
Abstract
It has been long established that not only the species but also the strain and supplier of rodents used in preclinical safety studies can have a significant impact on the outcome of studies due to variability in their genetic background and thus spontaneous pathologic findings. In addition, local husbandry, housing, and other environmental conditions may have effects on the development and expression of comorbidities, particularly in longer-term or chronic studies. More recently, similar effects related to the source, including genetic and environmental variability, have been recognized in cynomolgus macaques (Macaca fascicularis). The increased use of cynomolgus macaques from various sources of captive-bred animals (including nonnative, U.S./European Union-based breeding facilities or colonies) can affect study design and study results and outcome. It is important to acknowledge and understand the impact of this variability on the results and interpretation of research studies. This review includes recent examples where variability of preclinical animal models (rats and monkeys) affected the postmortem observations highlighting its relevance to study design or interpretation in safety studies.
Keywords
Introduction
It has been known for many years that the traditional rodent models (rats and mice) have considerable variability in their background pathology and responses to xenobiotics dependent on strain, source, time, and laboratory setting. As such, it is widely understood that historical control data for anatomic pathology end points in rodents are most reliable if it is as proximal to the study as possible, the best controls being the individual study control animals (Keenan et al. 2009). What has been less well documented or appreciated until recently is the considerable variability observed in cynomolgus macaques (Macaca fascicularis), not only as individuals but also dependent on their source and geographic background (Stevison and Kohn 2008, 2009). For a long time the cynomolgus macaque was considered by the biopharmaceutical research community to be relatively homogenous in their background observations and responses to xenobiotics, regardless of the source or supplier. Thus, animals from different geographic sources and/or suppliers/holding facilities were intermixed within programs and even within studies, either by the research facility or by the supplier themselves. This practice has resulted in some difficulties when trying to interpret the relevance and meaning of data produced within and between studies when using this species for nonclinical research. While this review covers the sources of variability within 2 nonclinical species, the rat and monkey, the same principles apply to mice and dogs. Examples of background variability observed in anatomic pathology end points as well as responses to xenobiotic administration, which resulted in challenges for pharmaceutical research and development, will be presented.
Sources of Variability
For all species, there are multiple sources of variability in anatomic pathology end points in nonclinical studies, ranging from genetic to environmental factors (Figure 1). In addition, it is now known there is a dynamic interaction between the genotype of an animal and the environment, both current and previous, through epigenetic mechanisms and microbiome alterations, which add significantly to the complexity involved (Barros and Offenbacher 2009; Jacobs and Braun 2014; Jostins et al. 2012).
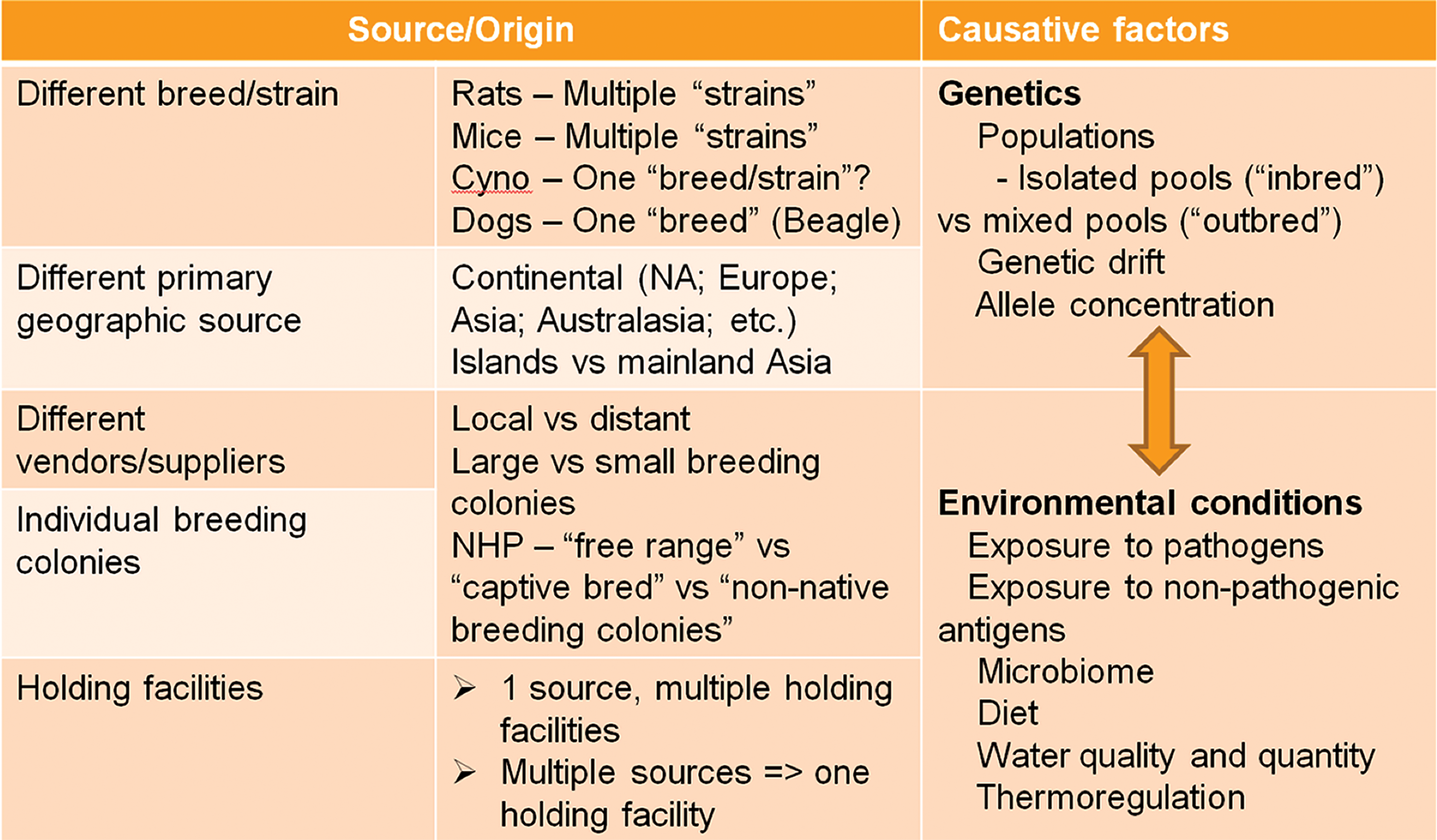
Overview of variables involved in the study design, results, and interpretation of nonclinical studies.
Rats
Most rats used in nonclinical research are derived from 1 species: Rattus norvegicus. However, there are many strains and sub-strains available, all with different phenotypes both in outward appearance and in physiology. There are many publications and online resources available outlining the phenotypic and physiologic differences and similarities between the various strains, and 3 useful resources are included in the bibliography (Cavigelli, Michael, and Ragan 2013; Heidrich 2000; Johnson 2012).
Researchers and suppliers often describe different strains of rodents as “outbred,” “inbred,” “mutant,” and “transgenic.” The various types are defined as follows: Outbred—deliberate choice of “unrelated” mating pairs to maintain heterozygosity within the colony.
Strains include: Sprague-Dawley (SD), Wistar (Wi), and Long-Evans.
Inbred—derived from a very small number of closely related animals. Often genetically tested to ensure the stock maintains genetic uniformity.
Strains include: Fischer 344 (F344), Lewis (LEW), and Brown Norway.
Mutant—naturally derived mutated strains (spontaneous mutations). These are often derived from SD strains/stocks.
Strains include Zucker rat, Zucker diabetic fatty rat), and spontaneously hypertensive rat.
Genetically engineered—(also known as transgenic, knock-in or knock-out) derived through intentional DNA manipulation to produce desired mutant qualities, including humanization of certain genes. Again these are often derived from SD strains/stocks.
Strains include RIP-HAT™ rat, HIP™ rat (both are humanized diabetic models).
It is important to realize that there are no truly outbred strains, however, since all breeding colonies are derived from a relatively small number of originating pairs. Outbred strains do have greater heterozygosity within their strain at a single breeding facility than the other 3 subtypes, although they are likely to have greater genetic drift within, and variability between, different breeding colonies due to the different allele concentrations and/or losses occurring at each breeding site over time. In all breeding facilities, regardless of the type of strain, there is the constant challenge of genetic drift over time, which is frequently evaluated these days through the use of routine genetic testing and restocking by breeders to minimize its effects.
