
Abstract
This review article is designed to serve as an introductory guide in neuroanatomy for toxicologic pathologists evaluating general toxicity studies. The article provides an overview of approximately 50 neuroanatomical subsites and their functional significance across 7 transverse sections of the brain. Also reviewed are 3 sections of the spinal cord, cranial and peripheral nerves (trigeminal and sciatic, respectively), and intestinal autonomic ganglia. The review is limited to the evaluation of hematoxylin and eosin–stained tissue sections, as light microscopic evaluation of these sections is an integral part of the first-tier toxicity screening of environmental chemicals, drugs, and other agents. Prominent neuroanatomical sites associated with major neurological disorders are noted. This guide, when used in conjunction with detailed neuroanatomic atlases, may aid in an understanding of the significance of functional neuroanatomy, thereby improving the characterization of neurotoxicity in general toxicity and safety evaluation studies.
Background
The National Toxicology Program (NTP) recently expanded the neuroanatomical subsites screened in studies investigating environmental and other potential toxicants based on the rapid rise in human degenerative neurological diseases and the exponential increase in various forms of childhood and juvenile learning deficits. The process for improved sampling of the nervous system started in 2010, when the NTP developed a compact disk–read only memory (CD-ROM; Sills and Little 2010) detailing the modified evaluation (including images and examples of lesions) of the nervous system in NTP studies (Sills and Little 2010). The modified NTP protocol (Rao et al. 2011) includes the evaluation of 7 sections of the brain (Figure 1) compared to the traditional 3-section approach. Following the publication by Rao et al. in 2011, the NTP organized a Neuropathology Symposium, held on April 23–24, 2012, at the National Institute of Environmental Health Sciences (NIEHS). The NTP approach is consistent with the recommended practices for sampling the nervous system by the Society of Toxicologic Pathology (Bolon et al. 2013).
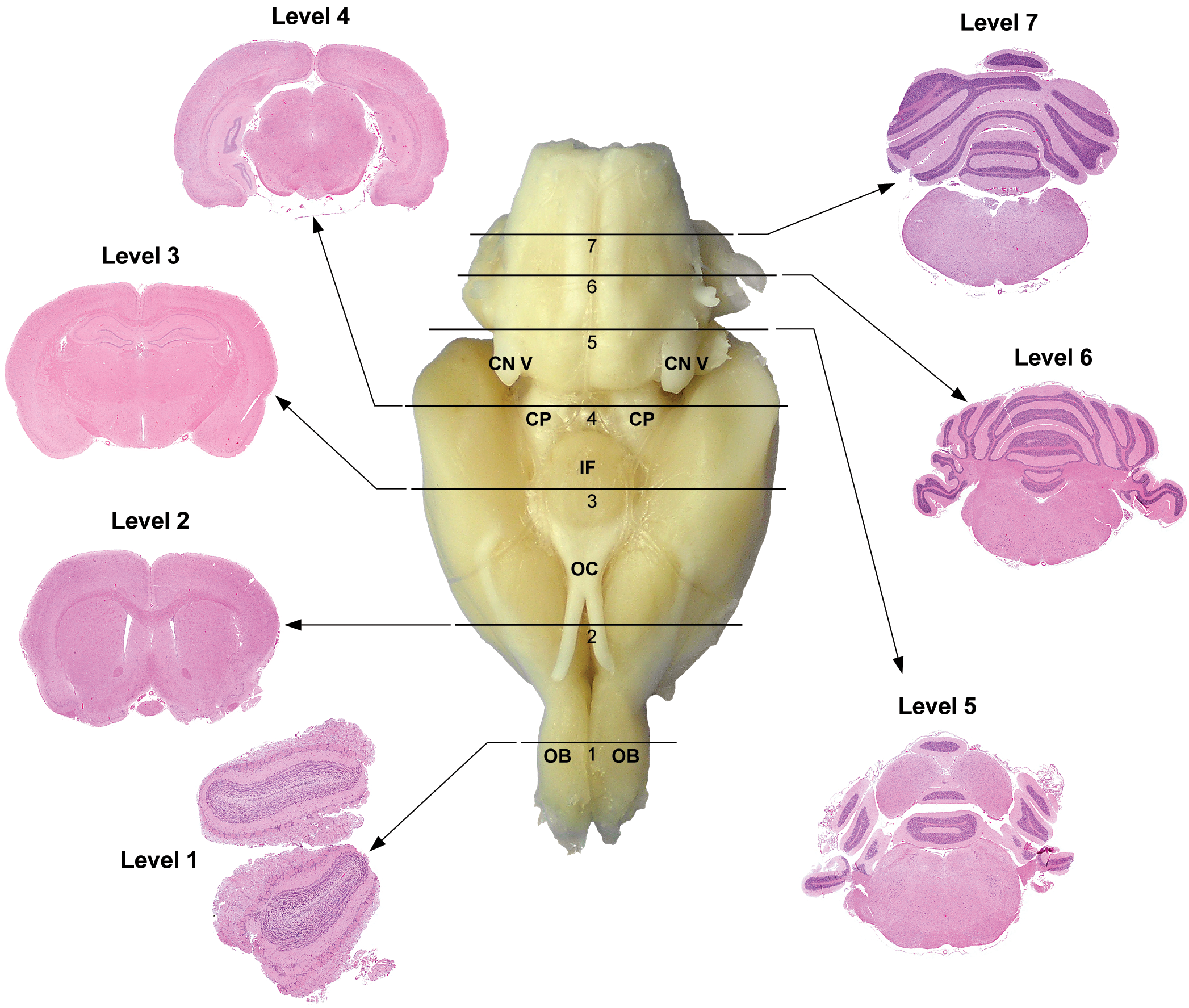
Rat: H&E. Seven transverse sections corresponding to levels based on anatomic target landmarks. Level 1 is at the mid-level olfactory bulb (OB). Level 2 is approximately 1 to 2 mm rostral to the optic chiasm (OC). Level 3 is taken at the infundibulum (IF) and/or median eminence. Level 4 corresponds to the superior colliculus, while level 5 corresponds to inferior colliculus. Level 6 is at the mid-cerebellum, at the root of the eighth or vestibulocochlear cranial nerve (CN VIII). Level 7 is taken 2 to 3 mm rostral to the caudal termination of the cerebellum. CP = cerebral peduncle.
