
Abstract
High concentrations of hexavalent chromium (Cr(VI)), captan, and folpet induce duodenal tumors in mice. Using standardized tissue collection procedures and diagnostic criteria, we compared the duodenal histopathology in B6C3F1 mice following exposure to these 3 carcinogens to determine whether they share similar histopathological characteristics. B6C3F1 mice (n = 20 per group) were exposed to 180 ppm Cr(VI) in drinking water, 12,000 ppm captan in feed, or 16,000 ppm folpet in feed for 28 days. After 28 days of exposure, villous enterocyte hypertrophy and mild crypt epithelial hyperplasia were observed in all exposed mice. In a subset of mice allowed to recover for 28 days, duodenal samples were generally indistinguishable from those of unexposed mice. Changes in the villi and lack of observable damage to the crypt compartment suggest that toxicity was mediated in the villi, which is consistent with earlier studies on each chemical. These findings indicate that structurally diverse agents can induce similar (and reversible) phenotypic changes in the duodenum. These intestinal carcinogens likely converge on common pathways involving irritation and wounding of the villi leading to crypt regenerative hyperplasia that, under protracted high-dose exposure scenarios, increases the risk of spontaneous mutation and tumorigenesis.
Introduction
In 2007 and 2008, the National Toxicology Program (NTP 2007, 2008) reported toxic and carcinogenic effects in mice and rats following 13 weeks and 2 years of exposure to hexavalent chromium (Cr(VI)) in drinking water . Exposure to Cr(VI) was associated with adenomas and carcinomas of the duodenum and jejunum in B6C3F1 mice (NTP 2008). The only other chemical that has clearly induced duodenal tumors in mice in an NTP study is the fungicide captan (NCI 1977; Stout et al. 2009). Folpet is structurally similar to captan and also induces duodenal tumors (Cohen et al. 2010). Like Cr(VI), neither captan nor folpet induces intestinal tumors in rats (Cohen et al. 2010; Gordon 2007). Despite evidence for in vitro genotoxicity, both captan and folpet are not genotoxic in vivo and appear to increase tumor risk in mice due to chronic mucosal wounding and regenerative hyperplasia (Cohen et al. 2010; Gordon 2007; Arce et al. 2010), and regulators have determined that these organochloride compounds induce intestinal tumors via nonmutagenic (i.e., threshold) mechanisms (United States Environmental Protection Agency [U.S. EPA] 2004; Eastmond 2012; International Programme for Chemical Safety (IPCS) 1995), in which chemically induced villous cytotoxicity and resulting crypt regenerative hyperplasia are nonneoplastic precursors that increase the chance of intestinal carcinogenesis. More recently, some scientists have concluded that Cr(VI) induces intestinal tumors through similar mechanisms (Haney 2015; HealthCanada 2015; TCEQ 2016; Thompson et al. 2013, 2014, 2017).
We have previously highlighted similarities between captan, folpet, and Cr(VI) in the context of 10 factors that regulators use to determine whether a carcinogen acts via a mutagenic or nonmutagenic mode of action (MOA; Thompson, Suh, et al. 2017). For example, all 3 carcinogens are negative for micronucleus induction in duodenal crypts following oral exposure (Arce et al. 2010; Chidiac and Goldberg 1987; O’Brien et al. 2013; Thompson, Wolf, et al. 2015). In addition, studies indicate that these carcinogens primarily affect the committed cells of intestinal villi as opposed to proliferating cells in the crypts (Cohen et al. 2010; Gordon 2007; Gordon, Cohen, and Singh 2012; O’Brien et al. 2013; Thompson et al. 2011; Thompson, Wolf, et al. 2015). To date, however, direct experimental comparison between captan, folpet, and Cr(VI) has been lacking.
Herein, we expose B6C3F1 mice to captan, folpet, or Cr(VI) in the same bioassay to compare histopathologic effects in the duodenum following exposure to carcinogenic doses. Recovery groups are included to assess the reversibility of intestinal lesions, which was previously cited as a data gap in the MOA literature/database for Cr(VI) (Becker et al. 2015). The findings herein, together with existing targeted mechanistic research on Cr(VI), can help risk assessors make informed decisions about the MOA for Cr(VI)-induced intestinal carcinogenesis in mice. More broadly, the findings for Cr(VI), captan, and folpet set the stage for additional research and adverse outcome pathway (AOP) development for intestinal carcinogenesis by structurally diverse carcinogens that act via nongenotoxic mechanisms.
Materials and Methods
Test Substances
Sodium dichromate dihydrate (SDD; 99.95% pure; CAS No. 7789-12-0) was obtained from Sigma-Aldrich Inc. (Milwaukee, WI) and was stored at room temperature and protected from light. Captan (98.3% purity; CAS No. 133-060-2) was obtained from AK Scientific, Inc. (Union City, CA), and folpet (98% purity; CAS No. 133-07-3) was obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). The dose formulations of Cr(VI) in drinking water were prepared weekly at concentrations of 514 mg/L SDD (∼180 ppm Cr(VI)) in tap water. All dose formulations were verified weekly at Eurofins Lancaster Laboratories Environmental (Lancaster, PA). Cr(VI) concentrations were verified analytically by method SW-846 7196A (Online Supplemental Table S1). The detection limit is 0.04 ppm Cr(VI). Dose formulations for both captan and folpet were prepared weekly in diet at concentrations of 6,000 and 16,000 ppm.
Study Design
The in-life portion of this study was conducted at Charles River (Spencerville, OH) using Good Laboratory Practice (GLP) standards. Female B6C3F1 mice were obtained from Charles River (Raleigh, NC). Female mice were selected to better match previous Cr(VI) studies, and because none of the agents demonstrated major differences in male and female mice in terms of effects. The mice were approximately 9 weeks old when they arrived, were acclimated for 16 days, and weighed between 16.4 and 23.1 g at initiation of dosing. NTP-2000 Irradiated Meal Diet (Zeigler Bros., Gardners, PA) was provided ad libitum throughout the study, except during designated procedures. Municipal tap water after treatment by reverse osmosis and ultraviolet irradiation was freely available to the captan- and folpet-treated mice during the dosing and recovery periods. Beginning 2 weeks prior to the study and throughout the dosing and recovery periods, the control and Cr(VI)-treated mice received water or 180 ppm Cr(VI) via water bottles. Animals were housed individually in wire mesh floor cages equipped with an automatic watering valve. Temperatures were maintained at 66°F to 71°F (19–22°C) with a relative humidity of 47% to 52%. A 12-hr light/12-hr dark cycle was used, except when interrupted for designated procedures. Housing and care were as specified in the United States Department of Agriculture Animal Welfare Act (9 CFR, Parts 1, 2, and 3) and as described in the Guide for the Care and Use of Laboratory Animals from the National Research Council. Control group animals were housed on a separate rack from the test substance–treated animals.
The study design is shown in Table 1. Each group contained 20 animals, of which 10 were allowed to recover. Mice were exposed to 180 ppm Cr(VI) in drinking water because it was the highest concentration employed in the NTP (2008) cancer bioassay and previously shown to induce intestinal lesions after 7 and 90 days of exposure. Mice were exposed to 16,000 ppm captan in diet because it was the highest concentration employed in an NCI (1977) cancer bioassay. The same concentration was selected for folpet due to structural similarity to captan. The low concentration for folpet (6,000 ppm) was chosen because it was previously shown to induce hyperplasia in a 28-day mouse study (Gordon, Cohen, and Singh 2012). The same concentration was selected for captan due to structural similarity to folpet. Due to poor health in the high-dose captan group after 1 week of exposure, the high concentration was reduced to 12,000 ppm for captan (see Results section).
