
Abstract
Cumene hydroperoxide (CHP) is a high production volume chemical that is used to generate phenol and acetone. Dermal exposure to CHP was hypothesized to result in systemic tissue toxicity, production of free radicals, and consequent decrease in plasma antioxidant levels. To evaluate the hypothesis and characterize the toxicity of CHP, male and female B6C3F1/N mice and F344/N rats were exposed to varying doses of CHP applied topically for 14 or 90 days. No significant changes in survival or body weight of mice and rats were observed following 14 days of exposure. However, 90 days of CHP exposure at the high dose (12 mg/kg) triggered a significant decrease (−15%) in the body weight of the male rat group only. Irritation of the skin was observed at the site of application and was characterized by inflammation and epidermal hyperplasia. In treated animals, histology of liver tissue, free radical generation, and antioxidant levels in blood plasma were not significantly changed as compared to the corresponding controls. Consistent with the lack of systemic damage, no increase in micronucleated erythrocytes was seen in peripheral blood. In conclusion, topical CHP application caused skin damage only at the application site and did not cause systemic tissue impairment.
Introduction
Cumene hydroperoxide (CHP) is a strong oxidizing agent that is produced mainly through the oxidation of cumene and used primarily in the production of acetone and phenol (Yadav and Asthana 2003). Other industrial applications of CHP include use as a catalyst, curing agent, or initiator for polymerization. It is a high production volume chemical with an aggregated production volume of 149 million pounds per year based on manufacturer information submitted to the U.S. Environmental Protection Agency (EPA; CAS number search of http://java.epa.gov/oppt_chemical_search/). CHP is listed on the EPA Final Third Drinking Water Contaminant List (CCL 3), which identifies contaminants that are currently unregulated but are known or anticipated to occur in public water systems and may require regulation under the Safe Drinking Water Act. As a likely product of petroleum photooxidation (Larson, Hunt, and Blankenship 1977), CHP could enter aquatic systems following oil spills.
Human exposure to CHP most likely occurs in an occupational setting via dermal contact or inhalation during the manufacturing process. There are currently no set regulatory guidelines for CHP occupational exposure limits. However, the Workplace Environmental Exposure Levels™ (WEEL) committee of the American Industrial Hygiene Association® (AIHA), which develops suggested exposure limits, lists a 2011 WEEL value of 1 ppm for an 8-hr time-weighted average exposure to the skin for CHP.
Toxicity and carcinogenicity data for estimating risks to human health associated with exposure to CHP are limited and largely derived from acute or short-term animal studies. CHP has exhibited genotoxicity in several in vitro studies. It was reported to be mutagenic in Salmonella typhimurium strains TA98, TA100, and TA102 (Lai et al. 1996; Yamaguchi and Yamashita 1980). In L5178Y tk+/− mouse lymphoma cells, CHP induced DNA damage as measured by the comet assay and micronuclei; it also induced gene mutations at the thymidine kinase locus in these cells (Brink et al. 2009). In contrast to these positive results, CHP did not induce DNA adducts (i.e., 8-oxo-7,8-dihydro-2-deoxyguanosine or 1,N6-etheno-2-deoxyadenosine) in L5178Y tk+/− cells (Brink et al. 2009).
In rats, as expected based on its reactivity, CHP has been found to be irritating to skin following dermal application (Brown, Box, and Simpson 1975) and to the respiratory system following inhalation exposure (Gage 1970). Data on carcinogenicity in rodent models is inconsistent. A single subcutaneous sarcoma and 11 incidences of malignant lymphomas were reported in a group of 50 C57B1 mice treated with 50-µM CHP via subcutaneous injection (experimental details not provided) with a 14-month time to appearance of first tumor (Kotin and Falk 1963). However, these results are difficult to interpret due to the absence of a concurrent control and inadequate study details. In contrast, no increase in tumor incidence was reported in a lifetime dermal application carcinogenicity study of CHP in Swiss Millerton mice (1% CHP dissolved in benzene, 3 times per week until natural death; Van Duuren, Orris, and Nelson 1965). Using a SENCAR mouse model to evaluate tumor initiation and promotion potential, dermal application of CHP for 4 weeks at 25 or 50 µmol dissolved in acetone caused significant increases in epidermal hyperplasia and dermal inflammation, indicating promotion potential, but did not cause an increase in c-Ha-ras mutations, which was the end point selected to indicate initiating activity (Hanausek et al. 2004).
Data characterizing the distribution of CHP following exposure in a rodent model were not identified in the literature. CHP is metabolized by the cytochrome P450 system via hemolytic cleavage of the O–O bond, producing singlet oxygen and free radicals including hydroxyl (•OH), alkoxyl (•RO), and alkyl (•R) radicals that can be detected and characterized by electron spin resonance (ESR) and spin-trapping techniques (Taffe et al. 1987). Furthermore, bioactivation of CHP by cytochrome P450s has been shown to be involved in lipid peroxidation in rat hepatocytes (Anari, Khan, and OBrien 1996) and other P450-containing biological systems (Weiss and Estabrook 1986). CHP has also been used experimentally to generate intracellular reactive oxygen species (Ayala et al. 1996). The ability of CHP to generate free radicals (Timmins and Davies 1993a, 1993b) and the association of free radicals with tumor promotion (Copeland 1983; Floyd 1990) have led to concern for a possible association between exposure to CHP and development of cancer.
The goal of this investigation is to study the toxicity of CHP by topical dermal exposure on systemic tissues, generation of free radicals, and consequently, the role of endogenous antioxidants. We performed 14-day and 90-day dermal toxicity studies with male and female B6C3F1/N mice and F344/N rats. Results from these studies provide useful data for characterizing dermal toxicity of CHP and establish whether further chronic toxicity and carcinogenicity studies are warranted.
Materials and Methods
Chemicals
CHP (Chemical Abstracts Service [CAS] No. 80-15-9, Lot No. 2529F) was procured from ICN Biomedicals (Aurora, OH). The chemical identity and purity were evaluated at RTI International (Research Triangle Park, NC). The identity of the chemical was confirmed by infrared spectroscopy, nuclear magnetic resonance spectroscopy, and mass spectrometry. The purity of the material as estimated by high-performance liquid chromatography (HPLC) with ultraviolet detection was 86%; identified impurities included acetophenone (1.5%), 2-phenyl-2-propanol (7%), and cumene (2.4%). Dose formulation development and stability studies were conducted at RTI International using HPLC coupled with ultraviolet detection at 210 nm. CHP (1 mg/ml) in 95% ethanol stored in amber vials was stable up to 42 days at room (23°C to 28°C), refrigerator (2°C to 8°C), and freezer (−20°C) temperature. Assessment of the dosing solution under simulated dosing conditions (exposed to air and light) for 3 hr revealed no significant loss of the test chemical. The vehicle used for all studies was US Pharmacopeia (USP)-grade 95% ethanol (Lot No. SO0121) procured from Spectrum (Gardena, CA).
Animals and Clinical Observations
All studies were conducted at the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) accredited facility of Battelle Columbus Laboratories (Columbus, OH). Male and female F344/N rats and B6C3F1/N mice were supplied by Taconic Laboratories Animals and Services (Germantown, NC). Mice and rats were held for 11 to 14 days before being randomly assigned to treatment groups to achieve approximately equal initial mean body weights among groups. Animals were identified by tail tattoo. Animals were approximately 5 to 6 weeks old at the start of the study (i.e., initiation of dosing).
Animals were housed 1 per cage in polycarbonate cages with irradiated Sani-Chips hardwood chip bedding (P. J. Murphy Forest Products Corp., Montville, NJ) and given NTP-2000 wafer feed (Zeigler Brothers, Gardners, PA) and municipal water (Columbus, OH) ad libitum. Mice and rats were treated with CHP in USP-grade 95% ethanol vehicle (Spectrum, Gardena, CA) via dermal application 5 days per week over a 14- or 90-day period. The animal room was maintained at a temperature of 69°F to 75°F and a relative humidity of 35 to 65%.