The present document is published to create wider awareness in the toxicologic pathology community of the new standards expected by NTP in its efforts to improve the detection of neurotoxicants in rodent test species. Specifically, this review is designed to complement the original article (Rao et al. 2011) by providing a navigational guide to the neuroanatomic localization and functional significance of selected subsites within the NTP-7. It is not intended to replace existing neuroanatomic texts by any means but is designed primarily for toxicologic pathologists involved in routine evaluation of hematoxylin and eosin (H&E)-stained sections in general toxicity and safety evaluation studies. Although there are several excellent detailed rodent brain atlases available, there is no known existing comprehensive rodent reference guide that depicts neuroanatomical subsites and/or functional neuroanatomy in specifically H&E-stained sections. This article would be most beneficial when used in conjunction with standard rodent brain atlases (Franklin and Paxinos 2008; Kruger, Saporta, and Swanson 1995; Paxinos and Watson 2009; Paxinos et al. 2008). In addition to toxicologic pathologists, it may be useful to experimental neurotoxicologists, neurobiologists, and regulatory scientists.
Introduction
The nervous system is complex from an anatomic, physiologic, and toxicologic standpoint. Although pathological changes are generally limited to a characteristic spectrum of cellular alterations, there is presently no way to reliably predict where neurotoxicity is likely to occur, especially in response to unknown toxicants. For example, some lesions such as astrocyte swelling can result in widespread changes observed in response to energy deprivation following hypoxia (Salkowski and Penney 1994). Other lesions may be subtle and specific changes limited to selectively small but functionally significant neuroanatomic subsites such as those seen with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in the substantia nigra (Switzer, Lowry-Franssen, and Benkovic 2011). Further, neuropathology can be daunting because of the numerous neuronal populations (at least ∼600 distinct subsites are noted in rodent brain atlases) and the inherent complexity of neuronal pathways and circuitry among the various neuronal populations.
A review of the literature demonstrates innumerable experimental studies on well-documented functional neuroanatomic circuits such as learning and memory, auditory, cognitive/attention, locomotor, and addiction pathways. These studies often include sophisticated and sensitive techniques including special stains, immunohistochemistry, in situ hybridization, and occasionally, electron and confocal microscopy. However, there is limited information on functional neuroanatomy of rodents relevant to general toxicity and safety evaluation studies to identify and capture potential neurotoxicants with undefined anatomical targets and mechanisms of action (Defazio et al. 2009; Switzer, Lowry-Franssen, and Benkovic 2011).
Results of a survey of current practices for sampling of the nervous system in general toxicity and safety evaluation studies were presented at the Joint Symposium of the Society of Toxicologic Pathology and the International Federation of Societies of Toxicologic Pathologists in Chicago in 2010 (Andrew-Jones et al. 2010). The survey results showed that, in rodents, all respondents (100%) included histopathologic examination, and the majority (97%) included gross examination of the brain as well. Histopathologic evaluation was primarily by coronal (i.e., transverse) sections (69% of respondents), while H&E (47%) was the most commonly used stain (Andrew-Jones et al. 2010).
Although brain sectioning varies across a spectrum that includes the traditional 3-section approach (Morawietz et al. 2004; Radovsky and Mahler 1999; Solleveld and Boorman 1990) at one end, and, up to ∼60 sections on the other end (Switzer, Lowry-Franssen, and Benkovic 2011); typically, between 3 (70% of survey respondents) and 8 (29%) sections (Andrew-Jones et al. 2010) have been commonly evaluated in toxicologic pathology (Bolon et al. 2013; Defazio et al. 2009). Typically, in NTP studies, the brain is trimmed per NTP specifications (NTP January 2011) to obtain multiple consecutive transverse brain slices (corresponding to 7 sections) that is then processed to obtain 2 (mouse) or 3 (rat) tissue blocks. With the exception of any remnant rostral olfactory bulb fragments, the advantage of this approach is that the whole brain is maintained within tissue blocks for any potential future evaluation.
The following review is limited to the identification and brief functional relevance (based on susceptibility to toxicity or association with neurological disorders) of approximately 50 neuroanatomic subsites (Table 1, 2 and Figure 2) noted in 7 transverse sections of brain adopted in the recently revised NTP protocol (Rao et al. 2011). The article is organized to begin with each of the 7 transverse sections of the brain (labeled as NTP-7, level 1 through NTP-7, level 7) taken progressively from the rostral to the caudal brain. Each NTP-7 level is identified with a target anatomical landmark used during brain trimming (Figure 1). It is important to note that modest variations in trimming planes are expected variations in brain sampling among animals within a single study, across multiple studies, and among institutions. In addition, an overview of the spinal cord, cranial and peripheral nerves (trigeminal and sciatic), and autonomic ganglia is included. While Figures 3 to 9 highlight the anatomic subsites at each NTP level, Figure 2 summarizes the anatomic subsites at each of the NTP levels in one figure and has been included for easy referencing. Table 2 summarizes the functional significance of prominent subsites across the 7 sections of the brain. The terminology for subsites that are identified based on stereotaxic orientation (such as the superior colliculus vs. anterior colliculus vs. rostral colliculus) were chosen to be consistent with the Paxinos rat and mouse brain atlases (Paxinos and Watson 2009; Paxinos et al. 2008) to allow easy cross-navigational reference. Similarly, abbreviations follow the Paxinos rat brain atlas for the most part. The references cited for functional significance were primarily from the rat and mouse, and rarely from other species when data in rodents were lacking.
