts of the largest diameter longest myelinated axons are most susceptible and this is reflected in the relative sensitivity of species based on axon size and length. Aggregates of neurofilament protein accumulate in the subterminal axon creating axonal swellings accompanied by myelin retraction that often occurs just proximal to the axonal constrictions at nodes of Ranvier. Electron micrographs of these swellings are consistent with an interruption of axonal transport with neurofilamentous aggregates proximal to and accumulation of vesicles distal to the nodal constrictions. With continued exposure, there is development of axonal atrophy followed by axonal degeneration distal to the swellings.
Hexane is probably one of the most studied toxic neuropathies and there is considerable knowledge regarding the necessary chemical properties of the toxic species and their reactions with proteins. However, there is still debate regarding the most relevant molecular targets and biological processes responsible for axonal degradation.
Investigations in several laboratories have helped to understand the neurotoxic mechanism, and based on these studies, the neurotoxic conditions are referred to as γ-diketone axonopathies. 64,121 –123 Both hexane and methyl-n-butyl ketone are metabolized through ω-1 oxidation to the same neurotoxic species, 2,5-hexanedione, possessing γ spacing of the ketone functions. The requirement for this spacing is well established and has been supported more recently through the use of aromatic analogs such as 1,2-diacetylbenzene. 124,125 The γ-diketones are able to react with amino groups of protein lysyl residues to generate 2,5-dimethylpyrrole adducts (Figure 6A). That a subsequent step involving oxidation of the pyrrole adduct to an electrophilic intermediate, that reacts with a protein nucleophile to generate protein cross-links has been supported experimentally. The 3,4-dimethyl analog of 2,5-hexanedione that forms pyrroles and cross-links more rapidly than 2,5-hexanedione was observed to be a more potent neurotoxicant that produced swellings in more proximal locations. 126,127 In contrast, the 3-acetyl analog of 2,5-hexanedione that forms pyrroles rapidly but does not undergo oxidation and cross-linking did not produce axonal swellings or clinical signs of neurotoxicity. 128 Considering their involvement in the axonal swellings, high content of lysyl residues, and long biological life time for cumulative derivatization, the neurofilament proteins have been proposed as a relevant molecular target. One hypothesis proposes that cross-linking of the neurofilament subunits limits their assembly and disassembly required for transport, particularly at the axonal constrictions occurring at nodes of Ranvier, leading to their accumulation and a blockade of transport. 122 However, whether neurofilament accumulation is a critical event to subsequent axonal degeneration has not been clearly established. Studies using crayfish and transgenic mice that did not express neurofilament proteins have exhibited neurotoxicity from 2,5-hexanedione, suggesting the involvement of additional mechanisms and that neurofilaments are not a requirement for axonal degeneration. 129,130 More recent studies have characterized changes in the transport motors kinesin and dynein and certain microtubule-associated proteins as potential targets contributing to γ-diketone neuropathy. 131
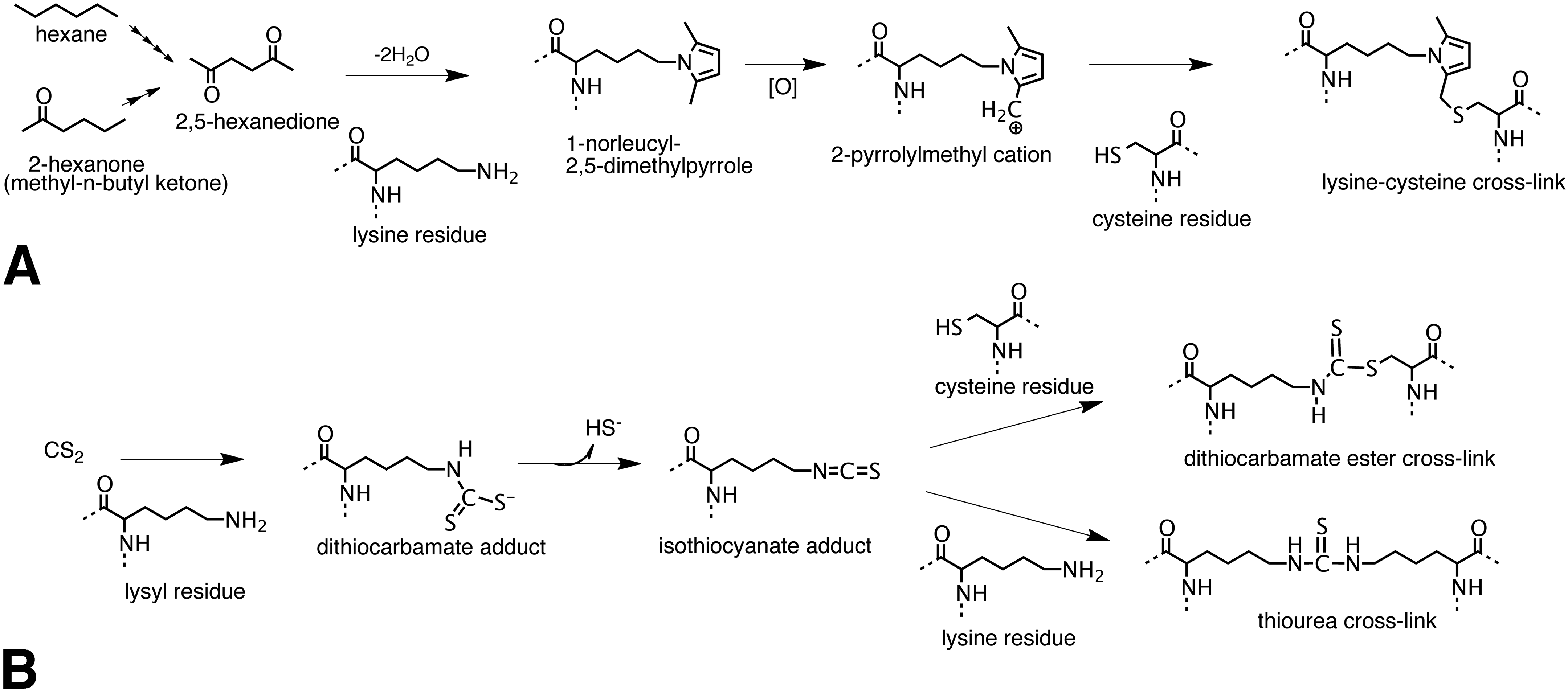
Protein cross-linking pathways for hexane, methyl-n-butyl ketone, and carbon disulfide (CS2). A, Hexane and methyl-n-butyl ketone are metabolized to 2,5-hexanedione through multiple steps of ω-1 oxidation. 2,5-Hexanedione reacts with protein lysyl residues to generate a 2,5-dimethylpyrrole adduct. In the presence of oxygen the pyrrole adduct oxidizes to a cationic species susceptible to nucleophilic addition by protein amino and sulfhydryl groups producing protein cross-links. Reaction of the oxidized pyrrole adduct with a cysteine residue to produce a possible lysine-cysteine cross-link is shown. B, Carbon disulfide reacts with protein lysyl residues to form a monoalkyldithiocarbamate adduct. The dithiocarbamate adduct undergoes facile elimination of sulfhydryl ion creating an electrophilic isothiocyanate adduct. The isothiocyanate adduct then undergoes nucleophilic addition by protein sulfhydryl or amino groups to generate dithiocarbamate ester or thiourea protein cross-links.
