
Abstract
Diacylglycerol acyltransferase 1 (DGAT1) catalyzes the final step in triglyceride synthesis. Since Dgat1 -/- mice fed a high-fat diet (HFD) are resistant to hepatic steatosis, DGAT1 inhibitors are expected to have antifatty liver effects. To evaluate the hepatic effects of DS-7250, a selective DGAT1 inhibitor, vehicle or 10 mg/kg of DS-7250 was administered orally to male Fisher 344 (F344) and Zucker fatty (ZF) rats fed a standard diet or HFD for 14 or 28 days. ZF rats showed slight hepatic steatosis regardless of feeding conditions. DS-7250 exacerbated hepatic steatosis in ZF rats fed an HFD compared with the vehicle control. Hepatic steatosis did not occur in F344 rats fed an HFD, in which systemic exposures of DS-7250 were comparable to those in ZF rats. There was a higher expression of genes involved in lipid uptake and fatty acid synthesis in ZF rats compared to F344 rats under HFD conditions. DS-7250 upregulated key genes involved in de novo lipogenesis, which causes hepatic steatosis independently of DGAT1, in ZF rats fed an HFD compared with the vehicle control. These data suggest that ZF rats were more susceptible to hepatic steatosis due to their genetic characteristics and DS-7250 exacerbated hepatic steatosis independently of DGAT1.
Triglycerides (TGs) are mainly synthesized in hepatocytes and enterocytes following the absorption of dietary fat. In cases where there is an excess of energy intake, large amounts of fat begin to accumulate as TGs in adipose tissue, and over time, an individual can be diagnosed as being “obese.” The final step in TG synthesis is the formation of an ester bond between acyl-CoA and diacylglycerol, which is catalyzed by diacylglycerol acyltransferase (DGAT). DGAT enzymes are encoded by two nonhomologous genes: DGAT1 and DGAT2 (Cases et al. 1998; Lardizabal et al. 2001). Both DGAT1 and DGAT2 are expressed in the tissues responsible for the synthesis of TG; DGAT1 is expressed mostly in enterocytes of the small intestine (Chen and Farese 2000), whereas DGAT2 is widely expressed with high levels in hepatocytes and adipocytes (Cases et al. 1998). DGAT1 plays a crucial role in enterocytes for the assembly of TGs into lipoproteins, which are transported between tissues.
DS-7250 was developed as an anti-obesity drug by Daiichi Sankyo Co., Ltd. (Tokyo, Japan). The 50% inhibitory concentration (IC50) for DGAT1 is 2.26 nM in humans and 3.24 nM in rats, while the IC50 for DGAT2 is >100 μM in humans. In addition, the drug does not show any specific binding property to other receptors, channels, or transporters, indicating that DS-7250 is a selective DGAT1 inhibitor. Since Dgat1 -/- mice were reported to be phenotypically lean and resistant to diet-induced obesity (DIO) and hepatic steatosis (Chen et al. 2003; Chen and Farese 2005), DS-7250 is expected to show pharmacological effects through the inhibition of TG synthesis in enterocytes, followed by the impairment of fat absorption into the circulation.
Obesity is one of the greatest concerns in the development of hepatic steatosis (Li et al. 2016). Indeed, obese rodent models such as ob/ob mice, DIO rats, and Zucker fatty (ZF) rats are known to develop hepatic steatosis (Takahashi, Soejima, and Fukusato 2012; Kucera and Cervinkova 2014). ZF rats are one of the most widely used animal models for genetic obesity. The fa mutation in ZF rats affects the extracellular part of the leptin receptor, which leads to a weaker affinity for leptin and alters signal transduction (White et al. 1997; Yamashita et al. 1997). As a result, ZF rats exhibit increased food intake and obesity accompanied by hyperlipidemia including higher TG and total cholesterol levels with age (Fellmann et al. 2013). Therefore, ZF rats develop early hepatic steatosis spontaneously and accelerate the progression of hepatic steatosis with high-fat diet (HFD) feeding (Carmiel-Haggai, Cederbaum, and Nieto 2005; Buque et al. 2010).
The inhibition of DGAT1 is known to protect against hepatic steatosis caused by HFD feeding in mice (Villanueva et al. 2009). Therefore, DS-7250 was expected to inhibit the development of hepatic steatosis in ZF rats fed an HFD by the suppression of fat absorption in the small intestine. The purpose of this study was to evaluate the effects of DS-7250 on hepatic steatosis in ZF rats fed an HFD by comparing to ZF rats fed a standard diet (STD) or Fisher 344 (F344) rats fed an HFD or STD. Furthermore, hepatic transcriptome analysis was conducted in HFD-fed F344 and ZF rats with or without DS-7250 administration.
Materials and Method
Drug
DS-7250 (chemical name: [trans-4-({[6-{[(4-methylphenyl) carbamoyl]amino}-3,4-dihydroisoquinolin-2(1 H)-yl]carbonyl}oxy)cyclohexyl]acetic acid; molecular formula: C26H31N3O5) was synthesized at Daiichi Sankyo Co., Ltd. DS-7250 was suspended in 0.5% methylcellulose solution (0.5% MC, Wako Pure Chemical Industries, Ltd., Osaka, Japan) at a concentration of 1 mg/ml.
Animals
Male ZF rats (Slc: Zucker-fa/fa, specific-pathogen free [SPF]) and F344 rats (F344/DuCrlCrlj, SPF) at ages of 6 to 7 weeks were purchased from Japan SLC, Inc., and Charles River Laboratories Japan, Inc., respectively. Rats were housed individually in a cage under controlled conditions (room temperature: 20–26°C; humidity: 30–70%; and 12-hr light–dark cycles) with ad libitum access to chlorinated water and a diet. All animal studies were approved by the Institutional Animal Care and Use Committee in Daiichi Sankyo Co., Ltd.
Study Design
The present study included three experiments as described in Figure 1. Each experiment was conducted on a different time period. The first day of dosing was described as day 1. All animals used in this study were euthanized at approximately the same time of day.