Experimental Design.
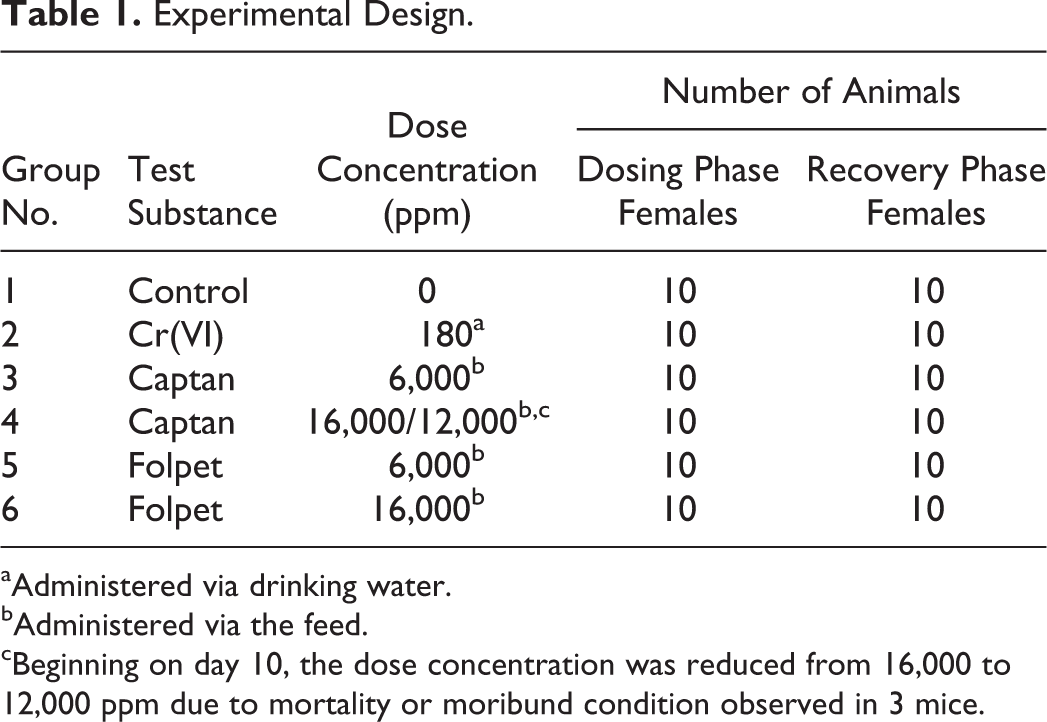
aAdministered via drinking water.
bAdministered via the feed.
cBeginning on day 10, the dose concentration was reduced from 16,000 to 12,000 ppm due to mortality or moribund condition observed in 3 mice.
Standard cage side and clinical observations were made per GLP guidelines. A final bodyweight was obtained on the day of necropsy. Food consumption was measured quantitatively for each animal daily. Water consumption in control and Cr(VI)-treated mice was measured quantitatively for each animal daily.
Histopathology
Mice were euthanized by CO2 asphyxiation, and duodena were excised by transecting the pylorus and proximal jejunum. A small gauge blunt-ended needle was inserted into the distal end of each duodenum to gently flush the specimen with phosphate buffered saline such that buffer slowly leaked from the proximal end. The pylorus was then stapled to small strip of cardboard, and syringe with a blunt-ended needle was used to instill 10% neutral buffered formalin (NBF) from the distal end, thereby gently inflating the intestine to approximately normal size. Hemostats were used to temporarily prevent escape of the formalin until the distal end could be stapled to the card. This process ensures that the specimen remains linear and untwisted (but not stretched). Each specimen (stapled to a card) was then submersed into a container of 10% NBF. After ∼24 to 48 hr, 10% NBF was exchanged with fresh 10% NBF. The duodenum samples were then sent to Experimental Pathologies Laboratory (Sterling, VA) for processing.
Three segments of duodenum were obtained at gross trimming, and these were designated as proximal, middle, and distal, respectively. These segments, each of which was 3-4 mm long, were acquired at the following distances from the still attached gastric pylorus: proximal 1 cm, middle 4 cm, and distal 7 cm. Following gross trimming, the lumen of each segment was filled with HistoGel™ to help maintain lumen patency and villus integrity (Joiner and Spangler 2012). All 3 duodenal segments were embedded in the same block and oriented for transverse sectioning, anterior end down, in a manner that ensured the identities of the proximal, middle, and distal segments were maintained. Each block was faced, following which 3 serial sections were obtained (each section contained proximal, middle, and distal segment profiles). Sections were placed on glass slides with the proximal segment profile located closest to the slide label and the distal segment profile located furthest. One section was stained with hematoxylin and eosin, whereas the other 2 serial sections were retained for future use. Slides were mounted and coverslipped as per routine.
All slides were evaluated using bright-field microscopy by an American College of Veterinary Pathologists board–certified veterinary pathologist (J.C.W.). To our knowledge, captan and folpet have not previously been studied concurrently, and thus, a consistent nomenclature has not been applied to these structurally related compounds. Given the reported similar phenotypic responses in the duodenum following treatment with captan, folpet, and Cr(VI), we applied nomenclature previously used to describe effects of Cr(VI) in the small intestine (Cullen et al. 2016) to all 3 compounds. These effects included villus histiocytic cellular infiltrates, villus atrophy/blunting, villous enterocyte vacuolation, villus single-cell necrosis, and crypt epithelial hyperplasia. Treatment-associated findings observed in the present study additionally included villous enterocyte hypertrophy, increased villous enterocytes, mononuclear cell infiltrates, and rarefaction of the villus lamina propria. Each duodenal segment (i.e., proximal, middle, and distal) was scored independently. The following scale was used to grade the severity of findings: grade 1 = minimal, grade 2 = mild, grade 3 = moderate, and grade 4 = severe. Findings were recorded into, and tabulated by, an electronic pathology data recording system.
Ki-67 Immunohistochemistry
Unstained duodenal sections (see above) from the dosing and recovery phases were subjected to immunohistochemical staining and evaluation. For antigen retrieval, the sections were incubated in 1:20 citrate buffer in a decloaking chamber for 5 min under pressure, followed by 10 min of depressurization. Sections were then stained for Ki-67 antigen with primary rabbit monoclonal antibody (Thermo Scientific, Fremont, CA, RM-9106) at 1:750 dilution and secondary goat anti-rabbit antibody (Vector, Burlingame, CA, BA-6100) at 1:500 dilution using routine avidin–biotin complex methodology (Vector, Burlingame, CA, PK-6100) and a diaminobenzidine chromagen (Biocare Medical, Concord, CA, BDB2004H).
Gastrointestinal Lumen Content Analysis
To maximize use of the experimental animals, lumen contents from the glandular stomach, small intestine (jejunum and ileum), and large intestine were collected analyzed for Cr(VI) via ion chromatography (U.S. EPA SW-847 Method 7199). This assay and related findings are described in Online Supplemental Material.