Carbon Disulfide
The neurotoxicity of this chemical was first recognized during the 19th century associated with the cold vulcanization of rubber in cottage industries and exposure to carbon disulfide is currently a concern in rayon and cellophane production. 132 Human cases and experimental models have established that carbon disulfide produces a sensorimotor neurofilamentous distal axonopathy identical to that of hexane (Figure 5). 133
The clinical similarities in these 2 neuropathies guided mechanistic studies to determine whether a parallel series of protein adduction and cross-linking steps contributed to carbon disulfide neurotoxicity. Using in vitro and in vivo systems, it was demonstrated that carbon disulfide can react with amino groups of lysyl residues to generate a monoalkyl dithiocarbamate adduct (Figure 6B). 134 –136 The dithiocarbamate adducts undergo a facile elimination of sulfhydryl ion to generate electrophilic isothiocyanate adducts followed by nucleophilic addition of lysyl or sulfhydryl functions to produce thiourea or dithiocarbamate ester cross-links, respectively. Similar to hexane, it has been postulated that the stability and long transit time of neurofilament proteins make them the toxicologically relevant target for the development of neurofilamentous swellings in chronic carbon disulfide neuropathy. 64,122
1-Bromopropane
Efforts to decrease the production of ozone-depleting solvents such as chlorofluorocarbons has led to the use of new chemicals including 1-bromopropane. Currently, 1-bromopropane is used as a solvent vehicle for spray adhesives and for cleaning and degreasing of metal components, electronics, optical instruments, and ceramics. 137 Clinical reports of human cases and animal studies have demonstrated the neurotoxicity of 1-bromopropane. 6,8,138 –141 The clinical signs reported included muscle weakness, paresthesias, numbness, urinary incontinence, and memory disturbances. Inhalation studies in rats demonstrated a dose-dependent neurotoxicity characterized by decreased forelimb and hind limb grip strengths, decreased motor nerve conduction velocity, increased distal latency, axonal degeneration in the muscular branch of the posterior tibial nerve, and axonal swellings in the gracile nucleus. 142 The mechanism of 1-bromopropane neurotoxicity has not been delineated, but biochemical studies have provided information on the interaction of 1-bromoproane in biological systems. 1-Bromopropane produces S-propyl cysteine adducts on globin in humans and rats and this protein modification has also been identified within the nervous system on neurofilaments of rats exposed to this chemical. 143 This alkylating effect could explain the low levels of protein sulfhydryl functions observed in the brains of rats exposed to 1-bromopropane and suggests that similar to the γ-diketones covalent protein modification may be a contributing process to neurotoxicity. 144,145 Further investigations will be needed to clarify the role of sulfhydryl modifications to the mechanism of 1-bromopropane.
Vulnerabilities of Schwann Cells and Myelin to Toxicants
Schwann cells ensheath most axons in peripheral nerve. Differentiated Schwann cells can be categorized into several groups based upon their morphology, biochemical composition, and the size and location of the axon or axons they associate with. These categories include the myelinating Schwann cells that each associate with a single large motor and certain sensory axons (Figure 7A), nonmyelinating Schwann cells that each wrap multiple small diameter axons of c-fibers from sensory and postganglionic sympathetic neurons to form Remak bundles (Figure 7B), paranodal Schwann cells whose compact myelin opens to form myelin villi near nodes of Ranvier (Figure 3), perisynaptic Schwann cells that are located distally at neuromuscular junctions where they partially wrap around the presynaptic terminal of motor axons, and the satellite cells that associate with neurons in the peripheral nervous system ganglia (Figure 7C). Our understanding of myelinating Schwann cells and their response to toxicants is the most advanced and will be the primary subject in this discussion.