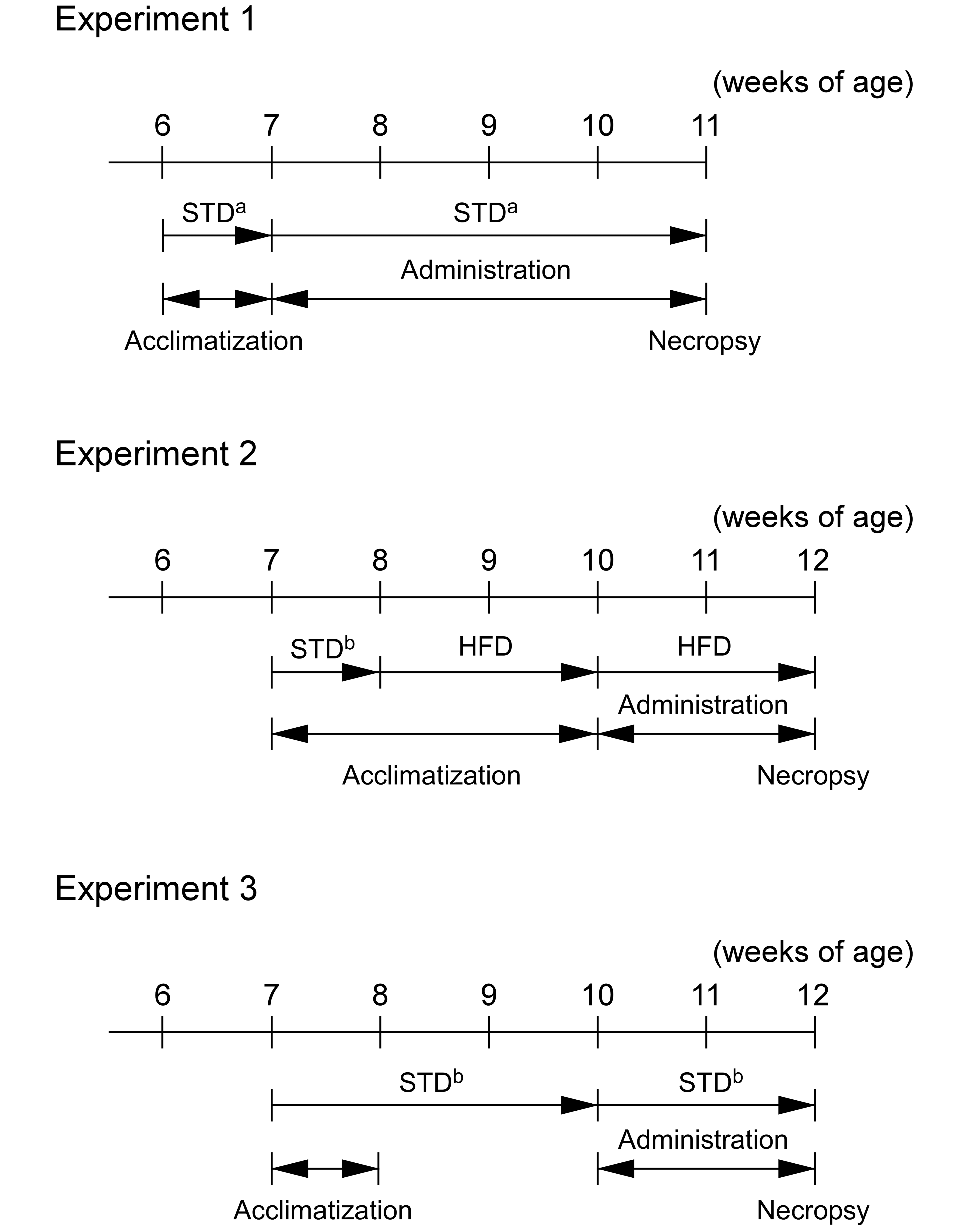
Experimental protocol. In experiment 1, vehicle (0.5% methyl cellulose) or 10 mg/kg of DS-7250 was administered orally to Fischer 344 (F344) rats for 28 days. A standard diet (STD) was given to animals during the study period including a week of acclimatization. In experiment 2, vehicle or 10 mg/kg of DS-7250 was administered orally to F344 and Zucker fatty (ZF) rats for 14 days. A high-fat diet (HFD) was given to animals from 2 weeks before the dosing period to the end of the dosing period. In experiment 3, vehicle or 10 mg/kg of DS-7250 was administered orally to ZF rats for 14 days. An STD was fed to animals during the study period. STD = standard diet (a. CRF-1, 14 kcal% fat; b. FR-2, 10 kcal% fat), HFD = high-fat diet (45 kcal% fat).
Experiment 1: 28-day dosing of DS-7250 in F344 rats fed an STD
Male F344 rats were fed an STD (CRF-1, Oriental Yeast Co., Ltd., approximately 14 kcal% fat) during the study period including a week of acclimatization to the laboratory environment. The rats received daily gavage of vehicle (0.5% MC) or DS-7250 (10 mg/kg) for 28 days. The dose level of DS-7250 was selected as a dose that showed pharmacological effects on obese model rats. All animals were necropsied on the day after the final dosing (day 29).
Experiment 2: 14-day dosing of DS-7250 in F344 or ZF rats fed an HFD
Male F344 rats or ZF rats were fed an STD (FR-2, Funabashi Farm Co., approximately 10 kcal% fat) for a week of acclimatization to the laboratory environment, followed by an HFD containing 45 kcal% fat (D12451, Research Diets, Inc., New Brunswick, NJ) for a 2-week acclimatization to an HFD. The rats received daily gavage of vehicle (0.5% MC) or DS-7250 (10 mg/kg) for 14 days. Animals were also fed an HFD during the dosing period. All animals were necropsied on the day following final dosing (day 15).
Experiment 3: 14-day dosing of DS-7250 in ZF rats fed an STD
Male ZF rats were fed an STD (FR-2) for 3 weeks before the initiation of dosing. The rats received daily gavage of vehicle (0.5% MC) or DS-7250 (10 mg/kg) under STD conditions for 14 days. All animals were necropsied on the day following final dosing (day 15).
Examinations
Body weight and food consumption
Body weight and food consumption were measured approximately twice a week during the dosing periods.
Blood chemistry
Blood was collected via the abdominal aorta under isoflurane anesthesia on the day following final dosing (day 15 or 29). Plasma total cholesterol, TGs, nonesterified fatty acids (NEFAs), and glucose were measured using TBA-2000FR (Toshiba Medical Systems Corporation, Tochigi, Japan).
Pathological examination
Animals were euthanized by exsanguination from the abdominal aorta under isoflurane anesthesia. The liver was collected and fixed in 10% neutral-buffered formalin. Left lateral and right medial lobes of the liver were embedded in paraffin via a routine procedure. Paraffin sections were stained with hematoxylin and eosin for light microscopy. Oil Red O staining was performed in hepatocytes using cryostat sections of formalin-fixed left lateral lobe. Lipid droplets in hepatocytes on Oil Red O slides were morphometrically evaluated using STEPanizer (http://www.stepanizer.com/; Tschanz, Burri, and Weibel 2011) in the single section from each animal. Densities of Oil Red O-positive droplets in hepatocytes were determined by point grids as lipid droplets per hepatocytes (%). Approximately 50% of each section was evaluated (a total of 50 fields at the 400× magnification per section).
Microarray analysis
The left lateral lobe of the liver obtained from HFD-fed F344 and ZF rats (experiment 2) was stored at −80°C until use. Microarray analysis was performed by GeneChip 3′IVT PLUS Regent Kit (Affymetrix Inc., Santa Clara, CA) for the synthesis of biotin-labeled cDNA. Every biotin-labeled cDNA target sample (approximately 10 μg) was individually hybridized to a GeneChip Rat Genome 230 2.0 Array (Affymetrix Inc.) at 45°C for 16 hr followed by washing and staining by streptavidin–phycoerythrin using Fluidics Station 450 (Affymetrix Inc.). The scanned image was analyzed with an Affymetrix MicroArray Suite 5 (MAS5) algorithm using GeneChip Operating Software (version 1.4; Affymetrix Inc.). All the MAS5-analyzed data were scaled by global normalization. The upregulated and downregulated genes were selected by the following criteria: (1) the detection call of each probe as “Present (P)” in >3/4 replicates and (2) the statistical significance from the concurrent control with a p value < .05 (t-test). The data were imported into BaseSpace Correlation Engine (Illumina Inc., San Diego, CA) to identify biogroups with the Broad Institute’s Molecular Signatures Database (MSigDB) based on canonical pathway analysis (Liberzon et al. 2011). In addition, Ingenuity Pathway Analysis (IPA; QIAGEN, Hilden, Germany) was utilized for gene annotations.
