
Abstract
Although the contributions of sitagliptin to endothelial function in diabetes mellitus were previously reported, the potential mechanisms still remain undefined. Our research was intended to explore the underlying mechanisms of protective effects of sitagliptin treatment on endothelial dysfunction in Zucker diabetic fatty (ZDF) rats. Male lean nondiabetic Zucker rats were used as control and male obese ZDF rats were randomly divided into ZDF and ZDF + sitagliptin groups. The significant decrease in endothelium-dependent relaxation induced by acetylcholine was observed in mesenteric arteries and thoracic aorta rings of ZDF rats. The administration of sitagliptin restored the vascular function effectively. The morphology study showed severe endothelial injuries in thoracic aortas of ZDF rats, and sitagliptin treatment attenuated these changes. The increased malondialdehyde levels and decreased superoxide dismutase activities in serum of ZDF rats were reversed by sitagliptin treatment. Sitagliptin also increased the expression of endothelial nitric oxide synthase and microtubule-associated protein 1 light chain 3 (LC3) and decreased the expression of inducible nitric oxide synthase, 3-nitrotyrosine, and p62 in ZDF rats. After giving Fe (III) tetrakis (1-methyl-4-pyridyl) porphyrin pentachloride porphyrin pentachloride (FeTMPyP, a peroxynitrite [ONOO−] scavenger) or sitagliptin to high-glucose (30 mmol/L, 48 hours) cultured human umbilical vein endothelial cells (HUVECs), the increased levels of Beclin-1 and lysosome-associated membrane protein type 2 were detected. Both FeTMPyP and sitagliptin also significantly increased the number of mRFP-GFP-LC3 dots per cell, suggesting that autophagic flux was increased in HUVECs. Our study indicated that sitagliptin treatment can improve the endothelium-dependent relaxation and attenuate the endothelial impairment of ZDF rats. The protective effects of sitagliptin are possibly related to antiperoxynitrite and promoting autophagy.
Introduction
Diabetes mellitus (DM) is a common metabolic disease which is based on increased fasting blood glucose level. 1 The main cause of death in diabetic patients is cardiovascular complications, and endothelial dysfunction is considered as an early event in the process of cardiovascular diseases. 2 However, until now, the underlying mechanisms involved in endothelial dysfunction in diabetes are not yet fully understood. Currently, there are many studies reported that some widely used medications in diabetes can lead to various cardiovascular diseases. 3 Therefore, effective therapeutics to restore vascular function in diabetes is in urgent need.
Quite a few potential mechanisms were reportedly related to endothelial dysfunction in diabetes, in which nitric oxide (NO) production and bioavailability were decreased. It is confirmed that NO has cardiac protective effect and influences vascular functions. 4 However, the increased contents of inducible nitric oxide synthase (iNOS)–derived NO in DM could produce peroxynitrite (ONOO−), 5 resulting in reduced NO bioavailability and bioactivity, as well as endothelial dysfunction, 6 which is the crucial initiation in vascular events of diabetes.
Meanwhile, there are different mechanisms to deal with endothelial dysfunction and to prevent the onset and the development of vascular diseases. One such mechanism is the autophagy, which can degrade damaged organelles by the lysosomal pathway. 7 Recent studies have indicated that in endothelial progenitor cells (EPCs) and vascular endothelial cells, autophagy can be an important means for the cell survival by removing damaged proteins and detritus. 8,9 Autophagy can keep intracellular environment steady and maintain normal cell structure. Inhibition of autophagy can induce the accumulation of mitochondrial reactive oxygen species (ROS), causing initiation of early diabetic nephropathy. 10 Thus, the enhancement of autophagy may be one of therapeutic choices for diabetic vascular complications.
Dipeptidyl peptidase-4 (DPP-4) inhibitors are the most recent available antidiabetic drugs by improving glucagon-like peptide-1 (GLP-1) and glucose-dependent insulinotropic polypeptide metabolism. 11 Sitagliptin, a highly selective DPP-4 inhibitor, can be used alone or in combination with other drugs, such as metformin, for the treatment of type 2 diabetes mellitus (T2DM). 12 Previous studies have provided a strong evidence for cytoprotective properties of sitagliptin in Zucker diabetic fatty (ZDF) rats. 13 Recently, many researches have turned attention to the vascular effects of GLP-1 receptor (GLP-1 R) agonists and DPP-4 inhibitors, and it has been found that GLP-1 R agonists and DPP-4 inhibitors could improve endothelial function in T2DM patients and rats, type 1 diabetic rats, and normoglycemic apolipoprotein-E-deficient mice. 14,15 Sitagliptin also increased circulating EPCs, 16 which participate in repair of vascular endothelial injury. Nevertheless, the effects of sitagliptin on endothelial function in diabetes and its underlying mechanisms remain largely undetermined. The aim of this study was to explore the effects and the potential mechanisms of sitagliptin treatment on endothelial function in ZDF rats, aiming to throw a novel light on the treatment of vascular complications in T2DM.
