
Abstract
A majority (∼80%) of human malignant mesotheliomas are asbestos-related. However, non-asbestos risk factors (radiation, chemicals, and genetic factors) account for up to 30% of cases. A recent 2-year National Toxicology Program carcinogenicity bioassay showed that male F344/N rats exposed to the industrial toxicant vinylidene chloride (VDC) resulted in a marked increase in malignant mesothelioma. Global gene expression profiles of these tumors were compared to spontaneous mesotheliomas and the F344/N rat mesothelial cell line (Fred-PE) in order to characterize the molecular features and chemical-specific profiles of mesothelioma in VDC-exposed rats. As expected, mesotheliomas from control and VDC-exposed rats shared pathways associated with tumorigenesis, including cellular and tissue development, organismal injury, embryonic development, inflammatory response, cell cycle regulation, and cellular growth and proliferation, while mesotheliomas from VDC-exposed rats alone showed overrepresentation of pathways associated with pro-inflammatory pathways and immune dysfunction such as the nuclear factor kappa-light-chain-enhancer of activated B cells signaling pathway, interleukin (IL)-8 and IL-12 signaling, interleukin responses, Fc receptor signaling, and natural killer and dendritic cells signaling, as well as overrepresentation of DNA damage and repair. These data suggest that a chronic, pro-inflammatory environment associated with VDC exposure may exacerbate disturbances in oncogene, growth factor, and cell cycle regulation, resulting in an increased incidence of mesothelioma.
Keywords
Introduction
Vinylidene chloride (1,1, diethylene chloride, VDC) is a comonomer widely used in the manufacture of synthetic polymers such as polyvinyl chloride (PVC), acrylonitrile, or acrylates, for the production of conduits and pipes, plastic profiles, films and sheeting, cables and cable sheeting, and flooring (Brandsch and Piringer 2008). Polymerization of VDC was originally used to produce flexible films for food covering (“Saran” wrap); however, this use has been discontinued due to concerns over decomposition at higher temperatures (>125°C), which leads to dehydrochlorination of the polyvinylidene chloride compound and release of hydrogen chloride (HCl; Wessling 1977). VDC is considered an eye, skin, and throat irritant; central nervous system depressant; and a potential carcinogen by the National Institute of Occupational Safety and Health (NIOSH 2010). Target tissues include liver, kidney, skin, respiratory system, and nervous system, although these vary by species and route of exposure (NIOSH 2010). Mice are more susceptible than rats to the toxic and carcinogenic effects due to differences in metabolic capacity (Hathway 1977; Jones and Hathway 1978). In rats, at lower doses, VDC is conjugated by glutathione and converted to metabolites (N acetyl S-(2 hydroxy ethyl) cysteine, thiodiglycolic acid, and chloroacetic acid), which are then excreted in urine. At higher doses, due to glutathione saturation, a percentage of VDC is excreted unchanged through expiration from the lungs, and some percentage is retained in lungs, liver, and kidneys (Hathway 1977; Jones and Hathway 1978). Results of a recent 2-year National Toxicology Program (NTP) carcinogenicity bioassay showed that exposure of male F344/N rats to VDC resulted in a marked treatment-related increase in the incidence of mesotheliomas compared to concurrent historical controls (NTP in press-a). Additionally, mesothelioma of the pleura and peritoneum occurred in 2 female F344/N rats. Aged control male F344/N rats develop spontaneous mesotheliomas at a low incidence (3.2%, all routes, all vehicles), and arise most commonly from the tunica vaginalis of the testes. Mesotheliomas are extremely rare in female control F344/N rats (0.17%, all routes, all vehicles), and typically arise from various locations within the peritoneal cavity, including adjacent to the ovary (Hall et al. 1990). Mesotheliomas have not been reported in vehicle control male or female F344/N rats in NTP inhalation studies (0%). Two VDC-exposed female rats developed mesothelioma; given the rarity of this tumor in females, this incidence isbiologically significant. Related to the incidences of mesothelioma, the NTP determined that there was clear evidence of carcinogenicity in male rats and equivocal evidence of carcinogenicity in female rats exposed to VDC (NTP in press-a). The exact reason for the increased proclivity of males to develop mesotheliomas is unknown, but an association with interstitial (Leydig) cell tumors of the testis, which are common in aged F344 rats (Tanigawa, Onodera, and Maekawa 1987; Turek and Desjardins 1979) has been suggested. Several compounds associated with mesothelioma development in humans and animal models are known mutagens (i.e., ethylene oxide, glycidol; Irwin et al. 1996; Lynch et al. 1984; Maronpot et al. 2009), while others such as potassium bromate and iron cause oxidative stress (Crosby et al. 2000; Hu et al. 2010; Peterson, Greenberg, and Buffler 1984), which can lead to genomic instability. Therefore, it is important to understand that non-asbestos-related chemical exposures can be an important and significant risk factor to the development of mesothelioma in humans.
Several studies have investigated the relationship between the high incidence of interstitial (Leydig) cell tumors of the testes and the development and pathogenesis of mesothelioma of the tunica vaginalis in aging male F344/N rats (Haber et al. 2009; Maronpot et al. 2009; Tanigawa, Onodera, and Maekawa 1987); however, a firm association between these 2 tumor types has yet to be established. One hypothesis proposed by Maronpot et al. (2009) suggests that the size of the interstitial cell tumors may cause physical pressure or mechanical stress on the mesothelial cells of the tunica vaginalis, leading to mesothelial cell stimulation and the release of growth factors. Another hypothesis briefly described subsequently is associated with age-related hormonal imbalance (Maronpot et al. 2009; Turek and Desjardins 1979). Testosterone levels decrease with age between 4 and 18 months in rats. Luteinizing hormone (LH) is increased and stimulates Leydig cell proliferation and increased testosterone production. Although Leydig cells produce testosterone, the hyperplastic Leydig cells and the subsequent Leydig cell tumors still do not produce enough circulating testosterone to correct the imbalance, so circulating LH remains elevated. Leydig cell tumors are associated with higher intratesticular testosterone levels compared to the circulation (Foster 2007). Exposure of the tunica vaginalis to this higher level of testosterone (Gerris and Schoysman 1984; Karpe et al. 1982) may trigger mesothelial cell proliferation via upregulation of growth hormones as suggested by Maronpot et al. based on this hormonal relationship in other tissues.
The objective of this study was to determine the gene expression profile of mesotheliomas in VDC-exposed rats, compared to spontaneously arising mesotheliomas. We recently characterized the global gene expression profile of spontaneous mesotheliomas in F344/N rats (Blackshear et al. 2013), which now provides a critical data set to better understand mesotheliomas induced by chemical exposure. We have now used this data set to investigate changes in gene expression in mesotheliomas that occur in VDC-exposed rats, in order to gain knowledge of mechanisms of mesotheliomagenesis due to VDC exposure, and to further understand the utility of the F344/N rat for the identification of potentially carcinogenic substances in hazard identifications. In addition, we explored the potential relationship between the presence of interstitial cell tumors of the testes and mesotheliomas in order to define a link to explain the gender specific nature of this lesion in the F344/N rat.
Materials and Methods
Sample Collection and Histopathology
Peritoneal mesothelioma samples were collected from VDC-exposed male F344/N rats from the 2-year NTP bioassay (TR582, NTP in press-a). Tumor samples were collected from the testes capsule (5), mesentery (2), and the peritoneum (2), from 9 animals: 6 from high-dose animals (100 ppm VDC), 2 from mid-dose animals (50 ppm VDC), and 1 from a low-dose animal (25 ppm VDC) (Table 1). One-half of each tumor sample was fixed in 10% neutral buffered formalin and the other half was flash frozen in liquid nitrogen. The morphology of each sample was reviewed to assure minimal necrotic tissue (<20%) and maximum tumor to normal tissue per section. Spontaneous mesotheliomas from control male F344/N or F344/NTac rats from other NTP studies (Codeine [TR 455, NTP 1996], Riddelliine [TR508, NTP 2003], Cobalt metal [TR581, NTP in press-b]) were available in the NTP frozen tissue repository for analysis (Table 1). A non-transformed, immortalized mesothelial cell line (Fred-PE) was used as a normal mesothelium control, as described in previous studies (Blackshear et al. 2013; Crosby et al. 2000; Kim et al. 2006).
Mesotheliomas from male F344/N rats in the NTP 2-year bioassay used for genomic profiling.
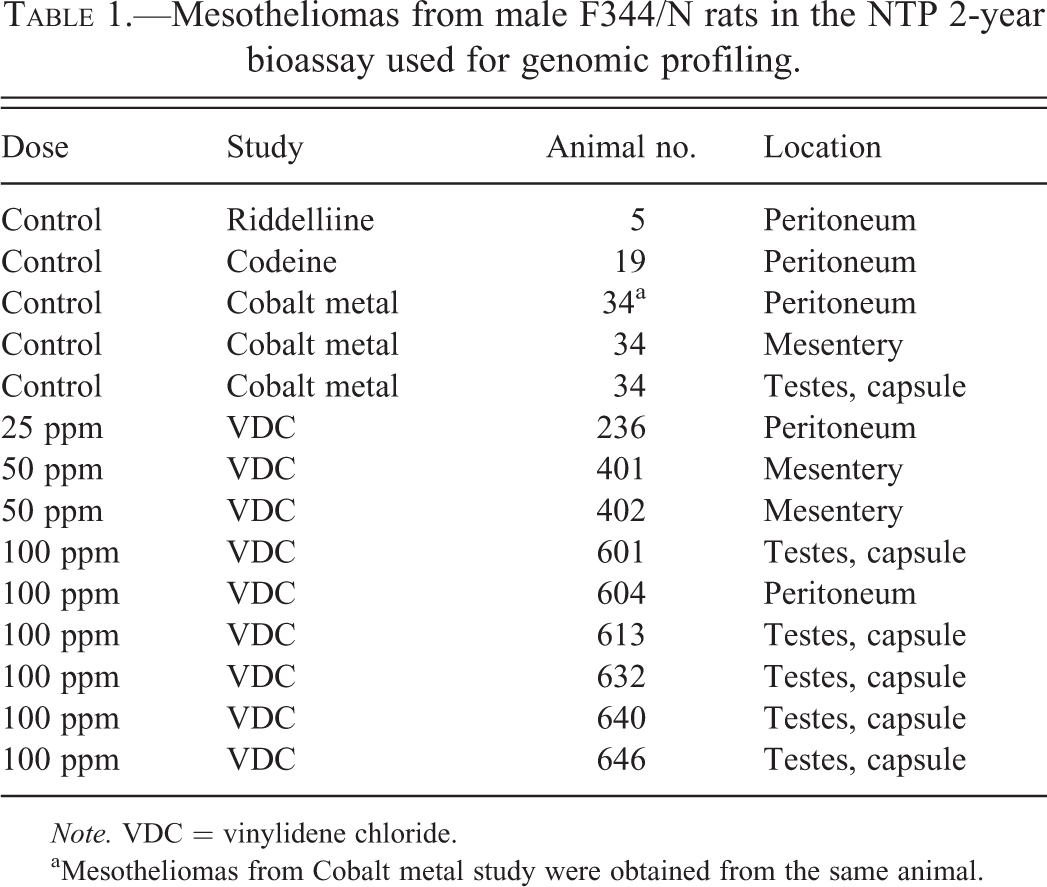
Note. VDC = vinylidene chloride.
aMesotheliomas from Cobalt metal study were obtained from the same animal.