Results
General Health
Two mice exposed to 16,000 ppm captan died during the first few days of exposure, and a third mouse was euthanized in moribund condition. Therefore, the captan dose was reduced to 12,000 ppm on day 10 of the study; no unscheduled deaths occurred thereafter. Four mice in the 12,000 ppm captan group exhibited signs of dehydration and hunched posture until day 28. Two mice exposed to 6,000 ppm captan, and one mouse exposed to 180 ppm Cr(VI) appeared thin. No clinical signs observed during the recovery phase were considered treatment related.
On day 28 of the study, the mean bodyweight in the Cr(VI)-treated mice was reduced significantly (5.7%; p < .05) relative to control mice. This is likely explained by the reduction in water intake (see below). Mean bodyweights in all other treatment groups did not differ significantly from controls.
Food and Water Intake
Mean food intake in the mice exposed to 6,000 ppm captan and 6,000 ppm folpet did not differ from controls. Mice in all other treatment groups generally exhibited sporadically reduced feed intake as compared to control mice. Mean water intake in the Cr(VI)-treated mice was significantly (p = .001) lower than controls by ∼25% during the 28-day exposure period. This decrease has been observed in previous Cr(VI) studies (NTP 2007, 2008; Thompson et al. 2011) and is likely due to poor palatability. Interestingly, during the recovery phase, mice that had been exposed to Cr(VI) continued to consume ∼25% less water (p < .0001) than untreated mice in the recovery phase. This may have been due to learned behavior, as mean bodyweights did not differ significantly between these 2 groups during the recovery phase.
Histopathology after Dosing Phase
Treatment-related findings in duodena at the end of the dosing phase (day 28) tended to be more prominent in the proximal segments as compared to the middle or distal duodenal segments. Results for the proximal duodenum are shown in Table 2 (results for middle and distal segments are shown in Online Supplemental Table S2). Mice exhibited crypt epithelial hyperplasia (minimal to mild), villous enterocyte hypertrophy (minimal to moderate), increased villous enterocytes (minimal to moderate), histiocytic cells in the villus lamina propria (minimal to mild), increased mononuclear cell infiltrates in villi (minimal to mild), and rarefaction of the lamina propria (minimal to mild). Except for increased mononuclear cell infiltrates in the duodenal villi of some control mice, all findings in mice killed at day 28 were exclusive to treated animals. No grade 4 (severe) effects were observed in any animals.
Prevalence and Severity of Findings in Proximal Duodenum (day 28).
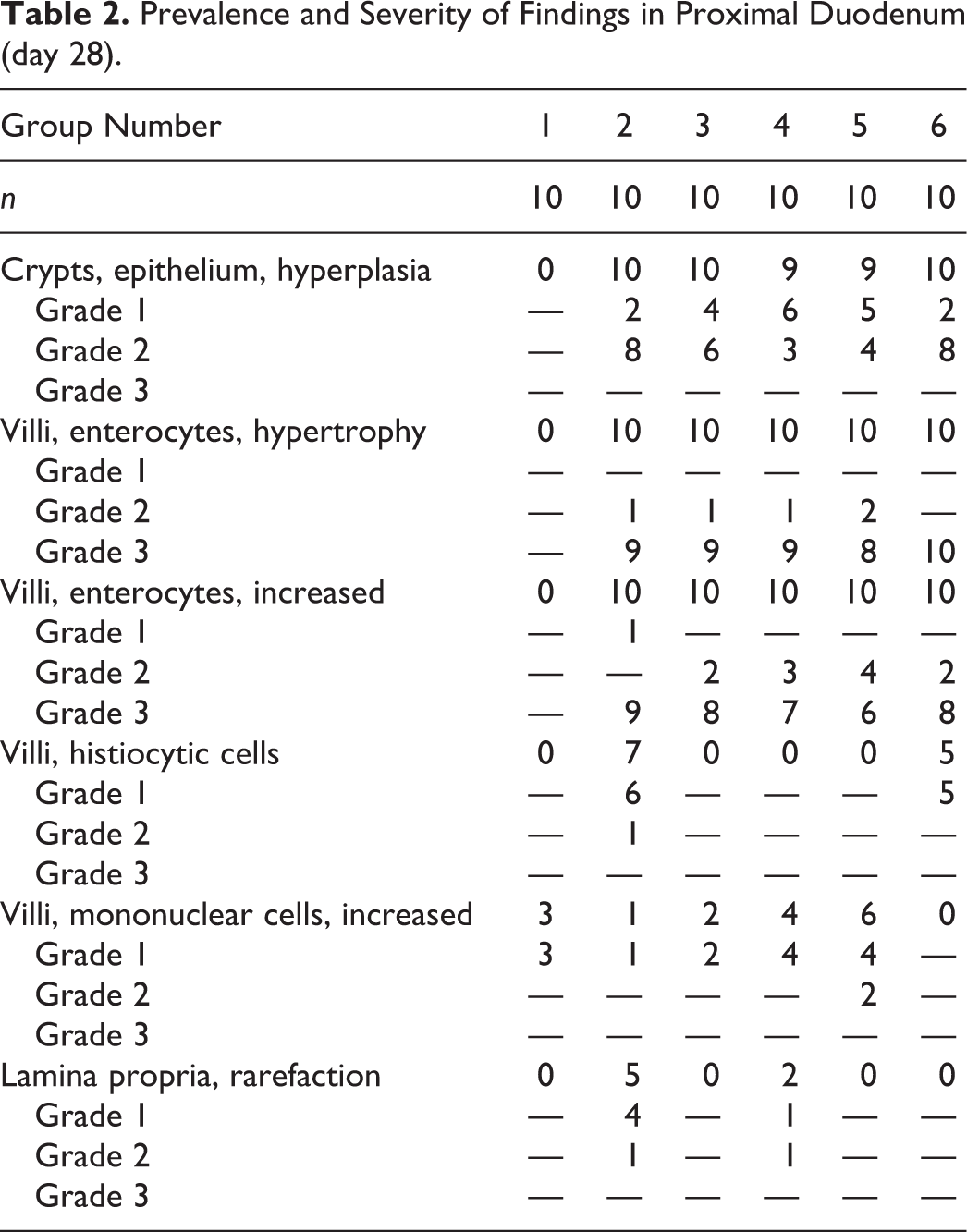
Histiocytic cellular infiltrates in villi occurred primarily in Cr(VI) and 16,000 ppm folpet mice. These infiltrates primarily occurred near the villus tips. Rarefaction of the lamina propria (possibly related to edema) occurred predominantly in Cr(VI)-treated mice. All other changes were qualitatively similar among treated mice, with only minor variations in prevalence and/or severity at a given level of duodenum. In control mice, duodenal villi were most frequently lined by a simple columnar mucosal epithelium, in which the pattern of enterocyte nuclei along the length of villi was only slightly staggered (Figures 1A and 2A). In contrast, the profoundly thickened villi of treated mice (Figures 1B–D and 2B–D) were lined by an extensively pseudostratified epithelium, with nuclei that were heavily crowded and piled up (increased villous enterocytes). However, villous epithelial cells did not display the characteristic appearance of crypt epithelial cells (i.e., cells with dense, hyperchromatic nuclei), and unlike crypts, mitotic figures were not evident in villous epithelial cells. Affected enterocytes generally had excessive amounts of amphophilic apical cytoplasm (villous enterocyte hypertrophy). In most instances, the degree of crypt epithelial hyperplasia, characterized by slight elongation of crypts, increased nuclear chromatin, increased cytoplasmic basophilia, and increased mitotic figures, was modest by comparison. Another relatively minor component of the treatment response was increased mononuclear cell infiltrates (plasma cells and lymphocytes) in villi, the prevalence and severity of which were greatest in the proximal segment of mice exposed to 6,000 ppm folpet.