Toxicokinetics
Blood was collected via the jugular vein prior to dosing (on the final day of dosing only), and 2, 4, 7, and 24 hr after dosing on day 1 and the final day of dosing (day 14 or 28) under conscious conditions. Plasma concentrations of DS-7250 were measured by liquid chromatography tandem–mass spectrometry (API 4000, AB Sciex, Tokyo, Japan). As toxicokinetic parameters, AUC0–24 hr, C max, and T max were determined.
Statistical Analysis
A Grubbs’ outlier test was performed on data of blood chemistry and liver morphometry. Quantitative data were expressed as mean ± standard deviation (SD). The comparisons were between the vehicle control and DS-7250-treated groups in each strain. The parameter was first analyzed by F test to evaluate the homogeneity of variance (5% significance level), followed by the Student’s t-test when the variance was homogeneous or by Welch’s t-test when it was not. The analyses by t-test were done at the 5% significance level, and the results were classified as significant at “less than 5%” and “less than 1%” based on the p values.
Results
Body Weight and Food Consumption
DS-7250 at 10 mg/kg did not impact body weight or food consumption in F344 rats regardless of feeding conditions (Figure 2). In contrast, DS-7250 did not affect body weight but reduced food consumption by 6 to 18% in ZF rats fed an STD compared to that of the respective vehicle-treated rats throughout the dosing period (Figure 2). In addition, DS-7250 caused decreases in body weight gain from day 8 to day 14 and food consumption by 20−30% in ZF rats fed an HFD as compared to that of the respective vehicle-treated rats (Figure 2).
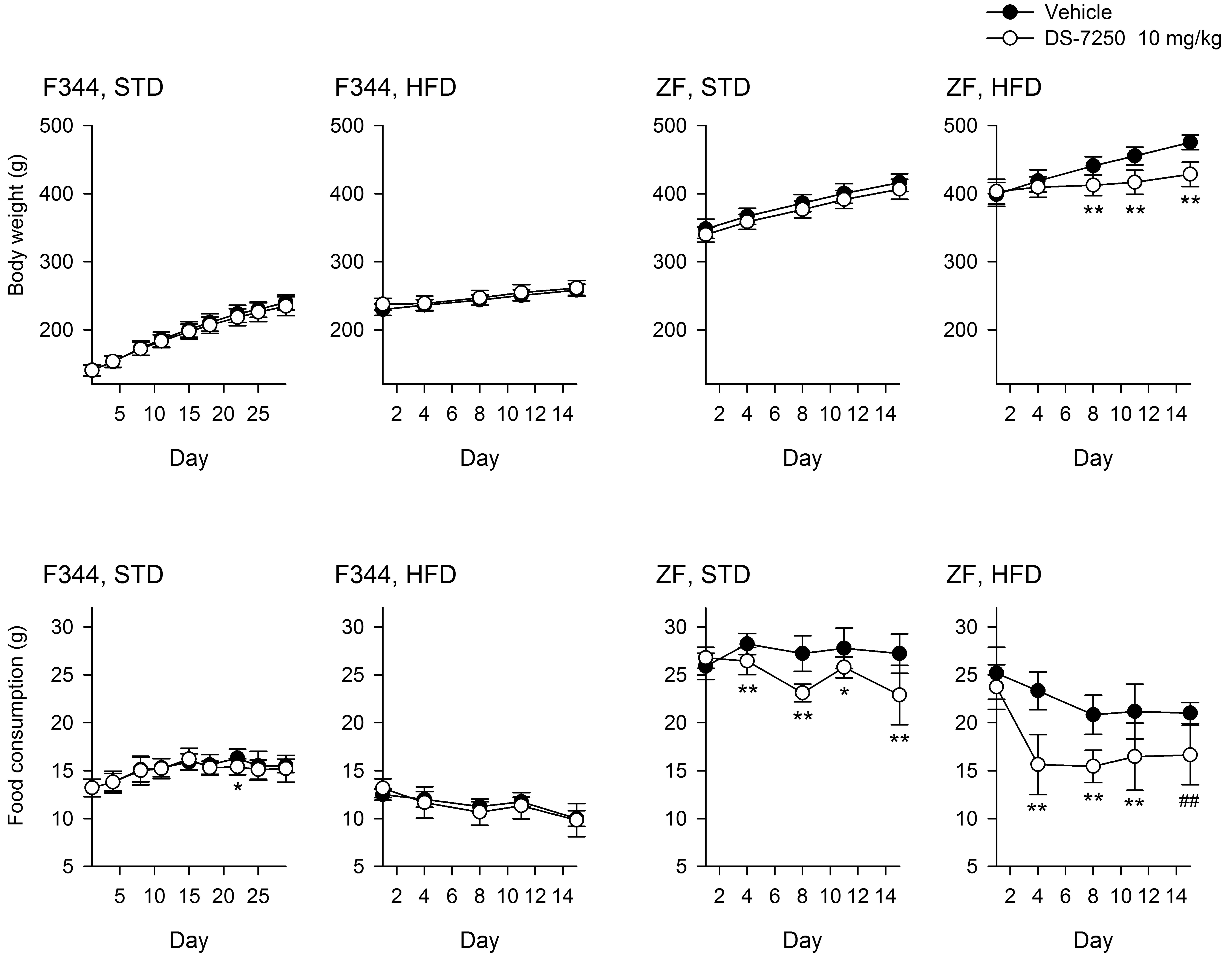
Effects of DS-7250 on body weights and food consumption in Fisher 344 (F344) or Zucker fatty (ZF) rats. STD = standard diet, HFD = high-fat diet. The first day of dosing was described as day 1. Data are expressed as mean ± standard deviation. Significantly different from the respective vehicle control: *p < .05, **p < .01 (Student’s t-test), ##p < .01 (Welch’s t-test).
Blood Chemistry
DS-7250 lowered plasma total cholesterol by approximately 10% in both F344 and ZF rats fed an HFD and increased plasma TG by approximately 45% and 120%, respectively, in F344 and ZF rats fed an HFD as compared to the respective control (Figure 3). DS-7250 increased plasma TG by ca. 17% and 10%, respectively, in F344 rats fed an HFD and ZF rats fed an STD and also increased NEFA by ca. 30% in ZF rats fed an STD as compared to those of the respective vehicle-treated rats (Figure 3).