For analysis of the correlation between mesothelioma incidence and interstitial cell tumors of the testes, vehicle control (n = 32) and VDC-exposed (n = 97) animals with testicular interstitial cell adenomas (TICA) were evaluated to examine the correlation between the size of TICA and the incidence of mesothelioma of the tunica vaginalis (MTV). The dimensions of each TICA were measured as the longest and shortest axis of each tumor, as well as the testis, and then the respective areas (π × R × r) were calculated. Based on the area (mm2) of the TICA and respective testis, tumors were categorized as “small” (testes < 130; TICA < 35), “medium” (testes = 130–160; TICA = 35–85), or “large” (testes > 160; TICA > 85;). The size and incidence of TICA were then compared to the incidence of mesotheliomas in vehicle control and VDC-exposed rats to determine if an association exists.
Extraction and Quantification of RNA
Extraction of RNA was performed as previously described (Blackshear et al. 2013) using the Invitrogen PureLink Mini Kit (Invitrogen cat no. 12183-081A). Frozen tissue samples were lysed and homogenized in TRIzol reagent (Invitrogen, Carlsbad, CA) using a rotor-stator homogenizer. Isolation of RNA was performed according to mini kit protocol. On-column DNase treatment was performed using the Invitrogen PureLink DNase kit (Invitrogen, Carlsbad, CA) to purify RNA samples. RNA quantification and RNA integrity number were measured on a bioanalyzer (Agilent Technologies, Santa Clara, CA). Samples were aliquoted and stored at −80°C until analyzed.
RNA Labeling, Microarray Hybridization, and Data Processing
Gene expression analysis was conducted as previously described (Blackshear et al. 2013) using Affymetrix Rat Genome 230 2.0 GeneChip® arrays (Affymetrix, Santa Clara, CA). Total RNA (100 ng) was amplified as directed in the Affymetrix 3′ IVT Express kit protocol. Amplified biotin-aRNAs (12.5 µg) were fragmented and 10 µg were hybridized to each array for 16 hr at 45°C in a rotating hybridization oven using the Affymetrix Eukaryotic Target Hybridization Controls and protocol. Array slides were stained with streptavidin/phycoerythrin utilizing a double-antibody staining procedure and then washed for antibody amplification according to the GeneChip Hybridization, Wash and Stain Kit user manual. Arrays were scanned in an Affymetrix Scanner 3000.
Probe intensity data from all arrays were entered into the R software environment (http://www.R-project.org) directly from .cel files using the R/affy package (Gautier, Bolstad, and Irizarry 2003). Data quality was evaluated using image reconstruction, intensity histograms, and boxplots. Image reconstruction of one vinylidene chloride sample was found to have a large smudge, and this sample was subsequently removed from the analysis. Normalization was performed on the remaining 19 samples (6 Fred-PE cell lines, 5 spontaneous mesotheliomas, 8 VDC exposed mesotheliomas) using the robust multiarray average (RMA) method to form one expression measure for each gene on each array (Irizarry et al. 2003). The RMA method adjusts the background of perfect match (PM) probes, applies a quantile normalization of the corrected PM values, and calculates final expression measures using the Tukey’s median polish algorithm.
Pairwise comparisons between treatment groups (normal, vinylidine chloride, and spontaneous tumor) were made for each probe set between Fred-PE cells and each tumor group (spontaneous mesothelioma, mesotheliomas from VDC-exposed animals) using a bootstrap t test while controlling the mixed directional false discovery rate (mdFDR; Guo, Sarkar, and Peddada 2010). The mdFDR procedure controls the overall false discovery rate across multiple comparisons and takes into account directional errors corresponding to upregulated or downregulated genes. The approach does not assume normality and is robust across different variance structures. Statistical calculations were performed in the ORIOGEN software package using 10,000 bootstrap samples (Peddada et al. 2005). The mdFDR significance level for differential gene expression was set to 5%. The resulting differentially expressed genes identified were significant at p < .001.
Ingenuity Pathways Analysis (IPA) Core Analysis was based on the lists of differentially expressed genes between spontaneous mesotheliomas and normal Fred-PE mesothelial cells as well as VDC-exposed mesotheliomas and normal Fred-PE mesothelial cells and used to determine biologic functions, canonical pathways, and transcription factor activation. Overrepresented categories of genes were determined from statistical outcomes by testing for association with gene product relationships from a curated database of biological networks (Ingenuity Pathways AnalysisTM [IPA] version 9.0; http://www.ingenuity.com/). The Ingenuity Pathways Knowledge Base (IPKB) consists of data with known biological relationships between genes and gene products. Enrichment of pathway members among differentially expressed probe sets were assessed with the one-tailed Fisher’s exact test for 2 × 2 contingency tables. Significantly altered pathways in the IPA Core Analysis were then grouped by pathways to account for upstream and downstream effectors as well as overlapping pathways. Upstream activation was based on an IPA Z-score of >2.0 with no bias. Gene expression data can be found in the GEO database at: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE47581.
Immunohistochemistry
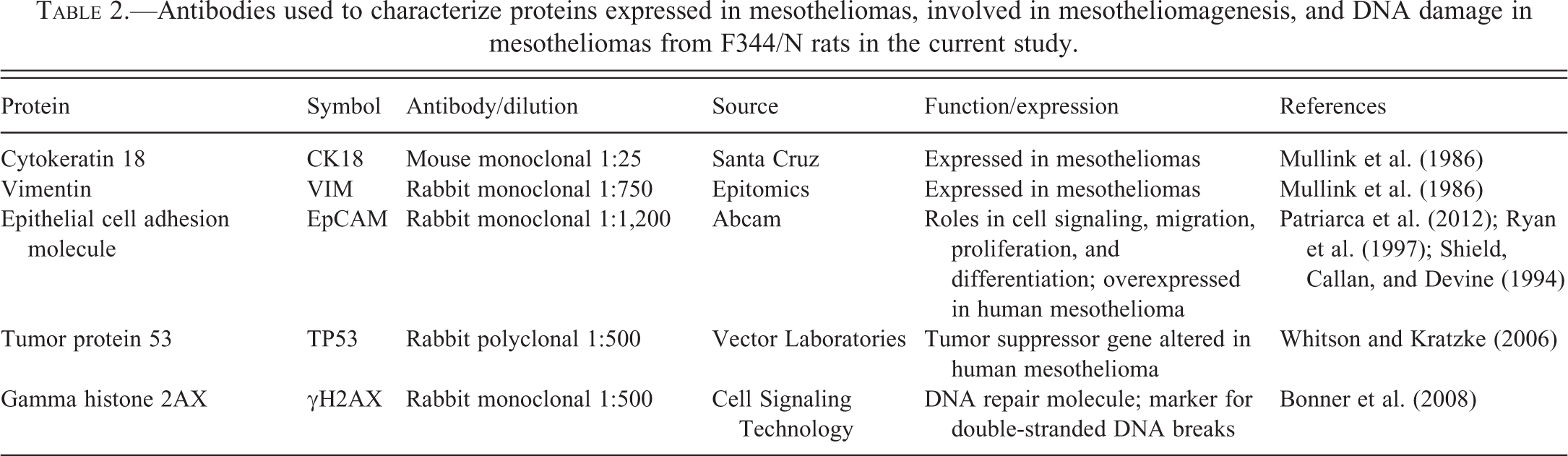
Immunohistochemistry was performed on unstained, formalin-fixed, paraffin-embedded sections of spontaneous mesotheliomas and mesotheliomas from VDC-exposed rats for a variety of proteins that play a role in mesotheliomagenesis or DNA damage (Table 2). Sections were deparaffinized, rehydrated, and counterstained with hematoxylin, then dehydrated, cleared, and coverslipped in a similar manner for all antibodies. Endogenous peroxidases were blocked with 3% hydrogen peroxide. Antigen retrieval was performed with heat and pressure, using citrate buffer (Biocare Medical, Concord, CA). Sections for H2AX staining were incubated with Dako block (Dako, Carpinteria, CA), whereas for the 4 other stains (Epithelial Cell Adhesion Molecule [EpCAM], Tumor Protein 53 [TP53], cytokeratin 18 [CK18], and Vimentin [VIM]), appropriate serum blocks were used, followed by the Avidin-Biotin Blocking Kit (Vector Laboratories, Burlingame, CA). The sections were incubated with EpCAM rabbit monoclonal antibody (1:1,200; Abcam, Cambridge MA), rabbit polyclonal TP53 antibody (1:500; Vector Laboratories, Burlingame CA), rabbit monoclonal H2A.X antibody (1:500; Cell Signaling Technology, Danvers, MA), mouse monoclonal CK18 antibody (1:25; Santa Cruz Biotechnology, Santa Cruz, CA), and rabbit monoclonal VIM antibody (1:750; Epitomics, Burlingame, CA). For negative controls, antibody was substituted with normal serum from the species the antibody was raised in (Jackson Immunoresearch Laboratories, West Grove, PA). The slides were then incubated with respective secondary antibodies and label incubation was performed using Vector R.T.U. Vectastain Kit (Vector Laboratories, Burlingame, CA) for 30 min at room temperature. Immune reactions were visualized using 3,3-diaminobenzidine (DAB) chromagen (Dako, Carpentaria, CA).
Antibodies used to characterize proteins expressed in mesotheliomas, involved in mesotheliomagenesis, and DNA damage in mesotheliomas from F344/N rats in the current study.