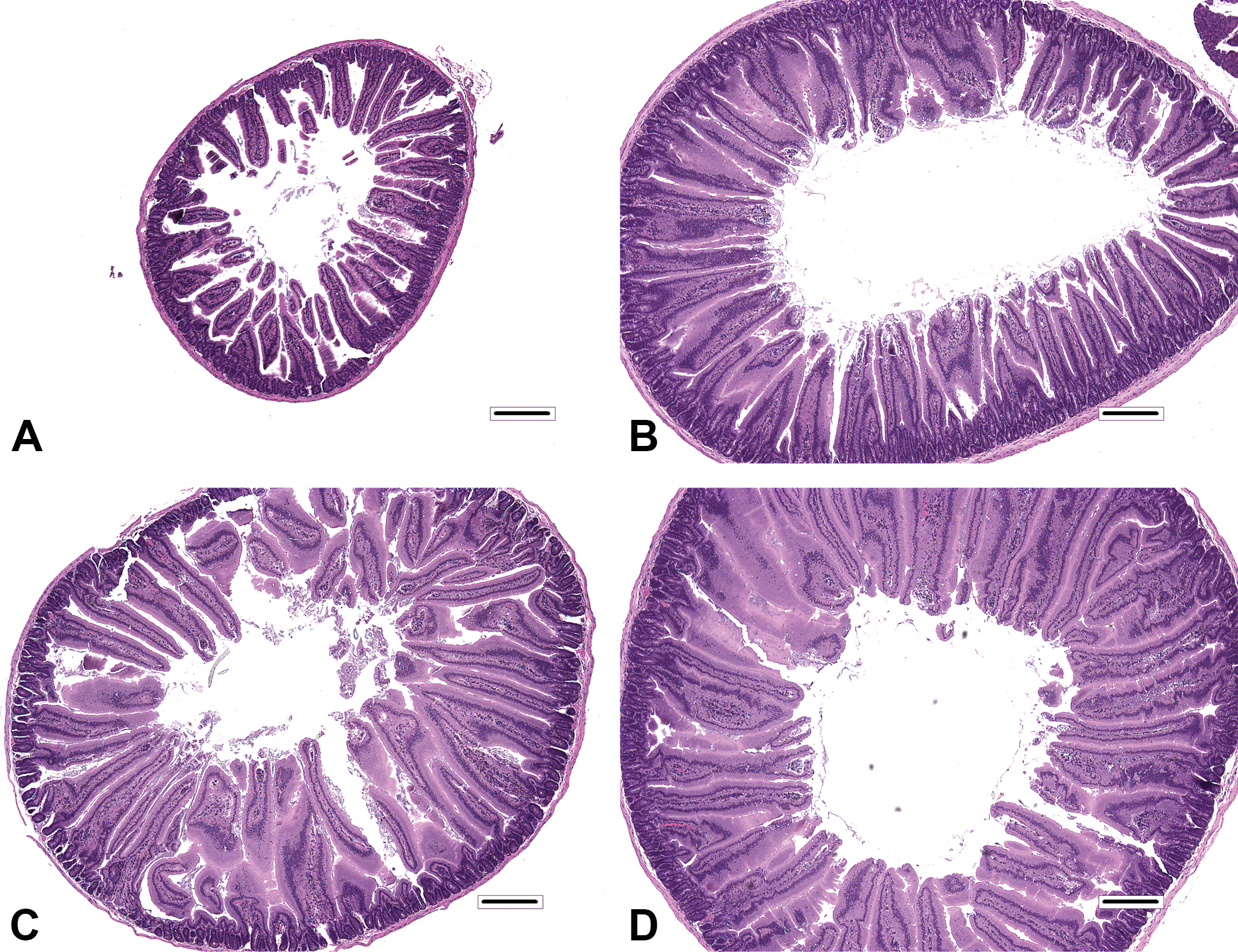
Histopathologic findings in mice at the end of dosing phase. (A) Proximal segment of duodenum from control mouse. Note the amount of separation between villi. (B) Proximal segment of duodenum from 180 ppm hexavalent chromium mouse. Note the thickening of villi evident at this magnification, which is attributable to increased villous enterocytes and villous enterocyte hypertrophy. As a consequence of villus thickening, the duodenal profile is also enlarged. (C) Proximal segment of duodenum from 12,000 ppm captan mouse. Changes are similar to those in B. (D) Proximal segment of duodenum from 16,000 ppm folpet mouse. Changes are similar to those in B and C. Original objective 4x. Bar = 250 μm.
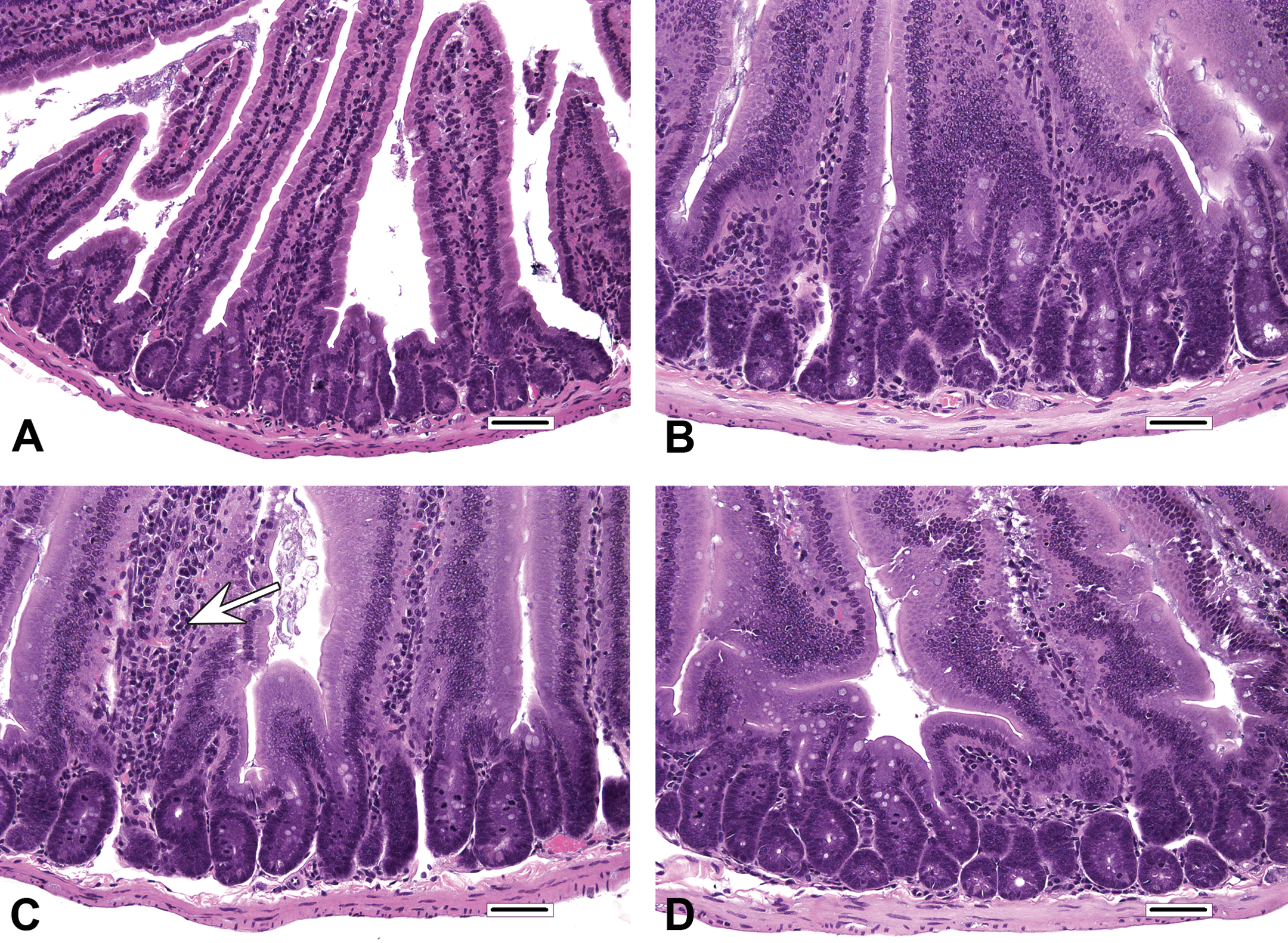
Histopathologic findings in mice at the end of dosing phase. (A) Higher magnification of proximal segment of duodenum from control mouse. Villi are covered by a single-layered epithelium, in which nuclei are only slightly staggered. Villous enterocytes have expected amounts of apical cytoplasm. (B) High magnification of proximal segment of duodenum from 180 ppm hexavalent chromium mouse. The villous epithelium is moderately pseudostratified, and villous enterocytes have abundant apical cytoplasm. Relative to the changes in the villi, the degree of proliferative hyperplastic response evident in the crypts is modest and is characterized by slight elongation, increased nuclear chromatin, increased cytoplasmic basophilia, and increased mitotic figures. (C) High magnification of proximal segment of duodenum from 12,000 ppm captan mouse. The proliferative changes in the villi