Animals were observed twice daily for signs of moribundity or mortality, and clinical observations were recorded once daily. Body weights were recorded prior to study start, on the first day of treatment, and weekly thereafter.
Dosing Method
For the 14-day studies in mice and rats, CHP in 95% ethanol was applied dermally to 5 animals/sex/species at doses of 0 (vehicle control), 6.25, 12.5, 25, 50, or 100 mg/kg in a volume of 2 ml/kg (mice) or 0.5 ml/kg (rats) 5 times per week on weekdays for a total of 12 days (rats) or 13 days (mice). The doses were applied directly to the dorsal skin using a pipette. The fur at the application site was clipped weekly. For the 90-day studies in mice and rats, doses of 0 (vehicle control), 0.75-, 1.5-, 3-, 6-, or 12-mg/kg CHP were applied dermally to 10 animals/sex/species. The high dose of 12-mg/kg CHP for the 90-day studies was selected based on effects observed in the 12.5-mg/kg dose and above in the 14-day studies (including extension of lesions into the dermis and the severity of necrosis in the epidermis at the site of application), which suggested that animals at doses above 12.5 mg/kg could develop ulcerations and/or infections over the course of the 90-day exposure. In addition to the animals in the standard 90-day studies, 50 male rats were treated with 0, 3-, or 12-mg/kg CHP for collection of plasma and tissues for the detection of radical generation (n = 5) and plasma antioxidant measurements (n = 5).
Blood and Plasma Collection
At the end of the 90-day studies, animals were anesthetized with a 70% carbon dioxide to 30% oxygen mixture, and blood was collected via the retro-orbital plexus (rats) or retro-orbital sinus (mice) for assessment of hematology, osmotic fragility of erythrocytes, and erythrocyte micronuclei (mice only). Blood was transferred to ethylenediaminetetraacetic acid–containing tubes. Plasma for the radical generation and plasma antioxidant studies in rats was harvested from blood collected via cardiac puncture after anesthesia with a 70% carbon dioxide to 30% oxygen mixture. The whole blood was placed into heparinized tubes that were then centrifuged. Plasma was collected in 2 vials and stored at −70°C until evaluation.
Hematology and Osmotic Fragility
Hematology parameters were analyzed using the Advia 120 hematology analyzer (Siemens Healthcare Diagnostics Inc., Tarrytown, NY). Parameters measured included red blood cells, hemoglobin, hematocrit, mean corpuscular volume, mean corpuscular hemoglobin, mean corpuscular hemoglobin concentration, reticulocytes, white blood cells, absolute lymphocytes, monocytes, eosinophils, basophils, and platelets. Osmotic fragility of red blood cells was determined by adding 20 µl of whole blood to a set of 16 test tubes containing 5 ml of 0.00 to 0.85% buffered NaCl in distilled water, inverting the tubes, incubating at room temperature for 30 min, centrifuging at 2,000 rpm for 10 min, and measuring the optical density of the supernatant at 540 nm.
Necropsy and Histopathology
At study termination for the 14-day studies and following blood collection for the 90-day studies, animals were euthanized in a carbon dioxide chamber (100% carbon dioxide) followed directly by necropsy. Organ weights recorded in the 14-day and 90-day studies included liver, lung, thymus, heart, right kidney, and right testis (male). Tissues collected for microscopic examination were fixed and processed according to National Toxicology Program (NTP) specifications resulting in 5- to 6-μm-thick paraffin-embedded sections stained with hematoxylin and eosin (H&E). In the 14-day studies in mice and rats, microscopic evaluation was performed on the liver, kidney, lung, and skin (both at the site of application and inguinal). Tissues were examined in all control animals and to a no observed effect level (NOEL) in the dosage groups. In the 90-day studies, complete necropsy and histopathologic evaluations were conducted on all control and 12-mg/kg mice and rats. At necropsy, all organs and tissues were examined for grossly visible lesions, and all major tissues were fixed and preserved in 10% neutral buffered formalin, processed and trimmed, embedded in paraffin, sectioned to a thickness of 5 to 6 μm, and stained with H&E for microscopic examination. All tissues including the skin at the site of application were examined in all control and high-dose animals and then to a NOEL in the remaining dosage groups. Semiquantitative grading of microscopic findings was performed according to the following scale: minimal (barely detectable alteration without functional significance) = 1, mild (change likely to be subclinical with only minor functional significance) = 2, moderate (possible clinical effects) = 3, and marked (approaching maximal severity of that lesion) = 4.
Bacterial Mutagenicity
Bacterial mutagenicity testing was conducted in 2 independent laboratories, and procedures were based on the protocol described by Mortelmans et al. (1986) and Zeiger et al. (1992), with some modifications. Briefly, CHP was incubated with S. typhimurium strains TA98, TA100, TA1535, and TA1537 and Escherichia coli strain WP2 uvrA/pKM101 (analogous to S. typhimurium strain TA102) with and without S9 mix (metabolic activation enzymes and cofactors from induced male Sprague-Dawley rat or Syrian hamster liver) for 20 min at 37°C. Following addition of top agar supplemented with
Mouse Peripheral Blood Micronucleus Test
A detailed description of the assay is presented in MacGregor et al. (1990). Peripheral blood samples from male and female mice were obtained at the end of the 90-day toxicity studies, and smears were immediately prepared and fixed in absolute methanol. The methanol-fixed slides were stained with acridine orange and scanned to determine the frequency of micronucleated erythrocytes per 2,000 normochromatic (mature) erythrocytes in each animal per treatment group. Data were evaluated as the mean number of micronucleated cells per 1,000 cells for each treatment group.
Specimen Collection and Measurement of Radical Formation and Antioxidant Status
Rat plasma preparation
Blood (∼5 ml) was collected via cardiac puncture following anesthesia with a 70% carbon dioxide to 30% oxygen mixture through single-draw vacutainer needles (21 Gauge) into open heparinized vacutainer blood collection tubes. The tubes were gently inverted 2 to 3 times for mixing and immediately placed on ice. Blood was centrifuged (2,000 rpm for 10 min at 4°C) no more than 30 min after collection, and the plasma supernatant was transferred to cryovials and stored at −70°C until analysis.
Analysis of antioxidant status
Ascorbic acid and uric acid were measured in plasma. Plasma (0.2 ml) was added to 0.6 ml of 4% perchloric acid, vortexed, frozen on dry ice, and stored at –70°C. Perchloric acid supernatants (27,000 ×g, 20 min) of plasma were assayed by high performance liquid chromatography with electrochemical detection (HPLC-ED) for uric acid and ascorbic acid according to methods described by Kutnink et al. (1987). The same isocratic method quantifies both antioxidants. Total glutathione (GSx, consisting of the sum of reduced glutathione [GSH] and oxidized glutathione [GSSG]) was analyzed by enzymatic recycling (Anderson 1985).
Detection of free radicals with ESR
Five male rats per treatment group (0, 3-, or 12-mg/kg CHP) were designated for a free radical detection study. On the 90th day of the exposure, rats were anesthetized with pentobarbital (50 mg/kg ip). Animals were given an ip (1 g/kg) injection of the spin-trapping agent α-(4-pyridyl-1-oxide)-N-tert-butylnitrone; POBN), and 1 hr later, blood (5 ml) was drawn through single-draw vacutainer needles (21 gauge) into open vacutainer blood collection tubes containing heparin. The tubes were gently inverted 2 or 3 times for mixing and immediately placed on ice. Blood was centrifuged (2,000 rpm for 10 min at 4°C) no more than 30 min after collection. Rats were sacrificed after blood collection. After centrifugation, the plasma samples were transferred to a flat cell, and ESR spectra of ascorbate radical were recorded at room temperature with a Bruker EMX spectrometer equipped with a Super High Q cavity. Computer simulation analyses of spectra were performed using a program by Duling (1994).