Results
Mesotheliomas in VDC-exposed F344/N Rats Are Morphologically Similar to Spontaneous Rat Mesotheliomas
Mesotheliomas in all VDC-exposed animals were morphologically similar to spontaneous mesotheliomas in F344/N rats, and were classified as epitheliomatous, characterized by papillary projections of one or more layers of atypical mesothelial cells overlying a fibrovascular stalk (Figure 1A). Some tumors had a more solid phenotype, forming sheets of cells interspersed with variable amounts of fibrovascular stroma, which frequently contained mixed to mononuclear inflammatory infiltrates. The mixed inflammatory component was variable from sample to sample but included eosinophils, mast cells, or neutrophils, and prominent macrophages laden with hemosiderin (Figure 1B). Although the majority of mesotheliomas in VDC-exposed rats arose from the tunica vaginalis of the testes, frequently other tissues such as epididymis, seminal vesicles, prostate, skeletal muscle, mesentery, peritoneum, spleen, stomach, pancreas, and liver were also involved. Eight of the 9 animals used for obtaining mesothelioma samples for microarray also had concurrent skeletal muscle involvement, but this was not associated with the microarray sample.
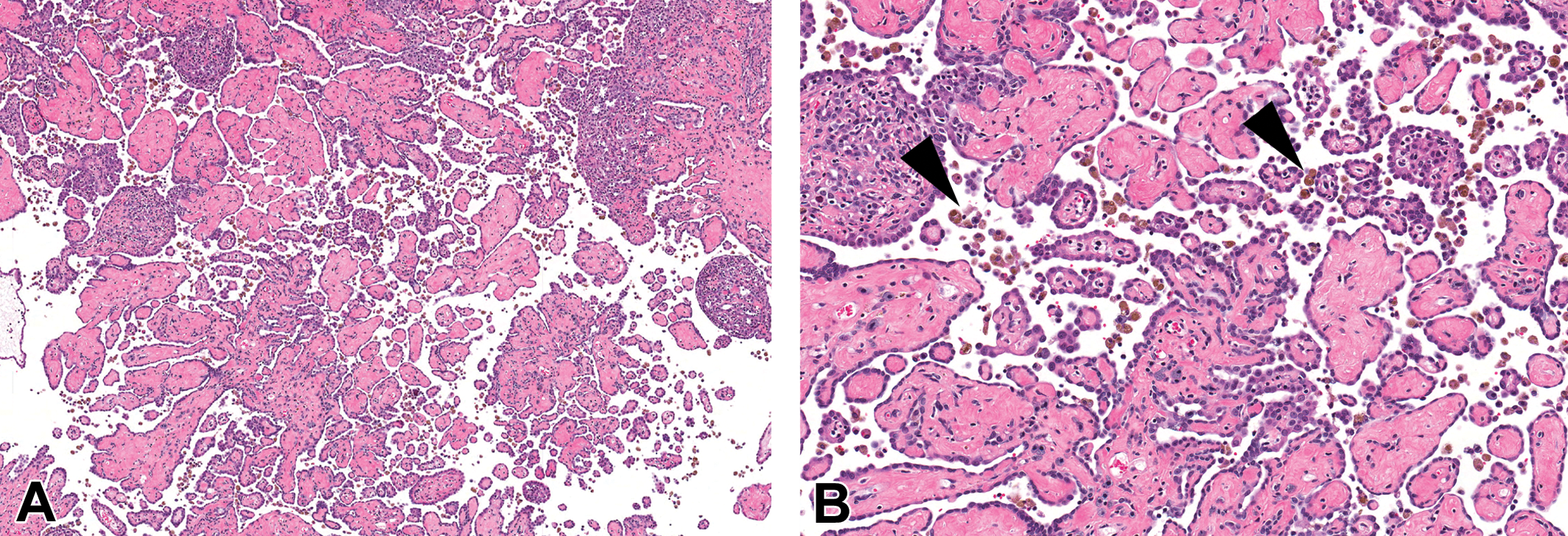
Mesothelioma arising from the abdominal wall in a vinylidene chloride-exposed male F344/N rat. (A) Mesotheliomas were characterized by an epitheliomatous phenotype composed of papillary projections of mesothelial cells supported by fibrovascular stalks. Hematoxylin and eosin (H&E). (B) Mesotheliomas were associated with a variable mononuclear cell infiltrate containing macrophages laden with hemosiderin (arrowheads). H&E.
Gene Expression of Mesotheliomas from VDC-exposed Rats Is Similar to Spontaneous Tumors, but Differentiated by Increased Cell Damage Response and Pro-inflammatory Response
Principal components analysis (PCA) of differentially expressed genes showed clear separation between experimental groups, based on significantly differentially expressed genes (Figure 2). Unsupervised hierarchical cluster analysis showed similarities in expression between spontaneous mesotheliomas and those from VDC-exposed animals, with small regional differences in global gene expression and clear differences in gene expression between tumors and Fred-PE mesothelial cell line (Figure 3). Of the 31,099 probe sets on the array, 14,136 were differentially expressed in VDC-induced mesotheliomas compared to Fred-PE. The top upregulated and downregulated, differentially expressed genes were largely associated with skeletal muscle biology and cellular damage; some overlap with genes found in spontaneous mesothelioma was observed. The genes associated with skeletal muscle biology and contractility in VDC-associated mesotheliomas were considered to be related to the presence of skeletal muscle within sections of mesothelioma obtained from the abdominal wall in VDC-exposed animals.
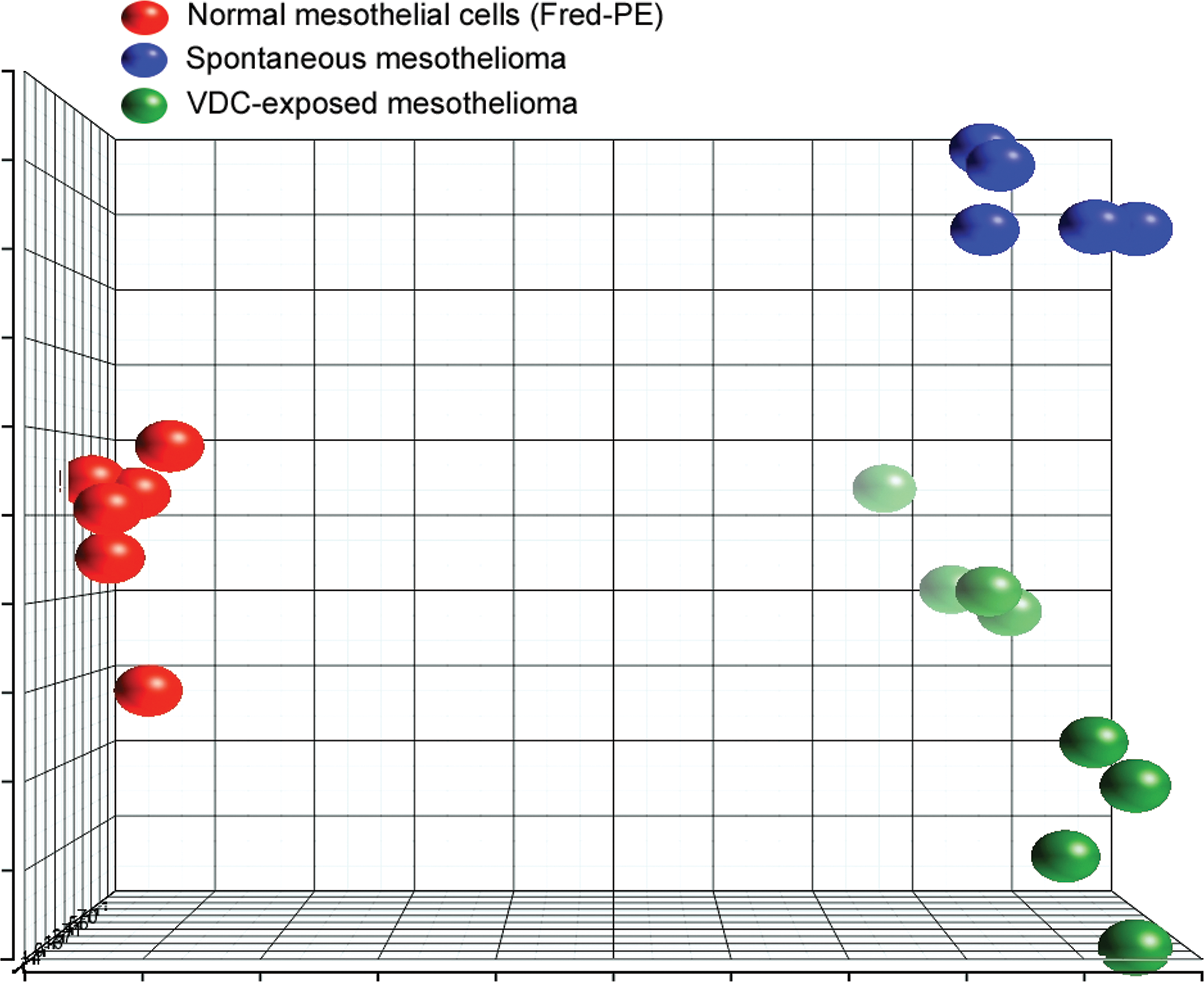
Principal component analysis (PCA) comparing gene expression profiles of Fred-PE mesothelial cells, spontaneous mesotheliomas from control rats, and mesotheliomas from vinylidene chloride–exposed rats for differentially expressed probe sets. PCA shows intergroup similarities in gene expression, and clear separation of experimental groups in space, indicating differences between groups in terms of their gene expression profiles.