In addition to the genetic variables between, and within, the different strains of rat, the environmental conditions under which the rats are bred, raised, and subsequently utilized at the individual research facility can affect the expression or development of various end points on any given study. Such factors can include diet, housing facilities (specific pathogen free vs. nonbarriered facilities), and individual cage environment (group vs. individual housing; enrichment practices; bedding). All these variable parameters can have a notable impact on the physiology and pathobiology of the animals, resulting in differential background lesions and/or responses to treatment with xenobiotics (Castelhano-Carlos and Baumans 2009; Wolf and Mann 2005).
Cynomolgus Macaques
The cynomolgus macaque is the most frequently used NHP species in biopharmaceutical research and can originate from a very wide range of locations throughout India, China, and Southeast Asia, including the distant island of Mauritius located in the Indian Ocean just east of the African continent. Increased demand for cynomolgus macaques for use in biomedical research has significantly affected wild-caught and native captive-bred populations and, thus, the breeding/sourcing practices for monkeys destined for biomedical research. For many years, it has been considered that different species of macaque monkeys used in nonclinical toxicology studies such as the cynomolgus (M. fascicularis) and the rhesus (Macaca mulatta) macaques were clearly distinct species with different behavioral and physiologic characteristics that impact their response to disease susceptibility and biomedical study outcome. However, what is now increasingly appreciated is that cynomolgus macaques, as a species, have marked genetic diversity, depending on primary origin, with notable overlap between rhesus and cynomolgus genotypes (and phenotype) in the so-called mainland cynomolgus monkeys (Kanthaswamy et al. 2008). More recently, it has become apparent that there is a greater genetic diversity between cynomolgus macaques originating from different geographic regions than between populations of cynomolgus and rhesus monkeys in some areas of mainland Asia (Kanthaswamy et al. 2013; Ferguson and Smith 2015). Furthermore, dedicated breeding colonies in China, the United States and/or the European Union (EU) lead to predictable decreases in genetic diversity and increased gene concentrations within each of the individual colonies as a consequence of unavoidable restrictions of the gene pool based on the founding animals, as has been observed in the naturally sequestered Mauritian cynomolgus macaques (Kanthaswamy et al. 2013; Krebs et al. 2005), while separation of these captive-bred breeding colonies into smaller, isolated enclaves results in increased divergence of the genotype of the animals between the different breeding facilities, even if derived originally from the same geographic location.
Lastly, but by no means least, environmental factors including early or later exposure to pathogenic or nonpathogenic infectious agents, microbiome, dietary, and other local ecosystem variability (such as ambient temperatures, water source, indoor/outdoor access), and effects of social hierarchies in group-housed animals can all contribute to the ultimate response of an animal to any given stimulus or a xenobiotic. Animals from the same original breeding colony but housed in varying holding colonies, or sourced from different vendors and mixed in one holding facility can have variable background incidences of, and exposures to, bacterial, viral, parasitic, and fungal agents, resulting in different susceptibility to pathogens and/or innate immune responses once placed together on study (Mansfield, Sasseville, and Westmoreland 2014; Sasseville and Mansfield 2010; Sasseville and Diters 2008; Sasseville et al. 2012; Saravanan, Sasseville, and Mansfield 2015).
Thus, the increased use of animals from various sources and/or suppliers of captive-bred animals (including nonnative, U.S./EU-based breeding facilities or colonies, and multiple holding facilities) due to current demand has had a significant impact on background observations, immunologic status, and responses to xenobiotics within and between studies.
Background Values and Data
Rats
There are many publications and private databases (e.g., registry of industrial toxicology) outlining the various different background data pertinent to the common strains of rat used in biomedical research. While genetic differences between the strains are usually considered the primary driver for these differences, as discussed earlier the environment has an equally important role. As a result of this complicated dynamic, individual breeding colonies or suppliers can have quite different historical background data, which can also change over time. Such references or databases can include incidences or prevalence of observations in untreated animals as well as trends in incidences over time. Differences in the onset of sexual maturity (Campion et al. 2013) and more recent focus on the onset of reproductive senescence between strains are also an important variable with the potential to impact the results and interpretations of nonclinical studies in rats. The multitude of general references and texts covering background histopathologic data pertaining to strains or sub-strains will not be reviewed in this manuscript nor included in the bibliography since they are too numerous, are updated or superseded rather frequently, may be included in other manuscripts in this publication, and can all be found easily online through the respective breeding companies, contract research organizations, or through literature searches using the strains and/or suppliers of interest. A few specific examples are provided to illustrate the variability in nonneoplastic (Table 1) and neoplastic (Figure 2) histopathologic end points between and within strains (Bellhorn, Korte, and Abrutyn 1988; Hashimoto et al. 2013; King-Herbert and Thayer 2006). Chadwick and colleagues demonstrated nicely the effects of strain (including disease models) and diet on the background incidence of histopathological findings, and select changes are summarized in Table 2 for illustrative purposes (Chadwick et al. 2014). Less commonly available, but equally important, are differences in growth curves and organ weight data between strains. Okamura and colleagues published a nice comparison between WI and SD rats in Japan (Okamura et al. 2011) and more recently Charles River Preclinical Services, Inc. (Montreal) provided data at the annual meetings of the Society of Toxicology (2015) and The American College of Toxicology (2012), showing differences in body weight growth curves and tumor incidences between Wister and SD rats (Figures 3 and 4). Although not anatomic pathology observations per se, inter- and intrastrain differences in metabolic enzyme activities and immune system physiology/biology can lead to considerable variation in the outcome when xenobiotics are administered in the biopharmaceutical setting and, therefore, are important to recognize and understand (Denucci et al. 2010; Sasaki et al. 2006; Reyes et al. 2006, 2004). Table 3 shows the relative susceptibility of strains to urinary tract bacterial infection as a reflection of the difference in innate immunity and/or immunologic competence between the strains evaluated with F344 rats most susceptible to infection and Wi and LEW rats least susceptible, with SD in between, suggesting a more robust immune response in Wi and LEW rats when compared to SD or F344 rats.
Inter- and Intrastrain Variability in Incidence of Corneal Dystrophy in Rats.
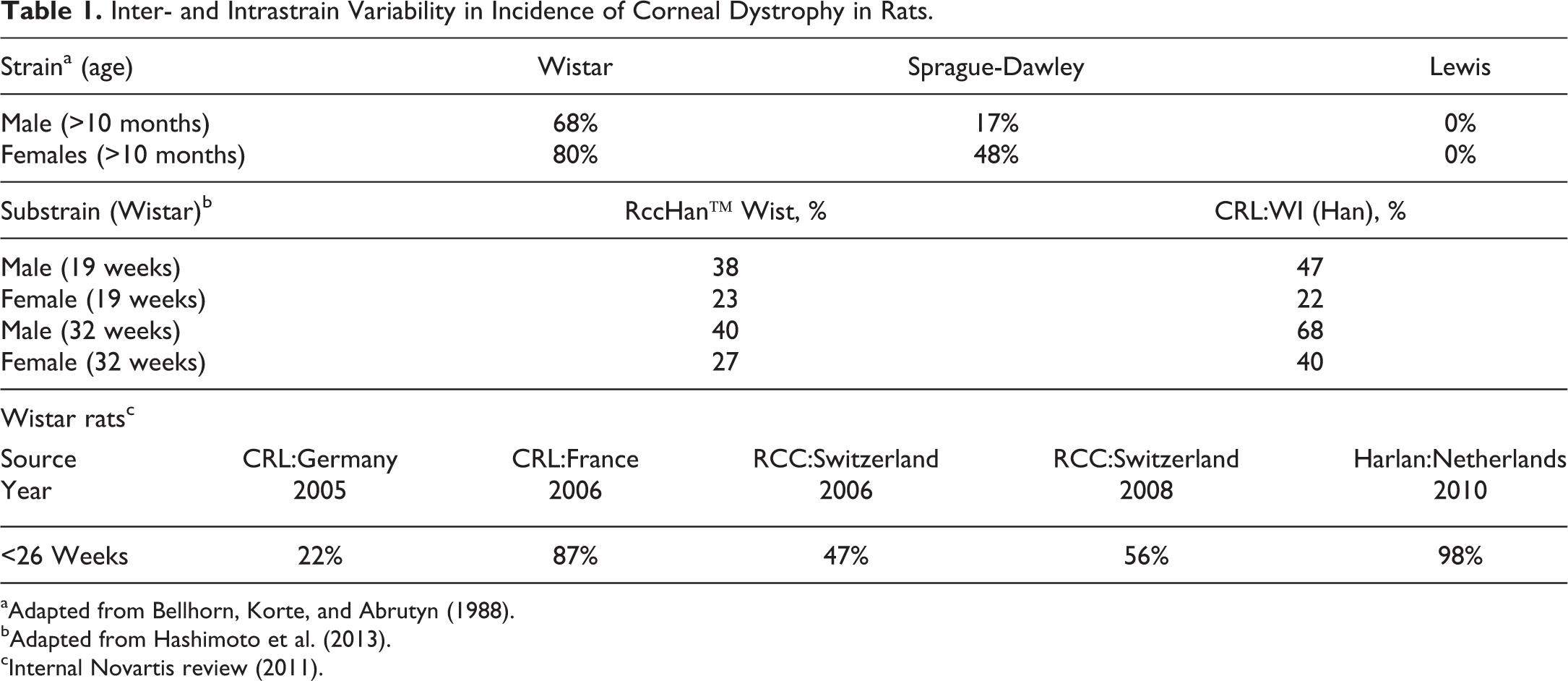
aAdapted from Bellhorn, Korte, and Abrutyn (1988).