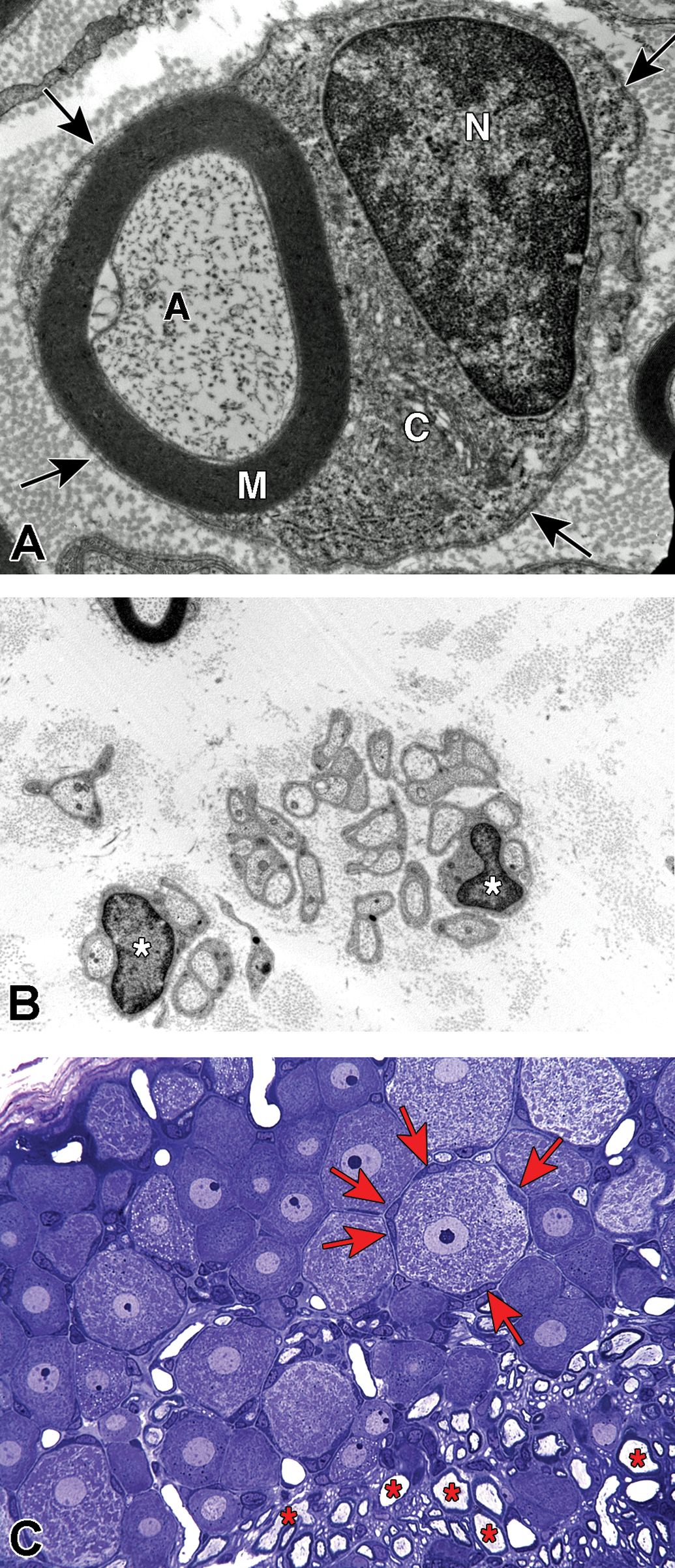
Schwann cell phenotypes. A, Transverse section through the cell body of a myelinating Schwann cell showing the nucleus (N), cytoplasm (C), compact myelin (M), axon (A), and basal lamina (arrows). B, Groups of nonmyelinated axons showing the cell bodies of 2 nonmyelinating Schwann cells (*) with their basal lamina wrapped around multiple axons. C, Plastic-embedded transverse section from a rat dorsal root ganglion showing neurons that have large central nuclei with prominent nucleoli and abundant Nissl substance in the cytoplasm, neurons encircled by multiple satellite cells (arrows) and myelinated axons (*).
Schwann cells associated with larger diameter axons produce myelin that increases conduction velocity and decreases energy requirements for membrane repolarization. 146 They form a lipid-rich wrapping of compact myelin that acts as insulation along axons. This insulation is interrupted at nodes of Ranvier where ion channels are expressed enabling localized depolarization and repolarization that is not perpetuated through the myelin but can depolarize adjacent nodes. The result is saltatory conduction having an increased speed and lower energy requirement than a continuous membrane depolarization along the length of the axon. To perform its insulating function, peripheral nerve myelin is ∼80% lipid as compared to most plasma membranes that are ∼50% lipid. 147 The polyunsaturated fatty acids in the myelin plasma membrane are susceptible targets for lipid peroxidation resulting from agents producing free radicals directly or that increase oxidative stress through other mechanisms. The formation and maintenance of compact myelin is dependent upon a number of membrane-associated proteins and the metabolism of specific lipids present within the lipid bilayers (Figure 8). The MBP is a myelin protein thought to stabilize the closely opposed inner cytoplasmic membranes that form the major dense line. P0 is a major protein component of myelin thought to stabilize the junction of the outer plasma membranes through homophilic interactions to form the intraperiod line of compact myelin. 148 Additionally, in order to maintain the integrity of compact myelin, energy-dependent ion pumps thought to include carbonic anhydrase are required to exclude water from the intraperiod line between the tight wrappings of plasma membrane. 149 The metabolic processes and structural features required for compact myelin provide unique targets for toxic agents, particularly those that are hydrophobic that can cross the blood–nerve barrier and accumulate in myelin.