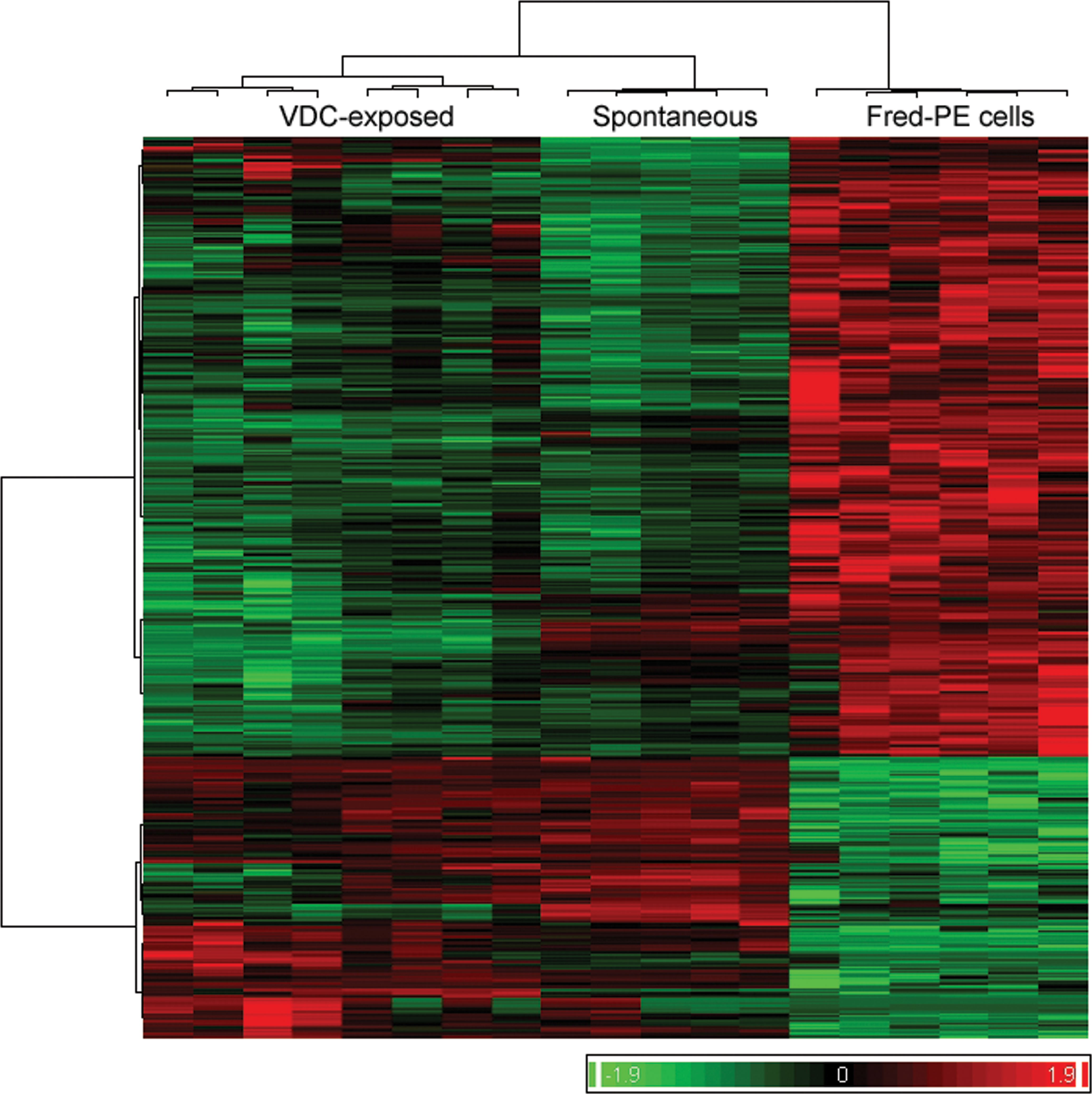
Hierarchical cluster analysis (HCA) comparing global gene expression profiles of Fred-PE mesothelial cells, spontaneous mesotheliomas, and vinylidene chloride–associated mesotheliomas. HCA clusters experimental samples based on global gene expression and shows relative expression levels across the genome for each of the 3 experimental groups.
To understand the similarities and the differences between VDC associated and spontaneous mesotheliomas, a comparison analyses of the differentially expressed genes generated from the analysis of VDC-exposed mesotheliomas versus Fred-PE and spontaneous mesotheliomas versus Fred-PE was conducted. There were 10,372 shared probe sets between spontaneous and VDC-induced mesotheliomas, 3,764 probe sets unique to mesotheliomas from VDC-exposed animals, and 3,612 unique to spontaneous mesotheliomas. The unique VDC probe sets were largely associated with skeletal muscle biology and not relevant to mesothelioma development. Of the probe sets shared in common, 9,568 were mapped in the IPA database, 7,454 of which were considered analysis ready according to IPA based on threshold of and filtering for duplicates. IPA core analysis was used to correlate the 7,454 probe sets with relevant biological functions. Biological functions representative of the top upregulated and downregulated genes in both tumor groups compared to Fred-PE cells (p < .05) included pathways associated with tumorigenesis, including cellular development, cell and tissue morphology, embryonic development, organ development, cellular growth and proliferation, cell cycle, and cell morphology, with many of the probe sets showing overlapping biological functions in multiple categories (Figure 4). There was significant differential expression of several oncogenes, growth factors, cell cycle regulators, embryonic genes, cell survival genes, and solute carrier molecules in both spontaneous and VDC-exposed mesotheliomas compared to Fred-PE cells (Table 3). There was significant overlap of genes associated with cell growth and tissue remodeling (Tgfβ2, Tgfβr1, Vegfc, Fgfr2 Igf1, Igfbp), cell cycle regulators (Cdkn1a, Cdkn1b), oncogenes and proto-oncogenes (Mafb, Fos, Junb, Lyn), Ras-Mapk pathway mediators (Rasd1, Rnd, Prkcb, Mapk12), tumor suppressor genes (Tp53, Lats2), adhesion molecules (Epcam, Cdh22, Ctnnb1, Itgb2), apoptosis genes (Gadd45b, Bcl2a1), developmental genes (Plac8, Wnt4, Plau, Gata5), transporters and solute carriers (Slc7, Slc28, Abc), mesothelial cell markers (Krt18/19, Des), and genes associated with oxidative stress (Duox, Gpx2), compared to Fred-PE cells. In many cases, the expression of these genes was fairly similar between spontaneous and VDC-exposed mesotheliomas, with both tumor types showing overlap of genes associated with multiple categories. However, there were noticeable differences in the expression of some genes in differentially represented pathways between each tumor group.
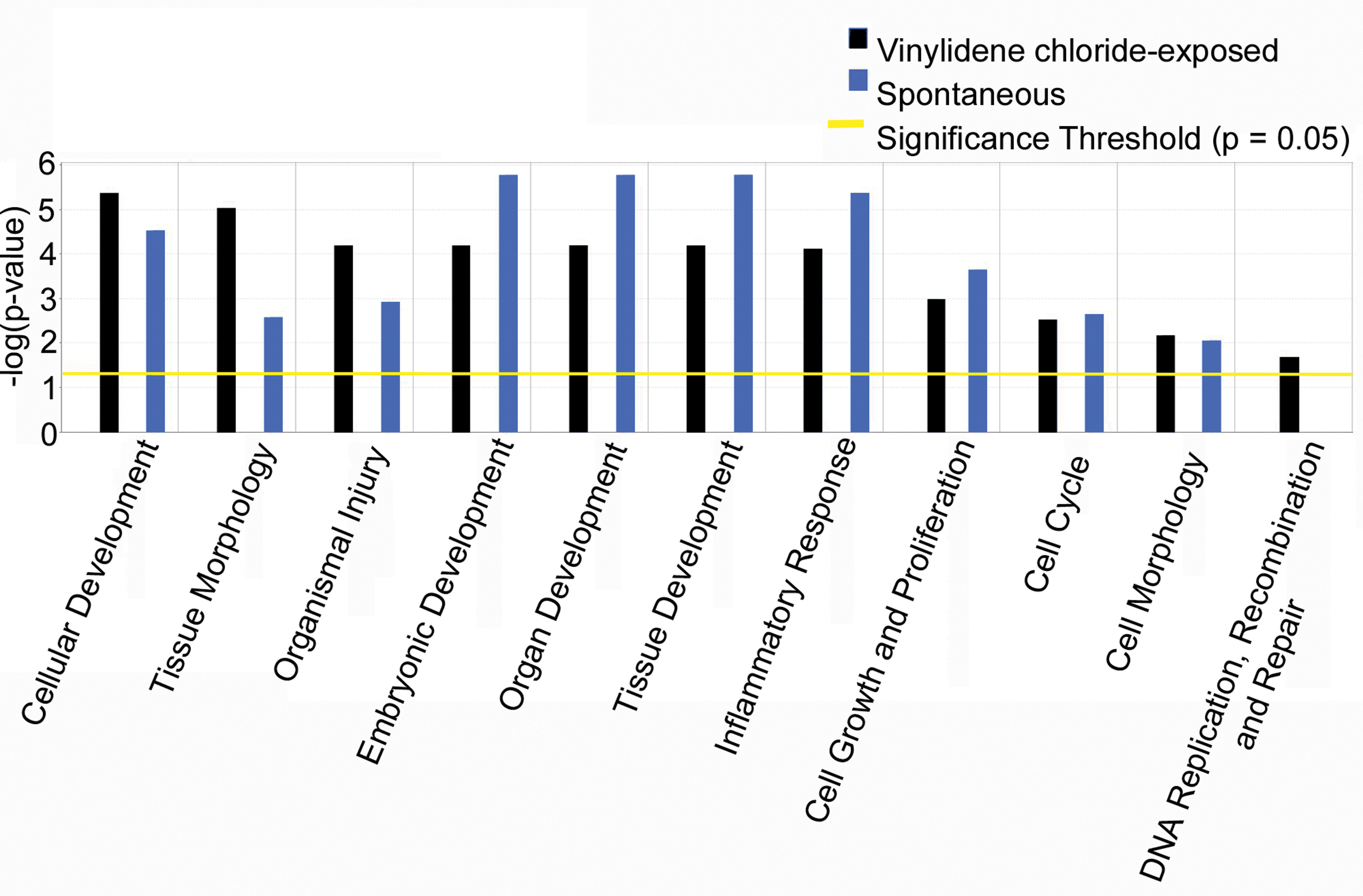
Top overrepresented biologic functions related to tumorigenesis in spontaneous mesotheliomas and mesotheliomas from vinylidene chloride–exposed male F344/N rats (p < .05).
Fold changes of selected genes from significantly altered pathways associated with tumorigenesis in mesotheliomas from VDC-exposed and vehicle control F344/N rats compared to Fred-PE cells.