bAdapted from Hashimoto et al. (2013).
cInternal Novartis review (2011).
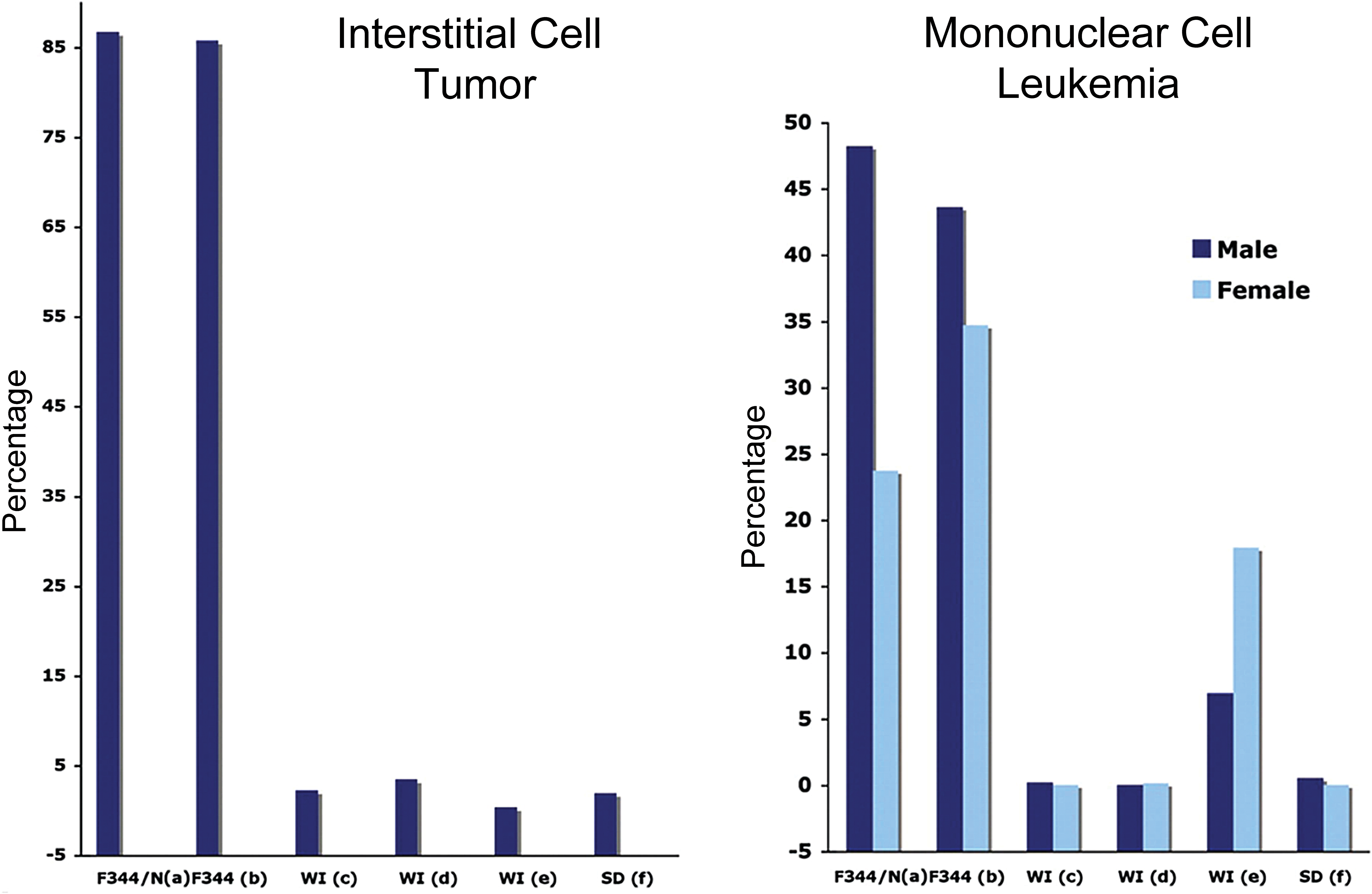
Incidences of interstitial cell tumors and mononuclear cell leukemia in F344, Wistar (Wi), and Sprague-Dawley (SD) rats from various sources. Data presented at the National Toxicology Program (NTP) workshop (http://ntp.niehs.nih.gov; see meetings and workshop). (a) NTP F344/N, (b) National Center for Toxicological Research (NCTR) F344, (c) Wi-Han Charles River Laboratories (CRL) data, (d) Wi-Han RCC data, (e) Wi proprietary data, and (f) SD proprietary data. Reproduced with permission from King-Herbert and Thayer 2006. Copyright © by Society of Toxicologic Pathology.
Incidence Table of Exocrine Pancreatic Inflammation—Strain and Diet Effects.
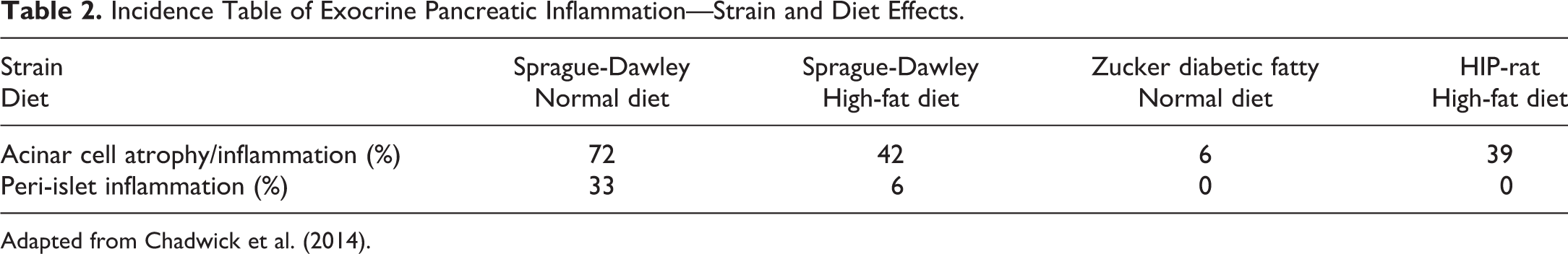
Adapted from Chadwick et al. (2014).