Materials and Methods
Animals and Cells
This study was performed in accordance with the “Guiding Principles in the Use and Care of Animals” published by the National Institutes of Health (NIH Publication No. 85-23, Revised 1996) and under the supervision of the Institutional Animal Care and Use Committee of Capital Medical University. Experiments carried out in male obese ZDF rats (ZDF/Gmi, fa/fa, 12 weeks old, n = 16) from Vital River Laboratories (Beijing, China). As the control, male lean nondiabetic Zucker rats (ZDF/Gmi, +/+, 12 weeks old, n = 8) were used. In the text, the ZDF/Gmi, fa/fa rats will be designated as ZDF rats, and, when under sitagliptin treatment, as ZDF+ sitagliptin rats. The ZDF/Gmi, +/+ rats will be designated as Lean rats. The rats were fed with Purina 5008 chow ad libitum and had free access to water. ZDF/Gmi, fa/fa rats were randomly divided into 2 groups: ZDF group and ZDF + sitagliptin group administered with 10 mg/kg/d of sitagliptin (Merck & Company, Kenilworth, New Jersey) by gavage for 12 weeks. 17 Lean and ZDF rats were treated with saline as vehicle. On the day of the experiment, animals were weighed and anesthetized with an intraperitoneal injection of 15% chloral hydrate (300 mg/kg). The right common carotid artery was isolated properly and was cut a small opening after the proximal part was clipped by the artery clamp. The PE-50 polyethylene plastic pipe (Becton Dickinson, New York, New Jersey) was inserted and fixed, the artery clamp was removed, and the sensor was connected to RM-6300 physiological grapher (Nihon Kohden, Tokyo, Japan) to record the blood pressure.
Human umbilical vein endothelial cells (HUVECs; CRL-1730; ATCC, Manassas, Virginia) were maintained in DMEM (dulbecco's modified eagle medium)/high-glucose media (Hyclone, Logan, Utah) containing phenol red supplemented with 10% fetal calf serum and were incubated at 37°C with 5% CO2/95% O2. The cells were treated with control (5.5 mmol/L) or high-glucose (30 mmol/L) medium for 48 hours. Besides, FeTMPyP (5 μmol/L; Cayman Chemical, Ann Arbor, Michigan) and sitagliptin (100 μmol/L; Selleck, Houston, Texas) were carried out in high-glucose medium for 48 hours. Both FeTMPyP and sitagliptin were dissolved in phosphate-buffered saline (PBS) just before treatment.
Biochemical Analysis
After blood was collected from abdominal aorta, serum was prepared by centrifuging the blood samples at 1000g for 15 minutes. The levels of serum blood glucose, total cholesterol, and triglycerides were determined by automatic biochemical analyzer (Hitachi 7020, Tokyo, Japan). Serum insulin was measured with rat insulin enzyme-linked immunosorbent assay kit (Crystal Chem, Illinois) using antirat/antimouse insulin antibody and a rat insulin standard. Serum malondialdehyde (MDA) levels and superoxide dismutase (SOD) activities were measured by colorimetry. The experiments were performed using commercially available kits, according to the manufacturer’s instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Measurement of Vascular Function
Measurement of vascular function was carried out in accordance with the previous description. 18 In brief, thoracic aortas and third or fourth branches of mesenteric arteries (2 mm in length) were connected to an isometric force transducer (DMT610 M; Danish Myo Technology, Aarhus, Denmark) and suspended in an organ bath chamber (37°C Hepes buffer containing 95% O2, 5% CO2). After the equilibration, segments of vessel were contracted with norepinephrine (NE, 10−5mol/L). Increasing concentrations (10−9-10−5 mol/L) of acetylcholine (ACh) and nitroprusside (SNP) were added to the chamber to obtain cumulative concentration–response curves after a stable contraction.
Histological Study
After fixed with 4% paraformaldehyde, thoracic aortas were embedded with paraffin, and paraffin section (5 μm) was stained using hematoxylin and eosin. In scanning electron microscopic observation, after cut open and fixed with 3% glutaraldehyde, the thoracic aortas were incubated with 2% OsO4 and dehydrated in ethanol. The aortas were then critical point dried, sputter-coated with gold, and observed using a JSM-6360LV scanning electron microscope (JEOL, Tokyo, Japan).
Flow Cytometry
To explore the effects of sitagliptin on endothelial dysfunction of ZDF rat, a LSRFortessa flow cytometer (Becton Dickinson, New Jersey) was used to evaluate the mobilization of EPCs. One hundred microliter of peripheral blood was incubated with a 4-fold volume red cell lysis on ice for 15 minutes. After centrifugation at 4°C, 450g, for 10 minutes and washing at the same condition, samples were then incubated with FITC (fluorescein isothiocyanate)-conjugated CD34 (sc-18917 FITC, 1:200; Santa Cruz, California) and phycoerythrin-conjugated CD31 (555027, 1:200; BD Biosciences, New Jersey) antibodies. Blank control was performed with PBS. Each analysis included 10 000 events. Circulating EPCs were counted with double positives for CD34 and CD31.