Statistical analysis
For the in vivo studies, group mean body weights were compared using Fisher’s least significant difference method. Homogeneity of variances across groups was assessed using Bartlett’s test. For data with homogeneous variances across test groups, Dunnett’s test was used to assess differences between the control and treated groups. For nonhomogeneous data, the Cochran and Cox’s modified 2-sample t-test was used to test for differences between control and treated groups. Incidences of nonneoplastic lesions are presented as the number of animals affected with each lesion among the number of animals examined microscopically. Incidences of nonneoplastic lesions were assessed using the Cochran–Armitage trend test for dose-related trend and Fisher’s exact test for pairwise comparisons with the control group. One-sided p values of .05 or less were considered statistically significant.
For the genotoxicity assay, data were analyzed for increasing trend using a 1-tailed Cochran Armitage trend test (significant at p < .025), and pairwise comparisons between each treatment group and the corresponding control were statistically evaluated (significant at p < .025/3 = .008). In addition, the mean percentage of polychromatic erythrocytes (reticulocytes) was determined among a population of 1,000 erythrocytes for each exposure group as a measure of CHP-induced bone marrow toxicity.
For the radical formation and antioxidant status data, spectra at the same time point were compared between each dose group and the controls using analysis of variance. When appropriate, the data were log transformed prior to analysis. p values < .05 were considered statistically significant.
Results
14-day Study
All animals survived to the end of the study. There were no significant differences in body weight of dosed mice or rats as compared to vehicle control animals (data not shown). Irritation was visible at the site of application and was observed at dosages of ≥12.5 mg/kg (male and female rats and female mice) or 50 mg/kg (male mice). Ulceration was observed upon gross examination at the site of application in 1 female rat at 100 mg/kg. Histopathological lesions were limited to the site of application (Table 1). There were statistically significant increases in a spectrum of nonneoplastic lesions in the skin of male and female rats and mice that occurred in most dose groups and in general the severity of the lesions increased with increasing concentration of the applied CHP. The spectrum of lesions (described in detail in the histopathology section below) included hyperplasia, degeneration, necrosis, ulcer and exudate in the epidermis, chronic active inflammation and fibrosis in the dermis, hair follicle hyperplasia, and sebaceous gland hyperplasia.
Body Weights and Histopathological Lesions Observed at the Skin, Site of Application (SOA) in the 14-day Dermal Toxicity Study of Cumene Hydroperoxide in F344/N Rats and B6C3F1/N Mice (n = 5).
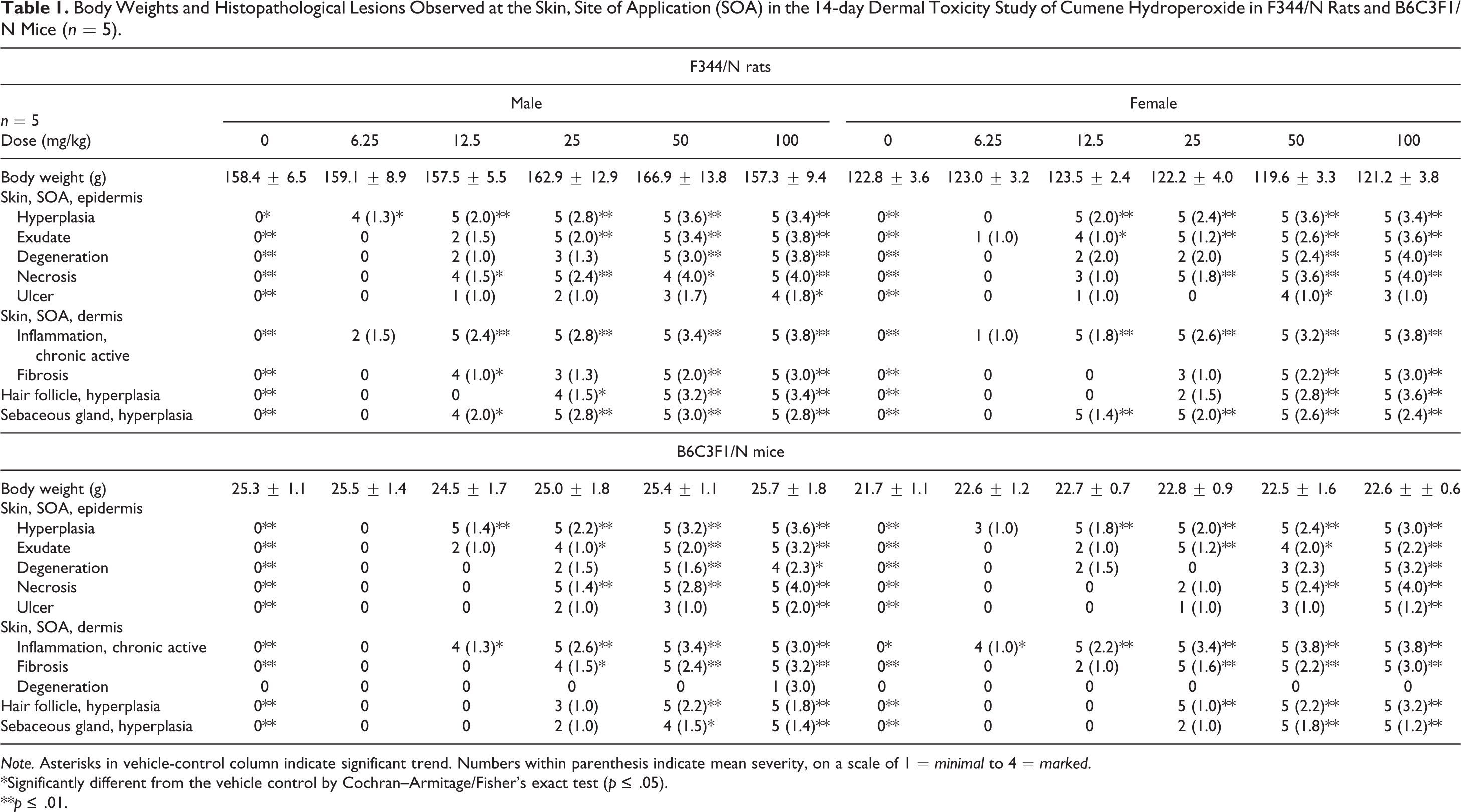
Note. Asterisks in vehicle-control column indicate significant trend. Numbers within parenthesis indicate mean severity, on a scale of 1 = minimal to 4 = marked.
*Significantly different from the vehicle control by Cochran–Armitage/Fisher’s exact test (p ≤ .05).
**p ≤ .01.
90-day Study
One rat from the main study (6 mg/kg) and 1 rat from the radical generation and plasma antioxidant studies (12 mg/kg) were sacrificed early due to chylothorax, which is characterized by lymphatic fluid accumulation in the pleural cavity. However, this affliction was relatively common in the F344/N rats during the period in which the study was conducted and was not thought to be treatment related. All other animals survived to the end of the study. Mean body weights of the 12-mg/kg CHP-treated male rats were significantly lower than controls (−15%), while there were no significant differences in body weight in female rats, male mice, or female mice.
Clinical observations and gross examination revealed irritation at the site of application in male (6 and 12 mg/kg) and female (12 mg/kg) rats, but not in mice. Increased numbers of neutrophils and white blood cells occurred in the 12-mg/kg female rats (data not shown) and were considered consistent with the inflammatory lesions that were observed histopathologically at the site of application. There were no other treatment-related effects observed in hematology or osmotic fragility parameters (data not shown) in mice or rats.
As in the 14-day studies, histopathological findings in the 90-day studies were limited to the site of application in both rats and mice (Table 2). Lesions observed in the 90-day studies were morphologically similar to those observed in the 14-day studies. At the highest applied dose in the 90-day studies (12 mg/kg), there were significant increases in the same spectrum of nonneoplastic skin lesions at the site of application in male and female rats that included hyperplasia, degeneration, necrosis, ulcer and exudate in the epidermis, chronic active inflammation in the dermis, hair follicle hyperplasia, and sebaceous gland hyperplasia. In male and female mice, statistically significant increases in treatment-related lesions were limited to hyperplasia in the epidermis and chronic active inflammation in the dermis at the 2 highest applied doses (6 or 12 mg/kg).