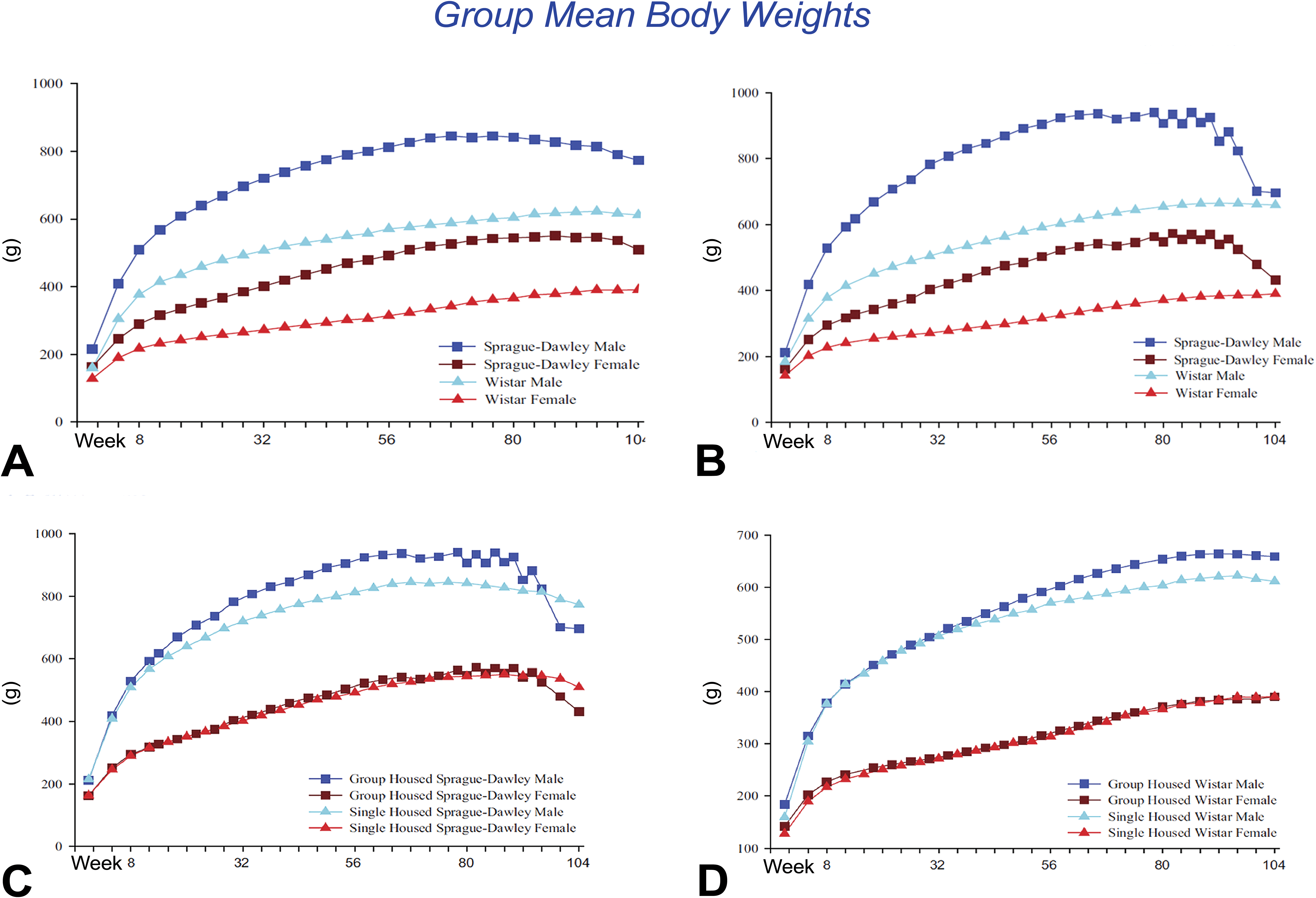
Comparison of body weight data in 2 strains of rat and in 2 different housing paradigms. (A) Individually housed rats: Sprague-Dawley (SD) versus Wistar (Wi). (B) Group-housed rats: SD versus Wi. (C) SD rats: individual versus group housing. (D) Wi rats: individual versus group housing. A clear difference can be seen for the weight curves between strains. No difference was observed between the 2 different caging paradigms for Wi rats, while some disparity may be evident with the SD rats. Data and graphs were provided by Charles River Preclinical Services Montreal Inc. and were originally presented in a poster presentation at the American College of Toxicology (2012).
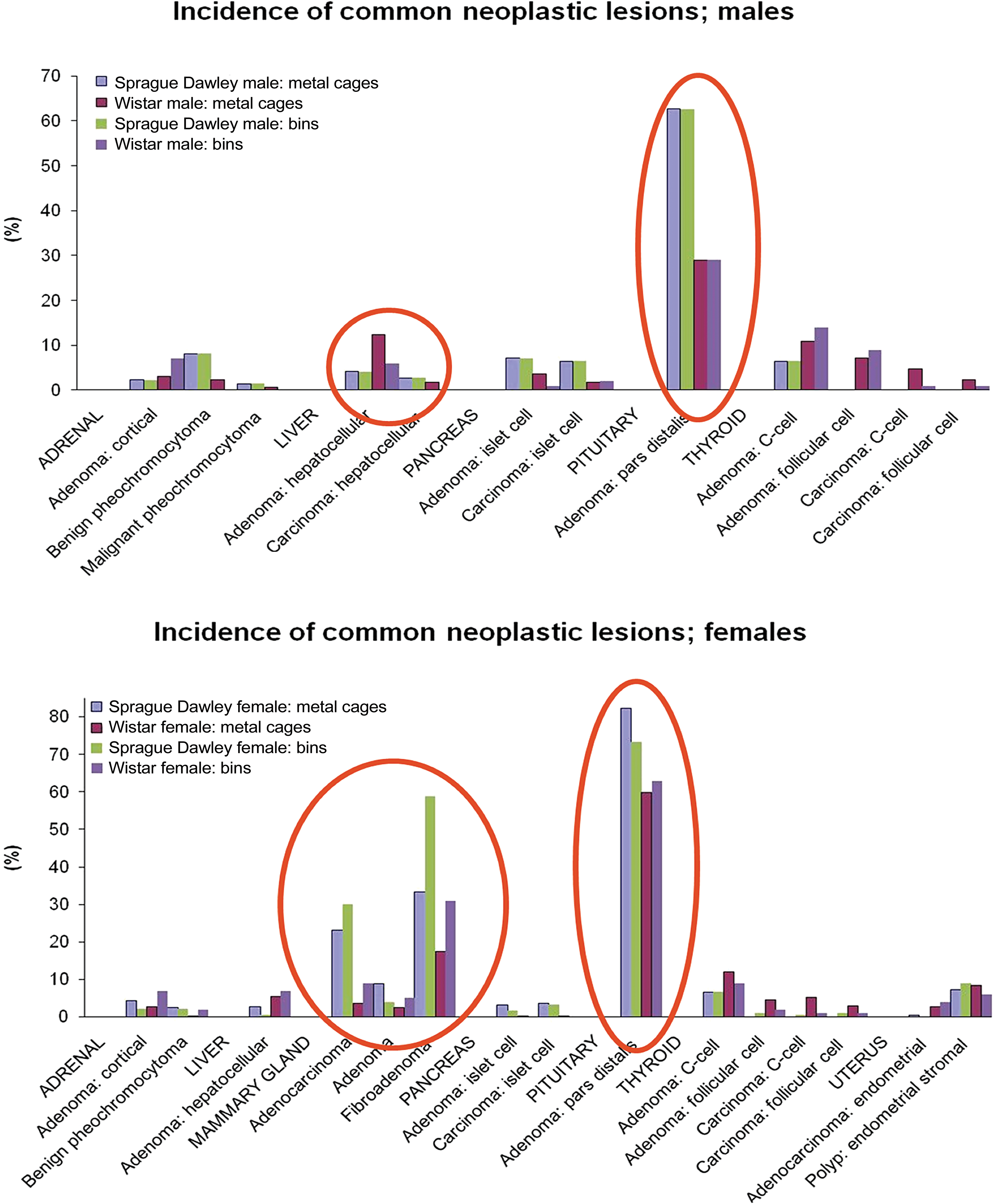
Comparison of tumor incidences in control animals in 2 strains of rat and in 2 different housing paradigms—an example of potential differences that can be observed (not intended to provide definitive differences for reference purposes). For the males, a clear separation of the strains can be seen in the incidence of pituitary tumors, with no impact of the housing, while in females there is a suggestion of an effect of housing, albeit in different directions in the 2 strains, with a less clear difference in incidence between the 2 strains. In the liver of males, evidence of a difference in incidence of tumors can be seen between strains, with no difference in incidence in SD rats between cages, while Wistar rats had a more prominent potential difference in incidence between the caging. For females, an effect of variability between strain and caging could be identified, particularly for adenocarcinoma and fibroadenoma. Data and graphs were provided by Charles River Preclinical Services Montreal Inc. and were originally presented in a poster presentation at the Society of Toxicology (2015).
