
Abstract
Male transgenic adenocarcinoma of the mouse prostate (TRAMP) mice are frequently used in prostate cancer research because their prostates consistently develop a series of preneoplastic and neoplastic lesions. Disease progression in TRAMP mouse prostates culminates in metastatic, poorly differentiated carcinomas with neuroendocrine features. The androgen dependence of the rat probasin promoter largely limits transgene expression to the prostatic epithelium. However, extra-prostatic transgene-positive lesions have been described in TRAMP mice, including renal tubuloacinar carcinomas, neuroendocrine carcinomas of the urethra, and phyllodes-like tumors of the seminal vesicle. Here, we describe the histologic and immunohistochemical features of 2 novel extra-prostatic lesions in TRAMP mice: primary anaplastic tumors of uncertain cell origin in the midbrain and poorly differentiated adenocarcinomas of the submandibular salivary gland. These newly characterized tumors apparently result from transgene expression in extra-prostatic locations rather than representing metastatic prostate neoplasms because lesions were identified in both male and female mice and in male TRAMP mice without histologically apparent prostate tumors. In this article, we also calculate the incidences of the urethral carcinomas and renal tubuloacinar carcinomas, further elucidate the biological behavior of the urethral carcinomas, and demonstrate the critical importance of complete necropsies even when evaluating presumably well characterized phenotypes in genetically engineered mice.
Introduction
The prostates of male transgenic adenocarcinoma of the mouse prostate (TRAMP) mice reliably exhibit a spectrum of preneoplastic and neoplastic lesions. Most notably, prostates of TRAMP mice develop poorly differentiated (neuroendocrine) carcinomas with widespread metastases (Shappell et al. 2004; Chiaverotti et al. 2008; Greenberg et al. 1995; Gingrich et al. 1996; Gingrich et al. 1999; Kaplan-Lefko et al. 2003; Ittmann et al. 2013). Strains of TRAMP mice are available for research on 2 different genetic backgrounds: pure C57BL/6 TRAMP mice and C57BL/6 TRAMP × FVB F1 animals. These 2 strains develop histologically similar lesions, although the disease progresses at a faster rate in C57BL/6 TRAMP × FVB F1 mice than in the C57BL/6 TRAMP mice (Gingrich et al. 1999). The histologic lesions of TRAMP mice have been described extensively, and grading schemes have evolved to improve descriptive and quantitative assessment of prostatic disease (Gingrich et al. 1999; Greenberg et al. 1995; Gingrich et al. 1996; Kaplan-Lefko et al. 2003; Berman-Booty et al. 2012; Suttie et al. 2003).
The lesions that develop within TRAMP mouse prostates result from prostate epithelial expression of the SV40 large and small T antigens (T Ag), which inhibit the tumor suppressor proteins p53 and Rb. Expression of T Ag in the TRAMP model is driven by the rat probasin promoter, an androgen-dependent promoter that largely restricts transgene expression to the prostate (Gingrich et al. 1999; Gingrich et al. 1996; Greenberg et al. 1995; Kaplan-Lefko et al. 2003). However, despite this relative specificity, a number of extra-prostatic lesions have been described in TRAMP mice, including neuroendocrine tumors of the urethra (Suttie et al. 2005), phyllodes-like tumors of the seminal vesicles (Tani et al. 2005), and renal tubuloacinar carcinomas (Suttie et al. 2005). Expression of T Ag has been demonstrated in all 3 of these lesions (Suttie et al. 2005; Tani et al. 2005).
In the present study, we report the identification and characterization of 2 novel extra-prostatic transgene-associated lesions in TRAMP mice, namely anaplastic tumors in the midbrain and poorly differentiated adenocarcinomas arising in the submandibular salivary gland. We describe the histologic and immunohistochemical features of these 2 new findings as well as provide insight regarding their biological behavior and incidence. In a parallel assessment, we also define in more detail the incidence and biological behavior of the previously reported urethral carcinomas and renal tubuloacinar carcinomas.
Material and Methods
Animals and Husbandry
Breeding pairs of C57BL/6 TRAMP mice were obtained originally from The Jackson Laboratory (Bar Harbor, ME), and FVB/NHsd mice were obtained from Harlan Laboratories (Indianapolis, IN). Both C57BL/6 TRAMP mice and C57BL/6 TRAMP × FVB F1 mice were subsequently produced at The Ohio State University (OSU) using the originally purchased breeding pairs and the FVB/NHsd mice. Mice were housed at OSU and genotyped as transgenic as previously described (Sargeant et al. 2007). All mice were provided food (specified subsequently) and water ad libitum. All protocols, experimental interventions, and procedures were approved in advance by OSU’s Institutional Animal Care and Use Committee.
Intact male mice (n = 141) on the C57BL/6 TRAMP × FVB F1 genetic background came from 3 dietary chemoprevention studies and 1 oral gavage study that were performed over the course of 3 years (2009–2012). In the dietary chemoprevention studies, mice were assigned to their respective experimental groups at 6 weeks of age and were fed the standard AIN-76A diet with or without an experimental compound (compound A, B, or C). The diets for compounds A and B were formulated by Research Diets, Inc. (New Brunswick, NJ). The AIN-76A diet containing compound C was acquired from Harlan Laboratories. Mice were fed their respective diets for 18 weeks or until a palpable prostate tumor greater than 1 cm in any 1 direction was noted; in addition, 3 mice with grossly visible masses in the submandibular region were removed before the intended study endpoint. For the oral gavage study, mice were randomized to either the vehicle group or experimental group at 10 weeks of age and then treated once daily with either vehicle or compound C for 8 weeks or until a palpable prostate tumor was detected; 1 mouse was removed at 14 weeks of age for a submandibular soft tissue mass. Mice surviving to the end of the 4 studies ranged in age from 14 to 24 weeks.
Two female C57BL/6 TRAMP mice were intact breeders being fed a standard rodent diet (8640 Teklad 22/5, Harlan Laboratories) ad libitum. Female 1 was euthanized at approximately 52 weeks of age due to seizures, while female 2 was euthanized at approximately 32 weeks of age due to a large submandibular mass.
Pathology Methods for Lesion Identification and Characterization
All animals were euthanized via carbon dioxide asphyxiation and then complete necropsies were performed. All tissues, excluding the spinal column, limbs, and carcass were fixed by immersion in 10% neutral buffered formalin, after which selected tissues were trimmed, processed by routine methods, and embedded in paraffin wax. Brains were fixed in situ, after which the intact heads were placed in Shandon TBD-2
Extra-prostatic lesions were found in multiple mice at 4 sites: midbrain, submandibular subcutis, urethra, and kidney. Brains were examined from 65 male mice from chemoprevention studies using compounds A and C and the oral gavage study, and also from the 2 female breeder mice. Submandibular salivary glands were examined in 46 male mice from chemoprevention studies utilizing compounds A, B, and C and the oral gavage study, and also in female breeder 2. Urethras distal to the prostate were collected and examined from 71 male mice from dietary chemoprevention studies that used compounds B and C and the oral gavage study. Kidneys were histologically evaluated in 141 male mice from chemoprevention studies employing compounds A, B, C, and the oral gavage study. The only clinical evidence for lesions at these extra-prostatic sites were seizures in 1 female mouse (indicative of brain involvement) and the grossly visible submandibular masses in 4 males and 1 female. None of the mice with submandibular masses exhibited dysphagia or dyspnea.