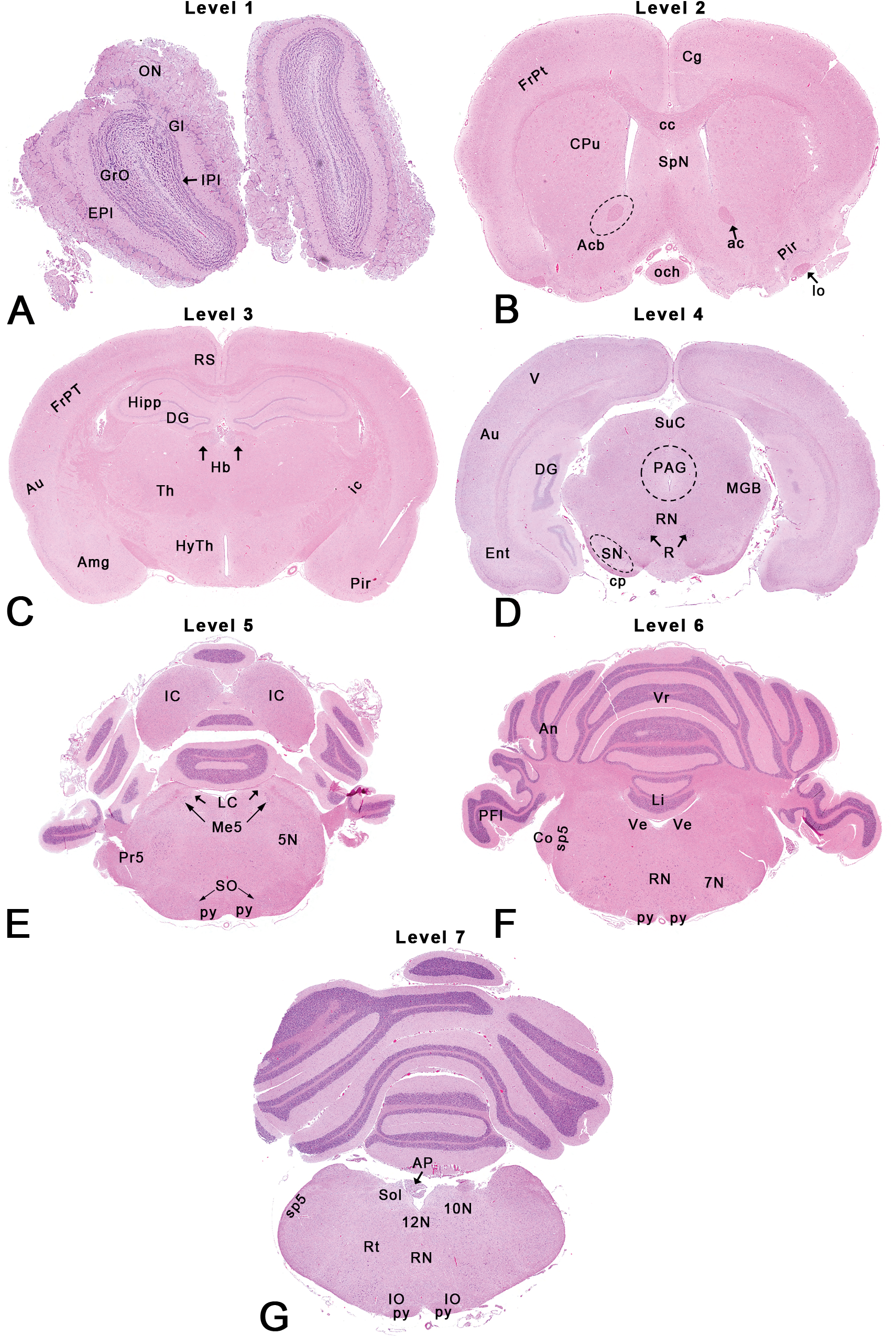
Rat: H&E. Seven transverse sections depicting neuroanatomical subsites. Abbreviations in italics below correspond to abbreviations used in the Paxinos rat brain atlas (Paxinos and Watson 2009). Dotted outlines are close approximations and use of an atlas is recommended for accuracy.
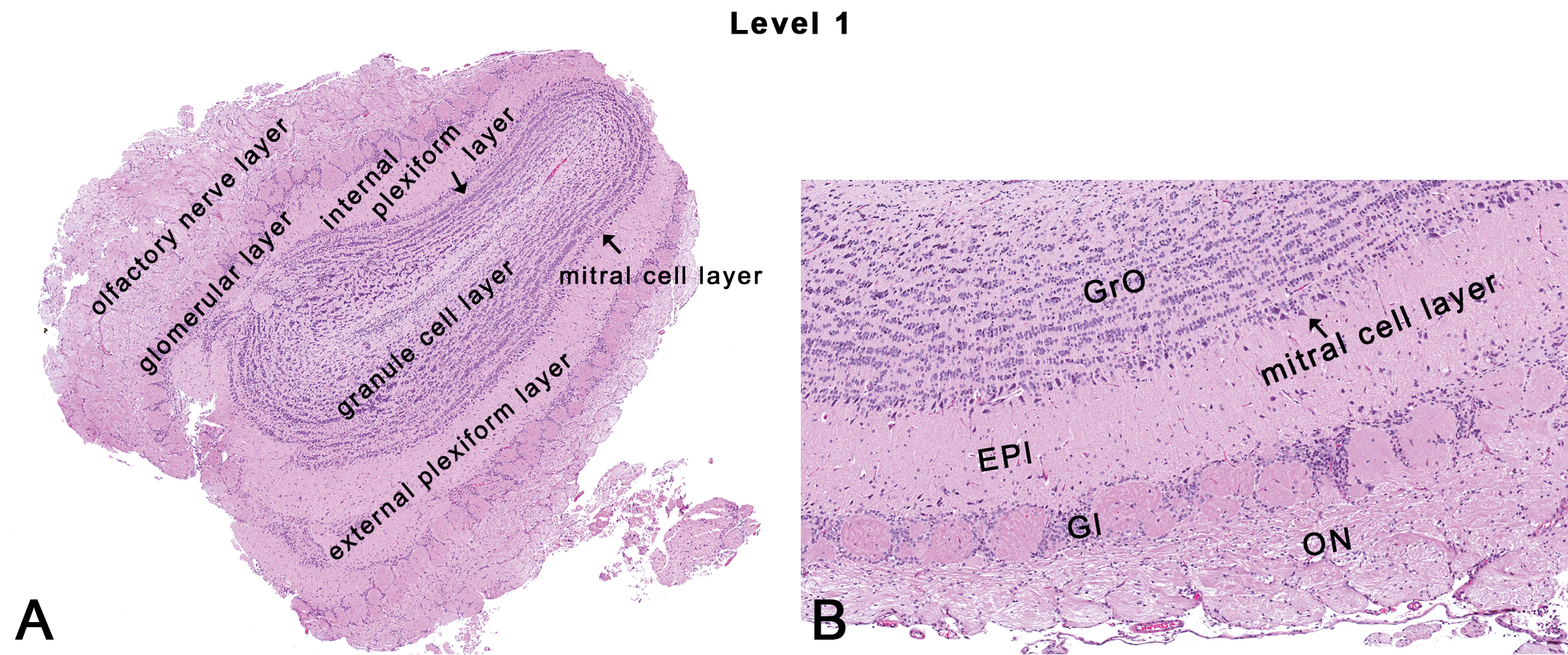
Rat: H&E. Transverse section at NTP-7, level 1. EPl = external plexiform layer; Gl = glomerular layer; GrO = granule cell layer; ON = olfactory nerve layer.