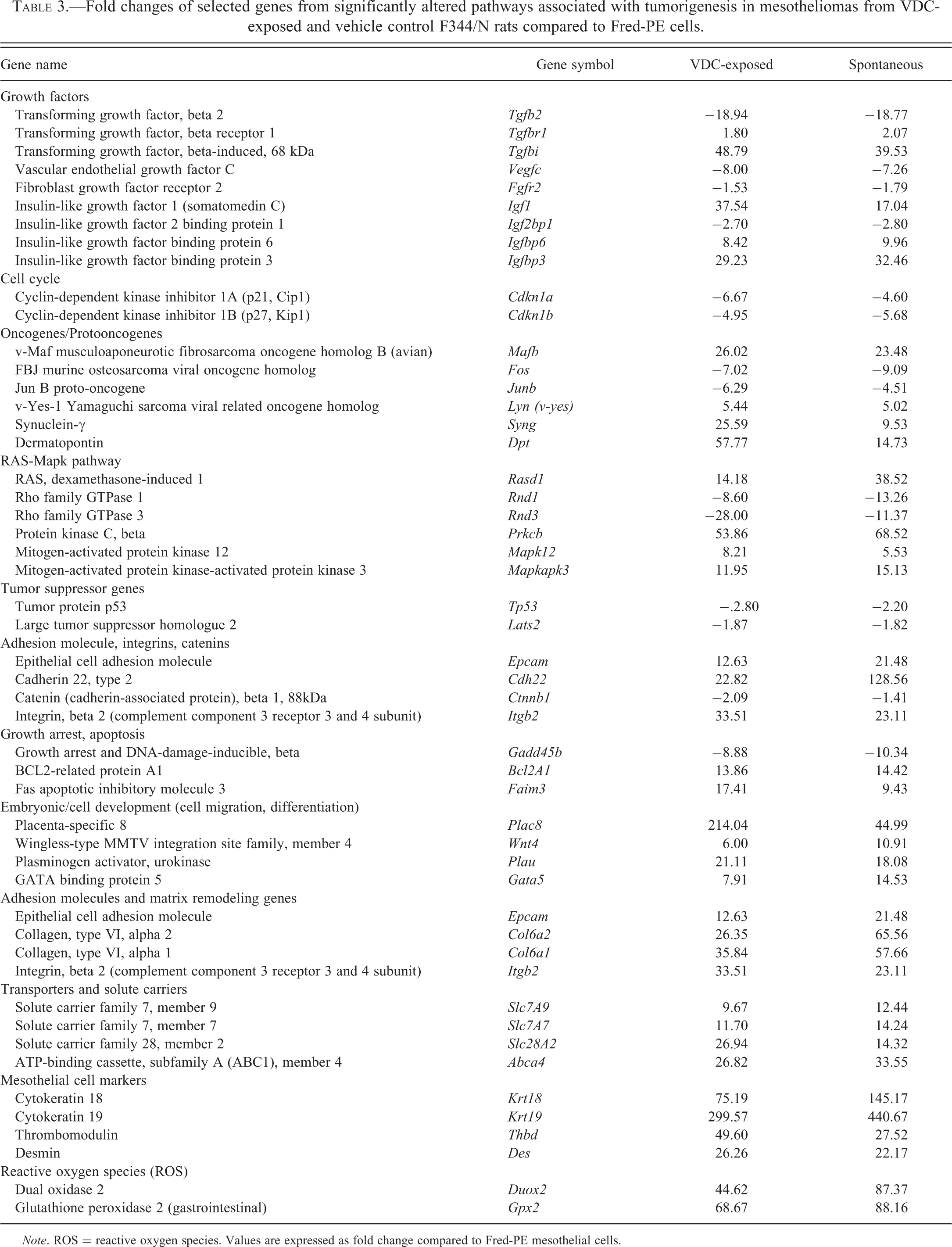
Note. ROS = reactive oxygen species. Values are expressed as fold change compared to Fred-PE mesothelial cells.
Mesotheliomas from control animals and VDC-exposed animals shared genes associated with the overrepresented biologic category inflammatory response as involved in tumorigenesis (Figure 4). Genes associated with inflammatory response included chemokines (Ccl5/6/11/27, Cxcl9/11), cytokines and cytokine receptors (Il1b, Il10/18/24/34, Il6r, Il7r, Il1rn, Tnfrsf11b, Cd40), Complement factors (Cfh, C1qa/b, Serping1), pattern recognition receptors (Tlr2/7/8, Mrc1), interferon pathway mediators (Ifngr1, Irf9, Ifr5, Ifitm1), activated macrophage products (Chi3l1, Sparcl1, C1qb, C1qa, S100a9), cell surface receptors (S1pr1, Fcer, Fcgr, Stab1, Cd163, Cd68, Cd53, Cd36, Clec), and a variety of inflammatory mediators (Aif1, Ptgds1/2, Lyz2, Mcpt10, Tdo2, Ubd, Ddx60, Cybb, Pla2g2a, Lyve1) compared to Fred-PE cells (Table 4). There was differential expression of several of these shared genes between tumor groups, including upregulation of pro-inflammatory chemokines (Ccl5, Ccl6, Cxcl9) and decreased expression of anti-inflammatory cytokines (Il10, Il18, Il24). There was higher upregulation of genes associated with tissue damage (Tlr2, Dpt, Mrc1, Pla2g2a) and damage-associated molecular pattern (DAMP) molecules (S100a8, S100a9, Mrc1, and Lyve1), in VDC-exposed mesotheliomas.
Fold changes of selected genes from significantly altered pathways associated with inflammation and immune response in mesotheliomas from vinylidene chloride-exposed and vehicle control F344/N rats compared to Fred-PE cells.
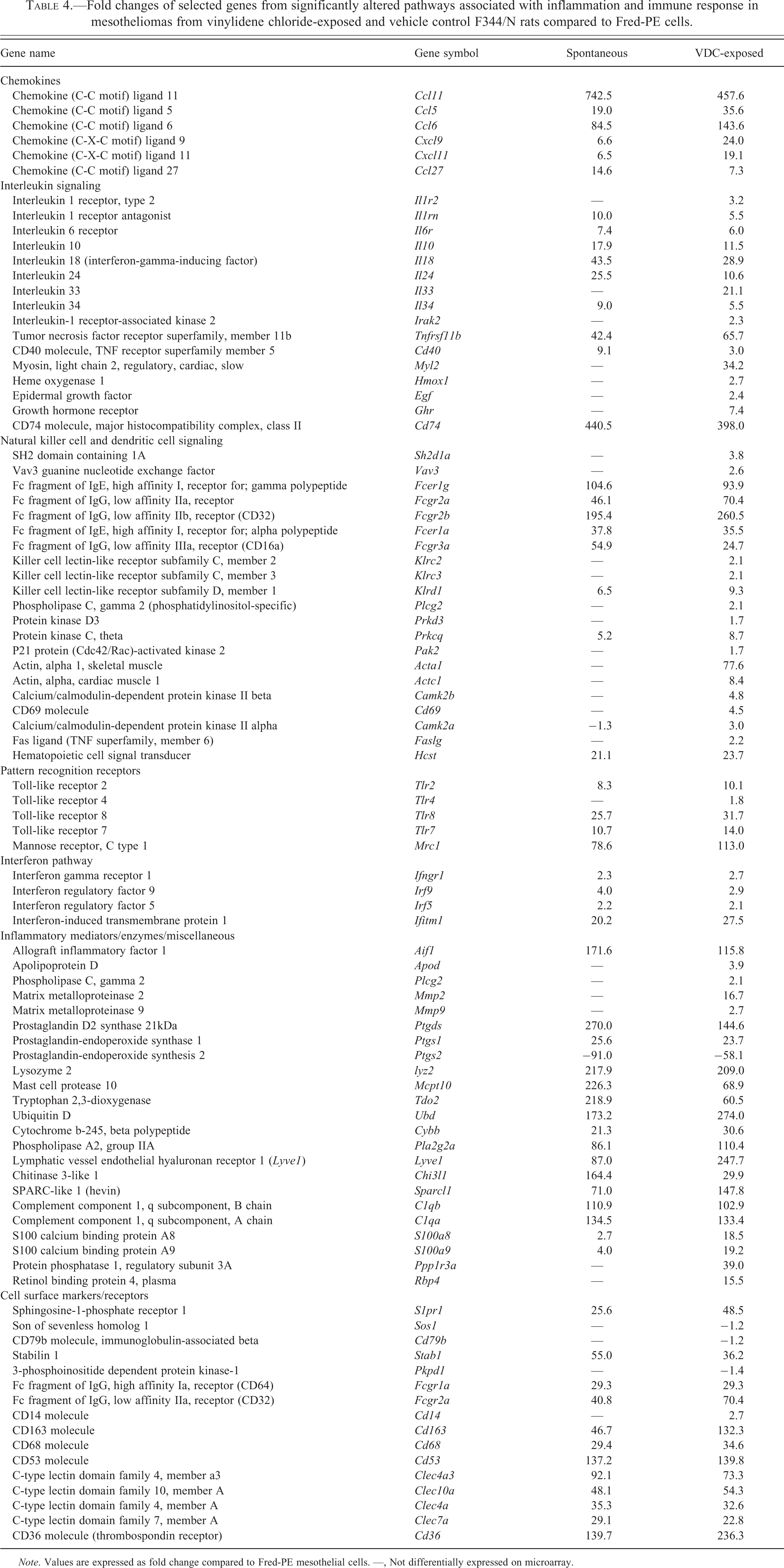
Note. Values are expressed as fold change compared to Fred-PE mesothelial cells. —, Not differentially expressed on microarray.
In contrast to the previously shared overrepresented pathways, there were several canonical pathways associated with inflammation and immune response that were significantly overrepresented in mesotheliomas from VDC-exposed animals compared to Fred-PE cell line control, which were not significantly overrepresented in spontaneous tumors. These included pro-inflammatory pathways such as the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling pathway (Il33, Ghr, Il1r2, Egf, Plcg2, Tlr4), interleukin (IL)-8 (Mmp2, Mmp9, Myl2, Gnai3, Hmox1, Irak2, Prkd3) and IL-12 (Rbp4, S100a8, Pik3r3, Prkce, Apod, Prkd3) interleukin responses, Fc receptor signaling (Vav3, Plcg2, Sos1, Fcgr2b, Mapk9, Cd79b, Pdpk1), PI3K/AKT signaling (Plac8, Ccl5, Gnai3, Nfkb2), and NK and DC signaling (Acta1, Actc1, Camk2b, Cd69, Faslg, Tlr4, Klrc2, Pak2, Pak4), among others (Figure 5 and Table 4). While spontaneous mesotheliomas and mesotheliomas from VDC-exposed animals shared similar expression of a number of genes in these categories compared to Fred-PE cells, based on canonical pathway involvement, there was significant overrepresentation of pathways associated with a pro-inflammatory phenotype and immune dysfunction in mesotheliomas from VDC-exposed animals. Selected genes from these pathways were validated by Quantitative real-time PCR (qPCR; Figure 6). Genes were selected based on known involvement with cancer progression (Cdnk1a, Sncg), embryonic development (Plac8), cell survival/decreased apoptosis (Tp53), repair (Dpt), or for their roles as tumor biomarkers (Epcam, Krt18). In all cases, RNA expression was consistent with the directional change observed on the microarray.