Western Blot Analysis
After isolated and lysed with RIPA (Radio Immunoprecipitation Assay) protein lysis buffer (100 mg/mL; Applygen, Beijing, China), 100 μg protein of thoracic aortas and 30 μg protein of HUVECs were loaded and subjected to 12% SDS-PAGE and then transferred to a polyvinylidene difluoride membrane. After blocking with 5% nonfat milk for 1 hour at room temperature, the blots were probed with the rabbit anti–endothelial nitric oxide synthase (eNOS) polyclonal antibody (ZS-6540, 1:500; ZSGB-BIO, Beijing, China,), the rabbit anti-iNOS polyclonal antibody (ZSGB-BIO), the mouse antinitrotyrosine antibody (#05-233, 1:1000; Millipore, Billerica, Massachusetts, 1:1000), the rabbit anti-p62 antibody (#5114, 1:1000; Cell Signaling Technology, Massachusetts, 1:1000), the rabbit anti-LC3 antibody (Cell Signaling Technology, Boston, Massachusetts), the rabbit anti-Beclin-1 antibody (#3495; Cell Signaling Technology, Boston, Massachusetts), the rabbit anti–lysosome-associated membrane protein type 2 (LAMP-2) antibody (#49067; Cell Signaling Technology, Boston, Massachusetts), or the control rabbit anti-GAPDH (glyceraldehyde-phosphate dehydrogenase) monoclonal antibody (sc-25778, 1:1000; Santa Cruz, California, 1:1000) at 4°C overnight. After washing with TBST (20 mmol/L Tris-base, 137 mmol/L NaCl, and 0.05% Tween-20, pH 7.4) twice, the membranes were then incubated with horseradish peroxidase–conjugated goat antirabbit IgG (immunoglobulin G) (sc-2004, 1:4000; Santa Cruz, California, 1:4000) or goat antimouse IgG (sc-2005, 1:4000; Santa Cruz, California, 1:4000) for 1 hour at room temperature. After washing with TBST 3 times for 15 minutes each, ECL Plus substrate (Thermo Scientific, Massachusetts) was applied to the blot, and images were captured in a gel documentation system. Relative optical density of protein bands was analyzed using gel software Image Lab 3.0.
Immunohistochemical Staining
Briefly, thoracic aortas were removed and stored in 4% paraformaldehyde for 48 hours. Fixed thoracic aortas were dehydrated and embedded in paraffin, and sections were cut into 5 μm thickness and mounted onto glass slides. After the antigen retrieval of the section, endogenous catalase was inactivated with 3% hydrogen peroxide for 10 minutes at room temperature. The slides were then blocked with 5% bovine serum albumin for 30 minutes at room temperature. The sections were stained with the mouse antinitrotyrosine antibody (#05-233, 1:100; Millipore,) at 4°C overnight and peroxidase-conjugated affinipure secondary antibody (PV-9001, 1:3000; ZSGB-BIO, Beijing, China,) at 37°C for 30 minutes, successively. Target proteins were detected with diaminobenzidine and then counterstained with hematoxylin.
Evaluation of Fluorescent LC3 Puncta
Human umbilical vein endothelial cells cultured on coverslips were transduced with Adenovirus tandem fluorescent mRFP (Red Fluorescent Protein)-GFP (Green Fluorescent Protein)-LC3 (Ad-tf-LC3, Hanbio, China) at 20 multiplicity of infection, 24 hours. After treatment, the cells were fixed with 4% paraformaldehyde, mounted and observed with a fluorescence microscope (ZEISS, Imager.A2, Baden-Württemberg, Germany). The number of GFP and mRFP dots was determined by manual counting of fluorescent puncta in random 5 fields from 3 different treated HUVECs.
Statistical Analysis
The results are presented as the mean (SD), where n equals the number of rats. The concentration–response data from isolated mesenteric arteries and thoracic aortas were fitted to a sigmoidal curve using nonlinear regression analysis (Prism version 6.0; GraphPad Software, San Diego, California) to calculate the pEC50. The maximum relaxation (Rmax) to ACh or SNP was determined as a percentage of the NE precontraction. Differences between Lean, ZDF, and ZDF + sitagliptin groups were compared and analyzed using 1-way analysis of variance (ANOVA) followed by Bonferroni post hoc test. Repeated-measures ANOVA followed by Bonferroni post hoc test was used to assess the statistical differences of vascular responses to ACh and SNP among groups. Differences were considered statistically significant at P < .05.
Results
Sitagliptin Improved Blood Glucose and Lipid Metabolic Disturbance in ZDF Rats
The serum levels of blood glucose, insulin, total cholesterol, and triglycerides were higher in the ZDF group than those in the Lean group. Compared with ZDF rats, sitagliptin treatment for 12 weeks significantly reduced the serum levels of blood glucose, insulin, and triglycerides without affecting total cholesterol, although their levels still significantly higher than that of Lean rats. Meanwhile, diastolic and systolic blood pressure was increased in ZDF rats, which was significantly decreased by sitagliptin treatment (Table 1).
Basic Parameters of Rats in Lean, ZDF, and ZDF + Sitagliptin Groups.a

Abbreviations: BW: body weight; BP: blood pressure; SD, standard deviation; Sit, sitagliptin; TG, triglyceride; Total-c, total cholesterol; ZDF, Zucker diabetic fatty.
aResults are shown as mean (SD). Comparisons between groups: ZDF/ZDF vehicle versus Lean/Lean vehicle and ZDF + sitagliptin versus ZDF vehicle.
b P < .05, as compared with the Lean group.
c P < .01 as compared with the Lean group.
d P < .01 as compared with the ZDF group.
e P < .05, as compared with the ZDF group.
f P < .001 as compared with the Lean group.
g P < .001 as compared with the ZDF group.