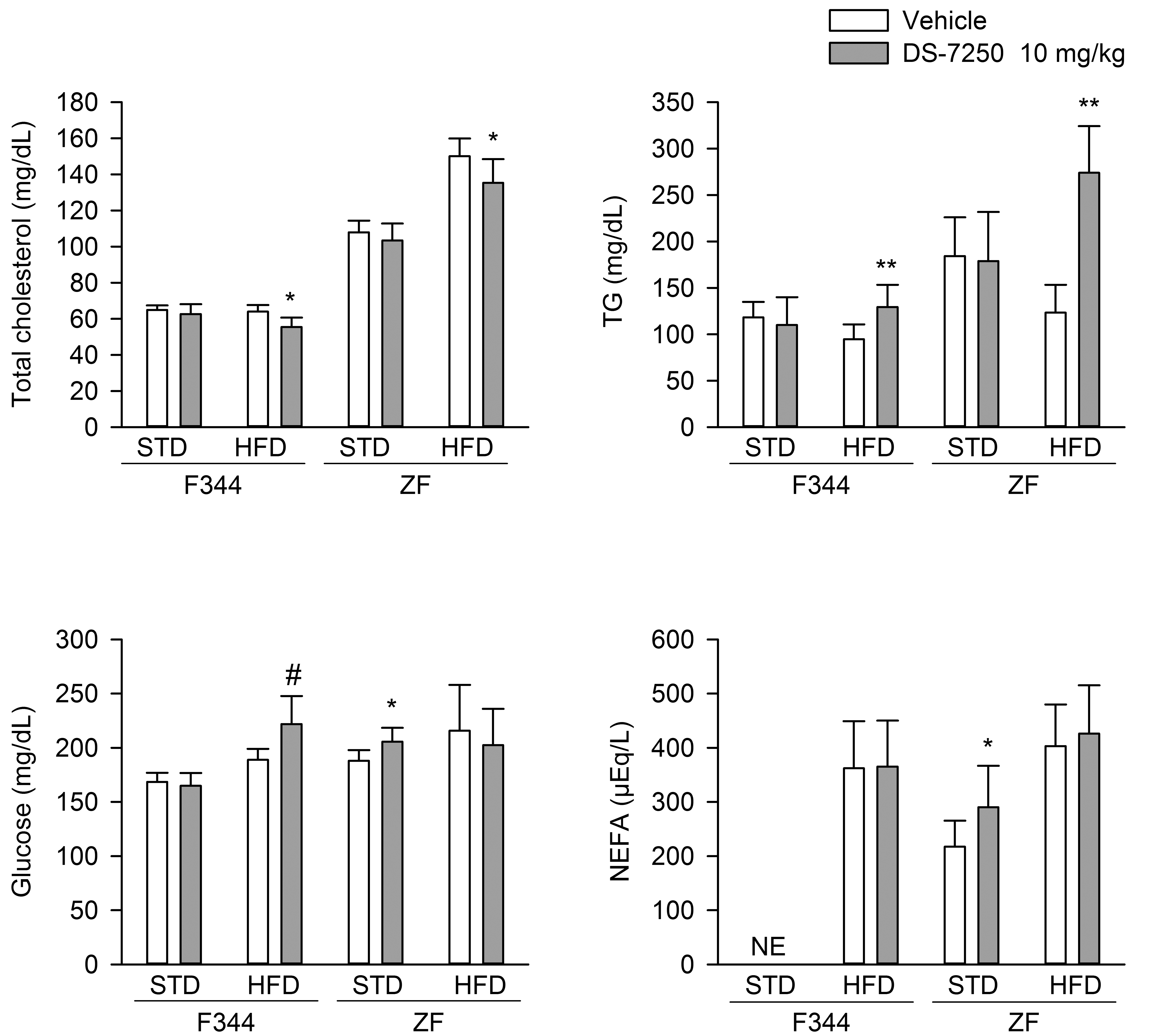
Effect of DS-7250 on blood lipid and glucose levels in Fischer 344 (F344) and Zucker fatty (ZF) rats. Plasma total cholesterol, triglyceride (TG), glucose, and nonesterified fatty acid (NEFA). STD = standard diet, HFD = high-fat diet, NE = not examined. Data are expressed as mean ± standard deviation. Significantly different from the respective vehicle control: *p < .05, **p < .01 (Student’s t-test), #p < .05 (Welch’s test).
Pathological Examination
DS-7250 exacerbated discoloration of the liver indicative of a fatty liver in ZF rats fed an HFD in gross pathology as compared to the control HFD-fed ZF rats (data not shown). In histopathology, ZF rats fed an HFD possessed hepatocytes that contained many vacuoles, and this was more remarkable in DS-7250-treated ZF rats as compared to the control rats (Figure 4). No hepatocellular necrosis or infiltrations of inflammatory cells were observed in any strain regardless of feeding conditions.
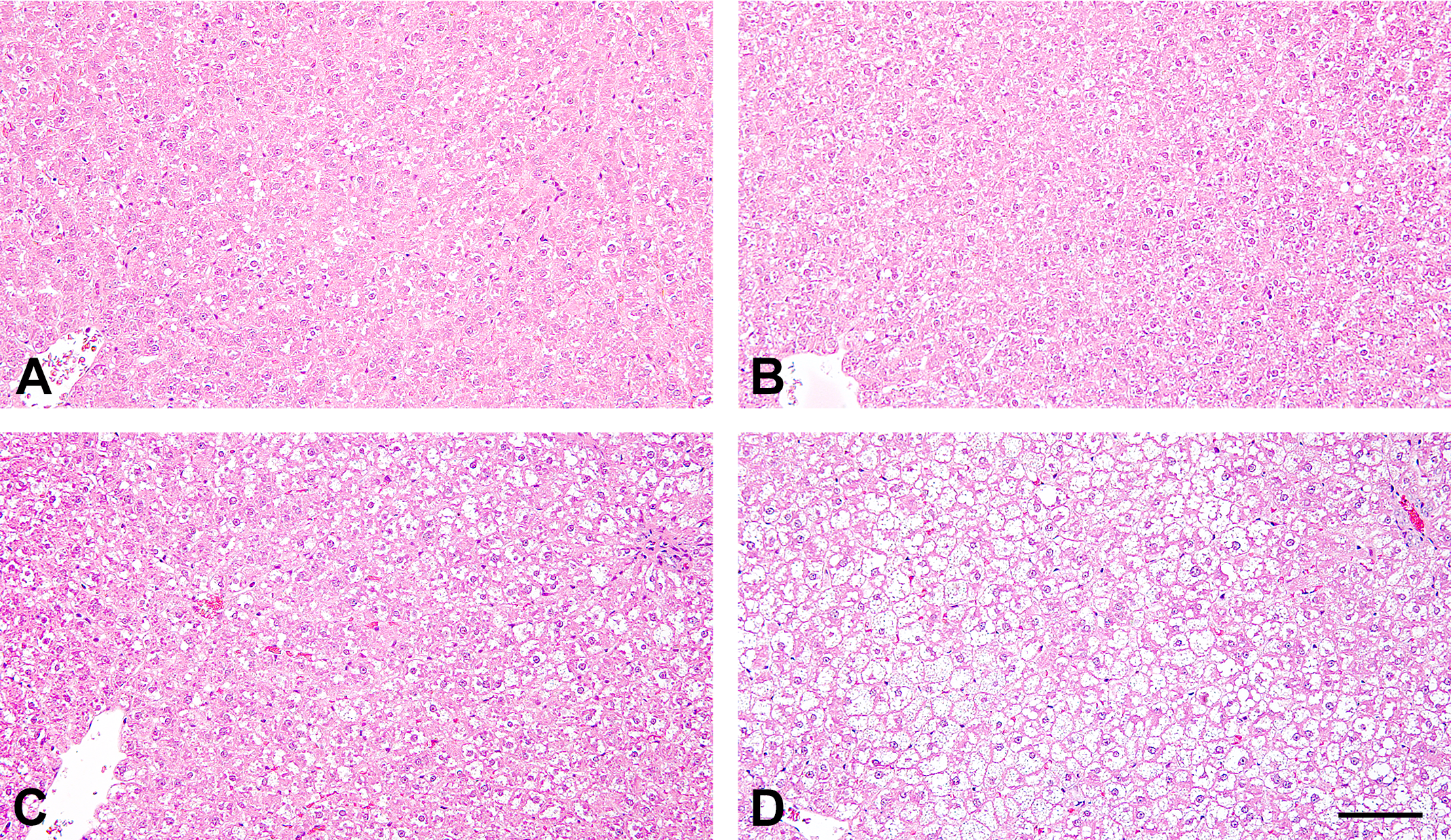
Representative photomicrographs of the liver sections stained with hematoxylin and eosin in Zucker fatty (ZF) rats. (A) Vehicle-treated ZF rats fed a standard diet (STD), (B) DS-7250-treated ZF rats fed an STD, (C) vehicle-treated ZF rats fed a high-fat diet (HFD), and (D) DS-7250-treated ZF rats fed an HFD. Scale bar = 100 μm.