Summary of the various neuroanatomic subsites that can be seen corresponding to each of the 7 NTP section levels of the brain.
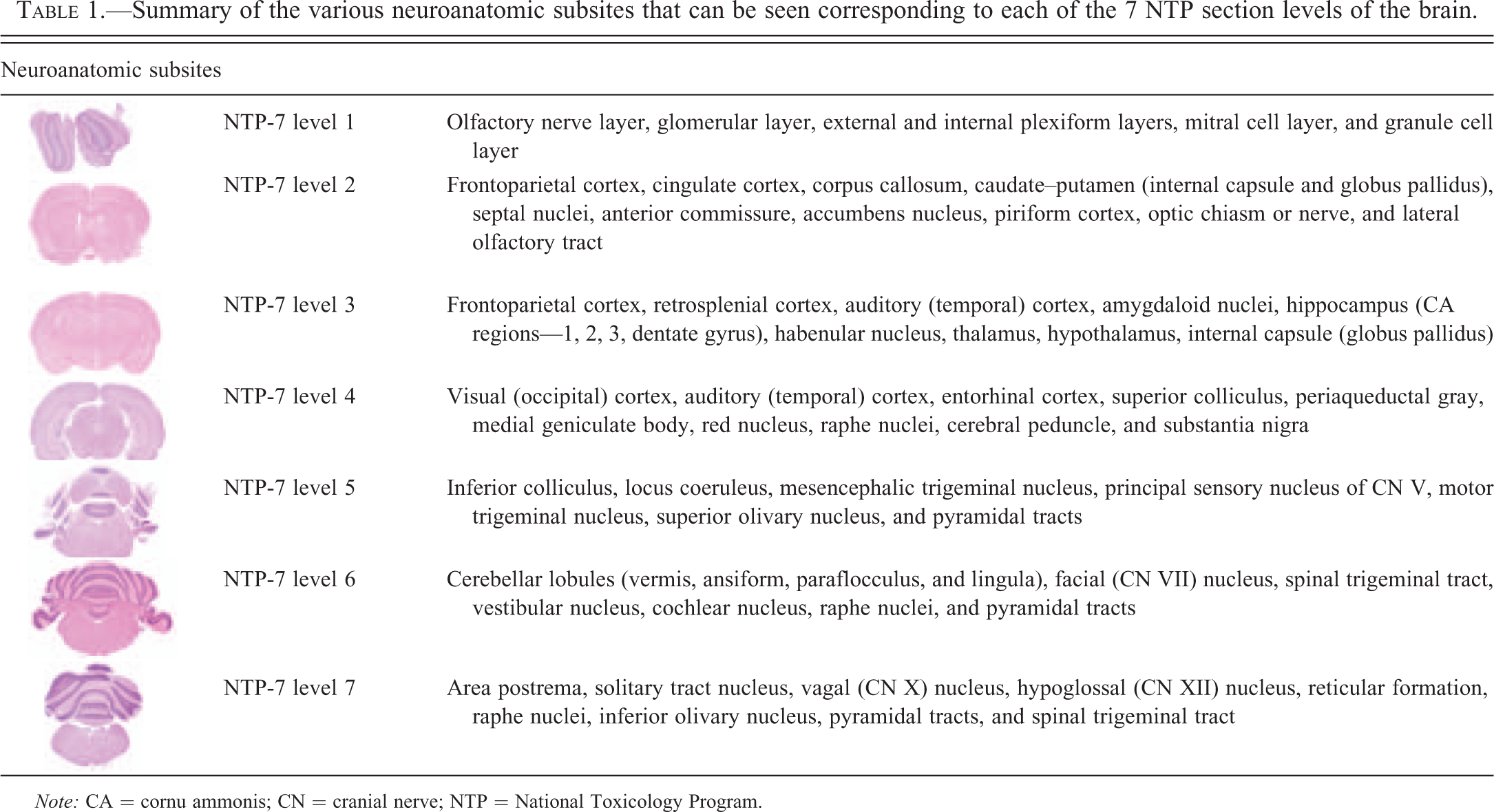
Note: CA = cornu ammonis; CN = cranial nerve; NTP = National Toxicology Program.
Summary of the functional significance corresponding to the various neuroanatomic subsites.