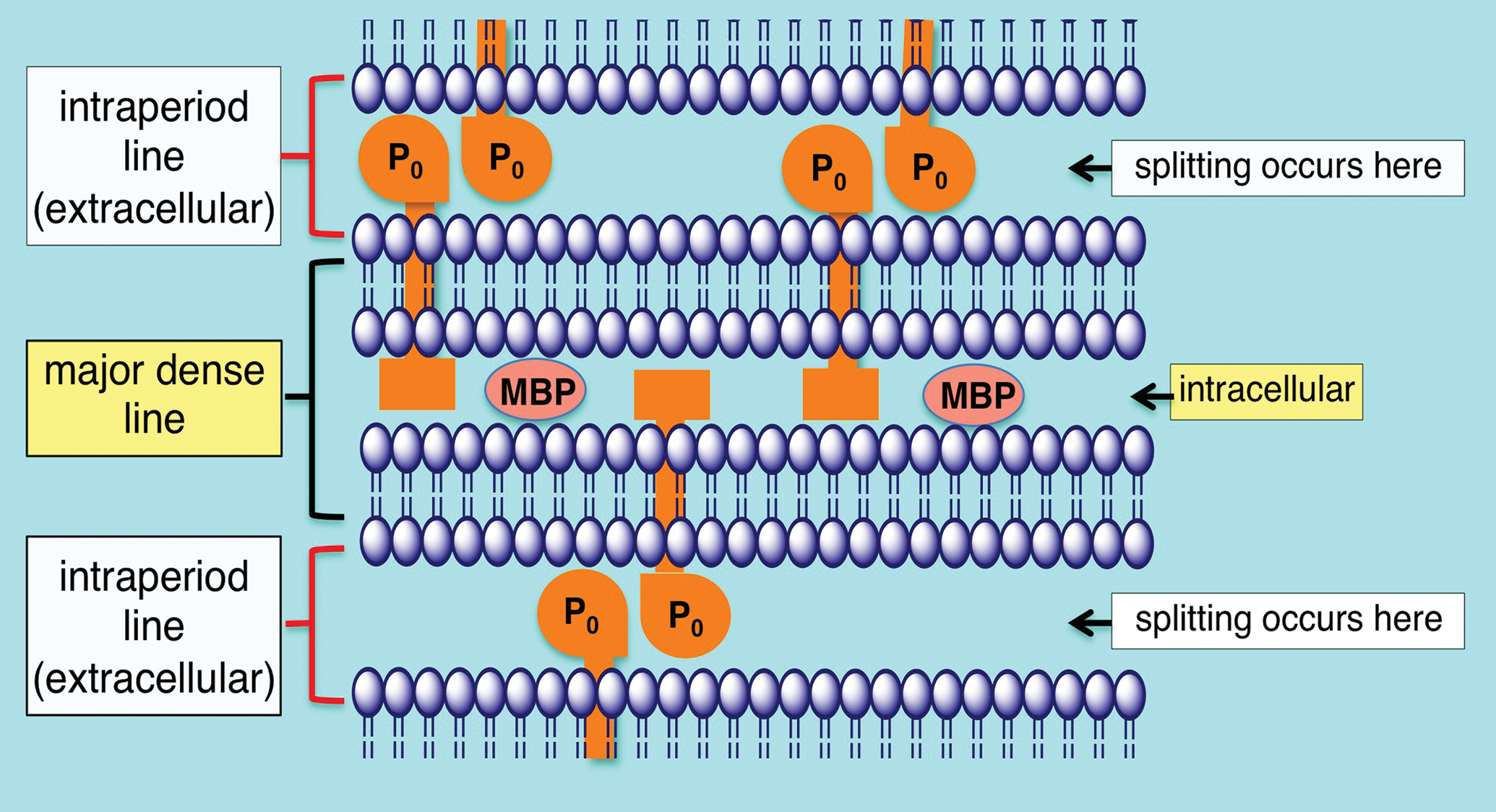
Schematic of peripheral nerve compact myelin. The intraperiod line is formed by the association of the exterior surfaces of the Schwann cell plasma membrane wrapped around the axon. The apposition of the external surfaces is stabilized in part by homophilic interactions of P0 protein. Energy dependent ion pumps thought to include carbonic anhydrase are required to exclude water from the intraperiod line, the typical location that myelin splitting occurs. The major dense line is formed through the exclusion of cytoplasm and close association of the interior surfaces of the Schwann cell plasma membrane. Myelin basic protein (MBP) is important in stabilizing the apposition of the interior plasma membrane surfaces.
Toxic Disorders of Schwann Cells and Myelin
The high lipid content of myelin introduces unique challenges to fixation and embedding. Therefore, when examining the peripheral nervous system for myelin lesions, it is particularly important to discern between lesions and processing artifacts. 150 A common artifact that can be misleading is myelin blebbing that results from inadequate dehydration for embedding in plastic. Experimental treatment-related changes can be distinguished from blebbing artifact by the symmetric distribution of the blebs around the axons and the uniform inclusion of all axons within a section. In contrast, nonartifact myelin lesions exhibit a range of severity among axons within a section and the lesions of affected axons are typically asymmetric. Other common artifacts include those resulting from crush during handling of the nerve and incomplete penetration of osmium used for the lipid fixation.
As described for axonopathies, myelinopathies can be classified as either primary or secondary. Once demyelination is initiated, the morphological changes including paranodal retraction, collapse of Schmidt-Lanterman incisures, lysosome induction, and certain molecular changes are shared suggesting the existence of a common demyelinating phenotype. 73 For the case of secondary myelinopathies resulting from axonopathies, similar stages of response are expected for all of the Schwann cells along the entire length of the affected axon section. In contrast, individual Schwann cells, typically those maintaining the longest internodes and thickest myelin sheaths of the largest axons, are affected initially in primary demyelination. This can result in segmental demyelination and is thought to result from the greater metabolic demands placed upon this population of Schwann cells (Figure 9A, B). Significantly, the loss of a single internode can be sufficient to interrupt saltatory conduction.
Segmental demyelination and intramyelinic edema in the sciatic nerve of rats administered disulfiram. A, Teased fiber from an animal exposed to disulfiram showing loss of myelin from an entire internode. An abrupt change to normal myelin thickness is seen on both sides of the demyelinated internode at each node of Ranvier. B, A demyelinated axon (A) surrounded by a Schwann cell sectioned through the cell body showing the nucleus (N) and increased cytoplasmic volume (C) filled with vesicular structures and debris. Processes of supernumerary Schwann cells are present (arrows). C, Several layers of the compact myelin lamella have separated and the space created is filled with fluid (intramyelinic edema). The axon (A) has a relatively normal density and distribution of mitochondria and neurofilaments but the axolemma (arrows) has separated from contact with the Schwann cell plasma membrane at the inner layers of compact myelin.