Body Weights and Histopathological Lesions Observed at the Skin, Site of Application (SOA) in the 90-day Dermal Toxicity Study of Cumene Hydroperoxide in F344/N Rats and B6C3F1/N Mice (n = 10).
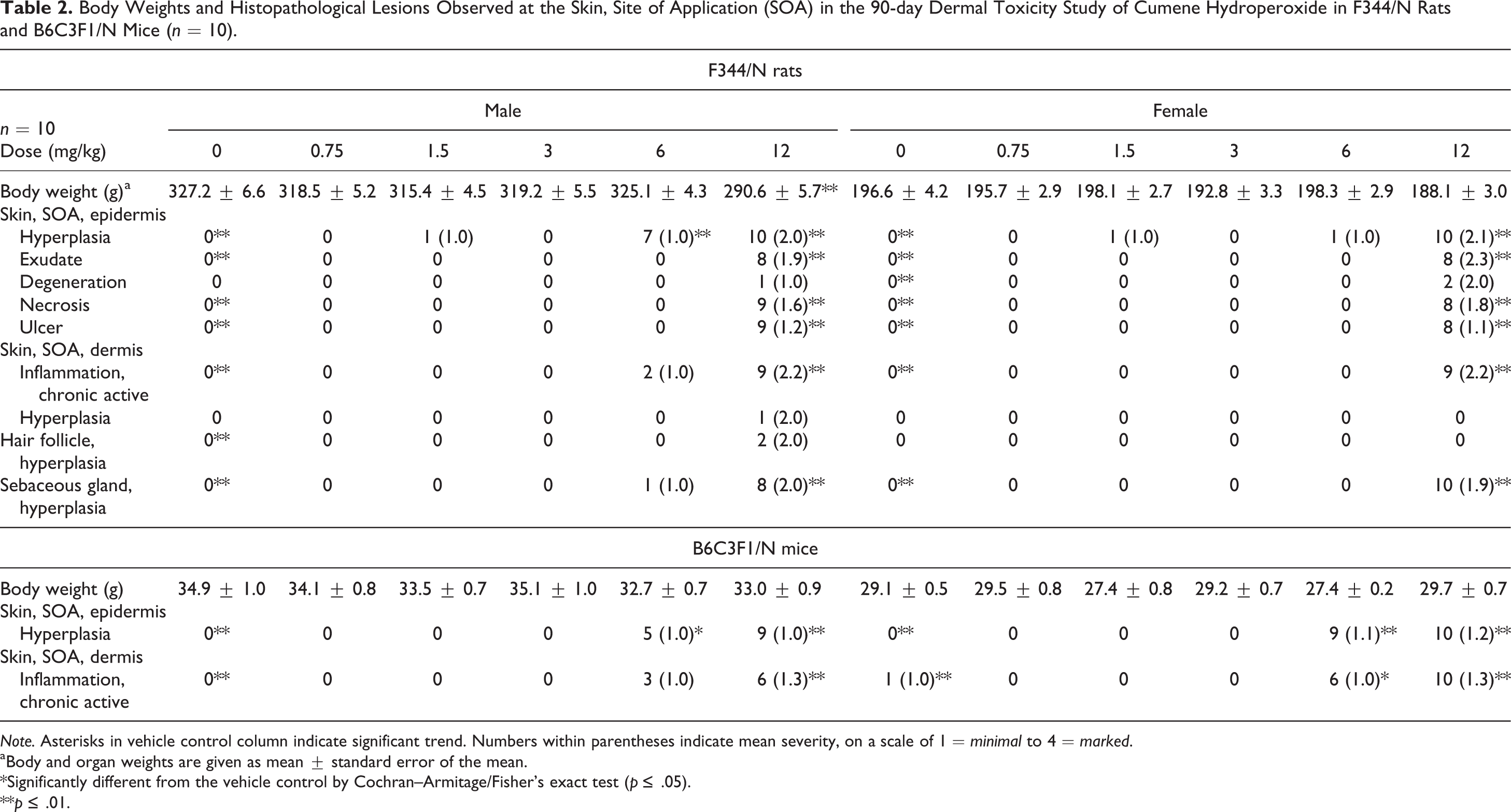
Note. Asterisks in vehicle control column indicate significant trend. Numbers within parentheses indicate mean severity, on a scale of 1 = minimal to 4 = marked.
aBody and organ weights are given as mean ± standard error of the mean.
*Significantly different from the vehicle control by Cochran–Armitage/Fisher’s exact test (p ≤ .05).
**p ≤ .01.
Detailed Descriptions of Histopathological Lesions from the 14-day and 90-day Studies
Microscopic changes observed at the skin at the site of application consisted of a spectrum of prominent, intimately associated lesions that in general were morphologically similar in the 14-day mouse and rat studies and the 90-day rat study (Figures 1 and 2). The lesions at the site of application in the 90-day mouse study were less severe and restricted to mild epidermal hyperplasia and dermal inflammation.
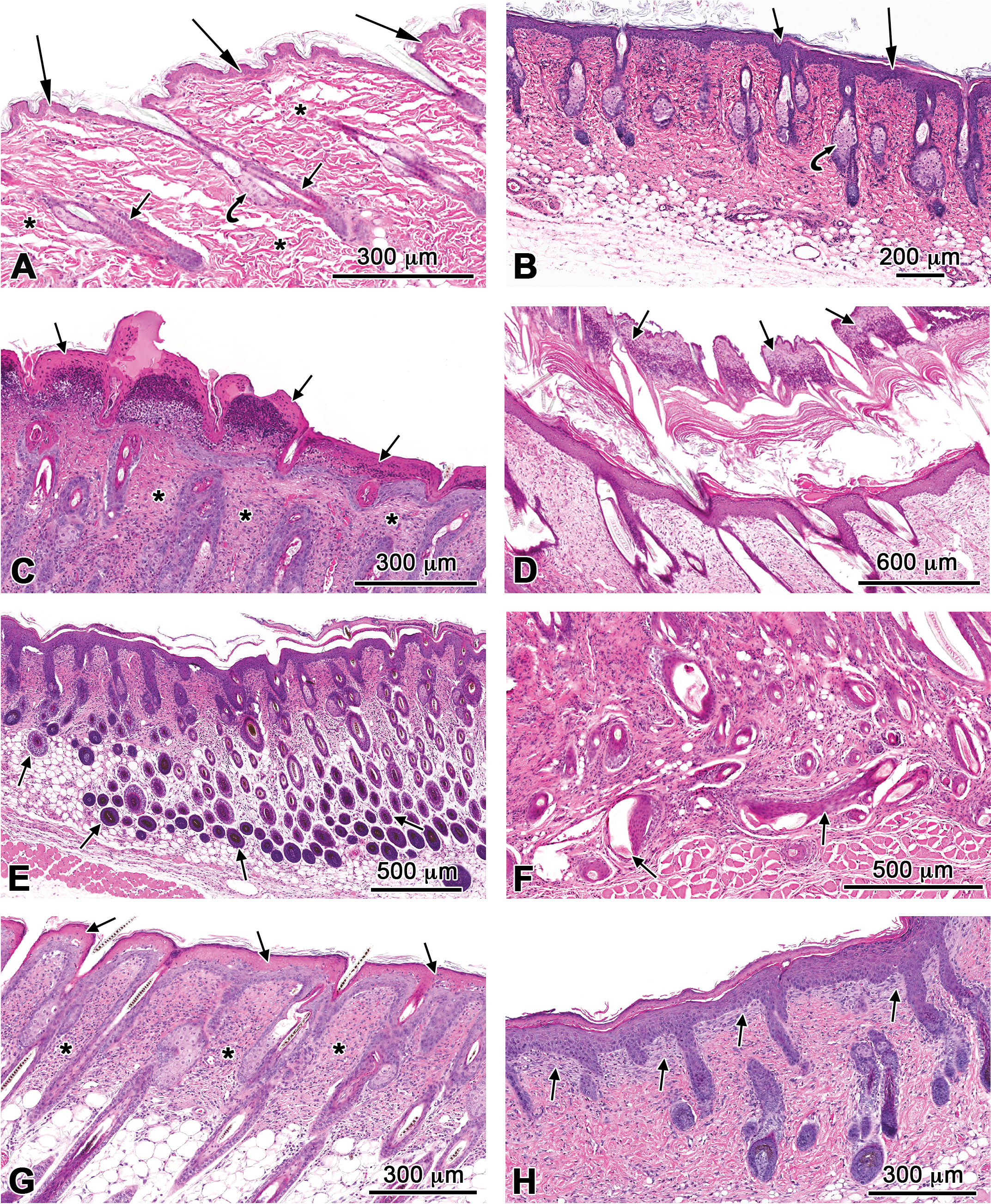
Histopathology of skin lesions at the site of application from the 14-day studies of cumene hydroperoxide in rats and mice. (A) Normal skin from a control rat. Epidermis (long arrows), hair follicles (short arrows), sebaceous glands (curved arrow), dermis (asterisks). (B) Epidermal hyperplasia in a male rat characterized by moderate thickening of the epidermis (long arrow) that extends into the hair follicles (short arrow). Note the hyperplastic sebaceous glands (curved arrows). (C) Exudate in a female mouse characterized by accumulation of a layer of eosinophilic proteinaceous fluid, keratin, and degenerate neutrophils on the surface of the epidermis (arrows). Note inflammatory cells within the dermis (asterisks). (D) Exudate in a male rat characterized by marked accumulation of successive layers of proteinaceous fluid, keratin, and degenerate neutrophils on the surface of the hyperplastic epidermis (arrows). (E) Hair follicle hyperplasia in a female mouse characterized by increased number of follicles within the dermis extending into the subcutis (arrows). (F) Aberrant hair follicle growth (arrows) in a male rat. Note deformed hair follicles growing perpendicular to the orientation of hair follicles. (G) Necrosis of the superficial epidermis characterized by hypereosinophilia and loss of cellular detail (arrows). Note inflammatory cells throughout the dermis (asterisks). (H) Fibrosis (arrows) in the papillary dermis of a male rat.