Rat Strain Variability of Positive Ureaplasma parvum Culture in the Urinary Tract at 2 Weeks Postinoculation with 109 Colony-forming Unit (CFU) of U. parvum in the Urinary Bladder.
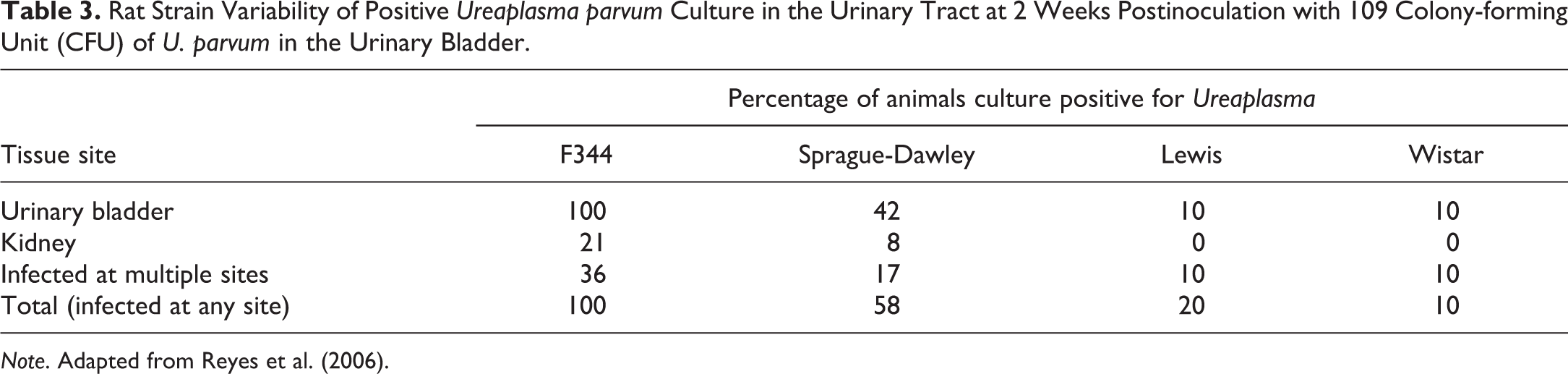
Note. Adapted from Reyes et al. (2006).
The effects of some environmental factors on background observations and metabolic enzyme activities are well established. Several authors have published on the impact of diet on background histopathology, both for neoplastic and nonneoplastic lesions, and on the effects of diet on metabolizing enzyme activities in rats (Eggleston and Krebs 1969; Haseman et al. 2003; Keenan et al. 1997, 1996; Laroque et al. 1997). More recent work related to the increased use of group housing instead of individual cages has identified some parameters where no or variably discernable differences have been observed in body weights or tumor profiles (Figures 3 and 4). There is, however, evidence that environmental alterations that increase or decrease subclinical stress levels may have an impact on anatomic pathology parameters, sometimes in a strain-related manner (Castelhano-Carlos and Baumans 2009; Sharp, Azar, and Lawson 2014). Therefore, although the literature and published data are conflicting with regard to possible effects of housing and other environmental factors on study results and data, it is essential to consider the environment an important part of the equation, primarily to reduce possible complications and also when evaluating confusing data during the interpretation of the results of studies (Wolf and Mann 2005).
Cynomolgus Monkeys
Comparisons of the prevalence of background observations in cynomolgus macaques from different primary (geographic) sources have been less well characterized, although several publications and texts are available giving background data on cynomolgus monkeys as a single species. More recent publications have compared various pathology data derived from animals originating in Mauritius and different Asian sources, identifying some trends with respect to the relative incidences of observations in the respective groups (Drevon-Gaillot et al. 2006; Kozlosky et al. 2015; Vidal et al. 2010). A few notable anatomic pathology differences between the sources are summarized together in Table 4, for illustrative purposes. Of particular note, Mauritian-sourced monkeys seem to have reduced incidence of lymphoid infiltrates and/or lymphoid activity in the gastrointestinal tract or associated lymph nodes, when compared to all sources of Asian monkeys. This finding is consistent with other data available, showing that Mauritian monkeys have different immune responses related to their major histocompatability complex allele concentrations (Menninger et al. 2002) and also exposures, or lack thereof, to certain pathogens such as malaria compared with monkeys obtained from Asian sources (Migot-Nabias et al. 1999). The prevalence of Plasmodium inui is 0% in Mauritian cynomolgus monkeys and those bred in closed, captive colonies outside of malaria endemic regions. The incidence of observed infections by P. inui is 10–14% in monkeys originating from Chinese and Indonesian facilities where malaria is endemic (Sasseville and Mansfield 2010), resulting in a more effective response to similar infectious agents and a generally more well-adapted immune system in these “Asian” populations. This difference in immune response between cynomolgus macaques obtained from different origins, breeding colonies, or even holding facilities can result in significantly different responses to the administration of immune-modulating xenobiotics or biopharmaceutic agents (the so-called biologics) in addition to published differences in background observations (Shenton, Henson, and Rebelatto 2015; Walter 2015).
Incidence Table of Select Microscopic Findings Showing Variations in Control Cynomolgus Macaques between Different Origins or Suppliers.
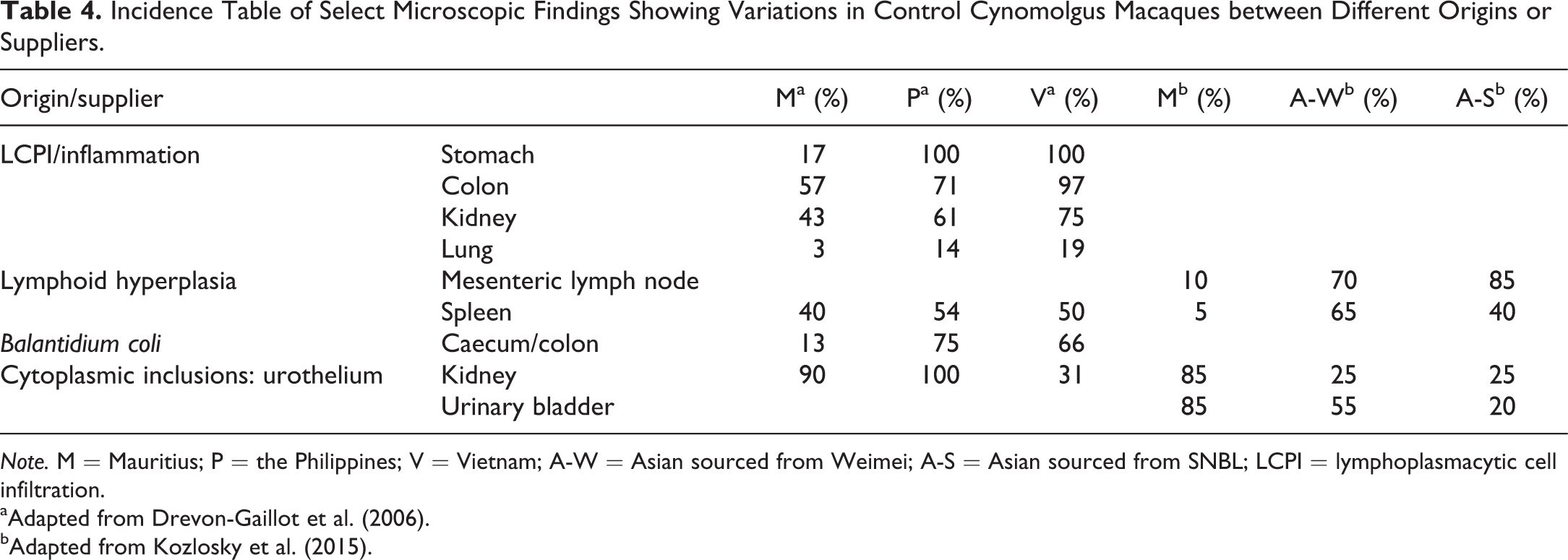
Note. M = Mauritius; P = the Philippines; V = Vietnam; A-W = Asian sourced from Weimei; A-S = Asian sourced from SNBL; LCPI = lymphoplasmacytic cell infiltration.
aAdapted from Drevon-Gaillot et al. (2006).
bAdapted from Kozlosky et al. (2015).