Fine- to small-sized lipid droplets, stained by Oil Red O, were observed diffusely throughout the hepatic lobule in vehicle-treated ZF rats regardless of feeding conditions and in DS-7250-treated ZF rats fed an STD. In addition, macrovesicular steatosis was observed in DS-7250-treated ZF rats fed an HFD (Figure 5). Histomorphometry revealed that DS-7250 increased lipid droplets per hepatocyte slightly (1.7-fold) or markedly (5.6-fold) in ZF rats fed an STD or HFD, respectively, compared to the respective vehicle control (Figure 6). On the other hand, DS-7250 had no effect on the liver of F344 rats fed an HFD.
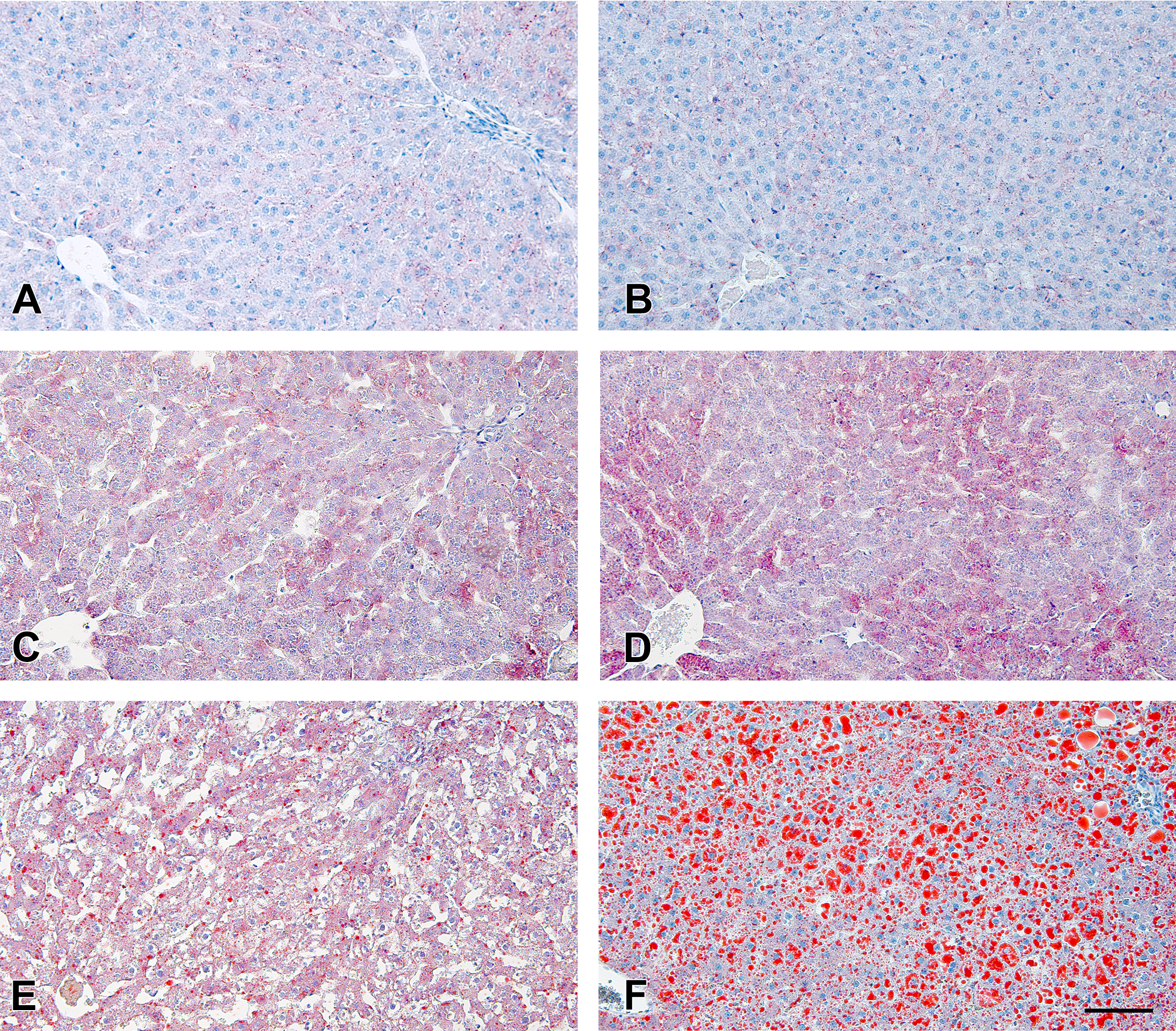
DS-7250 exacerbated hepatic steatosis in Zucker fatty (ZF) rats fed a high-fat diet (HFD). (A) Vehicle-treated Fischer 344 (F344) rats fed an HFD, (B) DS-7250-treated F344 rats fed an HFD, (C) vehicle-treated ZF rats fed a standard diet (STD), (D) DS-7250-treated ZF rats fed an STD, (E) vehicle-treated ZF rats fed an HFD, and (F) DS-7250-treated ZF rats fed an HFD. Oil Red O staining in the liver. Scale bar = 100 μm.