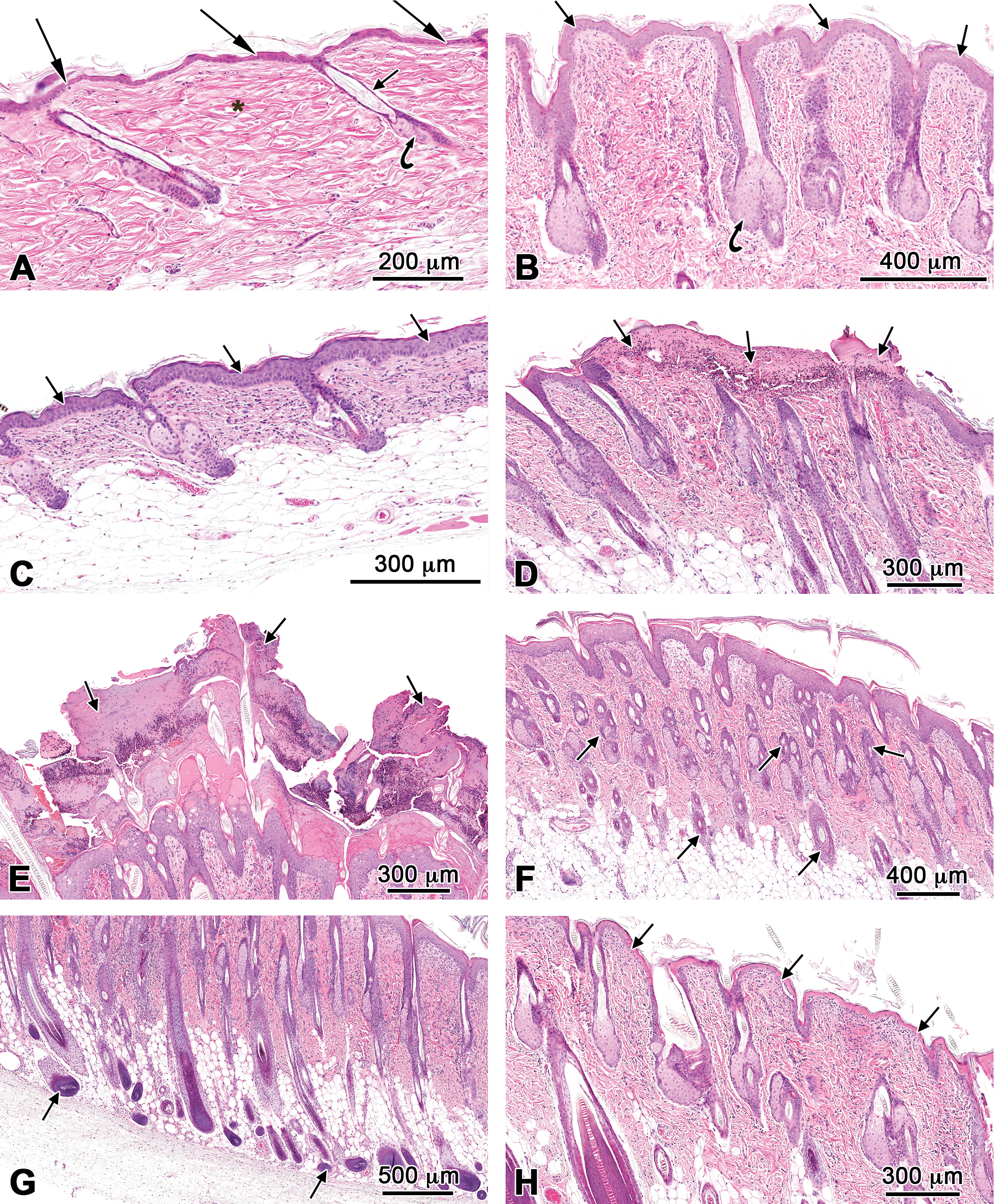
Histopathology of skin lesions at the site of application from the 90-day studies of cumene hydroperoxide in rats and mice exposed to 12-mg/kg cumene hydroperoxide. Lesions in male and female rats are generally similar to those observed in the 14-day studies. (A) Normal skin from a control rat. Epidermis (long arrows), hair follicles (short arrow), sebaceous glands (curved arrow), and dermis (asterisk). (B) Epidermal hyperplasia (arrows) and sebaceous gland hyperplasia (curved arrow) in a male rat. Note inflammatory cells throughout the dermis. (C) Epidermal hyperplasia (arrows) and chronic active inflammation in the dermis of a female mouse. (D) Exudate (arrows) on the epidermal surface in a female rat. Note complete loss of the underlying epidermis (ulcer). (E) Marked exudate (arrows) on the epidermis surface of a female. (F) Follicular hyperplasia (arrows) in a female rat. Note epidermal hyperplasia and sebaceous gland hyperplasia. (G) Aberrant follicular hyperplasia (arrows) in a female rat. (H) Necrosis of the superficial epidermis (arrows) in a male rat.
In both the 14-day and 90-day studies, epidermal hyperplasia was a prominent lesion at the site of application in most animals in both species and sexes. Hyperplasia was characterized by an increase in the thickness of the epidermis, thickened beyond the normal 1 to 2 nucleated cell layers typical of the skin of the vehicle controls (Figures 1A and 2A) to 5 or more cell layers (Figures 1B, 2B, and 2C). Invariably, the hyperplastic process extended into and involved the epithelium of the hair follicles. Additionally, there was hyperkeratosis characterized by a variable increase in the thickness of the keratin layer or stratum corneum of the skin.