Immunohistochemistry
The brain and submandibular lesions were further characterized using indirect immunoperoxidase methods and a battery of primary antibodies to define the origin of the cells and to characterize the patterns of molecular expression within them.
Transgene expression was evaluated in 2 brain tumors and 2 submandibular masses by immunostaining for SV40 T Ag using a mouse anti-mouse monoclonal antibody (catalog# PAb416, Calbiochem-Millipore, Billerica, MA) at a dilution of 1:100. The staining protocol utilized the DAKO EnVision+ System (Dako, Carpinteria, CA).
An epithelial origin for the submandibular masses was explored in 1 neoplasm using a rat anti-mouse monoclonal antibody directed against cytokeratin 8 (CK8/Troma-1) at a dilution of 1:500. This antibody was obtained from the Developmental Studies Hybridoma Bank after being developed under the auspices of the U.S. National Institute of Child Health and Human Development and maintained by the Department of Biological Sciences at the University of Iowa (Iowa City, IA).
A possible neuroendocrine phenotype for the brain and submandibular lesions was assessed using a mouse anti-bovine monoclonal antibody against synaptophysin (catalog# M0776, Dako) at a dilution of 1:100.
The severely atypical appearance of cells in the brain neoplasms necessitated a broad search for a potential histogenetic origin. Accordingly, serial sections of tumor were immunostained for the epithelial marker cytokeratin AE1/AE3 (catalog# M3515, monoclonal mouse antihuman, Dako) at a dilution of 1:50; the mesenchymal marker vimentin (catalog# M0725, monoclonal mouse anti-pig, Dako) at a dilution of 1:100; the astrocyte marker glial fibrillary acidic protein (GFAP, catalog# Z0334, polyclonal rabbit anti-bovine, Dako) at a dilution of 1:5,000; the microglial marker ionized calcium–binding adaptor molecule 1 (Iba1, catalog# ab107159, polyclonal goat anti-rat, Abcam, Cambridge, MA) at a dilution of 1:400; and the S-100 marker for neural crest derivation (catalog# Z0311, polyclonal rabbit antihuman, Dako) at a dilution of 1:5,000.
DakoCytomation Target Retrieval and a Decloaking Chamber (Biocare Medical, Concord, CA) were employed according to the manufacturer’s instructions to unmask the epitopes for cytokeratin, S100, synaptophysin, and vimentin, while a proteinase K incubation (Dako) was utilized to reveal GFAP. The Polink-1 HRP Polymer and AP Polymer Detection Kit for mouse antibody on mouse tissue (Golden Bridge International, Mukilteo, WA) were used for detection of mouse anti-mouse antibodies according to the manufacturer’s instructions. Nonmurine primary antibodies were localized using biotinylated polyclonal goat secondary antibodies (Vector Laboratories, Burlingame, CA) at 1:1,000 followed by a standard Avidin-Biotin Complex method kit (Vector). For all antibodies, signal was developed with 3,3′-diaminobenzidine and sections were counterstained with hematoxylin.
Results
Anaplastic Tumors in the Midbrain
Eight neoplasms were identified in the brains of 67 mice (65 males and 2 females), yielding an incidence of 11.9%. Histologically, tumors were unencapsulated, expansile, and mildly infiltrative masses located ventral or adjacent to the mesencephalic aqueduct in the midbrain (Table 1 and Figure 1A). Invasion and partial obstruction of the aqueduct by neoplastic cells resulted in dilation of the third ventricle and mesencephalic aqueduct in female 1 (i.e., the animal with seizures).
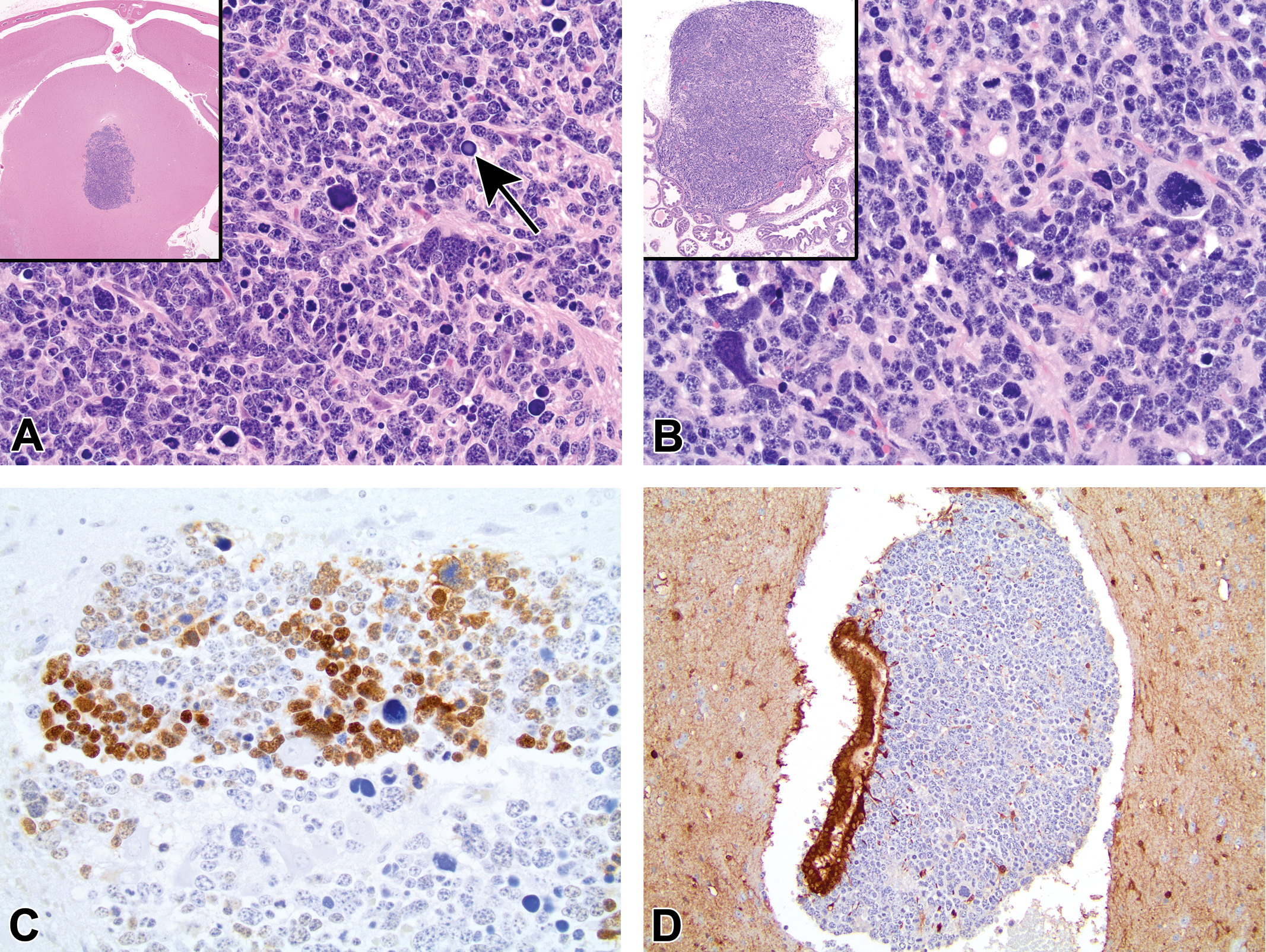
Histologic and immunohistochemical features of the anaplastic midbrain tumors. (A) The midbrain of affected mice contained unencapsulated, expansile, and mildly infiltrative masses located ventral to or at the level of the mesencephalic aqueduct (H&E, inset 20×). These masses were composed of sheets of markedly anaplastic cells with a marked degree of anisocytosis and anisokaryosis. Occasional neoplastic cells contained homogenous basophilic nuclei (black arrow). Mitotic figures were common. (H&E, 400×). (B) The histologic appearance of the neoplastic cells within the anaplastic midbrain tumors was morphologically indistinguishable from poorly differentiated prostate carcinomas with neuroendocrine differentiation of TRAMP mice (H&E, 400×, inset 40×). (C) Neoplastic cells of the midbrain tumors were multifocally immunopositive for the SV40 T Ag (SV40 T Ag, 400×). (D) Cells of the midbrain tumor invaded the mesencephalic aqueduct in female 1. Neoplastic cells were immunonegative for S-100, while the nearby ependymal cells were intensely immunopositive (S-100, 200×).