Sitagliptin Improved Endothelium-Dependent Relaxation of Mesenteric Arteries and Thoracic Aorta Rings in ZDF Rats
In order to explore the possible vascular effects of sitagliptin, we measured the vascular functions of ZDF rats after sitagliptin treatment. Norepinephrine (10−5 mol/L) induced the contraction of isolated mesenteric arteries and thoracic aortas in Lean, ZDF, and ZDF + sitagliptin rats. For both mesenteric arteries and thoracic aortas, the endothelium-independent relaxation induced by SNP (10−9-10−5 mol/L) was not affected in the ZDF group or the sitagliptin treatment group (Figure 1A, B). Conversely, the endothelium-dependent relaxation induced by ACh (10−9-10−5 mol/L) was significantly decreased in ZDF rats (Figure 1C, D). Both the pEC50 and Rmax of mesenteric arteries and thoracic aortas in the ZDF group were significantly decreased comparing to Lean groups. Compared with ZDF groups, sitagliptin treatment significantly enhanced ACh-induced relaxation (Table 2).

Sitagliptin improved endothelium-dependent relaxation of mesenteric arteries and thoracic aorta rings in ZDF rats. Endothelium-independent relaxation to SNP of mesenteric arteries (A) and thoracic aortas (B) rings from Lean, ZDF, and ZDF + sitagliptin groups. Endothelium-dependent relaxation to ACh of mesenteric arteries (C) and thoracic aortas (D) rings from Lean, ZDF, and ZDF + sitagliptin groups. **P < .01, # P < .05 as compared with the Lean group and ZDF group, respectively, n = 6. See Table 2 for pEC50 and Rmax values derived from these data. ACh indicates acetylcholine; SNP, nitroprusside; ZDF, Zucker diabetic fatty.
The Effects of Sitagliptin on Endothelium-Dependent Relaxation of Mesenteric Arteries and Thoracic Aortas from Lean, ZDF, and ZDF + Sitagliptin Groups.a
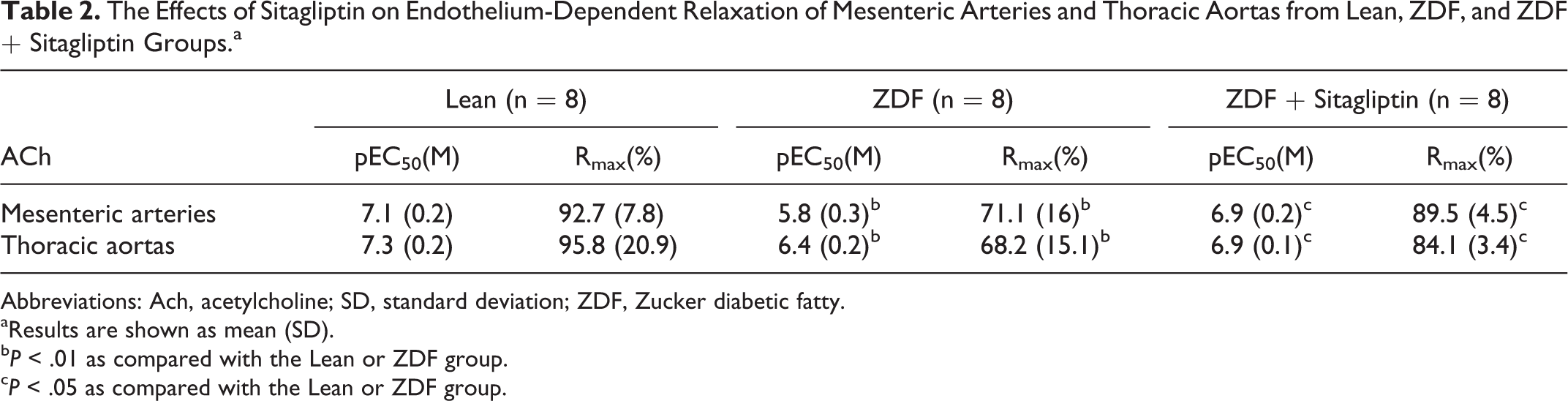
Abbreviations: Ach, acetylcholine; SD, standard deviation; ZDF, Zucker diabetic fatty.
aResults are shown as mean (SD).
b P < .01 as compared with the Lean or ZDF group.
c P < .05 as compared with the Lean or ZDF group.
Sitagliptin Alleviated Damage of Thoracic Aortas in ZDF Rats
Hematoxylin and eosin staining indicated that endothelium has smooth and continuous coverage in the elastic membrane in the Lean group. The endothelium in ZDF rats were severely damaged, with endothelial cells desquamating from the vascular wall. The vascular walls were slightly damaged in ZDF + sitagliptin group with smooth and intact tunica intima (Figure 2A). Besides, the smooth muscle cells among Lean, ZDF, and ZDF + sitagliptin groups were in normal structure, generally. Scanning electron microscopic observation indicated that endothelium was smooth and intact with flat endothelial cells in the Lean group. Endothelial cell injuries were extremely serious with the degenerated and desquamated endothelial cells adherent to the endothelium in the ZDF group. In the ZDF + sitagliptin group, there was slight damage of endothelium with almost intact tunica intima (Figure 2B). The eNOS expression of thoracic aortas was significantly decreased in ZDF rats. Treatment with sitagliptin could reverse this decrease of eNOS expression (Figure 2C).