Microscopically, exudate correlated with the gross lesion characterized as a “crust” that covered portions of the site of application. The character of the exudate varied. In mice, particularly, exudate occurred as an accumulation of a serocellular layer adherent to the epidermal surface and consisted of a mixture of eosinophilic proteinaceous fluid, keratin and numerous degenerate inflammatory cells, and primarily neutrophils (Figures 1C and 2D). More so in rats than in mice, exudate sometimes occurred as successive accumulations of 2 to 3 layers of dead tissue, each layer consisting of thickened keratin (ortho- and parakeratotic hyperkeratosis), necrotic epidermis and hair follicles, mixed with masses of degenerate inflammatory cells, and primarily neutrophils (Figures 1D and 2E). The exudate was loosely adhered to, or detached from, the underlying necrotic or viable (intact) epidermis.
The viable epidermis beneath and often adjacent to areas of exudate was thickened (epidermal hyperplasia) beyond the 1 to 2 nucleated cell layers typical of the skin of the vehicle controls. Invariably, epidermal hyperplasia extended into and involved the epithelium of the hair follicles. Focal aggregates of viable and degenerate neutrophils (pustules) often occurred within the exudate (Figure 1C).
A primary change was hair follicle hyperplasia characterized by a marked increase in the length and number of hair follicle profiles compared with unaffected sites or the skin of the vehicle-treated control rats (Figures 1E and 2F). Affected hair follicles frequently extended to abut the subcutaneous (panniculus) muscle. In the more severely affected skin at the site of application, there was often aberrant hair follicle growth (Figures 1F and 2G). In these sites, the orientation of hair follicles was often haphazard with many hair shafts growing horizontally and at odd angles in the dermis. Many hair follicles were abnormally shaped and enlarged with bending of the hair bulbs, as they reached the panniculus muscle. Often observed within the spectrum of hair follicle changes was abortive hair follicle development manifested as irregularly shaped blobs of keratin. Sebaceous gland hyperplasia often accompanied hair follicle hyperplasia and consisted of enlarged glands due to an increase in the number and size of the acinar cells (Figures 1B and 2B).
Epidermal necrosis was diagnosed when the epidermis beneath or adjacent to areas of exudate was hypereosinophilic and had loss of cellular detail (Figures 1G and 2H). Frequently, there was sloughing of the necrotic epidermis. Ulcers were characterized by complete loss of the epidermis (Figure 1C). Epidermal degeneration was diagnosed when areas of the viable epidermis contained focal areas in which the cells were disorganized and swollen and had clear or hyalinized cytoplasm, sometimes accompanied by evidence of intraepidermal edema that formed microvesicles.
Dermal chronic active inflammation in rats and mice consisted of diffuse infiltrates of variable numbers of mononuclear leukocytes (lymphocytes and macrophages), eosinophils, and neutrophils in the dermal interstitium with extension into the subcutis (Figures 1C, 1G, 2B, and 2C). Neutrophils tended to predominate in areas where the overlying epidermis was necrotic or ulcerated. Dermal fibrosis consisted of an increased density of collagen fibers and fibroblasts within the superficial dermis and around the adnexa, particularly in association with hyperplastic hair follicles (Figure 1H).
Genotoxicity Testing
CHP was positive in bacterial mutagenicity studies, although results differed between the 2 laboratories (Tables 3 and 4). In 1 study, CHP (5–200 µg/plate) was mutagenic in E. coli strain WP2 uvrA/pKM101 with and without rat liver S9 activation, and no mutagenic activity was observed in S. typhimurium strains TA98 and TA100 with or without S9. In the 2nd study, positive results were obtained with CHP (1–100 µg/plate) in S. typhimurium strains TA98 and TA100 with S9 activation (rat or hamster liver) only (Mortelmans et al. 1986). In vivo, no increases in the frequencies of micronucleated erythrocytes (biomarkers of numerical or structural chromosomal damage) were observed in peripheral blood of male or female mice sampled at the end of the 90-day dermal toxicity study with CHP. In addition, exposure to CHP did not cause changes in the proportion of polychromatic (immature) erythrocytes among total erythrocytes in the blood of either males or females, suggesting no bone marrow toxicity.
Mutagenicity of Cumene Hydroperoxide in Bacterial Tester Strains (Study 1).
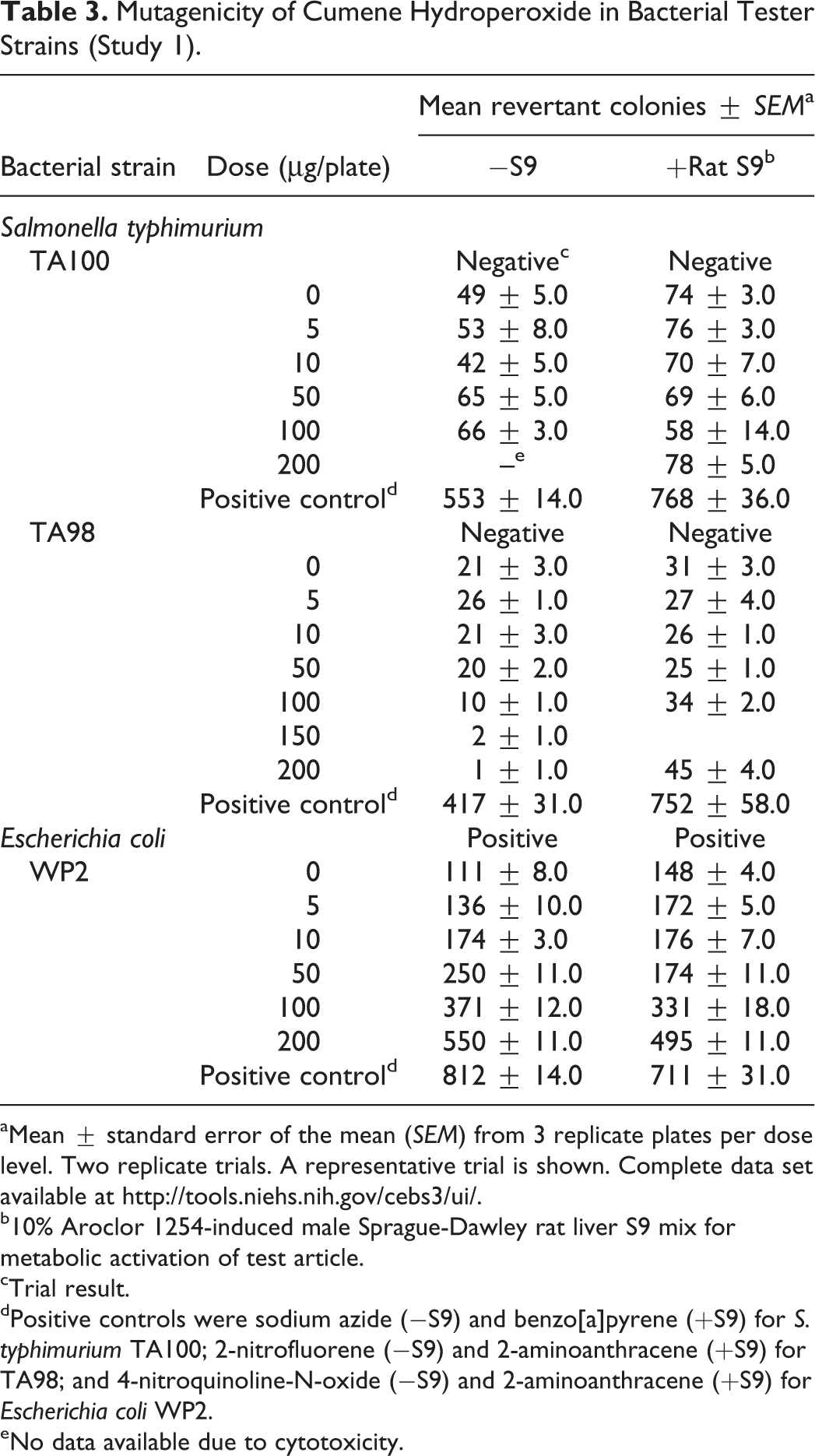
aMean ± standard error of the mean (SEM) from 3 replicate plates per dose level. Two replicate trials. A representative trial is shown. Complete data set available at http://tools.niehs.nih.gov/cebs3/ui/.
b10% Aroclor 1254-induced male Sprague-Dawley rat liver S9 mix for metabolic activation of test article.
cTrial result.
dPositive controls were sodium azide (−S9) and benzo[a]pyrene (+S9) for S. typhimurium TA100; 2-nitrofluorene (−S9) and 2-aminoanthracene (+S9) for TA98; and 4-nitroquinoline-N-oxide (−S9) and 2-aminoanthracene (+S9) for Escherichia coli WP2.
eNo data available due to cytotoxicity.