Examples of Effects of Variability on Responses to Xenobiotics
An excellent review of rat strain-related differences in response to the administration of xenobiotics was published in 1996 following a number of publications identifying clear strain-related variation in sensitivity to xenobiotics (Kacew and Festing 1996). This publication highlights examples of important divergent results (sensitive and resistant strains) and is a very useful tool when comparing “class effects” of similar therapeutic agents that utilize different strains for their evaluation. It may also have utility when choosing an appropriate rat model for any given program to ensure the highest relevance of any observations to the clinical patient.
Four recent examples of strain- or source-related challenges observed following the administration of a pharmaceutical are summarized below to highlight the importance of these variables in our nonclinical safety studies.
Example 1—Small Molecule Demonstrating Different Sensitivity to Adverse Effects in Rats (Table 5)
In this example, compound A was administered by oral gavage to SD rats in a number of routine investigational new drug-enabling studies and subsequently to Wi:Han rats in an investigative study. All studies were conducted in accordance with the established North American animal care regulations and standards (Association for Assessment and Accreditation of Laboratory Animal Care [AAALAC]–certified facilities) and governmental guiding principles in the use of animals. The salient results of the various studies are summarized in Table 5.
Example 1—Different Sensitivity to Adverse Effects in Rats (Strain and Gender).
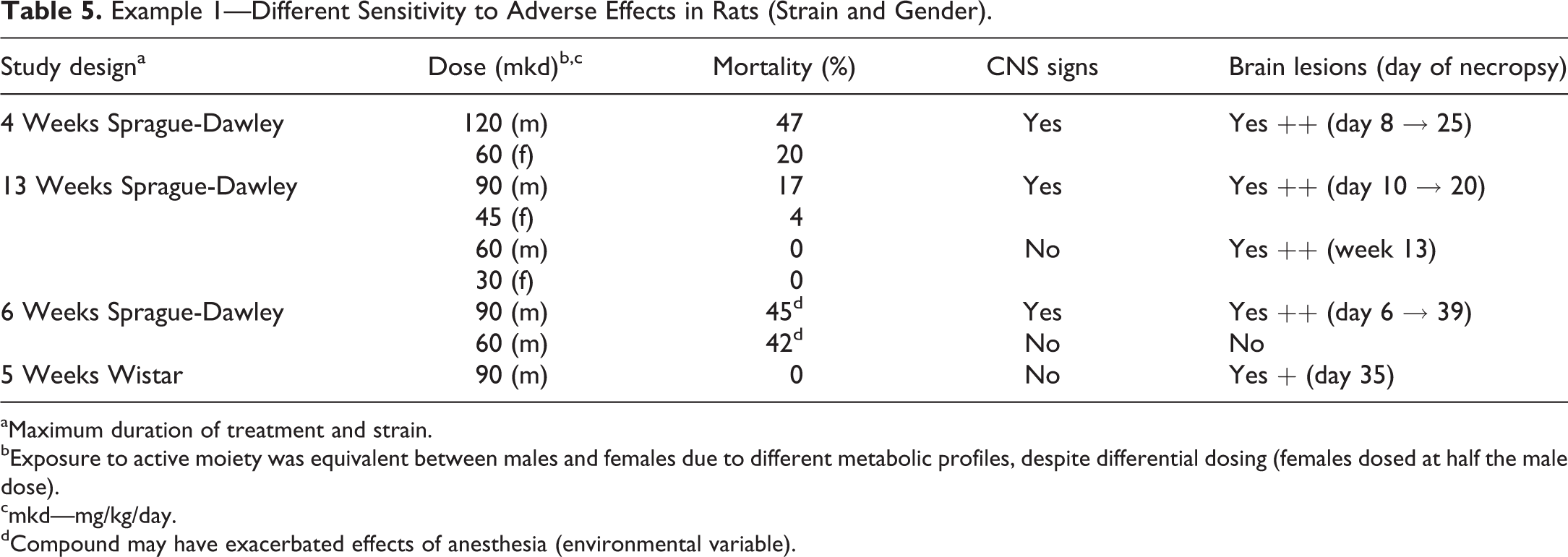
aMaximum duration of treatment and strain.
bExposure to active moiety was equivalent between males and females due to different metabolic profiles, despite differential dosing (females dosed at half the male dose).
cmkd—mg/kg/day.
dCompound may have exacerbated effects of anesthesia (environmental variable).
Exposures to the active moiety were equivalent in males and females with disparate dosing (females received half the dose administered to male for the same systemic exposure) and in Wis and SD rats at the same administered dose levels. Despite the equivalent exposures, males were more sensitive to the effects of treatment and SD rats considerably more sensitive to central nervous system (CNS) toxicity than Wi rats at the same exposure levels.
Example 2—Small Molecule Demonstrating Different Sensitivity to Adverse Effects in Cynomolgus Macaques (Table 6)
In this example, compound B was administered by oral gavage to cynomolgus macaques of Asian origin (unspecified precise geographic location) in a number of routine Good Laboratory Practice (GLP) studies for up to 13 weeks and subsequently to cynomolgus macaques of Mauritian origin for up to 5 days. All studies were conducted in accordance with Novartis animal care regulations and governmental guiding principles in the use of animals at AAALAC-accredited facilities. Select results of the various studies are summarized in Table 6.
Different Sensitivity to Adverse Effects Related to Geographic Origin of Cynomolgus Macaques.
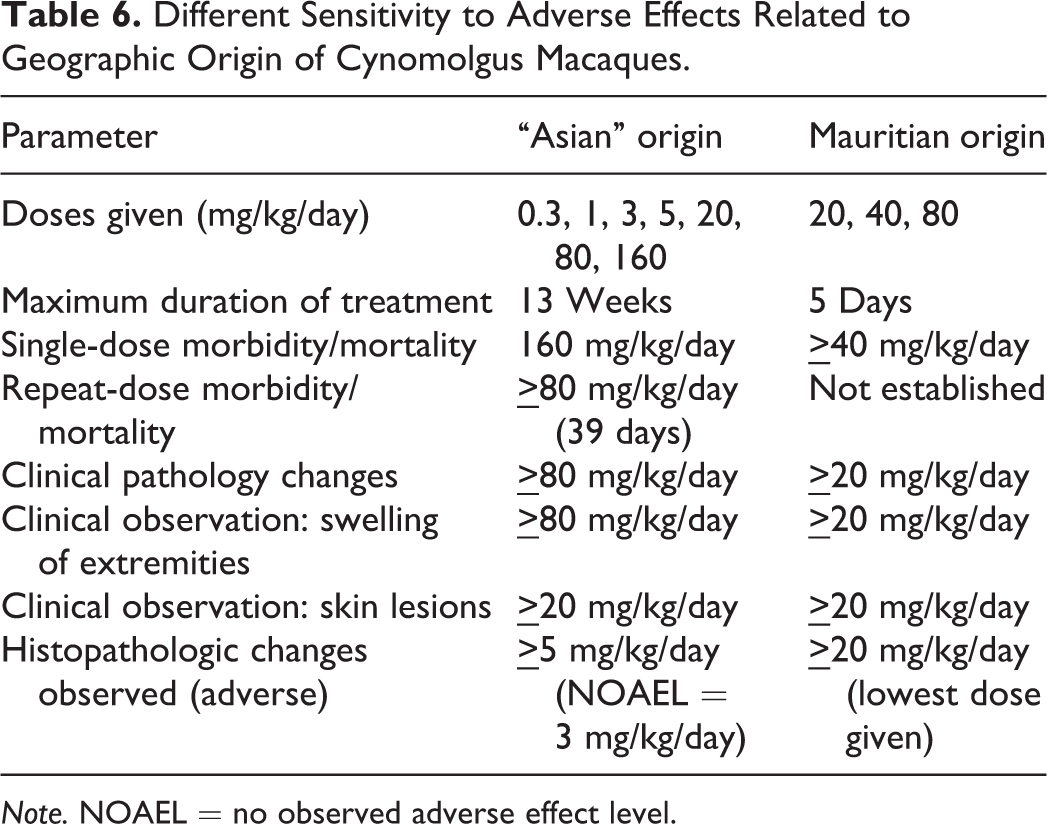
Note. NOAEL = no observed adverse effect level.