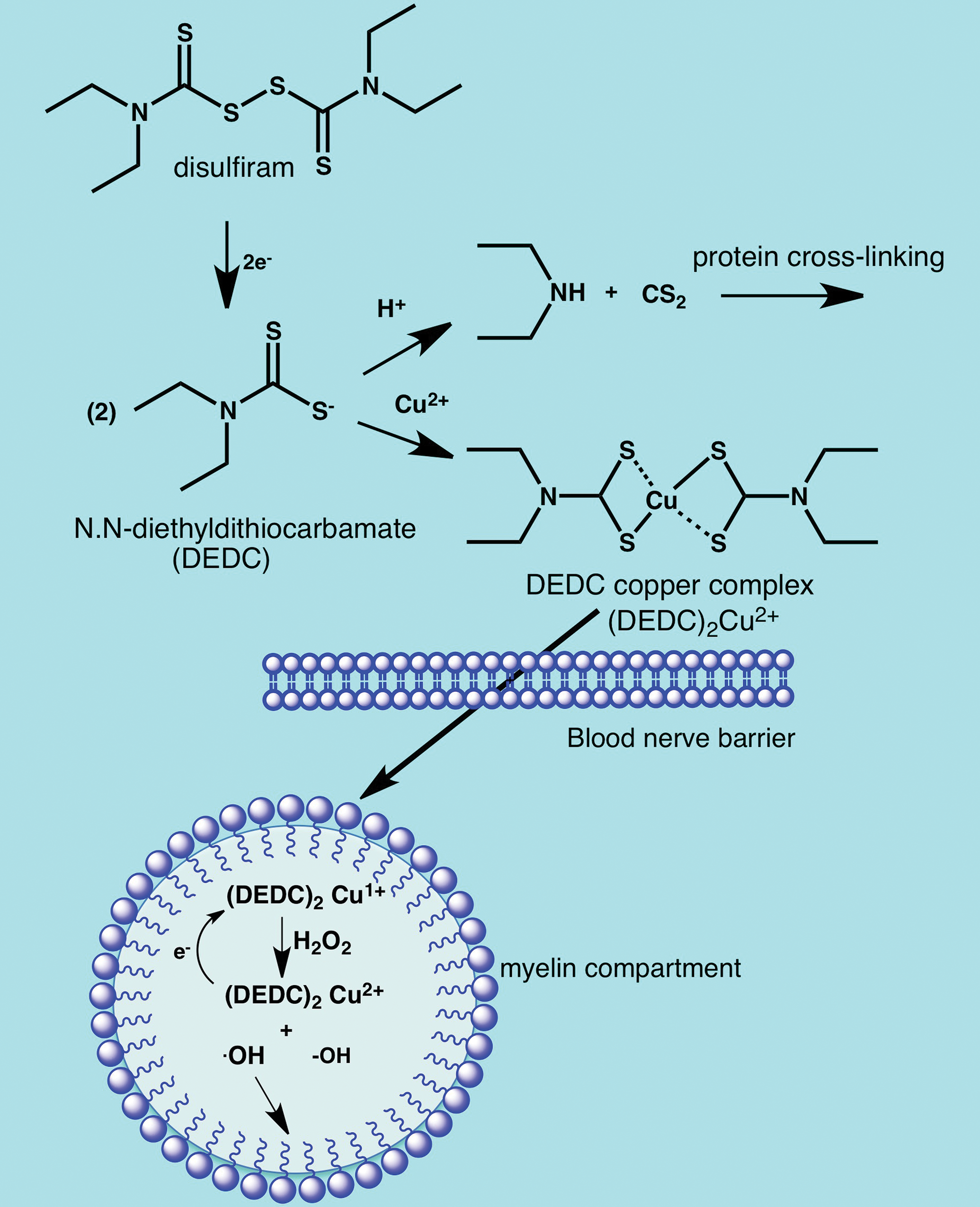
Disposition and mechanisms of neurotoxicity for disulfiram and N,N-diethyldithiocarbamate. Oral administration of N,N-diethyldithiocarbamate (DEDC) results in acid promoted decomposition to CS2 and diethylamine. The CS2 released produces a neurofilamentous axonopathy through protein cross-linking (see Figure 6). Disulfiram is more acid-stable and can be absorbed intact following oral administration and then reduced to DEDC in vivo. The DEDC derived either from oral disulfiram or parenteral administration of DEDC forms lipophilic redox active complexes with copper that cross the blood–nerve barrier and accumulate in myelin. Redox cycling of the copper complexes in myelin produces free radicals promoting lipid peroxidation that results in demyelination.
Integral to the structural integrity of compact myelin are the interactions of the associated membrane proteins and the energy-dependent processes required to exclude water. Agents that compromise these processes can produce myelin splitting or intramyelinic edema. This lesion is characterized by extracellular separation of the lamella at the intraperiod line formed by the closely opposed outer membrane protein layers of the Schwann cell membrane wrapped around the axon (Figure 9C). This type of splitting can be observed in the inner, middle, or outer lamella of the compact myelin.
Examples of Agents Producing Peripheral Myelinopathies
Hexachlorophene
The use of hexachlorophene in soaps and detergents as an antimicrobial has resulted in neurotoxicity in newborn humans and animals. This compound is hydrophobic, is absorbed through skin, and enters the nervous system where it accumulates in myelin. The greater susceptibility of newborns is thought to result from the incompletely developed blood–brain barrier during this period. A myelinopathy is produced in both the central and peripheral nervous systems that is characterized by splitting of myelin at the intraperiod lines. 151 –154 The largest axons appear to be the most susceptible and the optic nerve is particularly vulnerable. Mechanistic studies have supported the ability of hexachlorophene to uncouple oxidative phosphorylation and inhibit carbonic anhydrase. 155 –157 Both of these effects are thought to interfere with the ability of myelin to exclude water resulting in intramyelinic edema that separates the compact myelin at the intraperiod line.
Triethyltin
Neurotoxicity of triethyltin has been recognized through contamination of medications and as an industrial pollutant. It is most recognized for its central nervous system effects, although at high concentrations, it also affects the peripheral nervous system. 158 Splitting at the intraperiod line with myelin swelling is the characteristic lesion. 159 Triethyltin can concentrate in myelin due to its hydrophobicity and the presence of high-affinity binding sites for this compound on myelin. 160 Investigations have identified several potential mechanisms to contribute to the myelin lesions. Inhibition of oxidative phosphorylation resulting from inhibition of Mg2+-ATPase has been proposed as one mechanism. 161 –163 Triethyltin appears to alter ion transport across membranes including the mitochondrial inner membrane anion channel and may directly allow Cl− entry into the intraperiod line. 164,165 All of these mechanisms could disrupt the active processes required to exclude water or enhance osmotic pressure to draw an increased volume of water into the intraperiod line.