Features of TRAMP mice (n = 8) with anaplastic midbrain tumors.a

aAll mice are C57BL/6 TRAMP × FVB F1 with the exception of female 1, who was C57BL/6 TRAMP.
The midbrain tumors were composed of sheets of markedly anaplastic, polygonal cells with high nuclear-to-cytoplasmic ratios, mild amounts of eosinophilic cytoplasm, and a marked degree of anisocytosis and anisokaryosis (Figure 1A). Nuclei were oval to elongated and contained stippled basophilic chromatin and numerous prominent nucleoli. Mitotic figures, including highly irregular spindles, were numerous and often quite large. Multiple neoplastic cells had homogenous basophilic nuclei or were undergoing necrosis with nuclear fragmentation (Figure 1A). The tumor foci frequently entrapped normal elements of the neuropil, including large neurons. The histologic appearance of neoplastic cells within these anaplastic brain tumors was indistinguishable from the poorly differentiated prostate carcinomas with neuroendocrine differentiation that have been described in the prostates of TRAMP mice (Figure 1B). T Ag expression in the nuclei of tumor cells within the brain masses was mild to strong but tended to occur in patches (Figure 1C). Neoplastic cells did not express cytokeratin AE1/AE3, vimentin, GFAP, Iba1, or S-100 (Figure 1D), but rare cells were positive for synaptophysin.
Half (4/8, 50%) of the animals with midbrain tumors lacked a tumor in any other organ (i.e., prostate, urethra, kidney, and salivary gland; Table 1). No correlation was observed between the diet each mouse received (control vs. experimental) and the development of a midbrain tumor.
Submandibular Salivary Gland Adenocarcinomas
Grossly identifiable, firm, submandibular nodules (Figure 2A) were identified in 4 male mice and female breeder 2 (Table 2). Careful dissection revealed that the masses appeared to originate from the submandibular salivary gland (Figure 2B). The masses were large, with the smallest measuring 8 × 6 × 5 mm. The nodules generally were tan to white but often were speckled with dark red, small, and irregular foci.
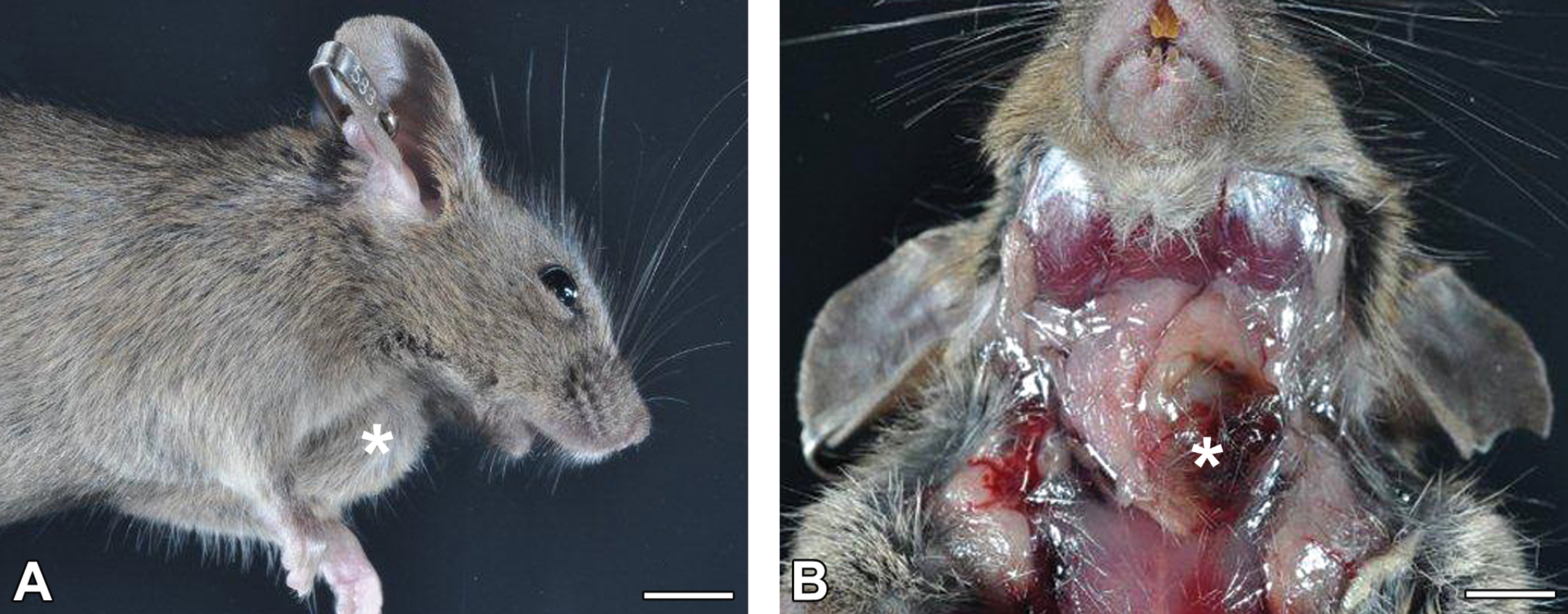
Gross appearance of the submandibular salivary gland adenocarcinomas. (A) The soft tissue of the ventrolateral neck of a male mouse was expanded by a large, firm mass. (B) Careful dissection of the mouse’s neck revealed an 8 × 6 × 5 mm nodule (*) originating from the left submandibular salivary gland. The bar for both panels represents 0.5 cm.