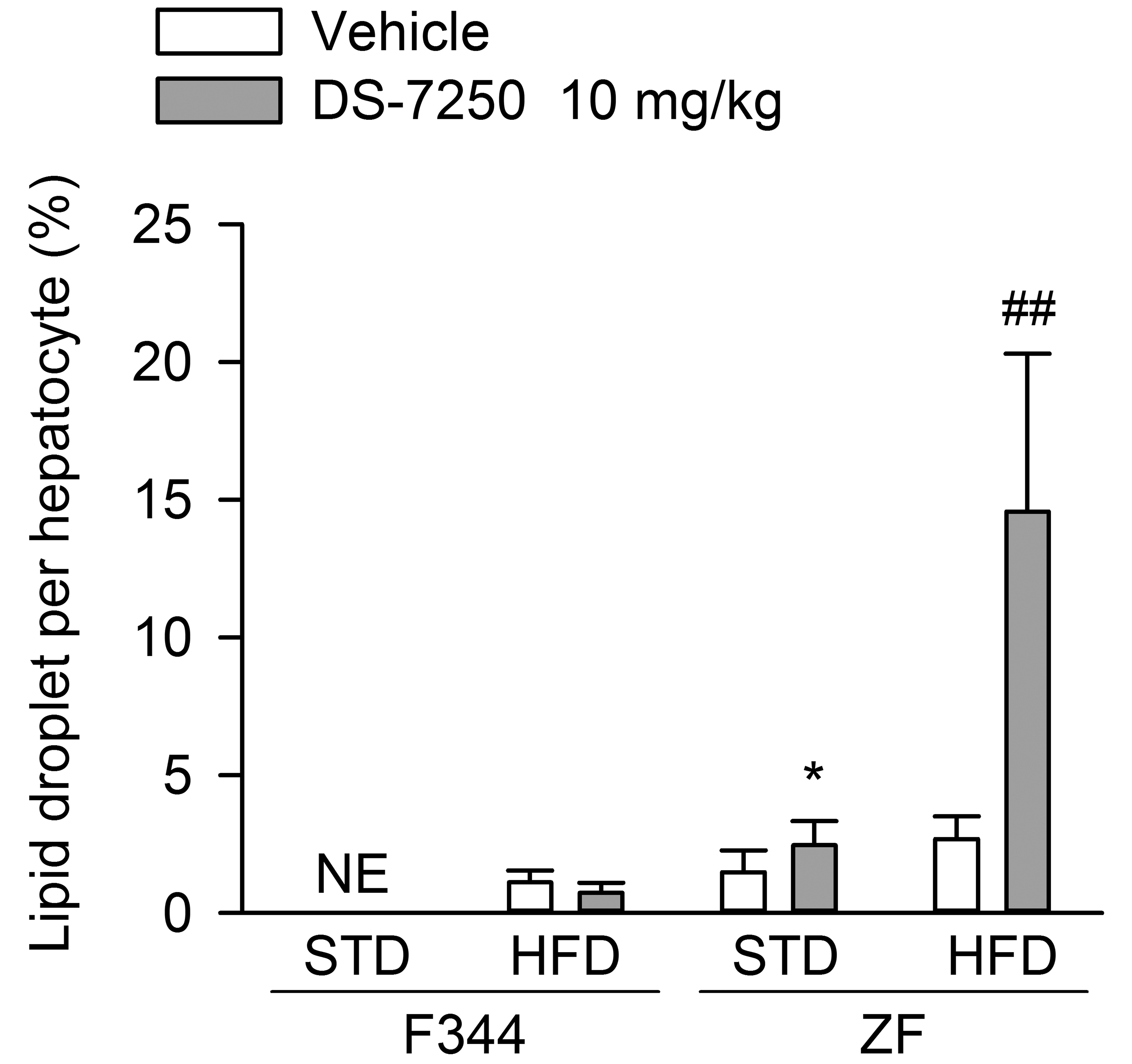
DS-7250 exacerbated hepatic steatosis in Zucker fatty (ZF) rats fed a high-fat diet (HFD). Histomorphometry of lipid droplets per hepatocyte. STD = standard diet, F344 = Fischer 344 rat. Data are expressed as means ± standard deviation. Significantly different from the respective vehicle control: **p < .01 (Student’s t-test), #p < .01 (Welch’s t-test).
Microarray Analysis
Trascriptome analysis identified 4,034 hepatic transcripts differentially expressed in vehicle-treated ZF rats fed an HFD relative to their expression levels in F344 rats fed an HFD. Specifically, 1,903 transcripts were upregulated and 2,131 were downregulated in ZF rats fed an HFD when compared to the F344 rats. In addition, transcriptome analysis identified differentially expressed transcripts associated with DS-7250 treatment relative to those of the corresponding control under HFD conditions. Specifically, 194 transcripts were upregulated and 322 were downregulated in F344 rats fed an HFD; 459 transcripts were upregulated and 417 downregulated in ZF rats fed an HFD (Table 1).
Number of Differentially Expressed Genes in Vehicle-treated and DS-7250-treated Fischer 344 (F344) and Zucker Fatty (ZF) Rats Fed a High-fat Diet.
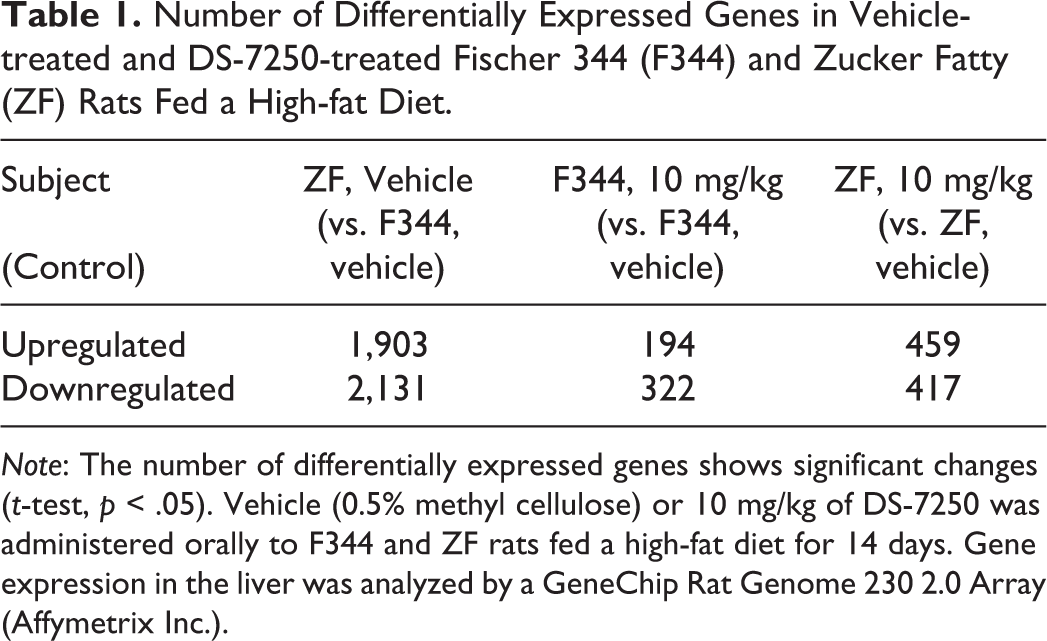
Note: The number of differentially expressed genes shows significant changes (t-test, p < .05). Vehicle (0.5% methyl cellulose) or 10 mg/kg of DS-7250 was administered orally to F344 and ZF rats fed a high-fat diet for 14 days. Gene expression in the liver was analyzed by a GeneChip Rat Genome 230 2.0 Array (Affymetrix Inc.).