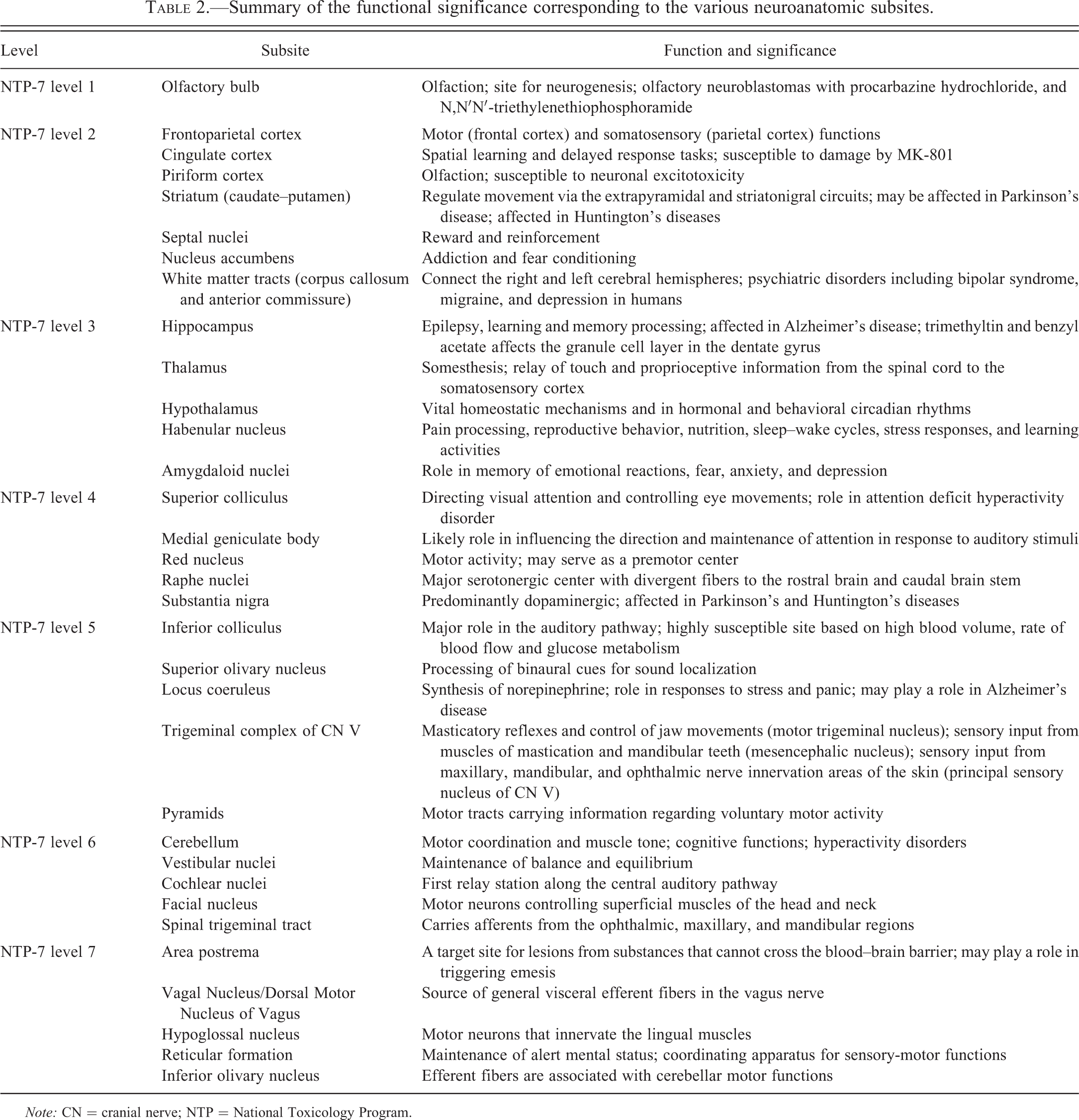
Note: CN = cranial nerve; NTP = National Toxicology Program.
Brain Sections
NTP-7, Level 1
This section is typically taken at the mid-level of the olfactory bulb (Figures 1, 2A, and 3). Grossly, the olfactory bulb is 6–7% of the total brain weight (Bolon et al. 2013) and lies immediately behind the cribriform plate of the ethmoid bone that separates the nasal and cranial cavities. In rodents, the olfactory bulbs are typically prominent. Histologically, there are multiple layers of the olfactory bulb that can be readily recognized and evaluated easily by light microscopy (Figure 3). These include the olfactory nerve layer, the glomerular layer, the external plexiform layer, the mitral cell layer, the internal plexiform layer, and the granule cell layer (Figure 3A). The olfactory nerve layer consists of nerve fibers that originate from the olfactory epithelium of the nose and terminate as a dense plexus on the surface of the olfactory bulb. The glomerular layer contains 1 or 2 rows of neuropil-rich spheroid structures surrounded by a distinctive shell of small neurons and glial cells (Figure 3B). The external plexiform layer is wide with low cell density and very dense neuropil. The mitral cell layer contains somata of triangular mitral cells (resembling a bishop’s miter) arranged for the most part, in a monolayer. The internal plexiform layer is thin, adjacent to the mitral cell layer (Figure 3A), with low cell density and many neurites. The internal granule cell layer contains 3 to 5 granule cells arranged in row-like aggregates of tightly packed somata (Shipley, McLean, and Ennis 1995). The cells in the various layers of the olfactory bulb form a synaptic circuit that extends from the nose to multiple areas in the brain such as the piriform cortex, the entorhinal cortex, and some amygdaloid nuclei (Genter and Bolon 2011; Shipley, McLean, and Ennis 1995). Additionally, in rats, the olfactory receptor neurons in the vomeronasal organ, an auxiliary olfactory sense organ connect with the accessory olfactory bulb. This accessory olfactory bulb can occasionally be seen in caudal sections of the olfactory bulb.
The olfactory bulbs are part of the olfactory pathway. Although the olfactory pathway is not as well delineated as the visual or auditory pathways, it is considered important for reproductive/maternal functions (Yoon, Enquist, and Dulac 2005), neuroendocrine regulation (Bruce 1959) and emotional and behavioral responses such as aggression (Guillot and Chapouthier 1996; Thorne 1993). Since the olfactory bulb is a site for neurogenesis (Lennington, Yang, and Conover 2003; Whitman and Greer 2009) in adults, the probability of identifying toxicants affecting cell migration and differentiation is increased here. It has been demonstrated that neuroblasts from the subventricular zone migrate along the rostral migratory stream into the olfactory bulb (Jacquet, Ruckart, and Ghashghaei 2010), where they differentiate mostly into granule cells. In toxicologic pathology, examination of the olfactory bulb is important in inhalation studies.