Features of TRAMP mice (n = 5) with submandibular salivary gland tumors.a

aAll mice are C57BL/6 TRAMP × FVB F1 with the exception of female 2 who was C57BL/6 TRAMP.
Histologically, the affected submandibular salivary glands contained densely cellular, partially encapsulated, expansile, and infiltrative adenocarcinomas (Figure 3A). The lesions consisted of sheets, lobules, and rarely small distorted acini composed of polygonal cells with large, oval, basophilic nuclei; multiple small nucleoli; modest quantities of pale basophilic cytoplasm; and indistinct cell borders (Figure 3A and B). Cells exhibited high nuclear-to-cytoplasmic ratios and moderate to marked degrees of anisocytosis and anisokaryosis. Mitotic figures were common and sometimes abnormal (Figure 3B). Irregular lakes of nascent or complete liquefactive necrosis were scattered throughout the neoplasms. A desmoplastic response was associated with cells that had invaded through the capsule. Neoplastic cells expressed T Ag (Figure 3C) and the epithelial marker CK8 (Figure 3D), while the acini and ducts of the adjacent nonneoplastic salivary tissue only expressed CK8. Neoplastic cells were immunonegative for synaptophysin.
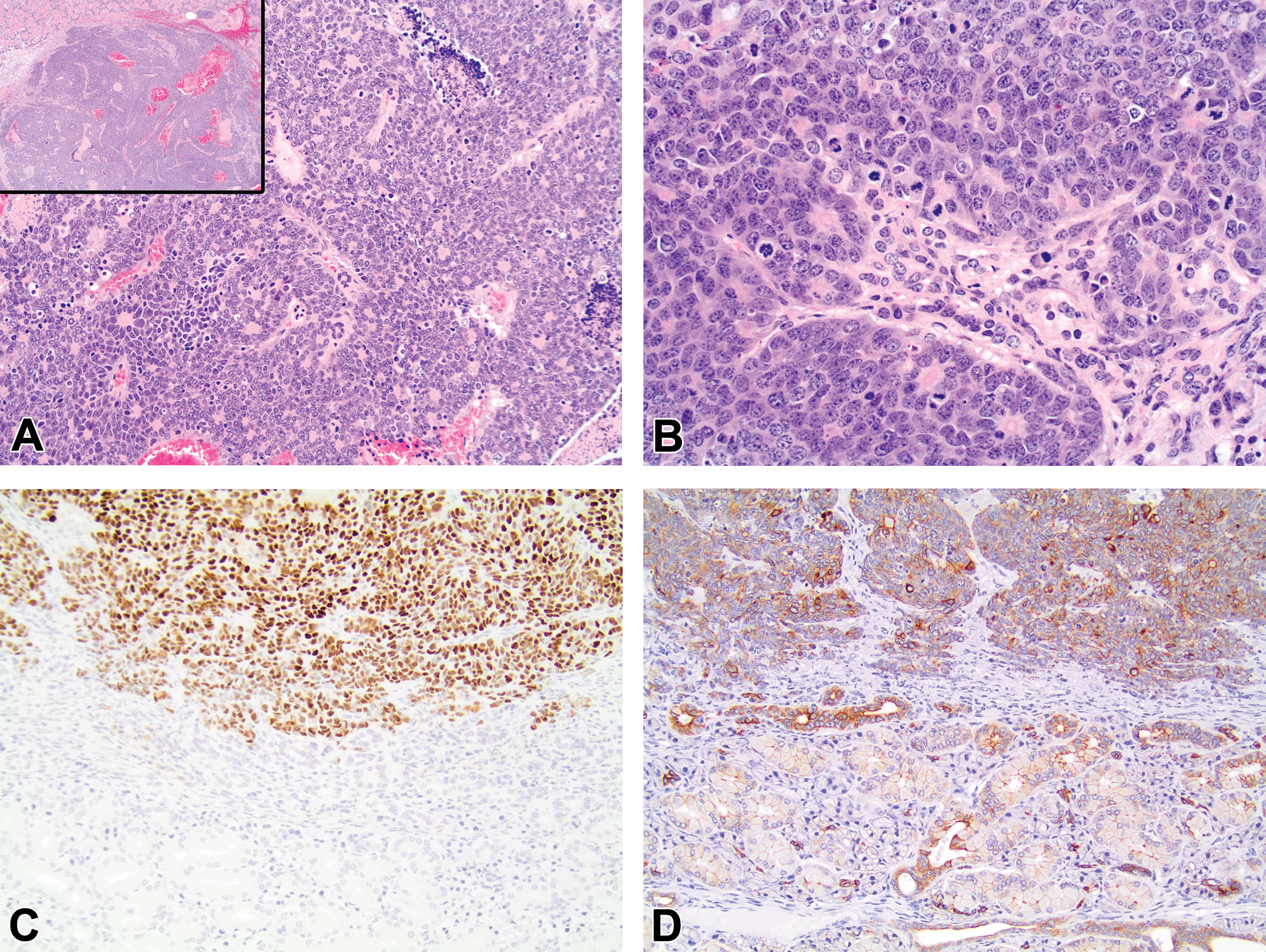
Histologic and immunohistochemical features of the submandibular salivary gland adenocarcinomas. (A) The affected submandibular salivary gland contained a densely cellular, partially encapsulated, expansile, and infiltrative mass that consisted of sheets, lobules, and, less frequently, acini (A: H&E, 200×, inset 40×). (B) Neoplastic cells contained large oval nuclei with prominent nucleoli and a moderate to marked degree of anisocytosis and anisokaryosis. Mitotic figures were common (H&E, 400×). (C) Neoplastic cells (top half of image) were immunopositive for SV40 T Ag, while normal salivary epithelium (bottom half) was immunonegative (SV40 T Ag, 200×). (D) Neoplastic cells (top half of image) and also the adjacent non-neoplastic salivary gland acini and ducts (bottom half) stained for cytokeratin 8 (CK8, 200×).
The incidence of poorly differentiated salivary gland adenocarcinomas in TRAMP mice was 10.6% (5 of the 47 animals). None of the mice with salivary gland adenocarcinomas had primary prostate or urethral carcinomas. Submandibular salivary glands were histologically within normal limits in the 42 male mice that had no salivary gland neoplasm.
Urethral Carcinomas and Associated Preneoplastic Changes
Histological examination of the urethras from 71 intact male TRAMP mice (age range, 16 to 24 weeks) demonstrated 19 urethral carcinomas (Table 3), yielding an incidence of 26.8%. Although the histological appearance and immunohistochemical features of tumors of the urethral submucosal glands have been described (Suttie et al. 2005), the urethral tumors in our studies were only occasionally identifiable as originating from either the submucosal glands (Figure 4A) or urothelium (Figure 4B). An urothelial origin was attributed to tumors that appeared to extend from the surface epithelium into the underlying submucosa with sparing of the periurethral glands. More commonly, the exact site of origin could not be determined, as these masses involved both the urothelium and the glands, greatly expanded the urethral submucosa, and extended into either the urethral lumen and/or urethral skeletal muscle (Figure 4C).