Tellurium
Toxicity in humans is rare, but tellurium provides an experimental model to study primary demyelination. When tellurium is administered to weanling rats, it produces segmental demyelination with the longest axons affected earliest leading to hind limb weakness. 166 –169 Typically, only peripheral nerve is affected and older animals are resistant. The myelin injury results from inhibition of squalene epoxidase, an enzyme involved in cholesterol synthesis. 170,171 The susceptible period coincides with a period of rapid myelination in the young animals and a correspondingly high requirement for cholesterol. 172 Inhibition of cholesterol synthesis in the rapidly growing peripheral nerve results in decreased expression of myelin-related genes including P0 and MBP. 173 The downregulation occurs in all Schwann cells and can lead to apoptotic cell death. 174 Because the exposure to tellurium is systemic, it is interesting that the effects are selective for peripheral nerve myelin. The liver is the major source of peripheral cholesterol and the activity of the hepatic rate-limiting enzyme for cholesterol synthesis, hydroxymethylglutaryl-CoA (HMGCoA) reductase, is upregulated in response to tellurium. As a result, the circulating levels of cholesterol are maintained for most organs but access of the circulating cholesterol to the nervous system is prevented by the blood–nerve barrier. 175 Because Schwann cells do not upregulate HMGCoA reductase, there is a localized deficiency of cholesterol in the most rapidly developing regions of the nervous system.
Diptheria Toxin
Corynebacterium diphtheria is an example of a microbe that produces a toxin targeting Schwann cells. This bacterium colonizes skin and mucous membranes where it produces localized necrosis and affects the upper respiratory tract initially. Necrosis results from the production of an exotoxin by 3 biotypes of this bacteria (mitis, intermedius, and gravis) that can also spread through the circulatory system to affect other organs including the heart and kidneys. 176 The peripheral nervous system can also be affected and the occurrence of polyneuropathy has been reported to be as high as 85% in affected children. 177 The first neurologic signs occurring within a week of infection involve sites nearest the infection such as paralysis of the soft palate and pharynx. Subsequently, there can be involvement of cranial nerves and then development of a motor sensory type of peripheral neuropathy occurring from 1 week to months afterward. The exotoxin is thought to not cross the blood–brain barrier but to gain access at peripheral ganglia. The pathology is characterized by paranodal and segmental demyelination with associated slowed and blocked conduction. Typically, axons are spared, but in severe cases, secondary axonal degeneration may also occur. The toxin is a typical AB type bacterial exotoxin of 62 kDa molecular weight comprised of 2 peptides joined by a disulfide bond. 178 The B segment provides selectivity through binding to the uncleaved precursor form of heparin-binding epidermal growth factor–like growth factor expressed on Schwann cells. 179 The toxin is internalized within an endosome through receptor-mediated endocytosis and then the 2 peptides are separated. 180 The A fragment catalyzes transfer of an adenosine diphosphoribose from NAD to elongation factor-2 that is essential for the translation of proteins. The toxin does not appear to promote degradation of myelin lipids or protein but produces necrosis in Schwann cells through inhibiting protein synthesis. 181,182 Young animals have been reported to be more susceptible to the demyelinating effects possibly due to their greater demand of myelin synthesis. 183
Disulfiram and N,N-diethyldithiocarbamate
These compounds find a wide array of applications in agriculture, medicine, and industry. The most generally recognized neurotoxicity of these compounds is the dose-dependent neuropathy associated with the use of disulfiram in alcohol aversion therapy. 184 –187 This compound is also currently being investigated as an adjunct treatment for glioblastoma. 188 The therapeutic basis for alcohol aversion therapy is disulfiram’s ability to inhibit aldehyde dehydrogenase-2 (ALDH2) that leads to elevated levels of acetaldehyde following ingestion of ethanol accompanied by flushing, headache, and nausea among other unpleasant symptoms and clinical signs. In animal models, oral disulfiram was observed to produce intramyelinic edema and segmental demyelination with accompanying secondary axonal degeneration. 189 Disulfiram is reduced to 2 molecules of N,N-diethyldithiocarbamate in vivo, and mechanistic toxicity studies have mainly focused on this metabolite that also finds considerable application on its own in agriculture and industry. Interestingly, N,N-diethyldithiocarbamate has been demonstrated to be capable of producing either a primary myelinopathy 190 or primary axonopathy 191 depending on the route of exposure. When administered orally, N,N-diethyldithiocarbamate undergoes acid promoted decomposition to carbon disulfide and N,N-diethylamine in the acidic environment of the stomach (Figure 10). 192 The CS2 released can be sufficient to produce a neurofilamentous axonopathy through protein adduction and cross-linking (Figure 6B). 136,193 In contrast, parenteral administration of N,N-diethyldithiocarbamate produced a primary myelinopathy characterized by myelin splitting and segmental demyelination similar to that reported for disulfiram. 190 Further investigation determined that the myelinopathy was dependent upon intact N,N-diethyldithiocarbamate forming lipophilic copper complexes that accumulated in peripheral nerve including the myelin compartment. The copper complexes are redox active and promote injury through promoting oxidative injury with lipid peroxidation (Figure 10). 194 –196 Analogs of N,N-diethyldithiocarbamate that also complex copper but have hydrophilic nitrogen substituents were not able to accumulate copper in nerve or produce a myelinopathy. 197 That intact parent dithiocarbamate is the proximate toxic species was supported by the report that oral administration of acid-stable pyrrolidine dithiocarbamate was able to produce the same myelin lesions as disulfiram. 198 Thus, the myelinopathy produced by disulfiram is interpreted to result from the acid stability of disulfiram facilitating systemic delivery of parent compound that subsequently undergoes reduction to N,N-diethyldithiocarbamate.
Lysolecithin and Ethidium Bromide
Injection or topical application of either of these compounds in the central or peripheral nervous systems produces an acute focal demyelination while sparring axons. 199 –201 Because total remyelination is usually achieved and the precise location of injury is known, these agents have been used as a model to test remyelination therapies and diagnostic methods for demyelinating diseases such as multiple sclerosis or hereditary peripheral nerve diseases. The mechanism for the demyelination produced by ethidium bromide is not known but recent investigations have provided insight into how lysolecithin (lysophosphatidylcholine) produces demyelination.