and crypts are similar to those in B. There is a minimal increase in mononuclear cell infiltrates in one villus (arrow). (D) High magnification of proximal segment of duodenum from 16,000 ppm folpet mouse. Findings of villous enterocyte hypertrophy and increased villous enterocytes are qualitatively comparable to those of duodena illustrated in B and C. Original objective 40x. Bar = 50 μm.
Histopathology after Recovery Phase
Treatment-related findings in the proximal duodenal segment at the end of the recovery phase are shown in Table 3 (results for middle and distal segments of the duodena are shown in Online Supplemental Table S3). The duodenal epithelium of recovery phase control mice tended to have a more proliferative appearance as compared to that of the dosing phase control mice (Figure 3A–D). Therefore, all findings in the duodena of recovery phase mice (both control and treated; Table 3) were scored relative to the appearance of those same diagnoses in dosing phase control mice. The prevalence and severity of findings in treated mice of the recovery phase were generally less than those of treated mice from the dosing phase (e.g., see Cr(VI) in Figure 3E and F). With minor variation, the prevalence and severity of crypt epithelial hyperplasia, villous enterocyte hypertrophy, and increased villous enterocytes were generally comparable among control and treated mice of recovery sacrifice. Findings of histiocytes and mononuclear cell infiltrates in villi tended to persist in Cr(VI)-treated mice.
Prevalence and Severity of Findings in Proximal Duodenum (Day 56).
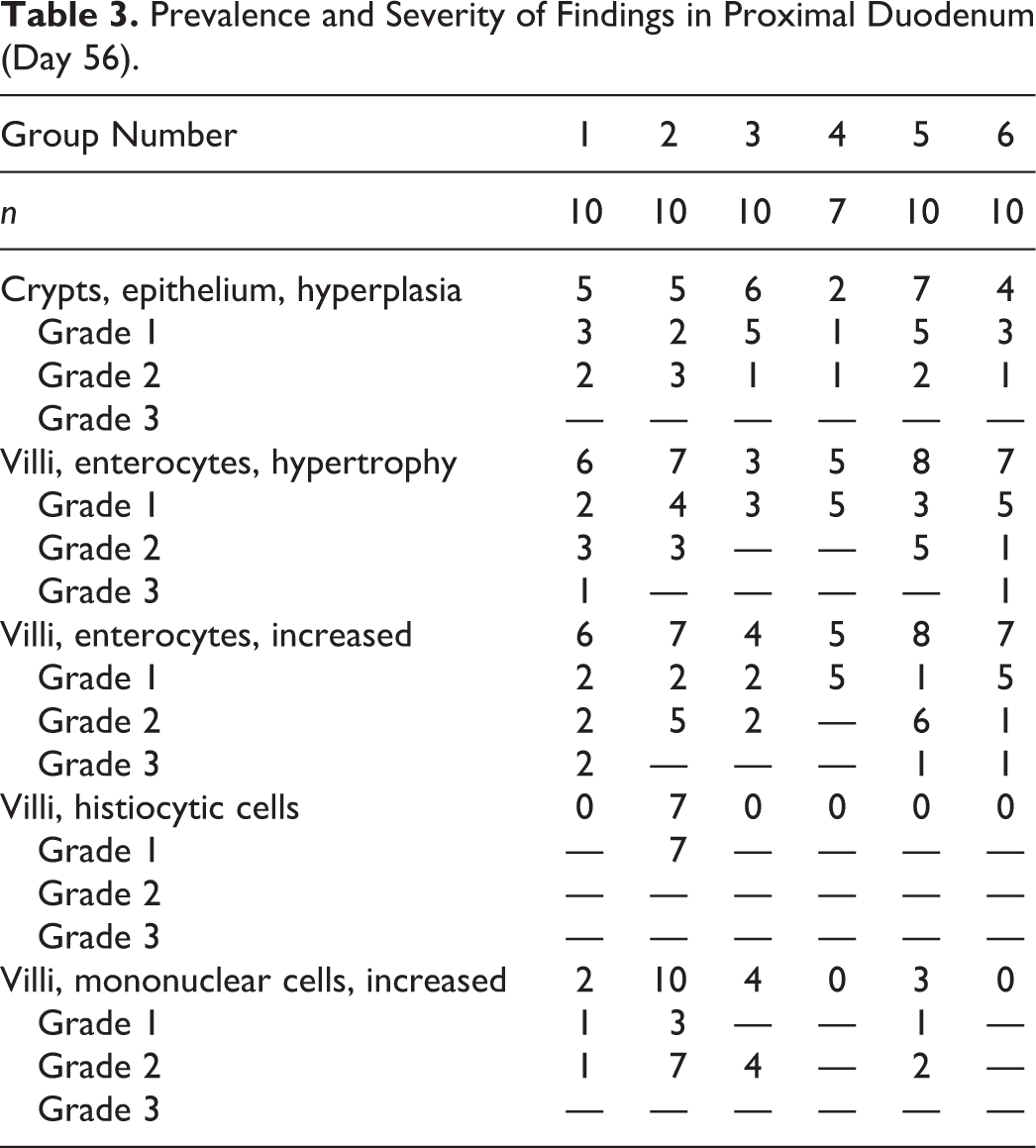
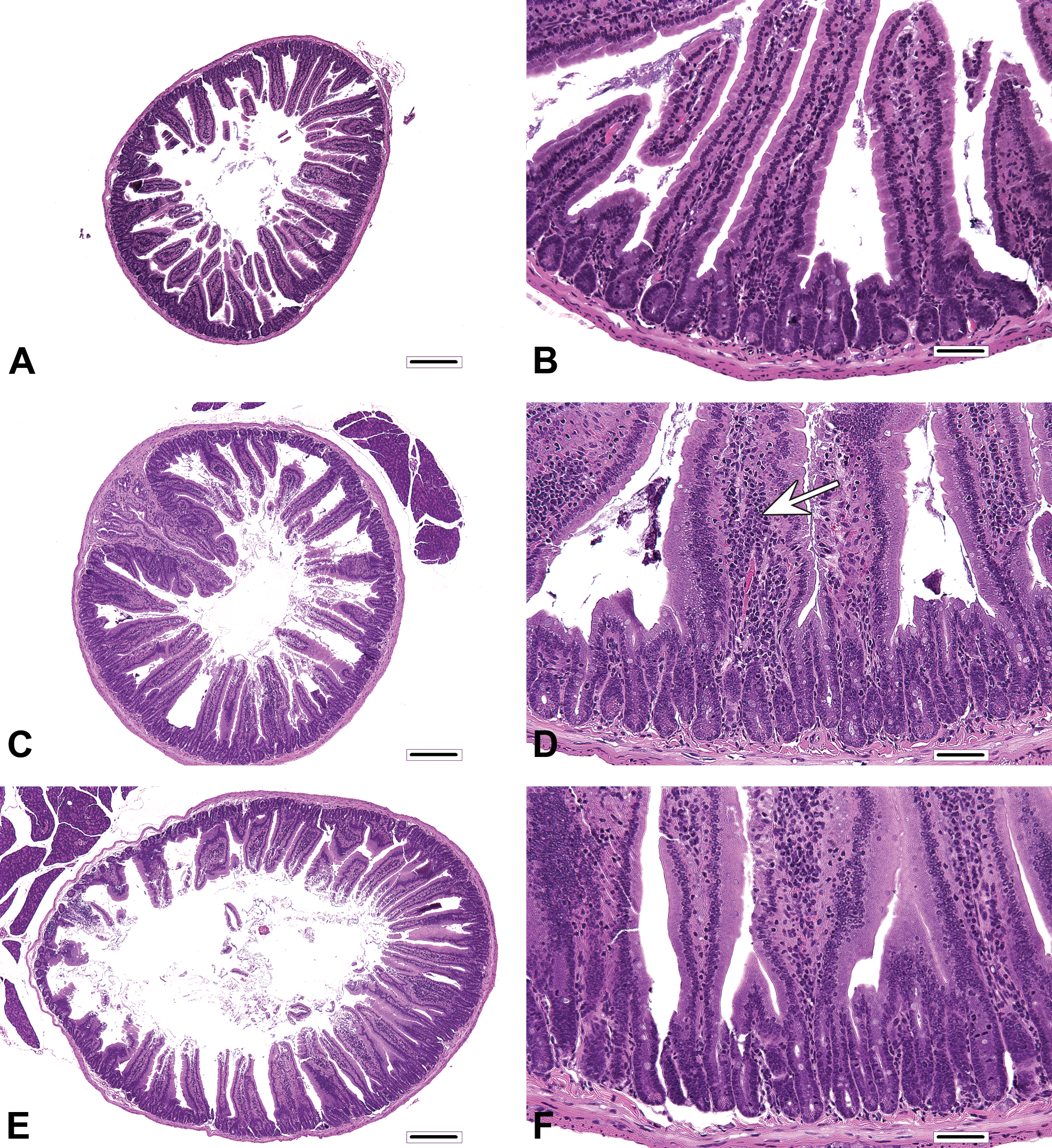