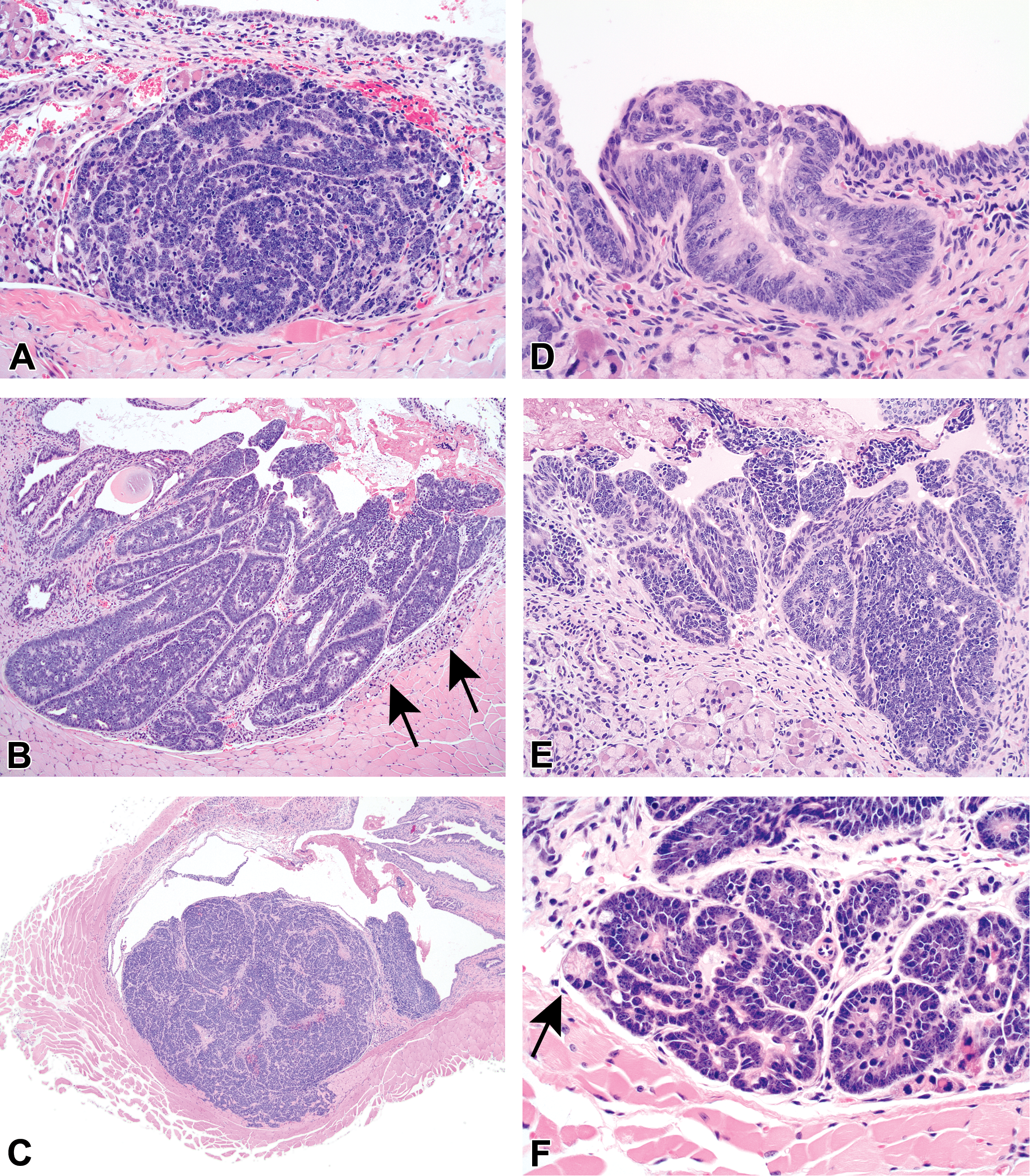
Histological appearance of the urethral lesions. (A–C) Urethral carcinomas occasionally were identifiable as originating from the submucosal glands (A, H&E, 200×) or the urothelium (B, H&E, 100×). Tumors originating from the urothelium were not associated with the prostate and completely spared the subjacent submucosal glands; compression of the unaffected submucosal glands by the urothelial tumor is delineated by the black arrows (B). More commonly, the urethral carcinomas involved both the urothelium and glands, so an exact site of origin could not be determined (C, H&E, 40×). In all cases, the carcinoma cells formed sheets or, less frequently, rosettes. Tumor cells were polygonal to elongated, contained oval to elongate nuclei with prominent nucleoli, and were extremely atypical as indicated by a marked degree of anisocytosis and anisokaryosis, high mitotic index, and many bizarre mitotic figures. (D) Atypical hyperplasia of the urothelium was characterized by proliferating urothelial cells with polygonal profiles, oval nuclei, and such evidence of pending neoplasia as high nuclear-to-cytoplasmic ratios, cellular atypia, and multiple mitotic figures. These atypical cells stratified but never invaded through the mucosal basement membrane (H&E, 400×). (E&F) Atypical hyperplasia with features of carcinoma in situ of the urothelium (E) or submucosal glands (F) featured proliferating cells exhibiting increased degrees of dysplasia, anisocytosis, anisokaryosis, and an increased mitotic rate compared to atypical hyperplasia. There was also loss of polarity and cells projected into the urethral lumen or filled the lumen of the affected gland without breaching the basement membrane (H&E, E: 200×, F: 400×). Note in the case of atypical hyperplasia with features of carcinoma in situ of the submucosal glands that atypical cells can be found in glands that are still partially composed of normal epithelium (F, black arrow).
Evolution and incidencea of urethral lesions in male TRAMP mice with concurrent poorly differentiated prostate carcinoma.

aSome mice had more than 1 urethral lesion. Values represent the number (percentage) of mice with a given lesion.
In addition to the overt urethral carcinomas, we observed a series of nonneoplastic and preneoplastic lesions in the urothelium and urethral submucosal glands that required a more detailed description (Table 3). These lesions included a nonneoplastic change that has been previously characterized (Suttie et al. 2005), namely proliferative tubules within the submucosal glands, as well as a novel subset of preneoplastic lesions: atypical hyperplasia of the urothelium or submucosal glands, sometimes with features of carcinoma in situ. In atypical urothelial hyperplasia, proliferating polygonal cells had high nuclear-to-cytoplasmic ratios, numerous mitotic figures, and stratified organization within the mucosa without invasion through the mucosal basement membrane (Figure 4D). In atypical hyperplasia of the urothelium or in submucosal glands where features of carcinoma in situ were evident, proliferating cells also exhibited higher degrees of dysplasia, anisocytosis, anisokaryosis, an increased mitotic rate with many bizarre mitoses, and loss of polarity with projection into the urethral or glandular lumens (Figure 4E and F).