The unique positioning of the primary olfactory receptor neurons in the nasal mucous membrane allows inhaled substances to potentially have direct contact with the brain, thus bypassing the blood–brain barrier (BBB). In fact, drug delivery strategies for therapeutic agents that circumvent the BBB for therapeutic agents in treating neurological disorders have been active areas of research (Illum 2002, 2003). There are several examples of inhaled toxicants impacting the olfactory bulb. For example, inhalation studies in rodents have shown that manganese deposited within the nose can undergo direct transport to the brain along the olfactory nerve (Aschner, Erikson, and Dorman 2005). Other metals such as uranium (Monleau et al. 2005) and cobalt (Persson, Henriksson, and Tjalve 2003) are known to accumulate in the olfactory bulb following inhalation exposure. Neoplastic lesions include nasal olfactory neuroblastomas extending into the brain following exposure to procarbazine hydrochloride (National Cancer Institute [NCI] 1979) and N,N′N′-triethylenethiophosphoramide (NCI 1978). Nonneoplastic lesions include loss of dendritic spines and neuronal vacuolation in the granule cells of the olfactory bulb (Colin-Barenque et al. 1999) following exposure to ozone. However, it is important to note that pathological changes in the olfactory bulb are not limited to exposure via the inhalation route (Colman et al. 2005).
NTP-7, Level 2
This section is taken 1 to 2 mm rostral to the optic chiasm (Figures 1, 2B, and 4). Neuroanatomic subsites include several cortical areas (frontoparietal cortex, cingulate cortex, and piriform cortex), the caudate–putamen (striatum), the septal nuclei, the accumbens nucleus, the anterior commissure, and the corpus callosum (Figure 4).
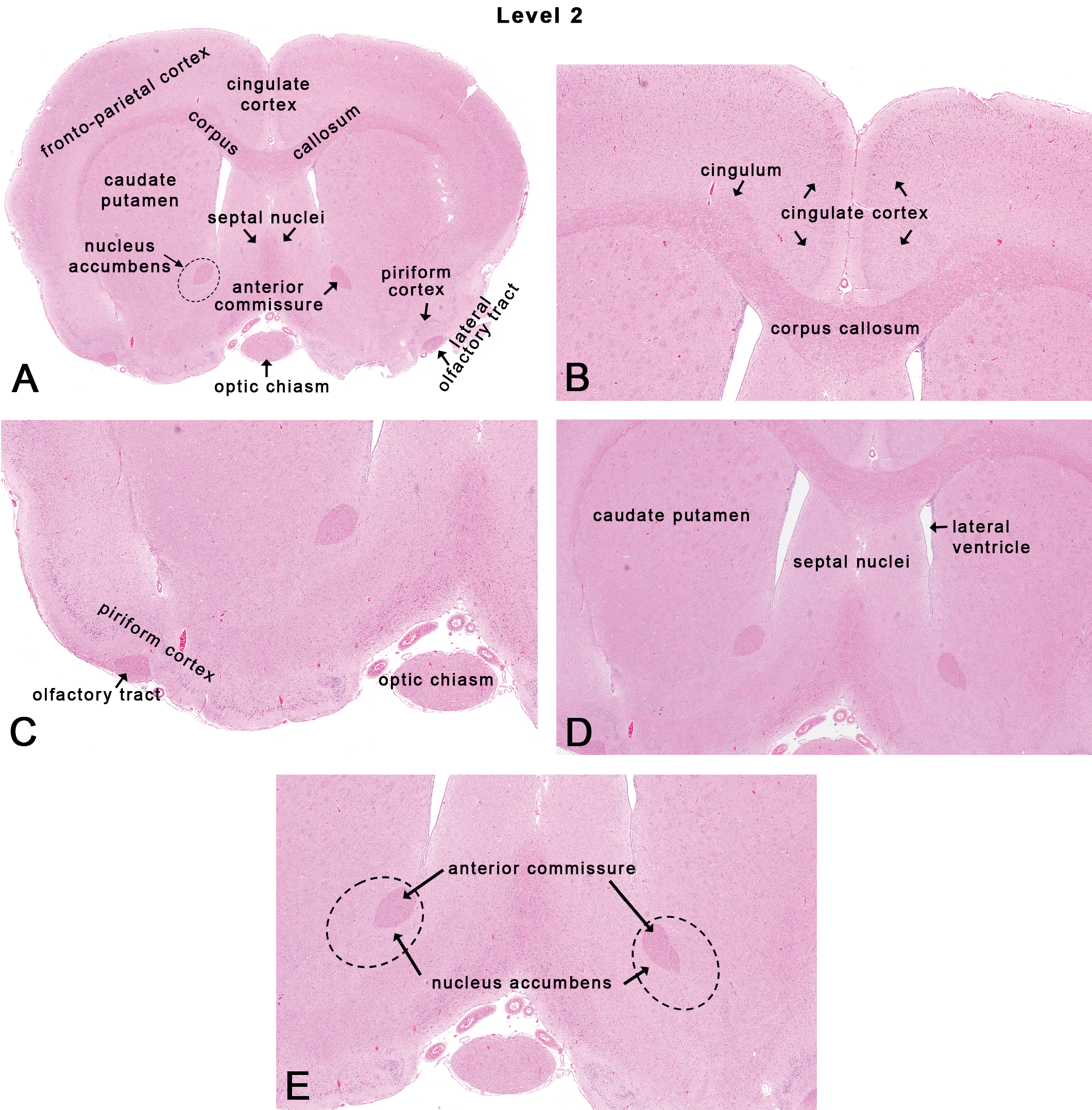
Rat: H&E. Transverse section at NTP-7, level 2. Dotted lines in Figure 4A and E depict the general area of the accumbens nucleus.
The cortex is a mantle of cells on the surface of the cerebrum. Based on phylogeny, the rat cortex is generally divided into an isocortex and an allocortex. The isocortex is synonymous with the neocortex (most evolutionarily recent) and comprises 4 major regions (frontal, parietal, auditory [temporal], and visual [occipital]) that serve specific functions consistent within each mammalian species. The allocortex is synonymous with the paleocortex (olfactory) and archicortex (limbic), and receives input from the olfactory bulb. Although the rodent brain is normally lissencephalic, the cortex is still organized in a somatotopic fashion, and maps of regions in the cortex are consistent between individuals of the same species. Histologically, the isocortex is generally comprised of 6 roughly comparable layers depicted traditionally by Roman numerals (I–VI), while the allocortex has a laminar structure that is quite heterogeneous. Toxicants are known to affect migration of cells to their respective layers during development. For example, developmental exposure to ethanol impacts neuroblast migration resulting in cortical dysplasia in offspring (Aronne et al. 2011).