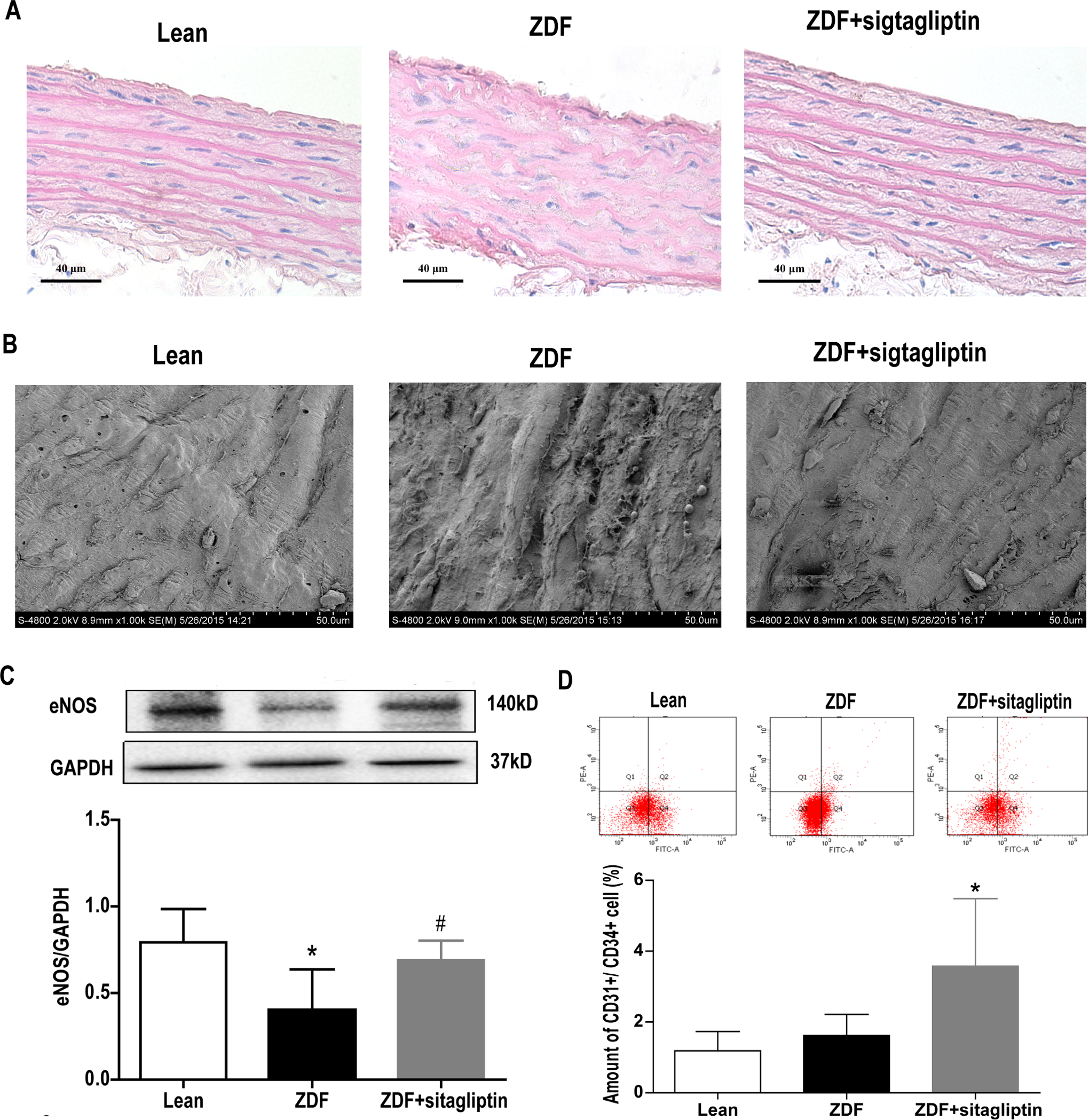
Sitagliptin alleviated damage of thoracic aortas in ZDF rats. A, HE staining (×400). B, Scanning electron microscopic observation (×1000). C, The expression of eNOS in thoracic aortas of Lean, ZDF, and ZDF + sitagliptin groups. *P < .01, # P < .05 as compared with the Lean group and ZDF group, respectively, n = 5. D, Sitagliptin enhanced the mobilization of EPCs in ZDF rats. The mobilization of EPCs (defined as CD31+/CD34+ cells) was determined using flow cytometry. # P < .05 as compared with the ZDF group, n = 8. eNOS indicates endothelial nitric oxide synthase; EPCs, endothelial progenitor cells; ZDF, Zucker diabetic fatty.
To explore the effects of sitagliptin on EPC mobilization in response to vascular endothelium dysfunction, levels of CD31+/CD34+ cells in peripheral blood of ZDF rats were determined using flow cytometry. Zucker diabetic fatty rats had an increased tendency in circulating levels of EPCs than the Lean group, but the differences did not reach statistical significance. Meanwhile, treatment with sitagliptin significantly enhanced the mobilization of EPCs in ZDF rats (Figure 2D).