In addition to the interest of lysolecithin in the experimental setting, it is also relevant as a possible contributor in demyelinating disease states. Lysolecithin is an endogenous lysophospholipid produced by phospholipase A2 and is elevated in a number of neuropathological conditions including ischemia, 202,203 spinal cord injury, 204 amyotrophic lateral sclerosis, 205 and multiple sclerosis. 206 Additionally, phospholipase A2 is upregulated in the reprogrammed Schwann cell demyelinating phenotype after injury. 83 Elevations of lysolecithin in these disease states suggest it may act as an endogenous demyelinating agent.
Using a combination of in vitro and in vivo systems, Plemel et al. 207 investigated the relative contributions of 3 demyelinating mechanisms: receptor-mediated toxicity, inflammation-induced injury, and nonspecific membrane disruption. From their studies, it was concluded that lysolecithin integrates into cellular membranes and, when it reaches a critical concentration, causes an increase in cell membrane permeability. Thus, the data suggest that lysolecithin acts as a nonspecific lipid-disrupting agent with the first stage of injury corresponding to the unraveling of the myelin sheath. 208 Supporting this interpretation is the ability to reproduce similar myelin lesions using 2 other lipid-disrupting agents Brij35 and Tween 20. 207
Vulnerabilities of Neurons to Toxicants
Certain peripheral neurotoxicants selectively affect the neuronal perikaryon resulting in injury or death, a condition termed neuronopathy. A significant consequence of neuron death is that it is irreversible and accompanied by the loss of all associated cytoplasmic extensions. Thus, when significant loss of axons is detected in the peripheral nervous system, there exists the potential that it is the result of a primary neuronopathy.
The selectivity of certain agents for neurons typically can be attributed to the ability of the agent to target one of their unique vulnerabilities. Neurons have a high demand for energy primarily derived through aerobic glycolysis that is required for repolarization of the membrane and axonal transport. Also unique to neurons is the magnitude of biosynthetic requirements necessary to support the volume of axoplasm that can be orders of magnitude greater than the neuronal cell body. Selectivity can also result from transport systems associated with neuronal populations that concentrate an agent to toxic levels within that cell type. 6-Hydroxydopamine demonstrates an example of such selective uptake by catecholaminergic neurons in the peripheral nervous system. Neurons in the peripheral nervous system may also demonstrate increased susceptibility due to decreased integrity of the blood–nerve barrier at sensory and autonomic ganglia where neurons reside in the periphery. 26 –29 Support for this concept is provided through the observation that agents producing peripheral neuronopathies produce more widespread toxicity in the nervous system when the blood–brain barrier is disrupted experimentally.
Toxic Disorders of the Neuron
Retrograde signaling of injury from the axon results in the chromatolysis response (“axonal reaction”) in the neuronal perikaryon characterized by neuronal enlargement, rounding of the cytoplasmic membrane, eccentric displacement of the nucleus, and loss of stainable Nissl substance. 209,210 Dissolution of Nissl substance (ie, free ribosome storage depots associated with rough endoplasmic reticulum) reflects recruitment of ribosomes for accelerated protein synthesis to support cell body and/or cell process repair. There is also increased metabolic activity and transcriptional changes to support regeneration of the injured axon. 211,212 However, alterations in expression of genes that contribute to cell death and inflammation also occur and it is likely a balance in the overall expression profile that decides the fate of the neuron. 213,214 The response of the neurons is signaled to support cells and corresponding gene expression and proliferation occurs in the ganglion satellite cells and macrophages. 215,216 Defining the gene expression profiles in the cells of ganglia in response to peripheral nerve injury holds promise for the development of diagnostic biochemical methods of peripheral nerve injury and the design of therapeutic interventions to prevent neuron cell death and facilitate regeneration.
Cell death of the neuron can proceed through necrosis or one of the programmed cell death pathways. Although there are a number of recognized cell death pathways including autophagy and necroptosis in addition to apoptosis, the currently recognized agents targeting the neuron appear to work through either necrosis or apoptosis. Biochemical and morphological methods can be used to determine which pathway is involved and have been described in detail in other reviews. 217,218 Morphologically, necrosis involves cell swelling and disintegration of cellular organelles from lipases, endonucleases, and proteases that are activated by increased intracellular Ca2+ levels that leads to rupture of the nuclear and cell membranes invoking an inflammatory response. Biochemically, there is a cessation of protein synthesis and the termination of energy-dependent processes due to the depletion of ATP. Pyridoxamine is an example of an agent targeting dorsal root ganglion (DRG) neurons that produce cell death through necrosis. 16 In contrast, apoptosis is an active process requiring energy and protein synthesis that proceeds through an orchestrated pathway with specific regulators that can either inhibit or activate the process. Apoptotic cells become shrunken with fragmentation of their chromatin but maintain the structure of their organelles. There is a translocation of phosphatidylserine to the cell surface that enables microglia or macrophages to remove the dead cell with its plasma membrane intact, thus precluding the development of an inflammatory response. The platinum-based chemotherapeutic agents produce cell death in DRG neurons through the apoptosis pathway.