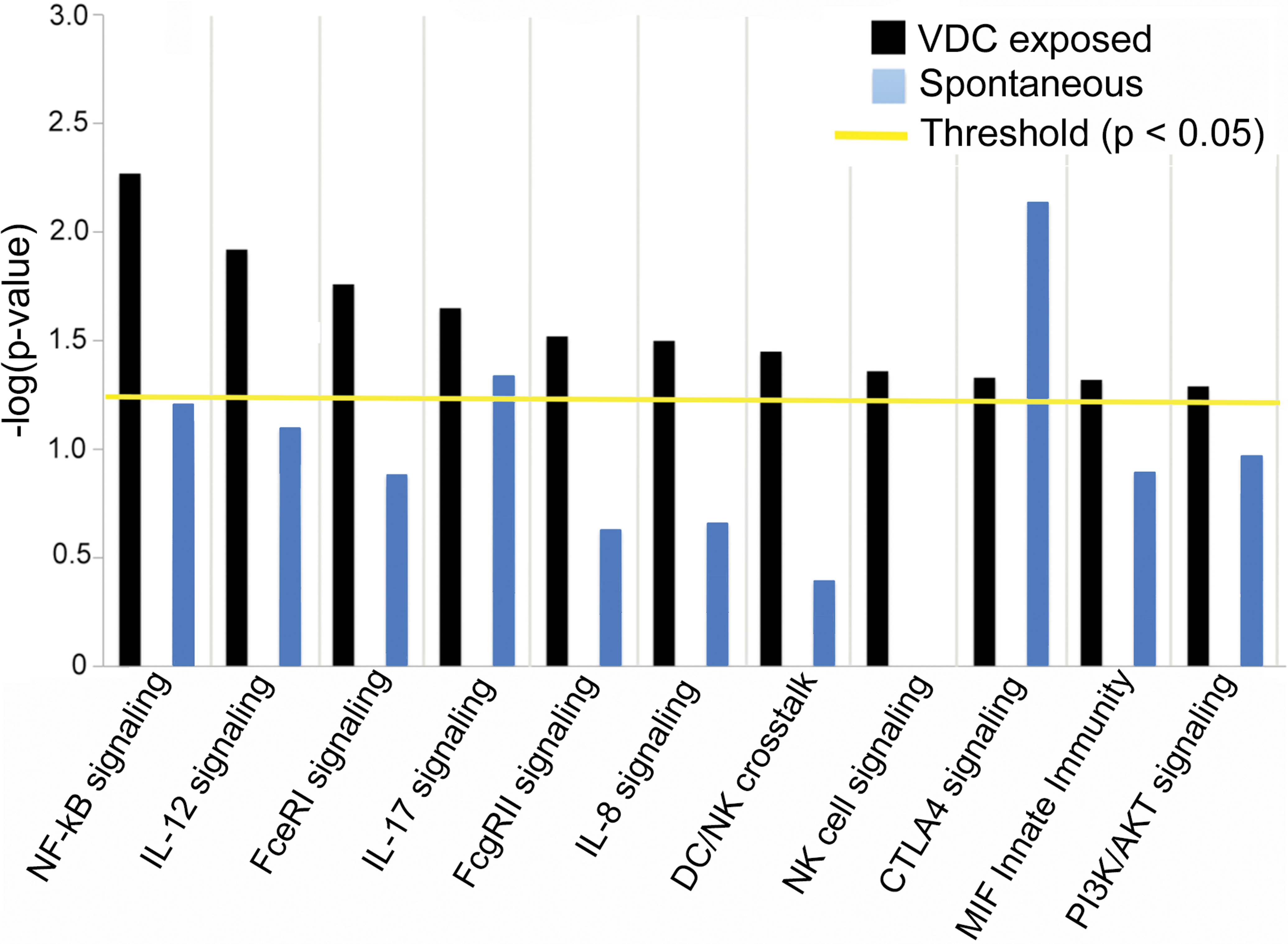
Significantly overrepresented canonical pathways related to inflammation and immune dysfunction overrepresented in mesotheliomas from vinylidene chloride–exposed male F344/N rats (p < .05).
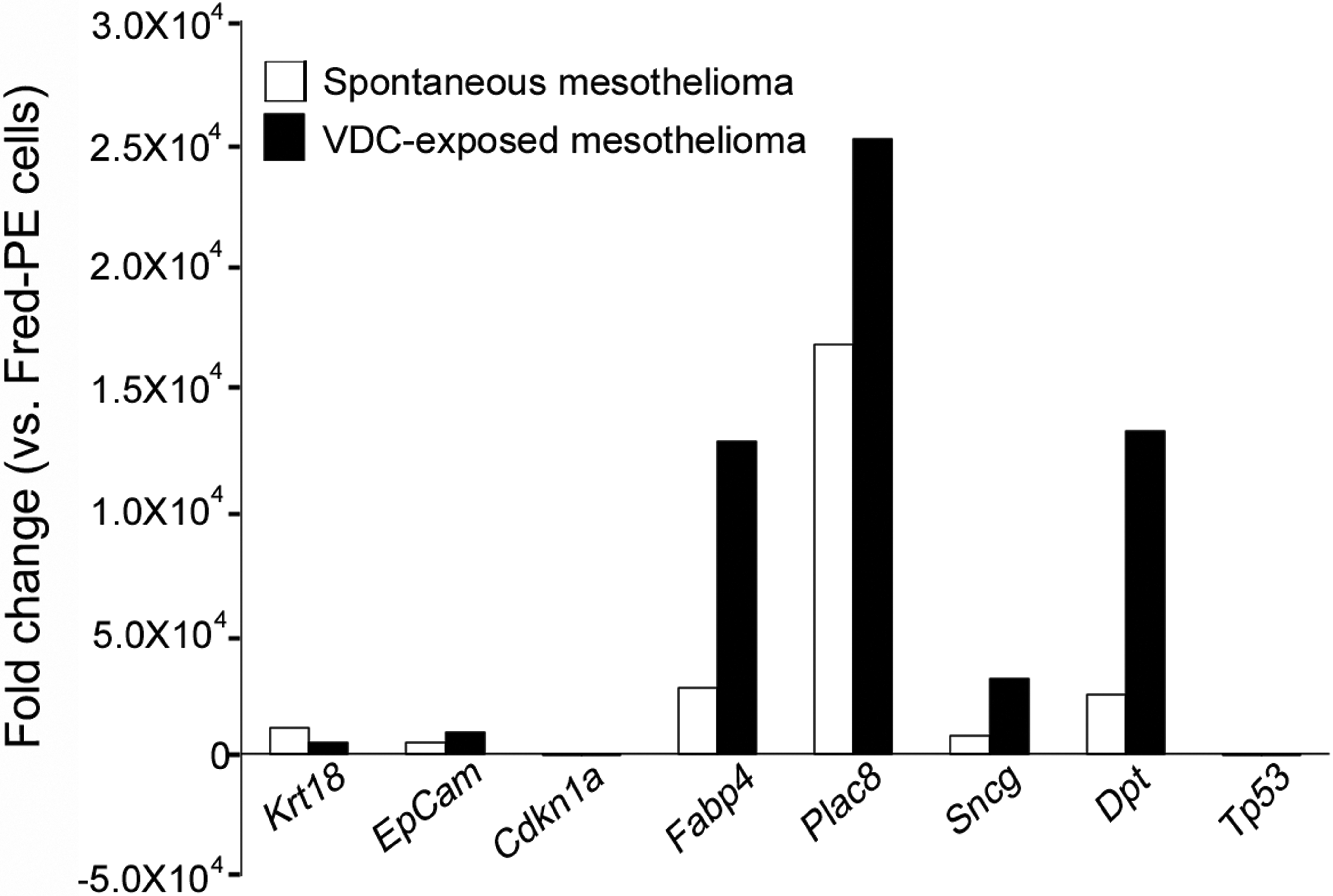
Quantitative real-time PCR validation of select genes differentially expressed on microarray, based on involvement with cancer progression (Cdnk1a, Sncg), embryonic development (Plac8), cell survival/decreased apoptosis (Tp53), repair (Dpt), or as tumor biomarkers (Epcam, Krt18).
Mesotheliomas from Vehicle Control and VDC-exposed F344/N Rats Are Biphasic and Show Similar Alterations in the Expression of Proteins Assayed for Tumor Progression and Slight Differences for DNA Damage
Expression of epithelial (cytokeratin, CK18) and mesenchymal (vimentin) proteins and expression of proteins involved in mesotheliomagenesis (EpCAM) and DNA damage (TP53, γH2AX) was assessed in spontaneous mesotheliomas and mesotheliomas in VDC-exposed F344/N rats by immunohistochemistry. In control samples (rat gastrointestinal tract mucosa), EpCAM was immunolocalized to the cell membrane (Figure 7A). In mesothelioma samples, EpCAM accumulated in the cytoplasm and in the nucleus of cells (Figure 7B). Gamma histone 2AX (γH2AX), a key molecule in DNA repair that is associated with DNA double-strand breaks (Bonner et al. 2008), was localized to scattered mesothelioma tumor cells and degenerate inflammatory cells and with expression levels slightly increased in mesotheliomas of VDC-exposed rats compared to spontaneous mesotheliomas (Figure 7B). There was no immunoreactivity of TP53 protein in spontaneous mesotheliomas or mesotheliomas in VDC-exposed animals (data not shown). Mesotheliomas from VDC-exposed animals as well as vehicle control animals had dual expression of VIM (Figure 7C) and CK18 (Figure 7D), demonstrating their biphasic nature (Mullink et al. 1986).