The frontoparietal cortex expands over the dorsolateral aspects of the rostral cerebral hemispheres (Figures 4A and 5A). Histologically, the frontal motor cortex lacks an inner granular layer IV when compared to the presence of a prominent layer IV in the somatosensory parietal cortex. In fact, in rats, layer IV is characteristically prominent in the portion of the somatosensory cortex that receives input from the vibrissae (via the thalamus), this being referred to as the “barrel cortex” (Welker 1976). Functionally, in the rodent, the frontal cortex reflects the motor functions, while the parietal cortex is the somatosensory cortex that receives projections from cutaneous mechanoreceptors and proprioceptors.
Rat: H&E. Transverse section at NTP-7, level 3 depicting subsites on a section taken at the midpoint of the infundibulum and/or median eminence. Dotted lines closely approximate the medial and lateral habenular nuclei.
The medial part of the frontal cortex contains the cingulate cortex (Figure 4B) that is part of the limbic system (allo- or archicortex). Destruction of the cingulate cortex in rodents results in deficits in spatial learning, including delayed response tasks (Eichenbaum, Clegg, and Feeley 1983). Also, neuronal vacuolation, necrosis, and gliosis were demonstrated in the posterior cingulate and retrosplenial cortex of rats administered MK-801 (an N-methyl-D-aspartate receptor antagonist) that is commonly referred to as Olney’s lesions in rodents (Fix, Wightman, and O’Callaghan 1995; Fix et al. 1995; Olney, Labruyere, and Price 1989).
The piriform cortex or primary olfactory cortex forms the ventrolateral edge of the piriform lobe (Figure 4C). The piriform cortex is part of the allo- or paleocortex that is characterized by fibers coming from the main olfactory bulb. Histologically, the piriform cortex is comprised of 3 to 4 tightly packed cell layers. Functionally, the piriform cortex represents the most extensive and important olfactory area in the brain for the integration and subsequent transmission of the incoming olfactory information (Switzer, De Olmos, and Heimer 1985). Efferent projections include but are not limited to the hypothalamus, thalamus, hippocampus, and amygdaloid nuclei (Switzer, De Olmos, and Heimer 1985). The piriform cortex is particularly susceptible in a variety of animal models to excitatory neurotoxic injury (Candelario-Jalil et al. 2001; Fatzer et al. 2000). Histologically, within the vicinity of the piriform cortex is a prominent narrow, triangular band of axons called the lateral olfactory tract (Figure 4C) that connects the mitral cells of the olfactory bulb to several target regions in the brain including the piriform cortex, the amygdaloid nuclei, and the entorhinal cortex. Also, on the ventral midline aspect is the optic chiasm (Figure 4C), a tract composed of partially crossed optic nerve fibers (Cunningham and Freeman 1977) that sends input to the lateral geniculate nuclei and the superior colliculi.
The basal “ganglia” include a group of several nuclei (ganglia is really a misnomer) located in the central regions of the cerebral hemispheres. Major nuclei include the caudate, the putamen, the globus pallidus, and their associated traversing white matter tracts that are collectively referred to as the corpus striatum. The corpus striatum is so named to reflect the “striated” mass of dispersed white matter tracts representing corticofugal or corticopetal projections that gradually collect to form the internal capsule (Heimer, Alheid, and Zaborszky 1985). In most mammals, the internal capsule separates the caudate nucleus from the putamen. However, in the rat, the indistinct separation of the caudate, the putamen, and the globus pallidus by the internal capsule precludes easy identification of each component, so that the caudate nucleus and the putamen are considered as one structure—the caudate–putamen (Heimer, Alheid, and Zaborszky 1985). In rodents, the caudate–putamen is generally referred to as the striatum or neostriatum. The striatum receives fibers from the cerebral cortex and substantia nigra and sends fibers to the globus pallidus. While the NTP-7 level 2 section of the brain clearly shows the caudate–putamen (Figure 4D), the NTP-7 level 3 often reveals the globus pallidus laterally adjacent to the internal capsule (Figure 5C).
The primary function of the corpus striatum is to regulate movement and influence various types of learning via the extrapyramidal motor circuit and the striatonigral circuits. The corpus striatum is relevant in neurodegenerative diseases such as Parkinson’s disease (Mehler-Wex, Riederer, and Gerlach 2006) where there is eventual loss of dopaminergic neurons that project to the striatum from the substantia nigra and ventral tegmental region. Loss of gamma-aminobutyric acidergic medium spiny neurons of the corpus striatum is also observed in Huntington’s disease (Ehrlich 2012) and other movement disorders.
The septal nuclei lie between the rostral horns of lateral ventricles, ventral to the corpus callosum, and dorsal to the midline portion of the anterior commissure (Figure 4A and D). The septal nuclei generally include medial and lateral components. Functionally, the septal nuclei are part of a pathway for fibers from the hippocampus and amygdaloid nuclei into and through the hypothalamus and brain stem. Along with the accumbens nucleus, the septal nuclei play a role in reward and reinforcement (Olds and Milner 1954).
The accumbens nucleus is a ventral extension of the head of the caudate nucleus (Figure 4E). It forms the main part of the ventral corpus striatum. This area has traditionally been studied for its role in addiction (Wolf 2010). In 1954, Olds and Milner (1954) found that rats with metal electrodes implanted into their accumbens nucleus and the septal nuclei repeatedly pressed a lever activating this region, and did so in preference to eating and drinking, eventually dying of exhaustion. In rats, the accumbens nucleus may also be involved in fear conditioning (Schwienbacher et al. 2004).