Sitagliptin Decreased the Level of Nitro-Oxidative Stress in ZDF Rats
To investigate the possible role of nitro-oxidative stress in the protective effects of sitagliptin against endothelial dysfunction in ZDF rats, we observed the effects of sitagliptin on the concentration of MDA, activity of SOD, and the expression of iNOS and 3-nitrotyrosine (3-NT) in the 3 groups.
Compared with the Lean group, the concentration of MDA in serum from ZDF rats was significantly increased with the decreased activity of SOD, indicating the production of free radical was increased while the clearance was decreased (Figure 3A, B). On the other hand, the ZDF + sitagliptin group showed lowered concentration of MDA and elevated activity of SOD, implying the lowered ROS production after sitagliptin treatment.

Sitagliptin decreased the level of nitro-oxidative stress in ZDF rats. Concentration of MDA (A) and activity of SOD (B) in serum from Lean, ZDF, and ZDF + sitagliptin groups. C, Expression of iNOS. D and E, Expression of 3-NT in thoracic aortas of Lean, ZDF, and ZDF + sitagliptin groups. **P < .01, # P < .05 as compared with the Lean or ZDF group, respectively, n = 5. iNOS indicates inducible nitric oxide synthase; MDA, malondialdehyde; 3-NT, 3-nitrotyrosine.
Compared with the Lean group, the expression of iNOS was significantly upregulated in the thoracic aortas of ZDF rats and sitagliptin significantly decreased the expression of iNOS (Figure 3C), implying that sitagliptin may deplete the excessive production of NO.
Hyperglycemia can increase the production of superoxide anion (·O2 −). Although ·O2 − itself is chemically inert, once combines with NO at a diffusion limited rate, it will become a highly reactive species, ONOO−. The formation of 3-NT was usually used for monitoring the generation of ONOO− in vivo. 19 Both immunohistochemistry and Western blot results indicated that the enhanced 3-NT expression in ZDF rats could be attenuated by sitagliptin treatment (Figure 3E, F). All these results implied that inhibition of nitro-oxidative stress might play a role in the protective effect of sitagliptin.
Sitagliptin Promoted Autophagy in Thoracic Aortas of ZDF Rats
To investigate the possible role of autophagy in the protective effects of sitagliptin against endothelial dysfunction in ZDF rats, we observed the effects of sitagliptin on autophagy, including detecting the protein expression of p62 and LC3 in thoracic aortas. Zucker diabetic fatty rats exhibited an increased expression of the autophagic marker, p62 (Figure 4A). Accumulation of p62 in ZDF rats implies a defect in clearance of protein aggregates. The transformation of light chain–associated protein 3 (LC3) from LC3-I to LC3-II by lipidation during the formation of autophagosomes is an indicator of autophagy. 20 This transformation was detected in the ZDF + sitagliptin group but not in the Lean or ZDF group (Figure 4B). These results implied that sitagliptin promoted autophagy in ZDF rats in vivo.

Sitagliptin promoted autophagy in ZDF rats. Expression of p62 (A) and LC3 (B) in thoracic aortas of Lean, ZDF, and ZDF + sitagliptin groups. *P < .05 was compared with the Lean group, # P < .05, and ### P < .001 as compared with the ZDF group, n = 5.
Sitagliptin Promoted Autophagy in High-Glucose Incubated HUVECs
To further test the mechanisms of sitagliptin’s protective effects on endothelial cells, we detected the autophagy level of sitagliptin-treated HUVECs in high-glucose medium. Compared with cells in control medium, HUVECs incubated in high-glucose medium (30 mmol, 48 hours) showed decreased Beclin-1, LAMP-2 and LC3-II/I expression, indicating lowered autophagy level. After treated with sitagliptin, the autophagy level was rescued with increased expression of Beclin-1, LAMP-2 and LC3-II/I (Figure 5A-C). To evaluate autophagic flux in cultured HUVECs, cells were transduced with Ad-tf-LC3. The images where more free red dots than yellow dots indicated the elevated autolysosome formation compared with autophagosomes (Figure 5D), suggesting that sitagliptin increased autophagic flux in high-glucose incubated HUVECs. Moreover, the administration of FeTMPyP showed a similar trend with sitagliptin treatment, indicating the promoted autophagy level might be realized in an ONOO− clearance manner.

Sitagliptin promoted autophagy in high-glucose incubated HUVECs. The HUVECs were incubated in high-glucose medium (30 mmol/L) for 48 hours in the presence or absence of sitagliptin (100 μmol/L) and FeTMPyP (5 μmol/L), respectively. A, B, and C, Expression of Beclin-1, LC3 and LAMP-2 in HUVECs, respectively. D, The HUVECs were transduced Ad-tf-LC3 for 24 hours and treated with the above-mentioned reagents and times. Mean numbers of autophagosomes and autolysosomes were represented by yellow and red dots in merged images per cell, respectively.*P < .05 as compared with HG(−)/FeTMPyP(−)/Sitagliptin(−) treated cells, and #P < .05 as compared with HG(+)/FeTMPyP(−)/Sitagliptin(−) treated cells, n = 5. HG indicates high-glucose; HUVECs, human umbilical vein endothelial cells; LAMP-2, lysosome-associated membrane protein type 2; LC3, light chain 3.