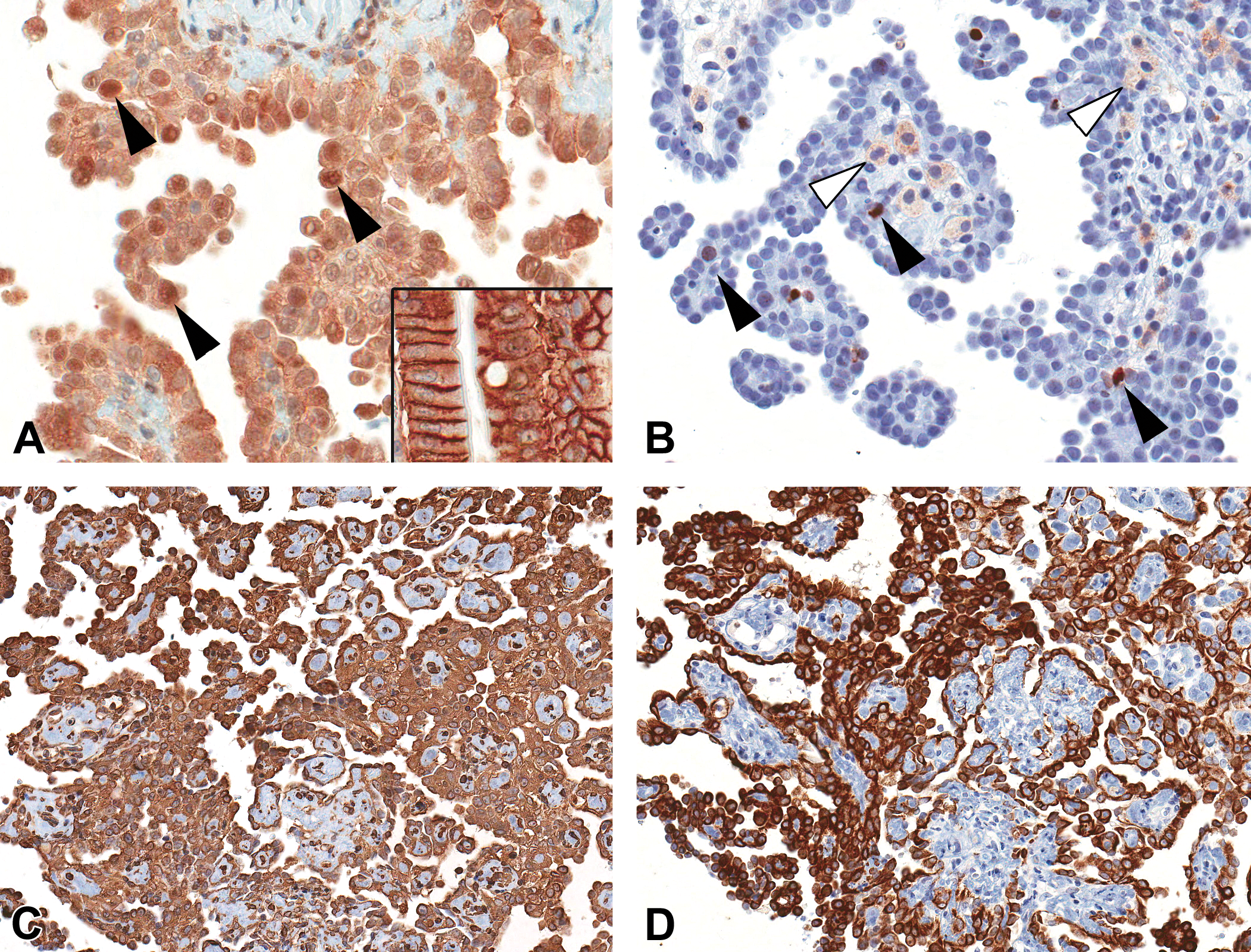
Immunohistochemical analysis of mesotheliomas from vinylidene chloride (VDC)-exposed animals. (A) Translocation of epithelial cell adhesion molecule from the cell membrane (control, rat gastrointestinal tract, inset) to the cytoplasm and nucleus (black arrowheads) in spontaneous and VDC-associated mesotheliomas. (B) Random expression of H2A.X protein in the nucleus of scattered mesothelioma cells (black arrowheads) and inflammatory cells (white arrowheads). (C) Biphasic nature of mesotheliomas illustrated by diffuse cytoplasmic immunoreactivity to cytokeratin and (D) vimentin antibodies.
Mesothelioma Incidence Is not Correlated with Interstitial Cell Tumors of the Testis
To address the hypothesis that the incidence of mesothelioma in rats is influenced by the presence and size of testicular interstitial cell tumors, analyses of testicular size, incidence and size of interstitial cell adenomas, and mesotheliomas were performed. No correlation was observed between testicular size and mesothelioma or interstitial cell adenoma incidence and mesothelioma (Table 5).
Correlation of interstitial cell adenoma size with mesothelioma incidence.
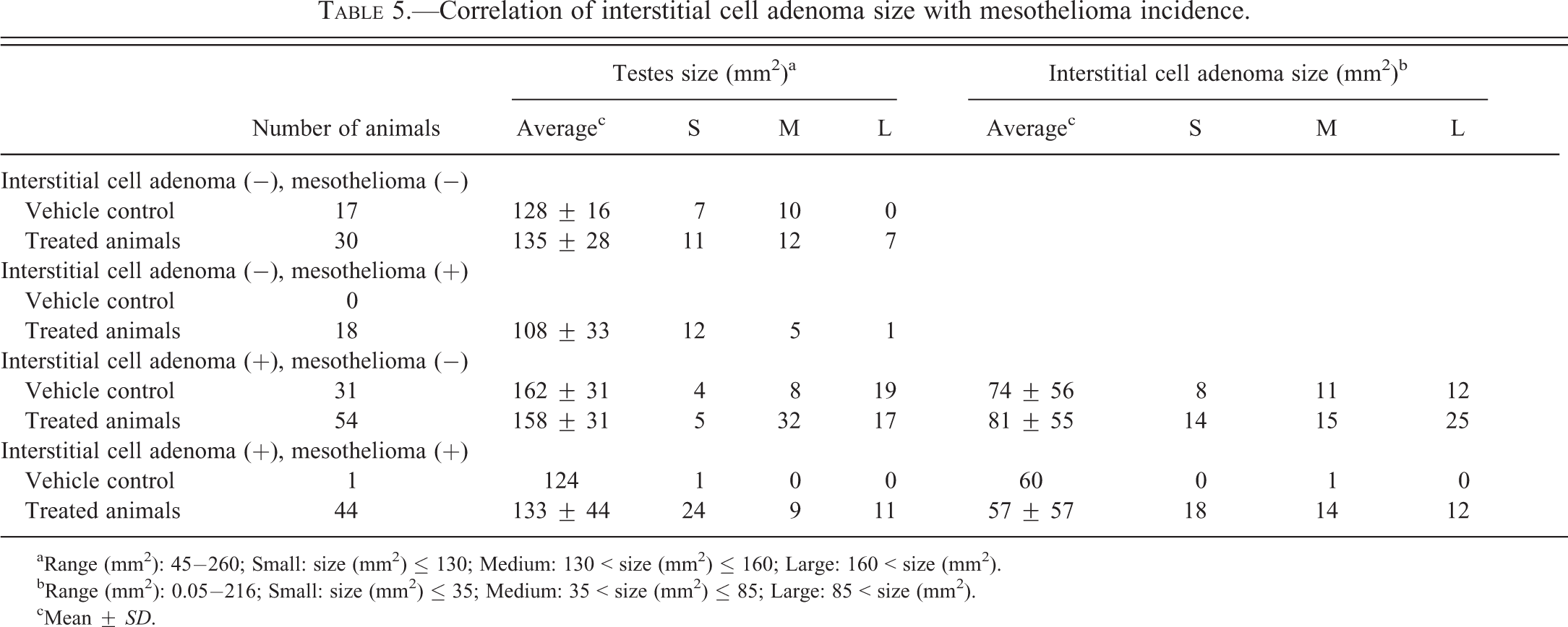
aRange (mm2): 45−260; Small: size (mm2) ≤ 130; Medium: 130 < size (mm2) ≤ 160; Large: 160 < size (mm2).
bRange (mm2): 0.05−216; Small: size (mm2) ≤ 35; Medium: 35 < size (mm2) ≤ 85; Large: 85 < size (mm2).
cMean ± SD.
Discussion
Comparison of global gene expression profiling of mesotheliomas arising in male F344/N rats exposed to VDC, spontaneous mesotheliomas in F344/N rats, and cultured rat mesothelial cells (Fred-PE cells) was performed in order to characterize the molecular features of these tumors and elucidate their chemical-specific gene expression profiles. Global gene expression profiling enabled the clear differentiation of mesotheliomas arising in VDC-exposed rats from background spontaneous mesotheliomas despite indistinguishable morphology and provided important information on the further understanding of the molecular features of these chemically induced tumors. Furthermore, based on differentially expressed genes, PCA suggested that variations in site, vehicle, and dose did not seem to bias the gene expression analysis in this study. Spontaneous mesotheliomas were obtained from studies utilizing a number of different routes of exposure and vehicle (gavage, feed, and inhalation), and 3 of the 5 spontaneous tumors were collected from different sites within the same animal (Cobalt metal). Regardless of these variables, spontaneous tumors showed tight clustering and overlap on the PCA based on differentially expressed genes. In addition, the 3 spontaneous tumor samples obtained from the single animal overlapped and admixed with the tumor samples from the other studies (Riddelliine and Codeine), indicating that the tumor samples from the single animal did not adversely bias the differential gene expression results. Likewise, mesotheliomas from VDC-exposed animals did not cluster based on dose or site of collection. Finally, in terms of the Fred-PE cell line control, it is well known that variations in RNA collection, time in culture, cell passage, and other in vitro factors when using these cells may contribute to variations in gene expression. However, the Fred-PE samples clustered very tightly together, suggesting these in vitro variables did not bias the differential gene expression in these samples.