Renal Tubuloacinar Carcinomas
Renal tubuloacinar carcinomas were identified in 3 of the 141 intact male TRAMP mice (age range, 18 to 24 weeks), resulting in a tumor incidence of 2.1% (Table 4). Two of the tumors were grossly visible as destructive renal neoplasms and were histologically confirmed to represent tubuloacinar carcinomas (Figure 5A and B). One early tumor was identified after routine histologic examination of the kidney (Figure 5C and D). The histologic appearance of these tumors was as previously described (Suttie et al. 2005).
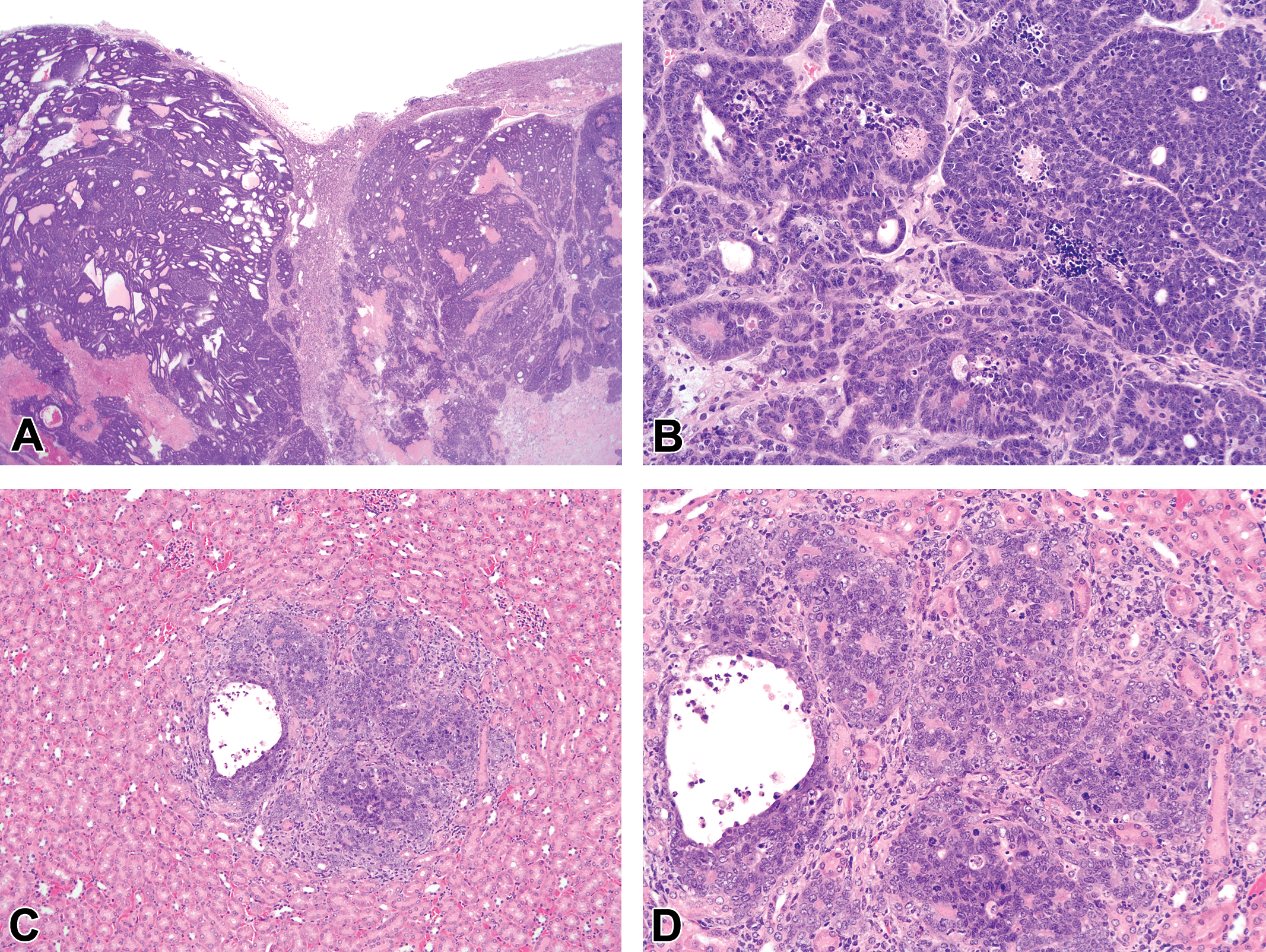
Histologic appearance of the renal tubuloacinar carcinomas. (A and B) The renal parenchyma in 2 mice was largely obliterated by an unencapsulated, infiltrative, expansile, and densely cellular mass composed of acini and tubules that consisted of extremely atypical cells (H&E, A: 20×, B: 200×). (C and D) In 1 mouse, the kidney contained a single focus of carcinoma characterized by a small, poorly demarcated nodule comprised of small tubules and acini that were similar in appearance to the neoplastic cells in more advanced renal tumors (H&E, C: 100×, D: 200×).
Features of TRAMP mice (n = 3) with renal tubuloacinar carcinomas.

Metastatic Behavior of Extra-prostatic Neoplasms in TRAMP Mice
Mice with anaplastic tumors of the midbrain did not exhibit histologic evidence of extra-cerebral metastatic disease. Of the 5 mice with salivary gland adenocarcinomas, pulmonary macrometastases were observed in 4 of the animals (80%; Figure 6A and B and Table 2), but no metastatic disease was found in any other organ. Of the 19 mice with urethral carcinomas, 3 (15.8%) had evidence of pulmonary micrometastases (Figure 6C and D). The prostates from these same 3 mice lacked histologic evidence of a concurrent poorly differentiated prostate carcinoma. Therefore, these pulmonary metastatic lesions were interpreted to be of urethral origin. The metastatic behavior of the renal tubuloacinar renal carcinomas has already been described (Suttie et al. 2005). We found similar results, with the 2 mice with grossly visible renal tubuloacinar carcinomas having metastases in the lung and liver.
