
Abstract
The female reproductive cycle is orchestrated by cyclical and coordinated hormonal changes under the direction of the hypothalamic–pituitary–gonadal (HPG) axis. Any disruption of the HPG axis may lead to functional and structural alterations in the female reproductive system. Test article–related disturbances in the estrous cycle can be recognized in nonclinical toxicity studies by staging the cycle based on microscopic evaluation of female reproductive organs. In chronic rat toxicity studies, an additional complication is the development of reproductive senescence, which is associated with natural alterations in the reproductive cycle leading to changes in the female reproductive system that can potentially be confused with test article effects. The current article describes the features of persistent estrus, one stage of reproductive senescence, in middle-aged Sprague-Dawley rats and discusses elements to help differentiate senescence from induced effects.
Rats are spontaneous ovulators, and ovulation occurs at a regular interval of 4 or 5 days in young adult female rats. With aging, reproductive senescence in female rats proceeds through sequential reproductive stages before ending in anestrus. The sequence of reproductive states during aging has been well characterized by examining histologic features of the stages of reproductive senescence (Westwood 2008) and vaginal smears (LeFevre and McClintock 1988). In brief, female rats undergo a transition from regular estrous cycles to irregular cycles, followed by persistent estrus, repetitive pseudopregnancy, persistent diestrus, and finally, anestrus. Various studies in female Sprague-Dawley rats have shown that they begin to develop irregular cycles upon reaching approximately 7 to 8 months of age although the data vary somewhat from study to study (LeFevre and McClintock 1988; Vom Saal, Finch, and Nelson 1994). More recently, Rudmann and Foley (2013) pointed out that Sprague-Dawley and Wistar female rats start entering reproductive senescence as early as 5 months of age.
The female reproductive cycle reflects cyclical and coordinated hormonal changes orchestrated by the hypothalamic–pituitary–gonadal (HPG) axis. Any disruption of the HPG axis may lead to functional and structural alterations in the reproductive system of the female rodent (Rudmann and Foley 2013). During nonclinical toxicity testing in female rats, test article–related disturbances in the estrous cycle can be recognized by staging the cycle based on microscopic evaluation of the female reproductive system (Westwood 2008). In longer-term rat toxicity studies, however, these evaluations may be complicated by the development of reproductive senescence due to natural alterations in the reproductive cycle with features that can overlap with test article–induced effects. In order to facilitate the differentiation of induced versus background aging changes in the female rat reproductive system, it is necessary for the pathologist to understand when age-related findings might occur and what those changes might look like.
Here, we describe the characteristic features and incidence of histologic findings indicative of persistent estrus that were observed in vehicle control rats in a long-term nonclinical toxicity study using Sprague-Dawley rats.
Female Sprague-Dawley rats were obtained from Charles River Laboratories (NC, USA) at approximately 10 to 11 weeks of age. Rats were housed individually in polycarbonate cages under standard vivarium conditions. Temperature (68°–79°F) and lighting (approximate 12-hr light, 12-hr dark cycle) in the animal rooms were controlled throughout the study. Rats had free access to municipal drinking water and to a commercial rodent diet (Certified Rodent Diet 5002, PMI Feeds, Inc., MN, USA) certified to be free of estrogenic compounds. In a 6-month exploratory toxicity study, a total of 30 female vehicle control rats were assigned to 3 groups of 10 rats each and administered 0.5% methylcellulose daily by oral gavage at a volume of 10 ml/kg/day for 29 or 57 days. At the conclusion of the dosing periods, 10 rats from groups 1 and 2 were euthanized on day 30 or day 58, respectively. The remaining group of 10 rats was withdrawn from treatment after the last dose on day 58 and observed for 4 months (122 days). All procedures were approved by the institutional animal care and use committee and performed in compliance with the US Animal Welfare Act, the Guide for the Care and Use of Laboratory Animal, and federal regulations. The ovaries, oviducts, uterus, cervix, and vagina were collected and processed for microscopic examination. The histologic stage of the estrous cycle was identified based on a practical histological guide to staging (Westwood 2008).