Previous studies have characterized gene expression profiles or molecular features of chemically induced mesotheliomas in F344/N rats or F344 crosses (Hu et al. 2010; Kim et al. 2006); however, these studies did not consider the background gene expression of mesotheliomas when interpreting chemical effects. By comparing gene expression of mesotheliomas in VDC-exposed animals to spontaneous mesotheliomas, the nonspecific oncogenic background cancer effects are normalized, and chemical effects in the context of oncogenesis can be more easily interpreted. Gene expression profiling and mutation analysis of human mesotheliomas has revealed a variety of genetic alterations including mutations in tumor suppressor genes (TP53, P16, BAP1, WT1, NF2, PTEN) and cell cycle genes (CDKN2A/2B and CDKN2C), and alterations in proteins associated with tumorigenesis (COL3, CCL2, LGALS; Bueno et al. 2010; Gordon et al. 2005; Gueugnon et al. 2011; Tsujimura et al. 2012). In large part, similarities between spontaneous mesotheliomas and those from VDC-exposed animals primarily involved genes associated with tumorigenesis. Commonly affected biologic functions involved a number of similarly expressed oncogenes, tumor suppressor genes, growth factors, embryonic genes, and apoptosis genes in both tumor groups. For example, there was consistent representation of the Tgf-beta and Igf pathways in mesotheliomas from VDC-exposed and control rats; these pathways are important in tissue remodeling of many organs during disease and tumorigenesis, including mesothelioma (Garlepp and Leong 1995). Mesotheliomas from both vehicle control and VDC-exposed rats showed downregulation of the critical tumor suppressor Tp53, which is lost or downregulated in several human cancers including mesothelioma (Whitson and Kratzke 2006). There was downregulation of Lats2, a downstream regulator of the Hippo pathway, which is implicated in the pathogenesis and promotion of human mesothelioma along with TGFβ (Transforming growth factor; Fujii et al. 2012). The oncogenes MafB and Yes were consistently upregulated in mesotheliomas from both vehicle control and VDC-exposed rats, which are mediators responsible for cell proliferation in human malignant mesotheliomas (Gordon et al. 2005; Sato et al. 2012). The gene expression of Epcam, a transmembrane p-glycoprotein that has multiple roles in cell signaling, migration, proliferation, and differentiation (Patriarca et al. 2012), was upregulated in rat mesotheliomas. This gene is a well-known epithelial tumor marker that is overexpressed in breast, bladder, colorectal, pancreas, prostate, and ovarian neoplasia, as well as epitheliomatous mesotheliomas in humans (Ryan et al. 1997; Shield, Callan, and Devine 1994). Alterations in EpCAM protein interfere with cadherin-mediated cell-to-cell adhesion (Litvinov et al. 1997); cytoplasmic and nuclear translocation of this protein, as seen in rat mesotheliomas in this study, is associated with stimulation of cell proliferation (Maetzel et al. 2009).
Differences in common pathways between tumors from control and VDC-exposed animals were sometimes noted; for example, overrepresentation of DNA replication, recombination, and repair were observed as a statistically significantly overrepresented biologic function only in mesotheliomas from VDC-exposed animals, suggesting an association with these genes with VDC exposure that may reflect differences in DNA repair between the 2 tumor groups. At the gene level, increased expression of certain oncogenes was observed, including synuclein-γ(Sncg), dermatopontin (Dpt), and the embryonic gene placenta-specific 8 (Plac8). Synucleins are neural proteins primarily expressed in neural tissues and cancers (George 2002). In several human cancers, including colorectal, esophageal, and breast cancer (Amsterdam et al. 2012), SNCG is associated with tumorigenicity, cell proliferation and invasion (Ahmad et al. 2007), and DPT is an extracellular matrix protein that has a stimulatory affect on tumor growth in prostate cancer (Takeuchi et al. 2006). Several embryonic genes (Plac8, Wnt4, Gata5, Plau) were increased in mesotheliomas of VDC-exposed rats compared to Fred-PE mesothelial cells, and Plac8 was increased in VDC-associated mesotheliomas compared to spontaneous tumors.
Exposure to chemicals can cause genomic instability, mutational events, or cellular damage through various mechanisms including inflammation, direct cytotoxicity or reactive oxygen species, well-known contributors to carcinogenesis. Direct mesothelial cell damage can lead to injury responses including cell proliferation and/or the activation of the mesothelial cell innate immune response. Spontaneous mesotheliomas and mesotheliomas from VDC-exposed animals both showed overrepresentation of pathways associated with inflammation, and many constitutively active mediators associated with innate immunity in mesothelial cells were upregulated in both VDC and spontaneous mesotheliomas. However, based on significantly overrepresented canonical pathways, mesotheliomas from VDC-exposed animals showed a significant overrepresentation of pathways associated with an exaggerated pro-inflammatory phenotype and immune dysfunction. Several of the pathways that were significantly overrepresented in mesotheliomas from VDC-exposed animals, which were not observed in spontaneous mesothelioma, involved well-known pro-inflammatory pathways such as the NF-κB pathway and IL-8/IL-12 pro-inflammatory pathways, Fc receptor signaling, dendritic (DC) and natural killer (NK) cell signaling, and PI3/AKT signaling. The NF-κB pathway, of which PI3K/AKT molecules are a component of, is a key orchestrator between pathways mediating cell stress, inflammation, and cancer (Mantovani 2010). IL-8 plays a major role in chemotaxis of inflammatory cells, phagocytosis, and angiogenesis (Mukaida 2003), and IL-12 stimulates production of interferon-gamma (IFNγ) and TNF-alpha (TNFα) from cytotoxic T cells and natural killer (NK) cells (Wang, Frank, and Ritz 2000). Fc receptor signaling plays a major role in immune system function, including dendritic cells, cytotoxic T cells, and macrophages among others (Bulliard et al. 2013). Dendritic cells and NK cells are important in host responses to pathogens as well as tumor cells; NK cells function as cytotoxic T cells and function in tumor cell immunosurveillance by directly inducing death of tumor cells (Vivier et al. 2011), and DC cells sample tumor antigens and in turn activate a variety of T cells including NK cells for tumor targeting (Dhodapkar, Dhodapkar, and Palucka 2008). Furthermore, mesotheliomas in VDC-exposed animals showed upregulation of pro-inflammatory (Il8, Il12, Ccl5, Ccl6, Cxcl9) and downregulation of anti-inflammatory (Il10, Il18, Il24) cytokine and chemokine responses compared to spontaneous mesotheliomas. Direct damage to mesothelial cells from inflammation or reactive metabolites can lead to cell proliferation and/or innate immune response activation, and pro-inflammatory cytokines have been demonstrated to cause mesothelial cell proliferation (Mutsaers et al. 1997). In addition, there was overrepresentation of macrophage migration inhibitory factor (MIF) innate immunity and PI3K/AKT signaling in mesotheliomas from VDC-exposed animals. MIF is a pro-inflammatory cytokine that is necessary for optimal activation of other pro-inflammatory mediators such as TNFα and IL1 (Leng et al. 2003). It also regulates TLR4 function (Roger et al. 2001) and inhibits TP53-mediated apoptosis (Mitchell et al. 2002). Conversely, there was overrepresentation of cytotoxic T cell lymphocyte antigen 4 (CTLA4) signaling in spontaneous mesotheliomas compared to those from VDC-exposed animals; CTLA4 is a negative regulator of T cell activation, essentially limiting effective immune responses (Alegre and Fallarino 2006; Rudd, Taylor, and Schneider 2009). These pro-inflammatory and immune dysfunction features helped differentiate mesotheliomas in VDC-exposed animals from spontaneous background tumors, and suggest that these chemically induced tumors are influenced in part by pathways associated with inflammation and immune dysfunction. While the exact effects of VDC on mesothelial cells are not known, it has been shown that exposure to VDC results in the generation of reactive metabolites (1,1-diethylene oxide, chloroacetyl chloride), which have the potential to cause tissue damage and inflammation (Hathway 1977), which is an important contributor to tumorigenesis in a number of cancers, including mesothelioma (Colotta et al. 2009; Hanahan and Weinberg 2000, 2011).
The increased incidence of mesotheliomas in the study resulting from VDC exposure suggests that long-term exposure to VDC directly or indirectly affects mesothelial cell function. Our data support the possibility that VDC exposure increases mesothelioma incidence in F344/N rats through inducing a pro-inflammatory environment and exaggerated or dysfunctional immune response, resulting in direct or indirect damage to mesothelial cells. Chronic cytotoxicity may result in the accumulation of DNA damage through direct or indirect mechanisms, and result in genomic instability that increases an organism’s susceptibility to the development of cancer. Consistent with this, protein expression of gamma H2A histone family member X (γH2AX), a key molecule involved in DNA repair, was observed in mesotheliomas in VDC-exposed animals. Phosphorylation of this histone protein at double strand break sites leads to the accumulation of γH2AX, a sensitive marker for DNA double strand breaks (Bonner et al. 2008).
Finally, we used this study as an opportunity to address the relationship between interstitial cell tumors of the testes and the incidence of mesotheliomas in male F344/N rats. Previous studies have suggested that the presence of interstitial cell tumors plays a role in the development of mesothelioma arising from the tunica vaginalis (Haber et al. 2009; Maronpot et al. 2009; Tanigawa, Onodera, and Maekawa 1987). Hypotheses regarding this association include decreased androgen levels due to the presence of interstitial cell tumors, that trigger growth factor production, or overall increased size of the testis due to the presence of interstitial cell tumors, generating mechanical stress or physiological pressure to the peritoneal surface of the tunica vaginalis, and subsequent physical irritation of mesothelial lining cells. Results of this study indicated that there was no statistically significant association between the incidence of interstitial cell tumors, size of tumors, and overall testicular size with the presence of mesotheliomas. These data suggest that other factors, such as strain-related sensitivity to chemical exposure, are more likely to play a role in the development of mesothelioma than the incidence of testicular tumors.
Conclusion
Global gene expression profiling of mesotheliomas arising in F344/N rats exposed to VDC distinguished these tumors from spontaneously arising background lesions, despite an indistinguishable morphology. Furthermore, this study was the first to characterize chemically induced mesotheliomas in the context of spontaneous, background mesothelioma, and provides important information on the molecular characterization and potential mechanism of tumorigenesis of mesothelioma in VDC-exposed F344/N rats. Key pathways associated with cellular proliferation and survival, apoptosis inhibition, expression of a variety of oncogenes, and tumor suppressor genes, were similar. This suggests that these pathways are central to mesotheliomagenesis in the F344/N rat. Overall, the overrepresentation of genes associated with immune dysfunction, inflammation, and tissue injury in mesotheliomas associated with VDC-exposure suggests that exposure to VDC may lead to cytotoxicity and DNA damage resulting in regenerative hyperplasia and genomic instability, which are important risk factors for the development of neoplasia. In concert with a pro-inflammatory phenotype and underlying immune function dysregulation, these factors may act to increase the susceptibility of VDC-exposed animals to the development of mesothelioma, a background lesion that normally arises at a low rate in male rats of this strain.
Footnotes
Acknowledgments
We would like to thank the NIEHS Microarray, Histology, and Immunohistochemistry Cores for their technical expertise. We would like to thank Norris Flagler and Beth Mahler of the NIEHS Imaging Core for their imaging expertise and assistance with figures.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.