Canonical pathway analysis using BaseSpace Correlation Engine with the MSigDB indicated that the top five biogroups with highly overlapped genes between vehicle-treated F344 and ZF rats fed an HFD included 155 genes involved in the metabolism of lipids and lipoproteins (Table 2). In addition, the canonical pathway analysis between vehicle-treated and DS-7250-treated ZF rats fed an HFD revealed that DS-7250 affected three biogroups (Table 2). Among them, 16 genes were involved in fatty acid, TG, and ketone body metabolism and 6 genes were involved in TG biosynthesis; overall, 42 genes involved in lipid metabolism and lipoproteins were identified. However, no significant change in the biogroups involved in lipid metabolism was determined between vehicle- and DS-7250-treated F344 rats fed an HFD (Table 2).
The Top Five Biogroups Identified for the Genes Altered in Vehicle- and DS-7250-treated Zucker Fatty (ZF) and Fischer 344 (F344) Rats Given a High-fat Diet.

Note: Analysis was done by the canonical pathway analysis using the BaseSpace Correlation Engine with the Broad Institute’s Molecular Signatures Database. List of genes showing statistically significant changes (t-test, p < .05) was imported to the BaseSpace Correlation Engine. The Overlap p values and the number of common genes were obtained by making a comparison between genes involved in each biogroup registered in the BaseSpace Correlation Engine and the differentially expressed genes in the present study. TG = triglyceride.
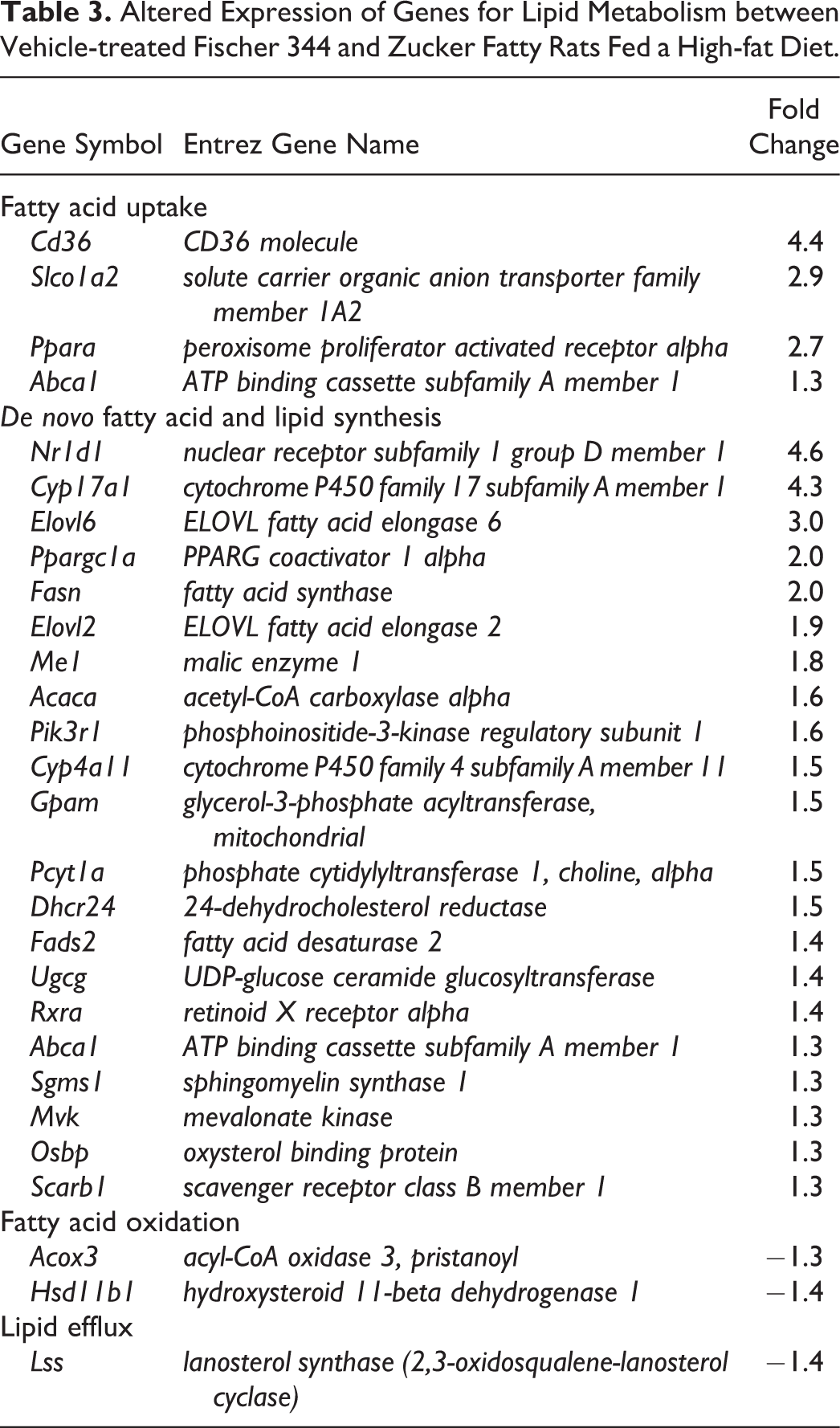
A list of 155 lipid metabolism genes differentially expressed in vehicle-treated ZF rats and 42 lipid metabolism genes in DS-7250-treated ZF rats were submitted to IPA for gene annotations. The genes were categorized into four groups central to hepatic lipid retention (i.e., fatty acid uptake, de novo fatty acid and lipid synthesis, fatty acid oxidation, and lipid efflux) based on functional annotation and manual curation (Angrish et al. 2016). Those genes that possessed a mean signal intensity value of at least 1.3-fold higher or 0.77-fold lower than a concurrent control value are shown in Tables 3 and 4. Key genes involved in fatty acid uptake and de novo fatty acid and lipid synthesis were upregulated in vehicle-treated ZF rats as compared with vehicle-treated F344 rats. These included critical genes for lipid metabolism such as CD36 (Cd36), ELOVL fatty acid elongase 6 (Elovl6), fatty acid synthase (Fasn), Elovl2, malic enzyme 1 (Me1), glycerol-3-phosphate acyltransferase, mitochondrial (Gpam), fatty acid desaturase 2, UDP-glucose ceramide glucosyltransferase, and mevalonate kinase (Table 3). In addition, DS-7250 increased Elovl6, Me1, Fasn, and Gpam mRNA levels in ZF rats. Furthermore, upregulation of sterol regulatory element binding transcription factor 1 (Srebf1) mRNA, which is a master regulator of lipid homeostasis, was noted only in DS-7250-treated ZF rats (Table 4).
Altered Expression of Genes for Lipid Metabolism between Vehicle-treated Fischer 344 and Zucker Fatty Rats Fed a High-fat Diet.
Altered Expression of Genes for Lipid Metabolism Vehicle-treated and DS-7250-treated Zucker Fatty Rats Fed a High-fat Diet.

Toxicokinetic Evaluations
Toxicokinetic parameters of DS-7250 are shown in Table 5. The AUC0–24 hr and C max values were comparable between F344 and ZF rats fed an STD. Similar results were obtained in F344 and ZF rats fed an HFD; however, the AUC0–24 hr and Cmax values in both strains fed an HFD were approximately 3.9- to 6.4-fold higher than those in animals fed an STD.