Microscopic examination revealed an absence of basophilic corpora lutea, reduced number and size of corpora lutea, and increased antral follicles in the ovary (Figure 1) compared with normal ovaries at estrus (Figure 2), along with characteristics of estrus in the uterus (Figure 3), cervix, and vagina (Figure 4) in 0/10, 1/10, and 4/10 rats on days 30, 58, and 180, respectively. These microscopic findings were indicative of a state of persistent estrus. Corresponding ages of rats euthanized on days 30, 58, and 180 were approximately 3.5, 4.5, and 8.5 months old, respectively. The development of a persistent estrus pattern in early reproductive senescence is known to be associated with the attenuation of gonadotropin-releasing hormone (GnRH)/luteinizing hormone (LH) surge (Downs and Wise 2009; Gore et al. 2000). Disruption of neural signals that regulate the pattern of GnRH release at the level of the hypothalamus triggers a failure to maintain regular estrous cycles (Downs and Wise 2009). Impairment of the preovulatory GnRH/LH surge allows ovarian follicles to persist and continue to produce estrogen. Concurrently, corpora lutea that produce progesterone do not develop. These hormonal changes consequently drive the reproductive tract (uterus and vagina) to a state of persistent estrus.
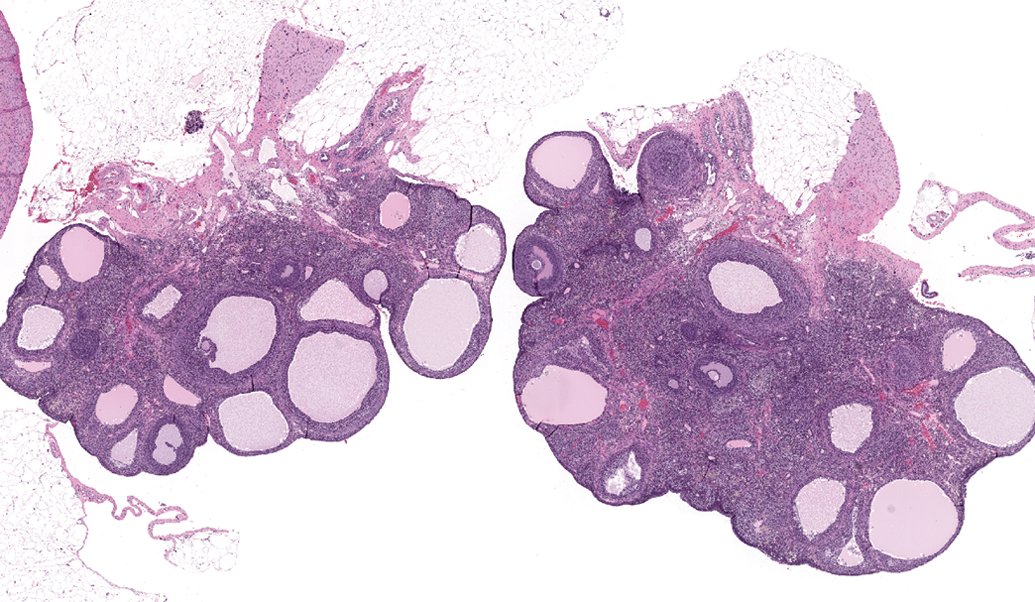
Sprague-Dawley rat, 8.5 months old. Photomicrograph of reproductive organs indicative of a state of persistent estrus: Ovaries characterized by increased antral follicles, absence of basophilic corpora lutea, and decreased corpora lutea. H&E = hematoxylin and eosin stain. Original magnification 16×.
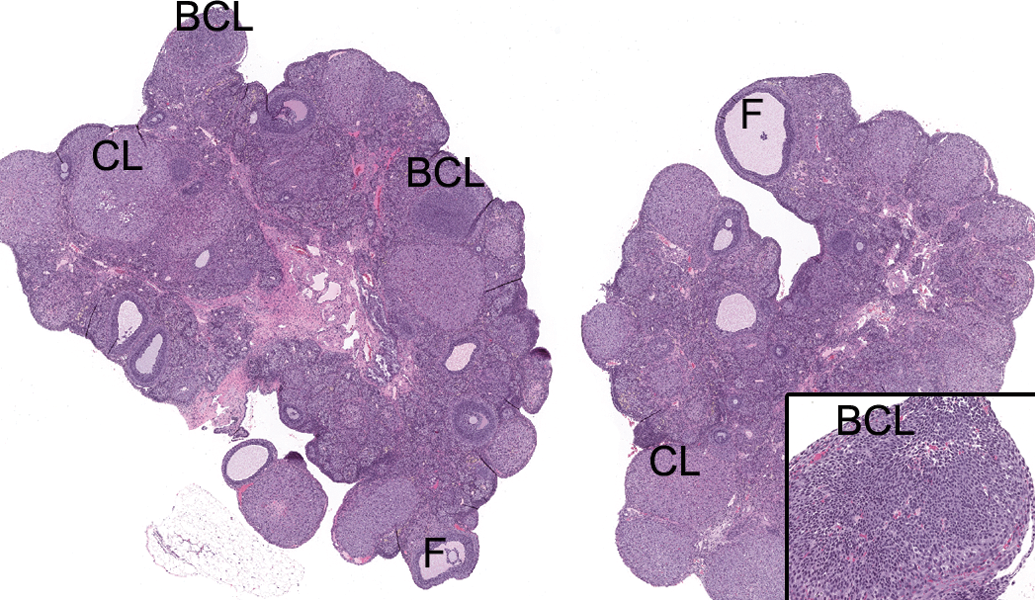
Sprague-Dawley rat, 8.5 months old. Photomicrograph of normal ovaries at estrus. BCL = basophilic corpus luteum; CL = corpus luteum; F = antral follicle; H&E = hematoxylin and eosin stain. Original magnification 16×. Inset: Higher magnification of BCL. Original magnification 100×.