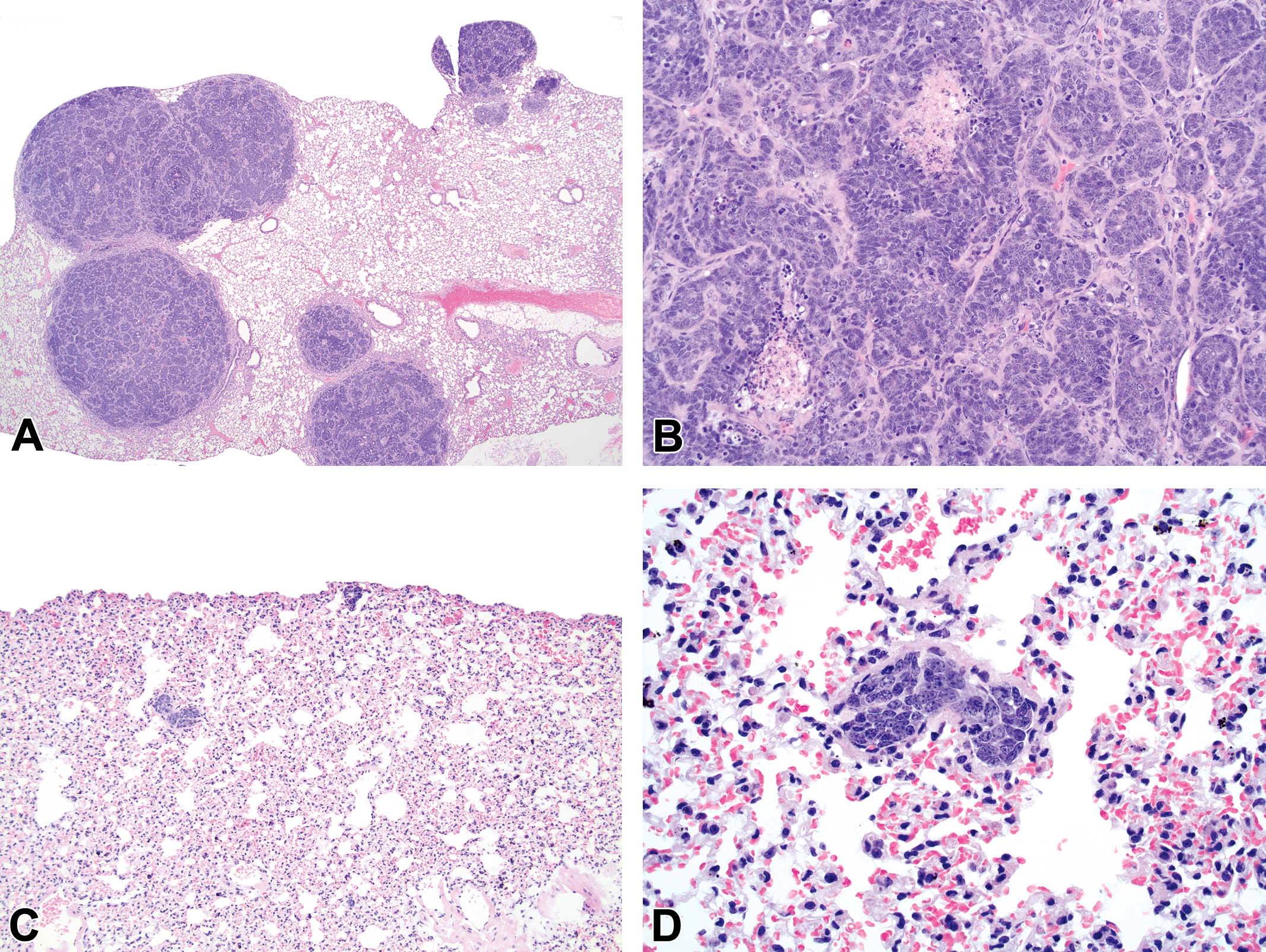
Metastatic behavior of the salivary gland adenocarcinomas and urethral carcinomas. (A) Pulmonary macrometastases were identified grossly and confirmed histologically in 4 of the 5 mice with submandibular salivary gland adenocarcinomas (H&E, 20×). (B) As in the primary submandibular salivary gland adenocarcinomas, neoplastic cells in the metastatic lesions formed sheets, lobules, and occasional acini (H&E, 200×). (C) Pulmonary micrometastases were identified upon routine histological evaluation of the lung in 15.8% (3/19) of mice with urethral carcinomas (H&E, 100×). (D) The pulmonary micrometastases were composed of tightly packed aggregates of neoplastic cells that closely resembled the cells within the primary urethral carcinomas (H&E, 400×).
Discussion
Male TRAMP mice are an extensively employed mouse model of prostatic neoplasia. The initiation of preneoplastic changes in prostate epithelium and the formation of poorly differentiated prostatic carcinomas result from the expression of SV40 T Ag under the control of the rat probasin promoter. Although this promoter originally was thought to be prostate-specific, extra-prostatic, T Ag-expressing primary neoplasms have been described in both the kidney and urethra of TRAMP mice (Suttie et al. 2005). In this additional characterization of the TRAMP mouse model, we have further investigated these urinary tract lesions. However, we have also detected and herein provide the initial detailed characterization of 2 novel transgene-expressing, extra-prostatic primary neoplasms, namely anaplastic tumors of the midbrain and poorly differentiated submandibular salivary gland adenocarcinomas. Single instance of tumors in the caudal brain (a brainstem/cerebellum medulloblastoma in a 272-day-old female C57BL/6 TRAMP mouse; The Jackson Laboratory [JAX] Mouse Tumor Biology Database [http://www.tumor.informatics.jax.org/mtbwi/index.do, Accession ID J: 94307]) and the submandibular region (a “neck tumor” requiring euthanasia, but not subjected to further analysis [Bonorden et al. 2009]) have been reported previously as incidental background lesions. However, since the SV40 T Ag expression status of these 2 tumors was not defined, we cannot assess whether or not these tumors developed spontaneously or actually were related to extra-prostatic transgene expression.
The transgene-associated nature of the brain and salivary gland neoplasms in our mouse colony was supported by tumor cell immunoreactivity for the SV40 T Ag. An acinar epithelial origin was proposed for the submandibular salivary gland adenocarcinomas since tumor cells formed occasional acini and were immunopositive for CK8 and immunonegative for synaptophysin. In contrast, the cell of origin for the anaplastic tumors of the midbrain could not be determined definitively despite an extensive immunohistochemical analysis for expression of cell type-specific biomarkers; our antibody battery ruled out epithelial, mesenchymal, astrocytic, microglial, and ependymal cells as potential lineages of origin. However, given the tumor location and the fact that occasional neoplastic cells were immunopositive for synaptophysin, we hypothesize that these tumors were of neuroendocrine derivation. It should be noted that the anaplastic midbrain tumors had an immunohistochemical profile similar to that of the poorly differentiated prostate carcinomas with neuroendocrine differentiation that develop in TRAMP mice (Kaplan-Lefko et al. 2003), with brain tumor cells lacking cytokeratin reactivity but demonstrating some reactivity against synaptophysin. This finding may also support the hypothesis that the anaplastic midbrain tumors arose from a neuroendocrine stem cell and bolsters the hypothesis that prostate carcinomas in TRAMP mice develop from a neuroendocrine lineage. Importantly, the anaplastic midbrain tumors we observed in our mouse colony lacked such stereotypical medulloblastoma traits as elongated primitive neurons forming pseudorosettes and whorls (Kaufmann et al. 2012). Thus, we are unsure whether the previously identified medulloblastoma (JAX Mouse Tumor Biology Database Accession ID J: 94307) represents a variant of our transgene-associated anaplastic midbrain tumors or alternatively is a distinct tumor type that arose coincidentally in the same general region.