Comparison of histopathologic findings in mice of dosing and recovery phase. (A) Proximal segment of duodenum from control mouse of dosing phase (same as Fig 1A). (B) Higher magnification of A (same as Fig. 2A). (C) Proximal segment of duodenum from control mouse from recovery phase. Villi and crypts in some mice appear more cellular than those in A. (D) Higher magnification of C. Findings include mild villous enterocyte hypertrophy, mildly increased villous enterocytes, mild crypt hyperplasia, and minimally increased mononuclear cell infiltrates in villi (arrow). (E) Proximal segment of duodenum from 180 ppm hexavalent chromium mice from recovery phase. (F) Higher magnification of E. A, C, and E: bar = 250 μm, original objective 4x; B, D, and F: bar = 50 μm, original objective 40x.
Overall Comparison of Histopathology
Visual comparisons of the severity scoring across the treatment groups during the dosing and recovery phases are shown as heat maps in Figure 4. During the dosing phase, mice from all treatments had similar scores for crypt hyperplasia, villus hypertrophy, and increased villous cells in the proximal, middle, and distal duodenum. Rarefaction was similar for Cr(VI) and high-dose captan mice, whereas villus histiocytic cellular infiltration was similar for Cr(VI) and high-dose folpet mice (in the proximal duodenum). During the recovery phase, the severity scores for most lesions were similar in treated and recovery control mice in all segments of the duodenum (Figure 4). Comparison of heat maps between the dosing and recovery phases clearly indicates a lessening of lesion severity among treated mice killed postrecovery.