Mutagenicity of Cumene Hydroperoxide in Bacterial Tester Strains (Study 2).
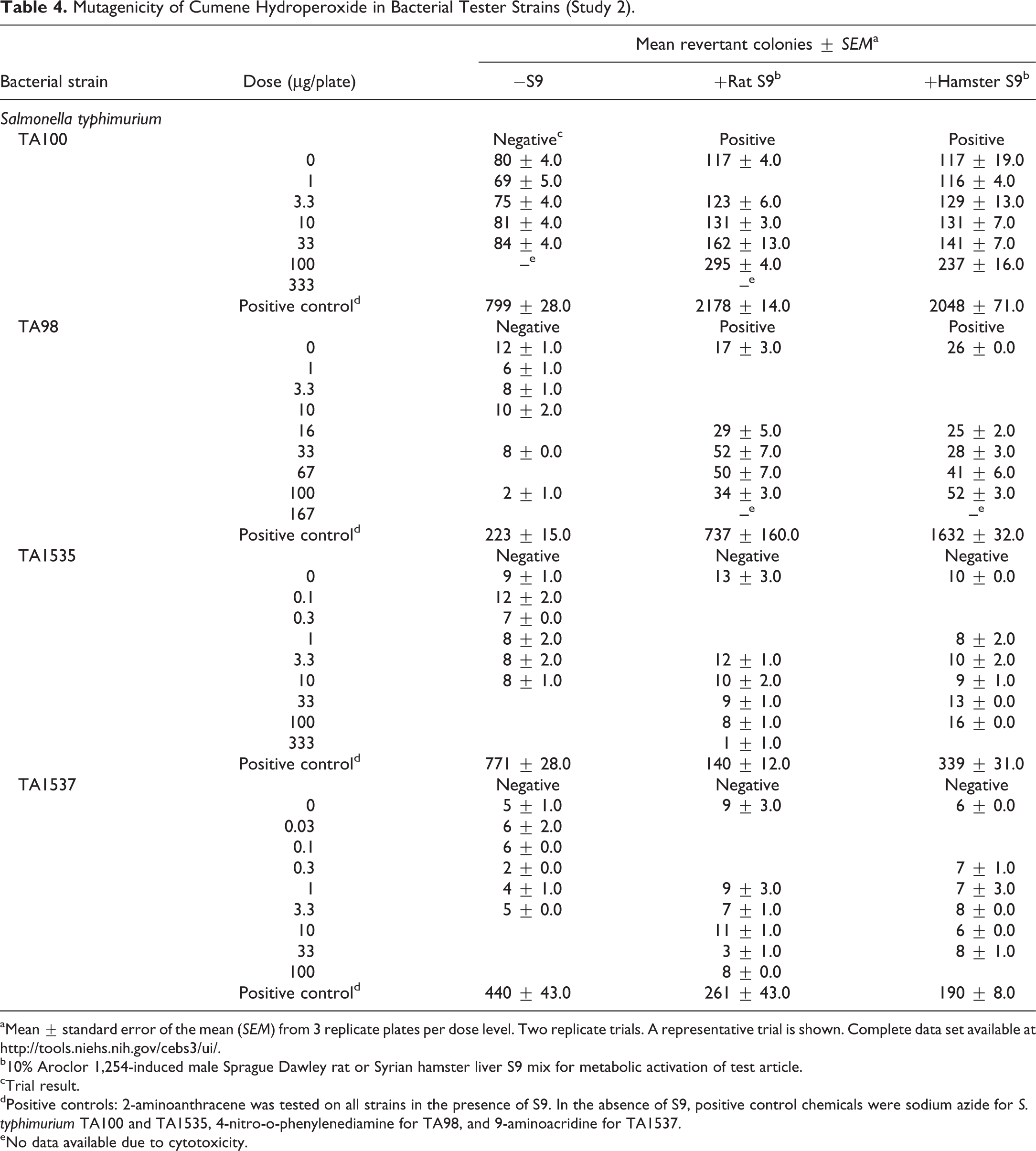
aMean ± standard error of the mean (SEM) from 3 replicate plates per dose level. Two replicate trials. A representative trial is shown. Complete data set available at http://tools.niehs.nih.gov/cebs3/ui/.
b10% Aroclor 1,254-induced male Sprague Dawley rat or Syrian hamster liver S9 mix for metabolic activation of test article.
cTrial result.
dPositive controls: 2-aminoanthracene was tested on all strains in the presence of S9. In the absence of S9, positive control chemicals were sodium azide for S. typhimurium TA100 and TA1535, 4-nitro-o-phenylenediamine for TA98, and 9-aminoacridine for TA1537.
eNo data available due to cytotoxicity.
Free Radical Generation
We used an ESR spectrometer and an in vivo spin-trapping technique with POBN to investigate free radical production in the present murine model. Plasma and tissue studies were conducted to detect free radicals caused by exposing rat skin to CHP at low- (3 mg/kg) and high-dose (12 mg/kg) concentrations for 90 days. Ascorbyl radical (Buettner and Jurkiewicz 1993) was detectable in rat plasma after the 90-day CHP treatment, but when the ESR signal intensities were measured, statistically significant differences between controls and CHP-treated rats were not observed at either dose. In addition, we were not able to detect ESR signals from free radicals in tissue lipid extracts of different organs (data not shown).
Markers of Antioxidant Status
Although the plasma level of ascorbic acid tended to be lower in the group exposed to 3-mg/kg CHP, significant effects of CHP exposure on ascorbic acid in plasma were not observed for either dose (Table 5). Concentrations of uric acid in plasma are shown in Table 5. Uric acid had a slight, but not significant, dose-dependent increase. Compared with control rats at the same time point, rats administered CHP exhibited no change for the low dose and showed a nonsignificant decrease in plasma total GSH level for the high dose (Table 5). Glutathione acts as an antioxidant both intracellularly and extracellularly in combination with enzymes that reduce hydroperoxides, as GSH is oxidized to GSSG and other mixed disulfides. In this study, neither GSH nor GSSG changed significantly (data not shown). The ratio of GSH to GSSG would be expected to be a more sensitive marker of oxidative stress, but there was no evidence of change in the ratio for either dose of CHP (data not shown).
Effect of Cumene Hydroperoxide (CHP) on Plasma Concentration of Antioxidants in Male F344/N Rats.
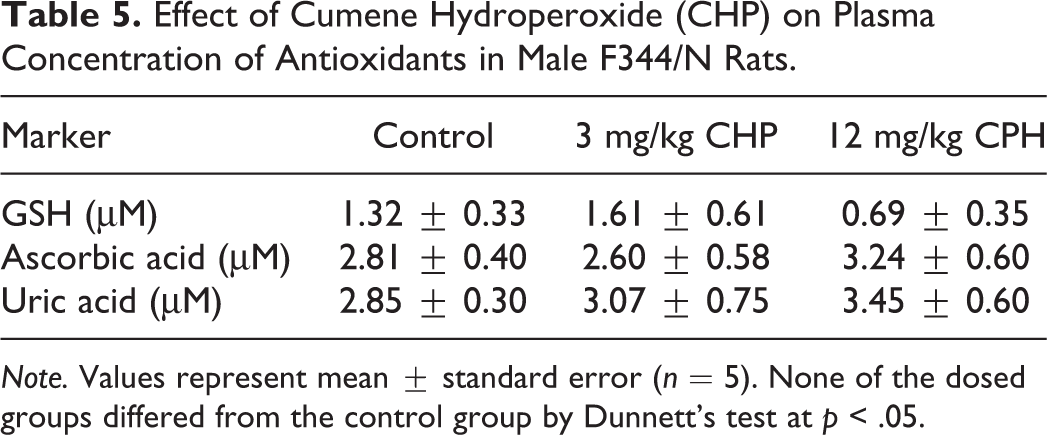
Note. Values represent mean ± standard error (n = 5). None of the dosed groups differed from the control group by Dunnett’s test at p < .05.