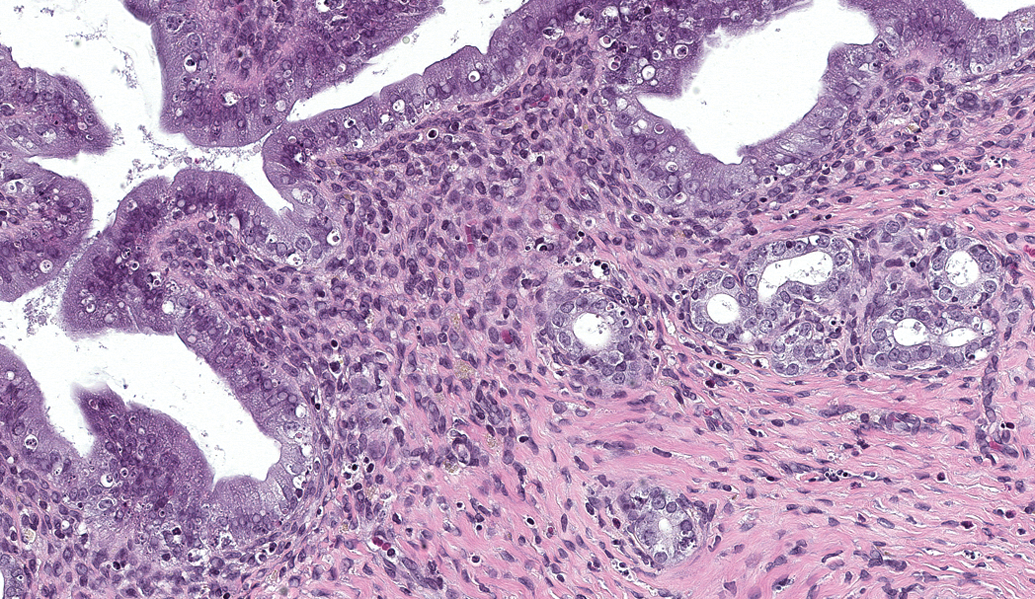
Sprague-Dawley rat, 8.5 months old. Photomicrograph of reproductive organs indicative of a state of persistent estrus: Uterus consisting of tall columnar epithelium with frequent vacuolar degeneration and single cell necrosis in luminal and glandular epithelium, and rare mitoses. H&E = hematoxylin and eosin stain. Original magnification 200×.