Discussion
Our present study indicated the importance of sitagliptin in improving endothelial dysfunction in ZDF rats. The major novel findings were included as follows: (1) treatment with sitagliptin in vivo can improve endothelium-dependent relaxation of ZDF rats, which possibly related to antiperoxynitrite to restore NO bioavailability; (2) sitagliptin treatment can promote autophagy, which possibly related to accelerating endothelial regeneration and maintaining endothelial integrity.
The leptin receptor-defective ZDF rat is characterized by insulin resistance and hyperglycemia, which is generally considered as a perfect experimental model of progressive development of T2DM. 21 We analyzed the effect of sitagliptin on basic parameters of rats (Table 1). The increased concentrations of blood glucose and insulin in ZDF rats indicated that the T2DM models were duplicated successfully. Furthermore, sitagliptin treatment significantly attenuated the levels of blood glucose and insulin, indicating its powerful effect of antihyperglycemia. Previous studies have ever suggested that sitagliptin is very comprehensive in the treatment of patients with T2DM, and in addition to lowering blood glucose, it also improves blood pressure and lipid profiles. 22 We didn’t observe significant difference in body weight among the 3 groups; however, diastolic and systolic blood pressure was increased in ZDF rats, which was markedly decreased by sitagliptin treatment. About the lipid profiles, the results of our study revealed significant increase in total cholesterol and triglycerides in ZDF rats. Sitagliptin treatment resulted in significant decrease in triglyceride concentration without affecting total cholesterol, which was corresponded with the study by Ferreira et al. 17 A research reported that DPP-4 inhibitors appeared to have an advantageous effect on levels of total cholesterol and triglycerides. 23 However, some researches also indicated that sitagliptin had no effect on total cholesterol and triglyceride levels. 24 The discrepancy in these studies may be due to the differences in duration of treatment time, types of drug administration, as well as experimental models. Until now, effects of sitagliptin, as well as other DPP-4 inhibitors, on the blood lipid profiles are still controversial.
Dipeptidyl peptidase-4 inhibitor, a kind of novel antihyperglycemia agent, has been widely used in recent years. Several reports have shown that sitagliptin confers cytoprotective effects on several tissues. 25 –27 Accumulating evidences also have demonstrated that the impaired endothelial function was generally considered as a preliminary indicator for future cardiovascular complications in T2DM. In present study, the weakened vascular response to ACh (endothelium-dependent relaxation) suggested that T2DM could induce severe endothelial dysfunction, which could be ameliorated in the presence of sitagliptin (Figure 1). Meanwhile, scanning electron microscopic observation also showed severely damaged vascular endothelium in ZDF rats, which could be attenuated by the treatment of sitagliptin (Figure 2). As for the EPCs, when mild injuries occur in endothelium, the bone marrow would be activated to release EPCs to the circulation to play the endothelium-protective roles. 28 Once the injuries became severe and decompensatory, the mobilization was lost or the EPCs were exhausted, then the amount of circulating EPCs was decreased. Therefore, the number of circulating EPCs depends on lots of factors, including the injury degree of the endothelium, the activity or function of the bone marrow, and the mobilization of EPCs. In this study, in ZDF rats, the number of circulating EPCs was a little greater while without significant difference compared to the Lean group, and sitagliptin treatment can improve the circulating EPCs (Figure 2D), which was similar with prior report. 29 In brief, vascular functional and histological studies have shown that sitagliptin treatment could alleviate the endothelial cell damage and improve vascular function in ZDF rats.
In diabetics, the accumulation of superoxide, a potent ROS, depletes NO to produce ONOO−, which binds with tyrosine residues of proteins to form 3-NT. ONOO− has a devastating impact on vascular function, which is a main damaging factor of nitro-oxidative stress on many organs. In our study, there was increased oxidative stress in the tissues of diabetes rats (Figure 3A, B). At the same time, the expression of iNOS was significantly upregulated in the thoracic aortas of the ZDF rats (Figure 3C), implying an excessive NO production. Then NO and ROS may concurrently produce additional tissue damage by formation of ONOO−. 4 Our present study also indicated that the expression of 3-NT (footprint of ONOO− in vivo) was increased in thoracic aortas of ZDF rats (Figure 3D, E), which was suppressed by sitagliptin, denoting that the excessive elevation of nitro-oxidative stress in T2DM rats may moderately be downregulated by means of sitagliptin.
Autophagy can degrade and recycle organelles (such as mitochondria) to maintain cellular homeostasis that is closely involved in the pathological process of diabetes. 30 If the reduction of autophagy sustained a period of time, mitochondrial dysfunction and the accumulation of ROS would result in cell damage and even cell death. Recent reports have suggested that the upregulated autophagy plays a protective role in renal injury in diabetic mice. 31 In our study, the accumulation of p62 in ZDF rats, known as an autophagic substrate, 32 implied a defect in clearance of protein aggregates (Figure 4A), and autophagy could be induced by sitagliptin as the content of LC3-II increased (Figure 4B). To consolidate the involvement of nitro-oxidative stress and autophagy in sitagliptin’s protective effects, we further detected the level of autophagy in high-glucose medium incubated HUVECs. The incubation of high glucose led to a decrease of autophagy level in HUVECs. However, with the treatment of sitagliptin, the expression of Beclin-1, LAMP-2, and LC3-II/I was increased, indicating the rescue of autophagy. At the same time, Ad-tf-LC3 33 was used for evaluating autophagic flux in HUVECs. The results with Ad-tf-LC3 showed elevated autolysosome formation compared with autophagosomes after sitagliptin treatment, indicating the increased autophagic flux in HUVECs, which were in accordance with increased LC3-II and decreased LAMP-2 expression. Besides, treatment with FeTMPyP also increased the level of autophagy, implying that the sitagliptin-induced elevation of autophagy might be realized in an ONOO− clearance manner (Figure 5). The role of autophagy in response to endothelial dysfunction indicates that sitagliptin treatment may be an effective therapeutic measure for vascular complications in T2DM.