Toxicokinetic Parameters of DS-7250.

Note: Toxicokinetic parameters were evaluated at the end of the dosing period in Fischer 344 (F344) and Zucker fatty (ZF) rats. Data are expressed as mean ± standard deviation. STD = standard diet; HFD = high-fat diet.
Discussion
In the present study, ZF rats aged 12 weeks showed slight hepatic steatosis regardless of feeding conditions; however, this was not observed in F344 rats. It was reported that ZF rats fed an STD spontaneously developed hepatic steatosis at 6 to 12 weeks of age, and its severity was well correlated with the upregulation of genes related to de novo fatty acid synthesis (Fasn, Scd1, and Me1) and fatty acid uptake (Cd36) compared with the corresponding lean control animals (Buque et al. 2010). Our data support this previous report and also reveal that additional genes associated with the uptake and synthesis of fatty acids in the liver were upregulated in ZF rats fed an HFD. Therefore, ZF rats are considered more vulnerable to hepatic lipid accumulation by enhanced lipogenesis and lipid storage compared with F344 rats due to genetic characteristics (Carmiel-Haggai, Cederbaum, and Nieto 2005).
In this study, DS-7250 exacerbated hepatic steatosis in ZF rats, which was more marked under HFD conditions, and significantly increased plasma TG in ZF rats fed an HFD. Other rat strains (i.e., F344 rats or Sprague-Dawley rats) or species (i.e., C57BL/6 mice) do not show similar events with DGAT1 inhibition and HFD conditions (Choi et al. 2007; Villanueva et al. 2009). Villaneuva et al. (2009) reported that the liver-specific knockout of the Dgat1 gene protected against HFD-induced hepatic steatosis due to hepatic uptake of exogenous fatty acids. However, they also suggested that hepatic steatosis was induced through a DGAT1-independent pathway controlled by liver X receptor and a transcriptional factorSREBF1 (Ferre and Foufelle 2010 SREBF1 regulates de novo lipogenic genes such as Fas and Elovl6 (Matsuzaka et al. 2007). Interestingly, DS-7250 increased hepatic mRNA levels of Srebf1, Gpam, Elovl6, and Fasn in ZF rats fed an HFD as compared with the corresponding control rats, indicating that hepatic steatosis in ZF rats was worsened through the increase in hepatic de novo lipogenesis, which was accompanied by the increase in plasma TG. Therefore, DGAT1 inhibition in the present study was not considered protective on hepatic steatosis in ZF rats, unlike the steatosis in Dgat1 -/- mice due to hepatic uptake of exogenous fatty acids. DGAT2, but not DGAT1, is a rate-limiting enzyme for de novo synthesis of TGs in the liver (Zammit 2013). In the present study, Dgat2 gene in DS-7250-treated ZF rats was not clearly upregulated compared to the vehicle-treated ZF rats under HFD conditions. Further studies are necessary to determine how DS-7250 contributes to the acceleration of de novo lipogenesis in the ZF rats under HFD conditions.
ELOVL6 catalyzes the elongation of saturated and monounsaturated fatty acids with 12, 14, and 16 carbons and is the only ELOVL isoform involved in de novo fatty acid synthesis (Shimano 2012; Matsuzaka et al. 2007). Matsuzaka et al. (2012) showed that steatohepatitis in Elov6 -/- mice fed an HFD was attenuated compared to wild type mice, even when those mice possessed the same degree of hepatic steatosis. Although DS-7250 induced marked hepatic steatosis and an increased expression of hepatic Elovl6 in ZF rats fed an HFD, hepatic injuries were not noted. TG synthesis has previously been demonstrated to protect against lipotoxicity by converting fatty acid substrates to a storage form of TGs (Matsuzaka et al. 2012; Li et al. 2009; Yamaguchi et al. 2007). Gene expression changes between vehicle-treated and DS-7250-treated ZF rats fed an HFD were not indicative of obvious inflammatory or oxidative stress responses (data not shown), which would support favorable effects of de novo lipogenesis.
DS-7250 reduced food consumption in ZF rats during the dosing period, and this was more significant under the HFD condition as compared with the STD condition. This effect is supported by the fact that DGAT1 inhibition in DIO mice delayed the intestinal absorption of lipids as reported by Cao et al. (2011). Furthermore, genetic deletion or inhibition of DGAT1 in mice was also shown to cause delayed gastric emptying associated with prolonged increases in incretin hormones, at least in part, resulting in the inhibition of postprandial TG absorption in the small intestine (Ables et al. 2012; Maciejewski et al. 2013). Although the effects of DS-7250 on the gastric emptying rate or postprandial TG absorption were not examined in the present study, these effects of DGAT1 inhibition may be one of the mechanisms for reduced food intake in ZF rats treated with DS-7250. Moreover, the reduced food intake in ZF rats fed an HFD is likely what prevented their body weight gain. It remains unclear whether or not the pharmacological action of DGAT1 inhibitors depends on the fat content per calorie of the administered diet (Cao et al. 2011).
The exposure levels to DS-7250 in F344 and ZF rats fed an HFD were higher than those in both rats fed an STD (3.9- and 6.4-fold higher in F344 and ZF rats, respectively). Previous studies have indicated that a high-fat meal can affect drug absorption, with increased AUC being observed for certain drugs with high lipid solubility including phenytoin, cyclosporine, and menatetrenone (Melander et al. 1979; Tan et al. 1995; Uematsu et al. 1996). DS-7250 shows relatively high lipid solubility (log D 2.8), and given this physicochemical property, DS-7250 was considered to show high exposure levels under an HFD condition. Notably, the exposure levels of DS-7250 were comparable between F344 and ZF rats fed an HFD. These results suggest that the systemic exposure to DS-7250 was not a causative factor for hepatic steatosis in ZF rats.
In summary, the present results indicate that ZF rats were susceptible to hepatic steatosis due to the activation of de novo fatty acid synthesis. Furthermore, DS-7250 administration appeared to accelerate hepatic steatosis, especially under HFD conditions, through the activation of key genes involved in hepatic de novo lipogenesis including Elovl6, Me1, Srebf1, and Fasn, which are not DGAT1 dependent. This phenotype is completely different from that observed in F344 rats and other mice strains (e.g., Dgat1 -/- and DIO mice); therefore, such unexpected results may only be noted in ZF rats, which possess unique genetic and metabolic characteristics.
Footnotes
Acknowledgments
The authors would like to thank Shunsuke Takada for the animal procedures, Takashi Yamaguchi for the clinical pathology, Tetsuya Ohsawa and Shinobu Hakamata for the preparation of histology slides, Yoshiko Oshima for the dose formulation, Miho Matsuoka for toxicokinetics, and Noriyo Niino for GeneChip analysis.
Author Contributions
Authors contributed to conception or design (KY, KKu, YT); data acquisition, analysis, or interpretation (KY, KKu, TI, YT, KKa, KM); drafting the manuscript (KY, KKu); and critically revising the manuscript (TI, YT, KKa, KM). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.