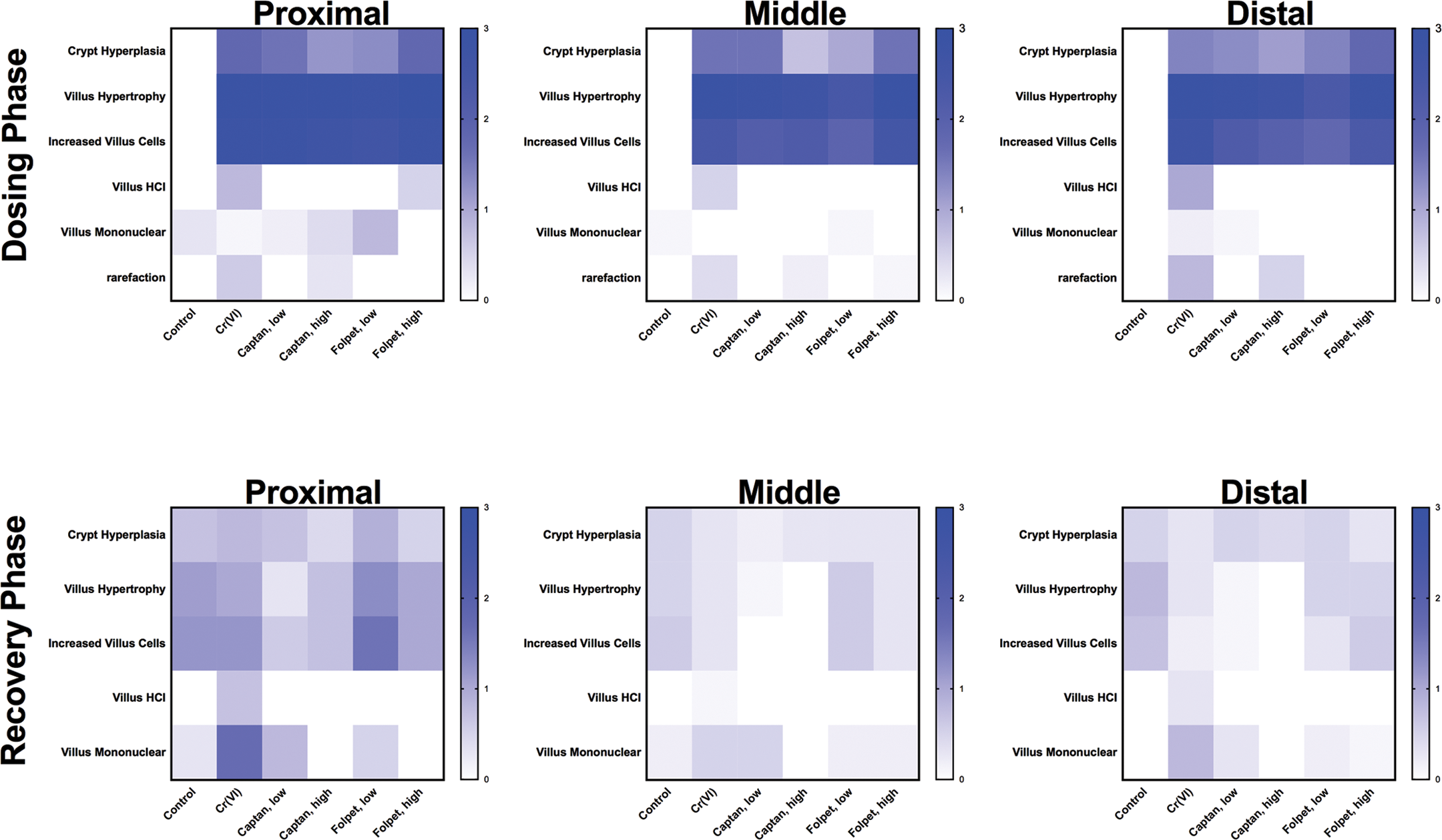
Heat map summaries of pathology responses. The scales represent average severity scores across lesions and treatment groups.
Ki-67 Immunohistochemistry
The distribution of Ki67 staining was found to be qualitatively similar in the duodena of treated and control mice. In all animals, Ki-67 staining occurred primarily in crypt epithelial cell nuclei, where virtually all enterocytes (other than Paneth cells) were positive. A relatively small frequency of positive immunostaining was also present in the nuclei of proximal villous enterocytes. Ki-67 immunostaining further highlighted a slight expansion of the crypt compartment in treated mice relative to controls (Figure 5). In the recovery phase, Ki-67 immunostaining was also comparable between treated and control mice (data not shown).
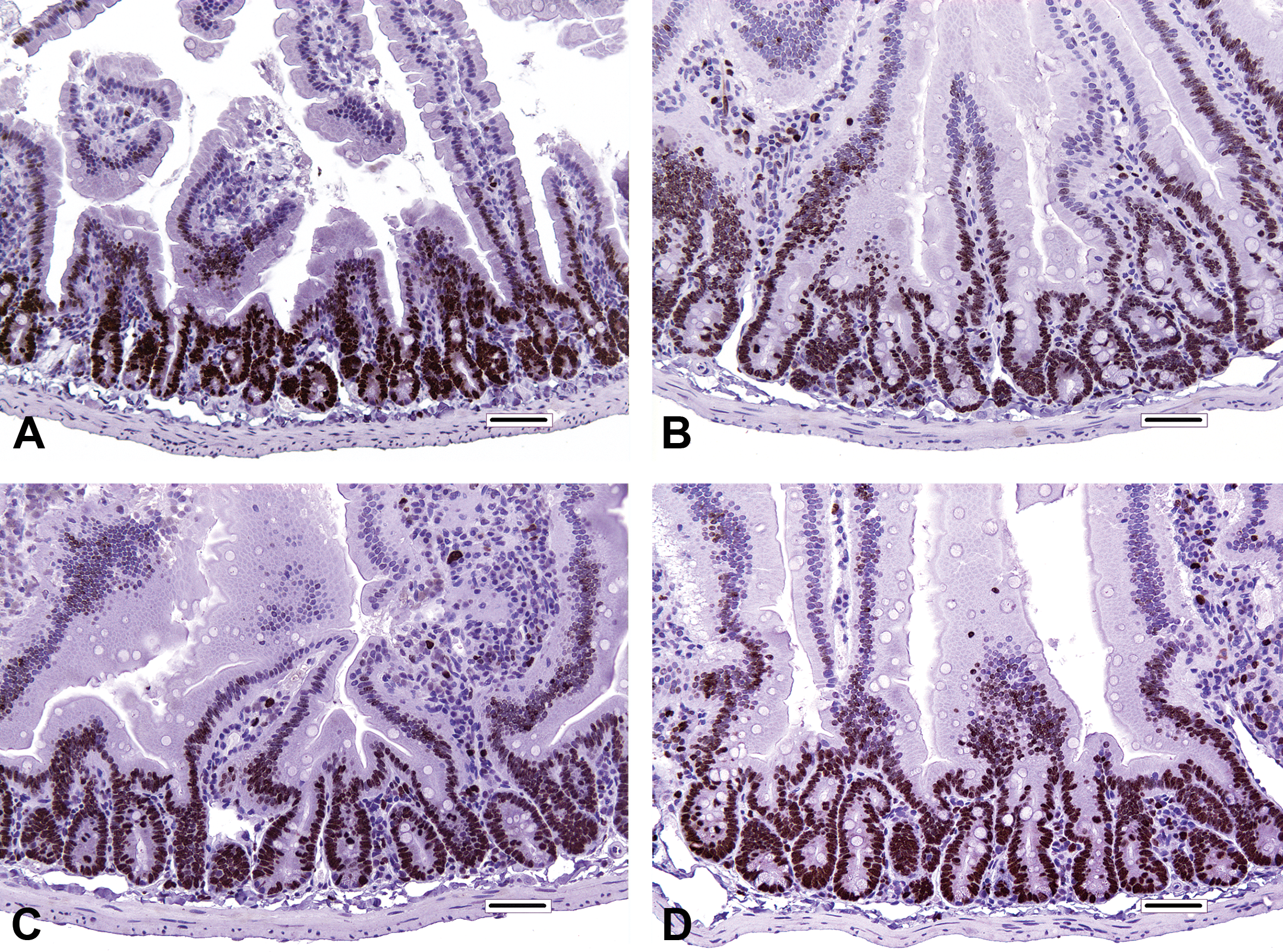
Ki-67 immunostaining in the duodenum from mice of the dosing phase. (A) Proximal segment of duodenum from control mouse. (B) Proximal segment of duodenum from 180 ppm hexavalent chromium mouse. (C) Proximal segment of duodenum from 12,000 ppm captan mouse. (D) Proximal segment of duodenum from 16,000 ppm folpet mouse. Bar = 50 μm.