The Mauritian-sourced macaques were considerably more sensitive to the toxicities observed than were the monkeys originating from Asia. This phenomenon was also observed with several other programs using the same class of compound (data not presented) and was considered related to the inherent differential physiology in the cynomolgus macaques from the 2 primary origins.
Example 3—Variable Antidrug Antibody (ADA) Response to a Novel Monoclonal Antibody (mAb) in Cynomolgus Macaques of Asian Origin
In this case, a humanized therapeutic mAb was administered to cynomolgus monkeys of Asian origin in 2 separate studies. Both studies were conducted at the same AAALAC-accredited Contract research organization (CRO) in accordance with Novartis animal care regulations and governmental guiding principles in the use of animals. Within the 2 studies, only a few animals demonstrated seroconversion with ADA production and with no clear pattern of response across the treatment groups. When the source of the animals was investigated more carefully, it became apparent that the all ADA-positive animals had been bred locally by the CRO performing the studies and were initially derived from parents of Vietnamese origin. All the ADA-negative animals had been captive-bred in Asia (China) and were of an undisclosed primary origin but stated not to be Vietnam. These 2 studies using the exact same biotherapeutic with the same confusing immunologic outcome demonstrate nicely the significant impact on study results of different primary geographic origin within Asia and sourcing from different breeding facilities. This difference in immune response between 2 different proximal sources of Asian monkeys also has the potential to result in variable histopathology profiles and variable risk of immune-complex disease within a study purported to use “single source,” for example, Asian origin, Cynomolgus macaques. The mixing of animals in this way can result in confusing or confounding results which may require additional resources to understand in order to evaluate the true risk of the therapeutic to the patient.
Example 4—Transmission of Pathogens between Cynomolgus Macaques from Different Proximal Sources during Study Conduct (Table 7; Figures 5 and 6)
This example is an extension of the previous example where monkeys from 2 different holding facilities at distant locations from one another but all from the same original breeding colony, are brought together in 1 study. This is not an unusual occurrence within larger studies.
Overview of Stool Consistency Scores and Mortality in Cynomolgus Macaques Given an Immune-modulating Therapeutic.

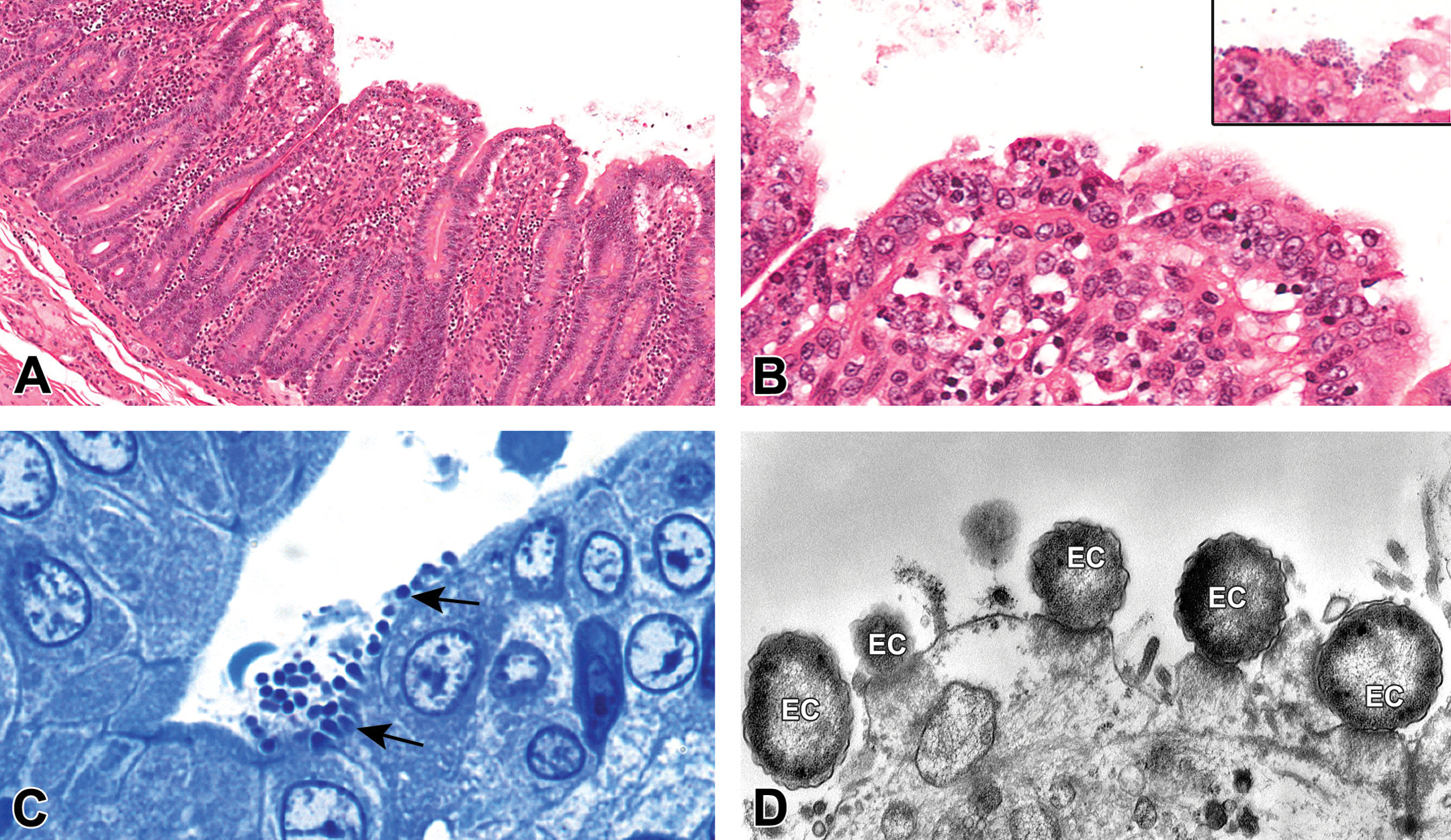
Photomicrographs of enteropathogenic Escherichia coli (EPEC) in the small intestine of an infected macaque. (A) and (B) H&E-stained micrographs showing typical EPEC attaching and effacing lesions and (inset) surface bacteria (original objectives: 5× and 20/40×, respectively). Note the villus clubbing and shortening with subepithelial vacuolar change. (C) Semithin toluidine blue section showing bacteria attached to the surface of the epithelial cells indicated by the arrows and concomitant loss of brush border (original objective 100×). (D) Electron micrograph showing bacteria (EC) with tight attachment to the cell surface and epithelial cell injury characterized by degeneration and loss of cilia in the brush border (107× magnification).