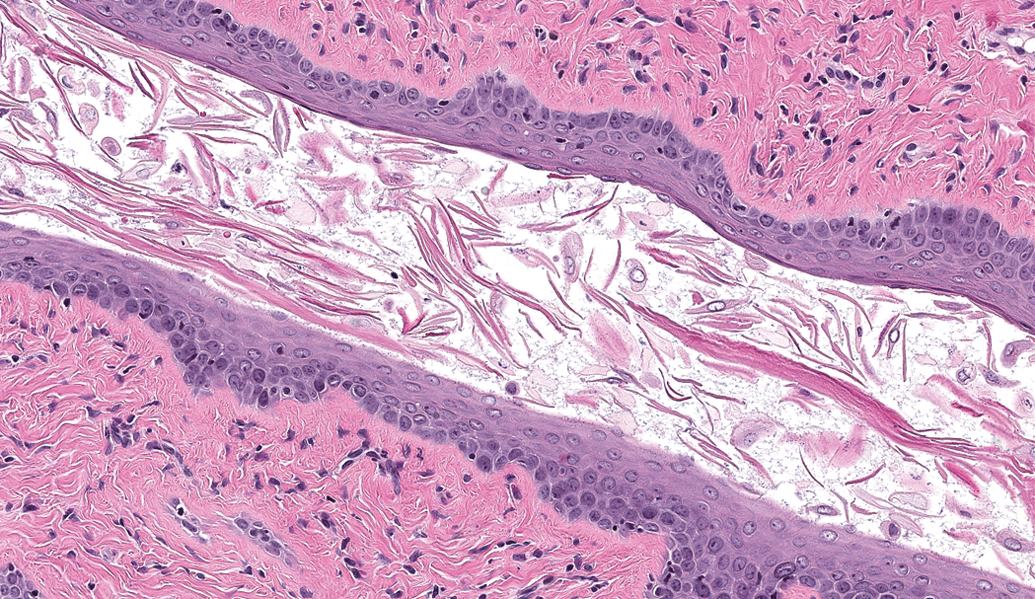
Sprague-Dawley rat, 8.5 months old. Photomicrograph of reproductive organs indicative of a state of persistent estrus: Vaginal mucosa lined by cornified squamous epithelium. A defined stratum corneum has been sloughed off. H&E = hematoxylin and eosin stain. Original magnification 200×.
The state of persistent estrus in early reproductive senescence may be differentiated from reproductive changes induced via pharmacologic effects on the HPG axis based on the character and pattern of the changes. For example, persistent vaginal cornification and increased large follicles that overlap, in part, with features of persistent estrus in early reproductive senescence have been reported to occur in young adult rats treated with antiprogestins. The ovaries from antiprogestin-treated rats, however, contained previously formed corpora lutea that were hypertrophied unlike those typical of early reproductive senescence (Tamura et al. 2009; Ruiz et al. 1996). Administration of an estrogen analogue ethinylestradiol or estradiol benzoate to young adult rats resulted in estrogenic effects on the vagina and uterus and increases in early stage follicles along with apoptotic corpora lutea or large basophilic corpora lutea in the ovary (Andrews et al. 2002; Ferre et al. 2013), the ovarian features of which do not match those of early reproductive senescence. Finally, administration of centrally acting agents such as chlorpromazine and sulpiride to young adult rats has been shown to produce not only increased large follicles in the ovary but also epithelial mucification and/or atrophy in the vagina (Ishii et al. 2009; Izumi et al. 2009). These vaginal patterns of change are inconsistent with the state of persistent estrus. However, in some cases, it may not be possible to easily discriminate the onset of reproductive senescence from an effect of a xenobiotic. For instance, atrazine, a chlorotriazine herbicide, inhibits the GnRH-induced LH surge in Sprague-Dawley rats, resulting in changes that mimic normal reproductive aging (Cooper et al. 2005).
The interpretation of histopathologic findings in the female reproductive tract can be difficult in the context of early reproductive senescence. One reason for this is that the incidence of irregular estrus cycles can abruptly increase in middle-aged female rats (Lu et al. 1979), although another is that there can be significant individual differences in progression through reproductive senescence. This latter point was characterized nicely in a longitudinal study by LeFevre and McClintock (1988) where they observed only about half of the female Sprague-Dawley rats developing persistent (constant) estrus while the remainder continued to exhibit irregular cycles prior to entering into persistent diestrus. These individual differences in reproductive aging and variability in the timing of the transition to reproductive senescence may influence the histopathologic incidence of background age-related reproductive findings. This is especially true in toxicology studies of ≥3 months duration, in which animals approach the age at which reproductive senescence is known to begin (middle age). In addition, nutritional factors may also be implicated in the onset of reproductive senescence. Severe food deprivation has been shown to provoke irregular estrous cycles and atrophy of the ovary in female Sprague-Dawley rats (Seki et al. 1997). Therefore, test article–related effects on food consumption and body weight must also be considered when interpreting estrous cycle and structural alterations noted in toxicology studies. These factors need to be taken into consideration when designing a study and interpreting cross-sectional data. The current report will help in the interpretation of histopathologic data by providing examples of female reproductive changes that can be used to distinguish persistent estrus of reproductive senescence from xenobiotic effects in middle-aged Sprague-Dawley rats and discussing additional factors to consider in the interpretation.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Author Contribution
Authors contributed to conception or design (NS, CH, MM); data acquisition, analysis, or interpretation (NS, CH, MM); drafting the manuscript (NS); and critically revising the manuscript (NS, CH, MM). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.