Discussion
Some regulatory authorities have concluded that captan, folpet, and Cr(VI) induce intestinal tumors in mice via chronic mucosal toxicity and regenerative hyperplasia (U.S. EPA 2004; Eastmond 2012; IPCS 1995; HealthCanada 2015; TCEQ 2016). These MOA determinations were based on multifaceted research efforts conducted for each agent (Cohen et al. 2010; Gordon 2007; Thompson et al. 2013). We report for the first time the results of a study in which direct histopathological comparisons of these 3 intestinal carcinogens were assessed using uniform diagnostic criteria and terminology. The major treatment-related findings in this study were villous enterocyte hypertrophy, increased villous enterocytes, and crypt hyperplasia. Qualitatively, these findings were similar among the 3 test articles, which suggest that these agents share a common AOP for intestinal carcinogenesis.
Interestingly, the effects observed in the present study appear milder than in the previous studies. For example, exposure of mice to 6,000 ppm folpet was previously reported to induce marked crypt hyperplasia in 23/24 mice (Gordon, Cohen, and Singh 2012), whereas mice exposed to 6,000 ppm folpet in the present study exhibited milder crypt hyperplasia. Moreover, increasing the dose to 16,000 ppm did not dramatically increase the effects (see Table 2). Mice exposed in previous studies to 180 ppm Cr(VI) exhibited more marked crypt hyperplasia as well as evidence for villus atrophy/blunting, villous single cell necrosis, and villous enterocyte vacuolation after both 7 and 90 days of exposure (Thompson et al. 2011; Thompson, Wolf, et al. 2015). The villous enterocyte hypertrophy observed in the current study appears consistent with the diagnosis of villous hypertrophy (widening of villi due to elongation of columnar epithelial cells) reported previously in mice exposed to these agents (Gordon, Cohen, and Singh 2012; NTP 2007, 2008).
There are several potential explanations for the disparate findings in this and prior Cr(VI) studies. Although mice in the current study were obtained from the same vendor as several previous studies, it is conceivable that genetic drift in the mice or their microbiomes may have affected their sensitivity to Cr(VI). Another important difference between this and previous Cr(VI) studies is that the earlier experiments used NTP 2000 “wafers” (i.e., pellets; NTP 2007, 2008; O’Brien et al. 2013; Thompson et al. 2011, 2012; Thompson, Seiter, et al. 2015a), whereas the current study used NTP 2000 meal to accommodate the captan and folpet dietary feed preparations. Digesta formed from consumption of the flour-like meal diet might be less abrasive to the intestinal mucosa than digesta formed from consumption of pellets. It is reported that digesta containing abrasive grasses and sedges can induce villus damage (e.g., blunting) in rodents (Wieczorek et al. 2015) and that digesta with lower water content might also be more abrasive to the intestinal mucosa (DeSesso et al. 2000; Greaves 2012).
Differences between meal and wafer feeds may also affect the dosimetry of Cr(VI), as the increased surface area of meal allows greater bioaccessibility of reducing agents present in feed to reduce Cr(VI) to Cr(III). In addition, the aqueous and solid phases of digesta likely differ between meal and wafer diets, leading to perhaps more rapid and greater gastric emptying of the aqueous phase in wafer-fed mice than in meal-fed mice. The increased residence time of Cr(VI) in the low-pH environment of the stomach would allow for more reduction in Cr(VI). Consequently, relative to mice in previous Cr(VI) studies, mice consuming the meal diet in the present study likely had lower Cr(VI) exposure in the intestine due to increased feed-mediated reduction in Cr(VI) to Cr(III) and slower gastric emptying.
During the recovery phase of the current study, the predominant changes of villous enterocyte hypertrophy and increased villous enterocytes were comparable among control and treated mice and were less severe than changes observed during the dosing phase, which suggests that effects were reversible following test article discontinuation. A previous study reported less than 50% recovery at 2 weeks following exposure to 6,000 ppm folpet in the diet for 4 weeks (Gordon, Cohen, and Singh 2012). The apparent persistence of histiocytes in villi and increased mononuclear cell infiltrates in villi into the recovery phase are not surprising, and it may be the case that these lingering inflammatory cell responses would also subside given further time to recover. For Cr(VI), X-ray fluorescence microscopy indicates the presence of chromium in the villus lamina propria (Thompson, Wolf, et al. 2015), perhaps relating to macrophage sequestration of chromium or free iron displaced by chromium. Histiocytic infiltration is often seen as a component of “accumulation enteropathies” (Gopinath, Prentice, and Lewis 1987), and such infiltrates may persist long after the original insult has subsided. Finally, the intestinal epithelium of control mice in the recovery sacrifice appeared generally more proliferative than that of the main sacrifice. Similar effects were reported in an earlier folpet study in which a higher incidence of villus hypertrophy was observed in recovery controls as compared to dosing controls (Gordon, Cohen, and Singh 2012). There appears to be no obvious reason for this morphologic difference between dosing and recovery controls.
The findings herein have particular relevance for Cr(VI). First, inclusion of a recovery phase in the present study addresses a perceived data gap in the MOA for Cr(VI), that is, the absence of demonstrated recovery from Cr(VI)-induced intestinal lesions (Becker et al. 2015). The reversibility of effects observed in the current study further establishes that the intestinal lesions are not caused by dysregulated cell growth due to transformation, which is consistent with the lack of in vivo genotoxicity that has been reported following Cr(VI) ingestion (De Flora, Iltcheva, and Balansky 2006; De Flora et al. 2008; NTP 2007; O’Brien et al. 2013; Thompson, Seiter, et al. 2015; Thompson, Wolf, et al. 2015, Thompson, Young, et al. 2015, 2017). Second, an earlier analysis (Eastmond 2012) identified 10 factors that influence regulatory determinations of whether a chemical causes cancer through a mutagenic or nonmutagenic MOA. In that analysis, captan and folpet were discussed as examples of 2 carcinogens that were determined to act through threshold mechanisms, whereas Cr(VI) was presented as an example of a carcinogen thought to have a mutagenic MOA. Using those same 10 factors, we recently concluded that the MOA for Cr(VI) was nonmutagenic and further highlighted several similarities between Cr(VI), captan, and folpet (Thompson, Suh, et al. 2017). The histopathologic results of the current study indicate that all 3 agents caused very similar phenotypic responses, and such results support an AOP for mouse small intestinal cancer that does not require carcinogens to be direct mutagens. Additional research is needed to further explore mechanistic similarities (or dissimilarities) between these agents. Development of this type of AOP in contradistinction to an AOP involving direct chemical-mediated crypt stem cell mutation could be beneficial to future risk evaluations for intestinal carcinogens.
Footnotes
Authors’ Note
ToxStrategies is a consulting firm providing services to private and public organizations on toxicology and risk assessment issues. CT, MS, DP, LH, CK, and MH have presented Cr(VI) research findings in meetings with regulators including public meetings.
Acknowledgments
We thank Rock Vitale and Erin Rodgers of Environmental Standards, Inc., for assistance with analytical methods for Cr(VI), and Nancy Harris of EPL, Inc., for performing the Ki-67 IHC.
Author Contributions
Authors contributed to conception or design (CT, MH, DP, LH, and JW), data acquisition, analysis, or interpretation (JW, CT, AM, MH, MS, and CK); drafting the manuscript (CT and JW); and critically revising the manuscript (MS, AM, CK, DP, LH, and MH). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Cr(VI) Panel of the American Chemistry Council.
Supplemental Material
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.