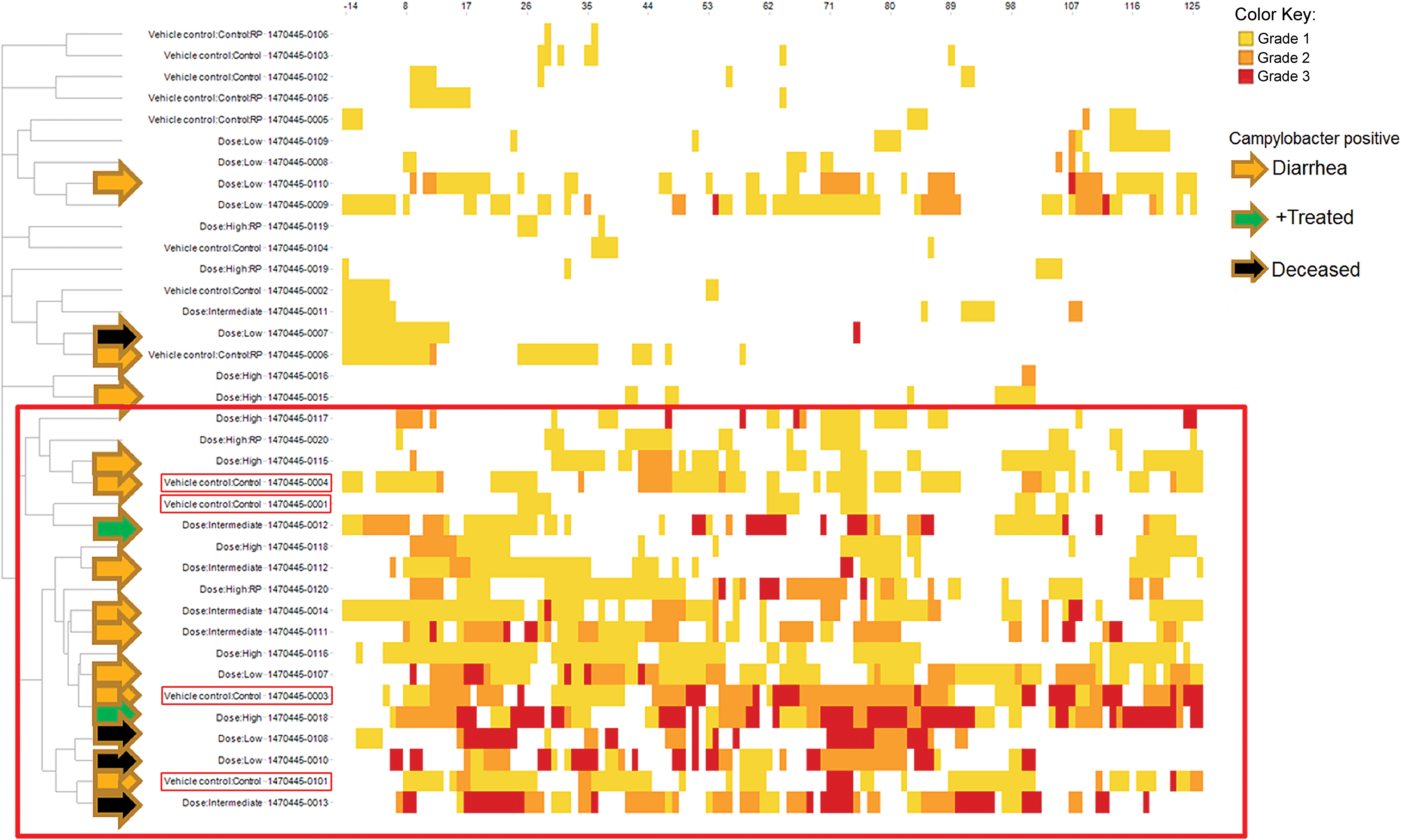
Hierarchical clustering of clinical diarrhea scores showing more and less affected groupings including controls. A higher rate of Campylobacter-positive results was identified through fecal culture in the more affected grouping (within the large red rectangle). Note the 4 vehicle control animals with a clinical pattern similar to treated animals in this more affected grouping (4 small red rectangles). The colored boxes in the graphic indicate the grades of clinical diarrhea observed on various days of the study (x-axis along the top). The severity grading for the colors used can be observed in the top right corner (1 = mild, 2 = moderate, and 3 = severe diarrhea). Animals with arrows on the y-axis are those with Campylobacter-positive results through fecal culture. Animals identified as treated (green arrows) were given symptomatic and antimicrobial treatment to alleviate the clinical signs, treat the infectious agents, and prevent further loss during the course of the study.
The study was conducted at an AAALAC-accredited CRO in accordance with Novartis animal care regulations and governmental guiding principles in the use of animals.
In this study, animals from 2 different holding facilities utilized by the CRO but originally from the same source colony were brought together due to the number of animals required for proper conduct of the GLP study. The 2 sets of animals had differing microbiomes consisting of disparate commensal flora and fauna on arrival at the CRO test site resulting in the transmission of organisms from 1 group to the other during study conduct, some of which were mildly pathogenic. The compound administered in this case was an immune-modulating small molecule and therefore had the potential to exacerbate or enable severe infections with low pathogenic microbes. As the study progressed, approximately 50% of the animals showed clinical signs of diarrhea with a higher incidence in treated animals, although with no dose response (Table 7). The clinical signs became so severe in 4 treated animals that they were euthanized during treatment (3 low-dose animals and 1 mid-dose animal), and these mortalities were attributed to direct effects of treatment on the gastrointestinal tract following initial histopathologic evaluation. A regulatory safety alert for new adverse findings was triggered and further work was initiated to try to identify the pathogenesis of the severe, unexpected, intestinal “toxicity.” Using polymerase chain reaction and electron microscopy, 2 primary pathogenic organisms were identified in conjunction with the clinical and histopathologic observations, Campylobacter spp., and enteropathogenic Escherichia coli (Figures 5 and 6). As the investigations progressed, it became clear that there were 2 populations of cynomolgus macaques on the study that could be divided based on clinical signs and prevalence of Campylobacter positivity (Figure 6) with 4 control animals also showing similar clinical observation of diarrhea and Campylobacter positivity as the 4 severely affected animals, although not requiring early euthanasia. By identifying the pathogenic organisms involved and eliminating a direct effect of the compound on the intestinal tract, while identifying through CRO sourcing records that there were 2 different holding facilities involved despite a single original source of cynomolgus macaques used on study, the study was completed and the clinical program was able to be continued with confidence in the results obtained.
Conclusions and Recommendations
It is important to realize the multitude of variables, some obvious and others not so apparent, that can be encountered when planning, designing, and executing nonclinical studies. As such, a sound understanding of the current genotype, microbiologic and immune status and background findings for the species, strain, geographic origin, and/or supplier of the animals used in nonclinical studies is essential. This will enable the best possible hazard identification, risk assessment, and translatability to human patients.
The following are some simple recommendations, in no particular order, to enable adequate interpretation of study and nonclinical program results. Ensure the consistent use of the same strain, supplier, and geographic origin (and, where possible, breeding colonies) for each individual drug development program to reduce the variability in possible response between studies. The sometimes significant differences in physiology, metabolism, and genetically programmed responses (including immune responses) between animals of the same species or strain originating from different sources or suppliers has been and continues to be underappreciated. Ensure that all animals on any one study are from the same strain, origin, and source (including holding facilities). It is not generally realized how often groups of cynomolgus macaques from different suppliers and/or holding facilities are placed in the same study. This is a result of the demand for cynomolgus macaques within biomedical research and is a practice that needs to be reconsidered. Less frequently cynomolgus monkeys from different origins are placed on the same study, creating even greater difficulties in interpretation and understanding the results produced. The use of Asian as an origin is too broad a category with notable differences between animals originating from different regions such as Vietnam, the Philippines, Indonesia, and now captive-bred colonies within China. Much more consistency is seen in Mauritian macaques, and this makes them more valued as a research model in many companies; however, they are not as widely or readily available as their Asian counterparts. Ensure adequate acclimation to new surroundings before placing on study. In order for the animals to adjust to their new environment on study, including the caging and social situation, diet, water source, and adjustments of their microbiome, and to allow for any necessary treatment prior to study start, should cross-contamination by pathogens between animals occur, the acclimation period should be of reasonable length. A minimum of 7–14 days prior to any study-related activities is recommended for rodents and large animals, respectively, with longer durations to be considered dependent on the proximal source of the animals (mixed or single source), class of agent to be administered, specific study requirements or end points, and/or evidence of infectious agent transmission between animals during the acclimation period. This prestudy period is extremely important with a significant role in the successful conduct of the study and deserves special attention. Where possible, select the most appropriate species, strain, substrain, and origin for the whole program early in the research and development process. Nonclinical species and strains should be selected for maximal translatability to the human patient. This can be achieved through genetic testing of patient populations and nonclinical species and/or strains for target distribution and target homology. Sound knowledge of the similarities and differences between the physiology, metabolic processes, and immune responses of the nonclinical species selected and the patient population is also important.
Footnotes
Acknowledgments
The author would like to thank CRL Montreal (Pierre Tellier, Gayle Hennig, and Peter Mansell) for graciously sharing their data and allowing its use in the presentation and this manuscript. In addition, thanks go to Chris Hayden, Keith Mansfield, Robert Kreutzer, Karen Killary, Peter Hoffmann, and Vito Sasseville for sharing their expertise, data, and/or providing images and tables. Finally, I extend a big thank-you to Vito Sasseville, Keith Mansfield, Emily Meseck, Theresa Boulineau, and Diane Gunson for their reviews and critical comments.
Author Contribution
All authors (KC) contributed to conception or design; data acquisition, analysis, or interpretation; drafting the manuscript; and critically revising the manuscript. All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.