Interestingly, the anaplastic brain tumors were only found in the midbrain, and the salivary gland adenocarcinomas were always located in the submandibular salivary gland. The consistent tumor locations and the fact that both tumor types were identified in male mice that lacked genitourinary tumors as well as in female mice strongly supported our interpretation that these neoplasms were not merely metastases seeded from primary prostate tumors. Instead, these neural and salivary masses likely arose from endogenous cell populations within their respective tissues of origin under the influence of extra-prostatic transgene expression. The consistent submandibular location of the salivary gland adenocarcinomas also lent credence to our conclusion that these tumors were not of mammary gland origin. It would be unlikely for a transgene-induced neoplasm of mammary origin to consistently affect only the most cranial glands and always invade the submandibular salivary tissue. Furthermore, after careful review of the literature, we could not find any reports of spontaneous mammary gland adenocarcinomas in male mice. Collectively, these findings attest to the local origin of these novel brain and salivary tumors. A topic for future study will be to ascertain the presumed neuroendocrine lineage from which the midbrain masses arise.
The development of transgene expressing primary tumors in the midbrain and submandibular salivary gland may be related to the fact that the SV40 T Ag is driven by an androgen-dependent promoter (Greenberg et al. 1995; Gingrich et al. 1999; Gingrich et al. 1996). Androgen receptor expression has been described in the submandibular salivary gland of mice (Sawada and Noumura 1995), and androgen induction of gene expression in the submandibular salivary gland has been clearly demonstrated (Treister et al. 2005; Senorale-Pose et al. 1998). Therefore, transgene expression could have been induced by androgens in the submandibular salivary gland resulting in neoplastic transformation. Androgen receptor expression has also been identified in neurons of the periaqueductal gray matter of the midbrain of rats (Loyd and Murphy 2008; Murphy, Shupnik, and Hoffman 1999). Although not described in mice, a similar location of androgen receptor-expressing neurons in the mouse midbrain could explain the localization of the anaplastic brain tumors. We could not explain why transgene-associated lesions have not been identified in other tissues that express androgen receptors, such as the testes. However, the possibility remains that aberrant SV40 T Ag expression may have occurred in an androgen-independent manner. In that case, extra-prostatic expression of the SV40 T Ag with subsequent neoplastic transformation may be due to the ability of unknown transcription factors to activate the rat probasin promoter, as was originally suggested in the case of neuroblastomas that arose from the taste buds of male and female SV40 T Ag-transgenic rats (Asamoto et al. 2001).
The anaplastic midbrain tumors and submandibular salivary gland adenocarcinomas were observed in both male and female mice, although the male mice developed these tumors at a younger age than did the females. This difference may relate to the genetic background of the TRAMP mouse strain. Both female mice were pure C57BL/6 TRAMP, while all the male mice were C57BL/6 × FVB F1. It is well known that male C57BL/6 × FVB F1 TRAMP mice develop poorly differentiated prostate carcinomas at a younger age than C57BL/6 TRAMP mice (Gingrich et al. 1999). Therefore, the difference in the age at which mice developed the extra-prostatic lesions may have been due to strain-related differences. Alternatively, if the development of these extra-prostatic transgene-associated lesions was dependent on the presence of androgen and/or androgen receptor, we would anticipate that female mice would develop lesions at an older age because of lower circulating androgen levels. Because neither the anaplastic tumors of the midbrain nor the adenocarcinomas of the submandibular salivary gland were identified in any male C57BL/6 TRAMP mice, we could not determine whether the differences in the age at which these lesions developed were related to variations in strain or sex. Additionally, a prospective evaluation of brains and salivary glands from transgenic C57BL/6 females across multiple ages is needed in order to better characterize the true incidence of these tumors in female TRAMP mice and compare it to similarly aged males.
In addition to characterizing 2 novel transgene-associated extra-prostatic lesions, our data also extend the previous report documenting the histologic appearances of the urethral carcinomas in a nonspecified number of TRAMP mice and renal tubuloacinar carcinomas in 2 mice (Suttie et al. 2005) by enhancing our understanding of their biological behavior. Our investigation of these organs in dozens of male TRAMP mice indicated that the incidence of the urethral neoplasms was 26.8% (19/71), while that of renal tumors was 2.1% (3/141). Therefore, these lesions should be anticipated as relatively common additional findings in this mouse model. Our work identified additional facets regarding the biological behavior of the urethral carcinomas that were not addressed in the original report (Suttie et al. 2005). First, we demonstrated that the urethral carcinomas could arise in 2 sites, the periurethral submucosal glands (as previously documented) and the urothelium (which represents a new variant). Second, we showed that these tumors metastasized widely to the lungs. Third, we described preneoplastic lesions leading to urethral carcinoma in the periurethral glands and urothelium. The only other described urethral lesion is the foci of proliferative tubules within the submucosal glands, which is believed to be nonneoplastic (Suttie et al. 2005). Taken together, these findings further our understanding of these 2 extra-prostatic neoplasms in this model.
In conclusion, our present work is chiefly important because it characterizes 2 novel transgene-associated, extra-prostatic, primary neoplasms that develop in the midbrain and submandibular salivary gland in male and female TRAMP mice, and it also further clarifies the biological behavior of previously described transgene-linked renal and urethral tumors. Given the histological similarity between some of these extra-prostatic lesions and TRAMP mouse prostate tumors, it is imperative that future TRAMP mouse studies include a thorough (i.e., lobe-by-lobe) evaluation of the prostate—as was done in the present study—for evidence of a small primary tumor, to exclude the remote possibility that the extra-prostatic neoplasms represent aberrant metastases. Extra-prostatic lesions found in mice with prostate tumors should be compared to the known histologic appearance, incidence, and location of previously described metastases. Lesions that differ in histologic appearance, incidence, or organ of identification should be further investigated, since they may represent novel extra-prostatic lesions. Furthermore, this article highlights the importance of performing complete necropsies followed by histologic evaluation of multiple organs in all genetically engineered mice even when the phenotype is assumed to be well understood. Narrowly focusing on only the “organ of interest” for a given genetically engineered model may result in ignoring or missing lesions at a distance from the known target organ. Routine histologic evaluation of a wide subset of organs (Bregman et al. 2003) should be performed beyond the initial phenotyping phase of new genetically engineered models, since the appearance and development of extra-target organ lesions is expected to be unpredictable. Finally, this work reiterates that researchers should be cognizant of potential leakiness in organ-specific promoters rather than believing that the “specific” promoter truly limits the expression of a transgene to the targeted organ of interest. This leakiness may be due to more widespread expression of the recognized factors that drive the expression of these promoters (e.g., widespread expression of the androgen receptor which is then able to bind androgens and drive the rat probasin promoter) or may be due to alternate transcription factors activating transgene expression.
Footnotes
Acknowledgment
The authors acknowledge Ms. Anne Saulsbery and Ms. Florinda Jaynes in the Comparative Pathology and Mouse Phenotyping Shared Resource (CPMPSR) at The Ohio State University for excellent histotechnological and immunohistochemical support, respectively.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a T-32 Institutional National Research Service Award in Mouse Models of Human Disease (Ruth L. Kirschstein-NRSA; T32RR007073) and NIH grant K01 OD010463 to LDBB and NIH grant CA112250 and Department of Defense Prostate Cancer Research Program grant W81XWH-09-0198 to C.-S.C. The Comparative Pathology & Mouse Phenotyping Shared Resource is supported in part by NIH grant P30 CA016058.