Examples of Agents Producing Peripheral Neuronopathies
Chemotherapeutic Platinum Derivatives
Cisplatin, oxaliplatin, and carboplatin are platinum-based compounds used to treat lung, breast, ovarian, and gastrointestinal cancers. In addition to other side effects, these chemotherapeutics are associated with a high prevalence of peripheral neuropathy. Large-diameter sensory fibers appear to be the most sensitive resulting in symmetrical glove-stocking distribution of sensory loss including numbness and paresthesias. Neurotoxicity is related to cumulative dose, can persist for months to years, and can even worsen for a period following cessation of treatment. In addition, oxaliplatin is recognized to produce an acute non-dose-dependent neuropathy very shortly after infusion exhibited by cold induced paresthesia and dysesthesia of distal extremities. These agents target DRG through their ability to accumulate and interact with DNA in this location. 219 –221 The high density of capillaries 27 that are relatively permeable 28,29 contributes to an incomplete blood–nerve barrier allowing platinum drugs preferential access to this region of the peripheral nervous system. Uptake by the sensory neurons may also be facilitated by 2 types of specific transporters, the copper transporter 1 and the organic cation transporter-2. 222 –224
These agents form platinum intrastrand adducts and interstrand cross-links. The DNA modifications compromise transcriptional activity in this population of cells noted for their high rate of transcription required to sustain the metabolic demands of maintaining the associated axon. The altered 3-dimensional structure of DNA along with activation of Cyclin D1 and MAPKs are thought to promote entry of these neurons into the cell cycle, which results in a compensatory process of apoptosis in these postmitotic cells. 225,226 In addition to nuclear DNA, mitochondrial DNA is also modified. The interactions within mitochondria result in increased oxidative stress, increased p53 tumor suppressor protein (p53) activity, release of cytochrome c and activation of p38 mitogen-activated protein kinases (p38) and extracellular signal-regulated kinases 1 and 2 (ERK1/2). 114,227 Compromised mitochondrial function also leads to a depletion of energy stores vital to this population of cells. Collectively, these processes work to activate recognized intrinsic signaling to initiate apoptotic pathways. 225,228 –230
Oxaliplatin can also exert an effect on the kinetics of the neuron voltage-gated sodium ion channels. 231 –234 It acts directly at the channel and possibly indirectly through chelation of Ca2+ to increase the duration of the refractory period. 231 This effect is thought to contribute to the acute clinical signs of cold paresthesia observed for oxaliplatin from the sensory neurons becoming hyperexcitable. Sensory neurons express transient receptor potential channels that contribute to the neuropathic pain. Oxaliplatin activates the transient receptor potential cation channel subfamily A member 1 and the transient receptor potential cation channel subfamily M member 8 providing another mechanism through which this compound may promote cold allodynia. 235 –237
Methylmercury
Environmental mercury is an important pollutant that exists in both inorganic and organic forms, both of which can produce neurological and developmental effects in humans and wildlife. Methylmercury (organic form) has been responsible for a number of human poisonings and has been the focus of considerable investigation. 238 The clinical signs associated with methylmercury intoxication include sensory impairment of the extremities, cerebellar ataxia, and visual field constriction. 239 In addition to neuron loss in the visual area of the calcarine cortex and granule cells of the cerebellum, methylmercury also produces degeneration of neurons in the DRG along with their associated axons while sparing the peripheral nerve motor axons. 239 –243
Methylmercury exerts its neurotoxic affects in DRG partly through the ability to accumulate there due to the high density of fenestrated capillaries. 26 –29 Although the molecular targets and mechanisms of methylmercury are still being delineated, the experimental evidence supports contributions from several processes including disrupted calcium homeostasis, increased oxidative stress, and altered glutamate homeostasis leading to excitotoxicity. 238,244 Methylmercury alters intracellular calcium levels through disrupting sequestration of intracellular calcium pools and through increasing permeability of the plasma membrane for calcium. 245 Elevated oxidative stress may be mediated through the ability of methylmercury to interact with sulfhydryl and selenocysteine functions of antioxidants and antioxidant proteins. Oxidative stress could also be elevated through the accumulation of methylmercury in mitochondria that decreases ATP production and depolarizes the mitochondrial membrane potential. 245 Consistent with the effects on mitochondria, the activation of signaling pathways involving release of cytochrome c and apoptosis-inducing factor leading to both caspase-dependent and caspase-independent apoptotic cell death pathways have been observed in methylmercury-treated cells and apoptosis has been observed in granule cell and DRG cells. 244,246 Collectively, the number and diversity of significantly altered cellular processes produced by methylmercury suggest the involvement of multiple mechanisms that may act additively or synergistically.
Catecholamines
If not processed correctly through sequestration and storage in a low pH environment, catecholamines can oxidize to form quinones that can via redox cycling produce reactive oxygen species which cause deleterious effects in neurons. 247 Dopamine is particularly sensitive to oxidation and an analog of dopamine, 6-hydroxydopamine, has been used to lesion the central and peripheral nervous systems experimentally. When administered peripherally, this compound does not cross the blood–brain barrier and is taken up by transporters at the sympathetic nerve terminals producing a sympathectomy. 248 The loss of sympathetic neurons results from retrograde transport of 6-hydroxydopamine to the neuronal perikaryon and increased oxidative stress from the oxidation of 6-hydroxydopamine and subsequent redox cycling similar to that observed with dopamine. 249,250 It has also been proposed that 6-hydroxydopamine can inhibit mitochondrial respiration chain complexes I and IV. 251 Over expression of copper/zinc (CuZn) superoxide dismutase or glutathione peroxidase has been shown to protect against 6-hydroxydopamine neurotoxicity. 252,253
Conclusion
There are many recognized peripheral nervous system toxicants but our understanding of their mechanisms is limited and there are few examples where a single mechanism is generally agreed upon to account for all the pathological events observed in a given toxic neuropathy. For those that have been investigated, there are usually a number of potential contributing events recognized. In addition, the diversity of toxicants and the structural complexity and spatial distribution of the peripheral nervous system present a formidable challenge for screening neurotoxicity. However, the capacity of an agent to target the peripheral nervous system can usually be attributed to its ability to circumvent the blood–nerve barrier and compromise one of the vital and unique characteristics the nervous system has evolved to perform its function of rapidly conducting information over distance. Once these requirements have been met, there are several general pathogenic processes through which most toxic agents ultimately mediate their effects including covalent or noncovalent modifications, compromised energy or protein biosynthesis, oxidative injury, and disruption of ionic gradients across membranes. Because programmed neuronal cell death, demyelination, and axonal degeneration proceed through active programmed pathways, the triggers for these pathways may be the final targets of neurotoxicants that become activated through the shared general pathogenic processes. Thus, although the initial critical actions of neurotoxicants may be quite varied and multiple for any given agent, there is the potential that common final pathways can be identified to help guide the development of biochemical based methods to complement morphological analyses.
Footnotes
Acknowledgments
I wish to thank Douglas Anthony and Brad Bolon for their help with reviewing electron micrographs and proof reading of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received the following financial support for the research, authorship, and/or publication of this article: The studies on 1-bromopropane, carbon disulfide, disulfiram and dithiocarbamates were supported in part by NIEHS Grants ES06387 and P30 ES00267.