Discussion
CHP is a high production volume chemical that has multiple industrial applications and a dearth of available toxicity data. It was selected for study by the NTP as an organic peroxide class representative based on potential for exposure, lack of toxicity data, and the likelihood that free radicals formed as reaction products of CHP are expected to be relatively stable, based on the parent structure. CHP is an alkyl hydroperoxide and is metabolized by cytochrome P450 via hemolytic cleavage of the O–O bond, producing singlet oxygen, hydroxyl radical, and other radical species that can be characterized by ESR spin-trapping techniques (Chiarpotto et al. 1984; O’Brien 1988; Taffe et al. 1987). In our study, dermal exposure to CHP was used as a model system to explore the link between free radical formation, oxidative stress, and toxicity.
The histopathological lesions observed in mice and rats were indicative of skin irritation and were limited to the dermis and epidermis at the site of application. Lesions were consistent across species and sex and were characterized by inflammation and hyperplasia. These findings were comparable to results from 14-day dermal toxicity studies of tert-butyl hydroperoxide in F344/N rats and B6C3F1/N mice (Behl et al. 2012) and 4-week dermal toxicity studies of 9 organic peroxides, in which benzoyl peroxide, t-butyl peroxybenzoate, p-t-butyl isopropylbenzene hydroperoxide, CHP, dicumyl peroxide, and methyl ethyl ketone peroxide induced sustained hyperplasia in the epidermis at the site of application (Hanausek et al. 2004).
In the current study, CHP was found to be mutagenic in certain Salmonella strains and E. coli WP2, although the results were not consistent across tests (Tables 3 and 4). These results are in accordance with other studies that have found CHP (Levin et al. 1982) and other alkyl hydroperoxides (Ball et al. 1992) to be genotoxic in some Salmonella strains and not others. Since CHP exposure did not result in chromosomal damage as measured by the production of micronuclei in mouse erythrocytes following a 90-day exposure, it is possible that CHP elicits DNA damage without causing chromosomal damage. Alternatively, the lack of positive response in the in vivo micronucleus assay could result from a lack of systemic response to CHP. Consistent with the lack of in vivo genotoxic effects observed in the current study, CHP did not produce an increase in oxidative damage in DNA as measured by 8-hydroxy-deoxyguanosine in treated skin of SENCAR mice in a study comparing carcinogenic potential of multiple organic peroxides (Hanausek et al. 2004). Furthermore, CHP did not induce tumors in a lifetime skin-painting study with Swiss Millerton mice (Van Duuren, Orris, and Nelson 1965).
According to the theory of multistage carcinogenesis, an initiation phase involving irreversible genetic damage is followed by a promotion phase that requires clonal expansion of initiated cells (Digiovanni 1992). Furthermore, sustained hyperplasia in mouse skin is strongly associated with tumor-promoting potential (Digiovanni 1992; Gimenez-Conti et al. 1991; Slaga et al. 1982). As in the current study with CHP, many alkyl peroxides have been shown to induce hyperplasia following dermal exposure (Hanausek et al. 2004; Slaga et al. 1981). For example, using a 2-stage (initiation–promotion) model in mice with dimethylbenzanthracene as the initiator, repeated CHP treatment induced progression of benign skin tumors to carcinoma in 2 separate studies (Bhasin, Kauser, and Athar 2004; Murray et al. 2007).
In addition to assessing toxicity, biomarkers of oxidative stress were measured to evaluate oxidative potential of CHP on tissues. Many studies assessing the oxidative status of biological systems have used detection of endogenous long-lived free radicals as markers of oxidative stress, such as ascorbyl radical (Spasojevic 2011). Ascorbyl radical is one of the most stable radicals that can be detected by ESR spectroscopy (Buettner and Jurkiewicz 1993). Studies have shown that topical application of CHP to murine skin can generate ascorbyl radical detectable by ESR (Timmins and Davies 1993a). Other reports have demonstrated the CHP-mediated production of free radicals in vitro (Athar et al. 1989; Taffe et al. 1987; Timmins and Davies 1993a) and in vivo (Shvedova et al. 2002). In vivo free radical production was also assessed at the site of administration immediately following application (Shvedova et al. 2002). Although such results strongly imply free radical generation by CHP, we were not able to detect them in lipid extracts of tissues from various organs or in plasma following repeated dermal application of CHP over a 90-day period.
Despite their reactivity, free radicals can react selectively with targets at critical cell locations and may diffuse to some distance from the site of their generation (Buettner and Jurkiewicz 1996). It should be mentioned that although the involvement of ascorbyl radical in biological reactions is extensive, animal and plant cells contain a reduced nicotinamide adenine dinucleotide (NADH)-dependent semidehydroascorbate reductase to remove this radical (Lane and Lawen 2009). Since ascorbyl radical release is dependent on ascorbate concentration, the observation that there was no ESR evidence of ascorbyl radical in this study may imply a lack of effect of CHP on the concentration of ascorbate measured in plasma or low free radical generation that is not detectable by ESR. Most animals, including rats, synthesize ascorbate and because exogenous ascorbate confers protection against oxidative stress, we tested whether endogenous ascorbate levels in plasma would be affected by CHP exposure. In addition, the high cellular concentration of GSH makes it difficult to isolate the antioxidant role of ascorbate, especially because GSH has been shown to recycle ascorbate from its oxidized form, dehydroascorbate. In the present study, we show that plasma levels of ascorbate and GSH in rats administered CHP were not significantly different from the controls.
Some studies have shown that CHP may exert toxic effects on several cells and tissues (Anari, Khan, and OBrien 1996; Poot et al. 1987; Vroegop, Decker, and Buxser 1995). For example, in vitro studies with CHP (10–200 µM) resulted in redox imbalance of murine peritoneal macrophages, with significant release of proinflammatory cytokines (tumor necrosis factor-α, interleukin-1β, and prostaglandin E2; Kaur, Alam, and Athar 2009). In vitro studies with keratinocytes (HaCaT) demonstrated expression of cyclooxygenase-2 (COX-2) and increased levels of prostaglandin E2 following 18-hr treatment with CHP (Shvedova et al. 2004). Furthermore, inhibitors of COX-2 were able to suppress oxidative stress and enzyme expression in the cells treated with CHP, leading the authors to suggest involvement of COX-2 in CHP-induced inflammatory responses (Shvedova et al. 2002). Other authors have reported an antioxidant role for endogenously produced ascorbate that protects isolated rat hepatocytes from CHP-mediated oxidative stress (Chan et al. 2005). While these studies suggest that CHP has the potential to elicit oxidative stress and an inflammatory response, compatible with the lesions observed at the site of application in the current study, the absence of free radical detection and lack of plasma ascorbate and GSH changes in this report do not support the hypothesis of a stable free radical capable of eliciting systemic damage.
In summary, findings from these studies indicate that topical skin exposure to CHP causes mainly skin lesions characterized by inflammation and hyperplasia at the site of application. Therefore, further testing of CHP in chronic studies is not currently being pursued.
Footnotes
Acknowledgments
The authors thank Kay Crissman and Dr. Gary Hatch for their technical assistance with the antioxidant assays. The authors would also like to thank Drs. Mamta Behl and Chad Blystone for their review of this manuscript and Dr. Ann Motten for her editorial assistance.
Author Contribution
Authors contributed to conception or design (PC, GT, and MK); data acquisition, analysis, or interpretation (CR, RH, GK, LF, MH, KW, SW, GT, and MK); drafting the manuscript (CR); and critically revising the manuscript (CR, PC, RH, GK, LF, MH, KW, SW, GT, and MK). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Authors’ Note
Po Chan has retired from National Institute of Environmental Health Sciences.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the NIH, National Institute of Environmental Health Sciences.