As an antihyperglycemia agent, sitagliptin can breakdown DPP-4 activities and then elevate GLP-1 levels. Pancreatic β cells could be activated by GLP-1 to secrete insulin, and the blood glucose would be decreased. 34 Besides its antihyperglycemia effects, sitagliptin also showed beneficial effect toward vascular functions. According to the researcher’s data, GLP-1 itself has direct vasodilatory effects. 35 As reported, the elevated GLP-1 and GLP-1 receptor expression induced by sitagliptin can increase cAMP (cyclic adenosine monophosphate) level, therefore to activate AMPKα/eNOS pathway, then NO bioactivity would be improved and the endothelium-dependent relaxation would be restored. 36 In addition, GLP-1 receptor agonists, exenatide, also exert vasoprotective effects via eNOS/NO signalings. 37 Furthermore, the GLP-1-induced insulin secretion might also be involved in sitagliptin’s vasoprotective effects. It’s reported that insulin itself has direct vasodilatory effects in healthy subject. 38 –40 Insulin can enhance the NO production and eNOS expression to exert vasoprotective effects, 41,42 which can be suppressed by tumor necrosis factor α (TNF-α). 43 Meanwhile, sitagliptin can inhibit inflammation and reduce TNF-α level, 44 which is also a possible mechanism involved in sitagliptin’s vasoprotection. Except the GLP-1-dependent vascular function restoration, we are focusing on the involvement of nitrative stress and autophagy in vasoprotective effects of sitagliptin. In vivo, we observed the changes of nitrative stress and autophagy levels in ZDF rats administered with sitagliptin, and then in cultured endothelial cells, the increased autophagy level in FeTMPyP-treated cells hinted us that sitagliptin-promoted autophagy might be related to the antiperoxynitrite effect. At present, it is not sufficient enough to draw a conclusion that sitagliptin can promote autophagy level through antiperoxynitrite, which just demonstrating that nitrative tress and autophagy were correlated to some extent. In our current study, whether the vasoprotective effect of sitagliptin is lowered glucose dependent or not still cannot be confirmed. However, in some other researches, nondiabetic experimental animal models were constructed to illustrate the antidiabetic independent impact of sitagliptin. For instance, sitagliptin exerts vasoprotective effect in both experimental hypertensive and atherosclerotic animal models. 45,46 Other mechanisms involved in sitagliptin’s protection toward DM-induced vascular injuries were also reported, including decreasing oxidative stress, inactivation of apoptotic signals, and induction of angiogenesis in the development of DM. 47 –50
Besides sitagliptin, some other interventions and agents can also improve DM-induced vascular dysfunction. As reported by Focardi, low-carbohydrate diet can improve endothelium-dependent relaxation via producing H2O2 in a NO-independent way. 51 Rosuvastatin, a kind of statins, can improve vasodilatation by 2 possible mechanisms. One is NO dependent, the other is Ca2+/K+ channel dependent. 52 In our current work, sitagliptin can also promote vascular function by eNOS/NO pathway, however, the impact on Ca2+/K+ channel still needs to be further investigated. Meanwhile, cannabinoids and rosiglitazone can improve diabetes-induced cardiovascular complications. However, the side effects (such as cannabinoids-induced shock or rosiglitazone-caused endothelial dysfunction in humans) were also reported, 53,54 which didn’t occur in the presence of sitagliptin.
Conclusion
In brief, our present study indicates that sitagliptin can improve endothelium-dependent relaxation and attenuate the endothelial impairment of ZDF rats, which are possibly related to antiperoxynitrite and promoting autophagy. This study may provide a novel sight on the management of vascular endothelial dysfunction in DM.
Footnotes
Acknowledgments
The authors thank Dr Qi Pan from Department of Endocrine, Beijing Hospital (Beijing, China) for her support in ZDF rats.
Author Contributions
Wen Wang, Huanyuan Wang, and Yi Zhou contributed to conception and design, contributed to acquisition, analysis, and interpretation, drafted the article, critically revised the article, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy. Huirong Liu, Haixia Huang, and Zhiying Guo contributed to conception and design, contributed to acquisition, analysis, and interpretation, critically revised the article, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy. Yu Dong and Jiahui Xu contributed to acquisition, analysis, and interpretation, critically revised the article, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Natural Science Foundation of China (No 81370450) and Scientific Research Foundation of Beijing Key Laboratory of Metabolic Disorders Related Cardiovascular Diseases (No 2015DXWL03).