There are three main interhemispheric commissural tracts connecting both cerebral hemispheres: the corpus callosum (Figure 4B), hippocampal commissure, and the anterior commissure (Figure 4E). The corpus callosum is composed of millions of axons that integrate neural activity between the two cerebral hemispheres and is usually larger in male rats than female rats (Berrebi et al. 1988). Another tract that facilitates interhemispheric communication is the anterior commissure. Specifically, it connects the temporal lobes and amygdaloid nuclei between the hemispheres. Like the corpus callosum, the anterior commissure is larger in male rats than in female rats (Noonan et al. 1998). More recently, lesions in white matter tracts have been associated with psychiatric disorders including bipolar syndrome, migraine, and depression in humans based on imaging studies (Alves et al. 2012; Mahon, Burdick, and Szeszko 2010; Yu et al. 2013).
NTP-7, Level 3
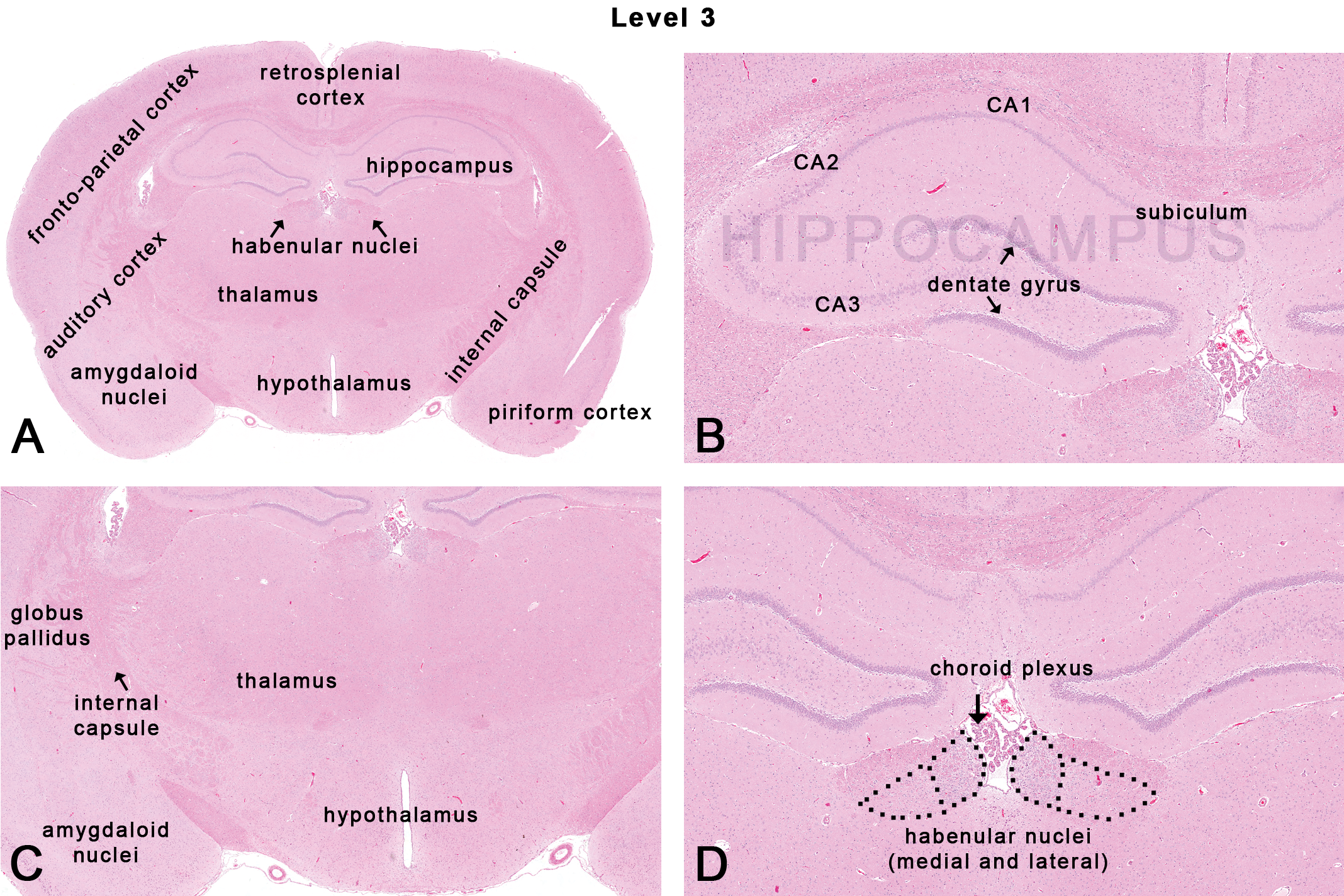
This section is taken through the infundibulum and/or the median eminence (Figures 1, 2C, and 5). Neuroanatomical subsites include the hippocampus, the thalamus, the hypothalamus, the habenular nuclei, and the amydaloid nuclei (Figure 5). The cortex at this level includes a continuation of the frontoparietal cortex, the auditory (temporal) cortex, the piriform cortex (Figure 4A), and the retrosplenial cortex (Figure 5A).
The hippocampus is one of the most studied areas of the brain by neuroscientists, especially in the field investigating epileptogenesis, learning, and memory processing. Neuroanatomically, the hippocampus is composed of the cornu ammonis (CA) and the dentate gyrus. The dentate gyrus receives afferent input from the entorhinal cortex (Figure 6A) and sends efferents to the hippocampal pyramidal cells. The CA is subdivided into regions CA1, CA2, and CA3. In rodents, a fourth region located in the hilar area facing the dentate gyrus is considered as the CA4 or the ‘end blade’ of the hippocampus by some neuroscientists. The CA region of the hippocampus receives afferent inputs from the dentate gyrus and the septal nuclei and sends efferent fibers to the subiculum and septal nuclei. The subiculum (Figure 5B) is a transitional area between the hippocampus and entorhinal cortex (Figure 6A) and is the main output of the hippocampus. Functionally, the hippocampus is involved in short-term memory function and is one of the first areas to show histopathologic changes in Alzheimer’s patients (Van Hoesen and Hyman 1990). The hippocampus is also relevant in studies of neuronal plasticity, which has been documented in the subgranular zone of the dentate gyrus, suggesting that the hippocampus is a site for adult neurogenesis (Cosgrave et al. 2010; Ming and Song 2005). The CA1 region, also known as Sommer’s sector, is particularly vulnerable to hypoxic or ischemic damage (Somjen et al. 1990), while the CA2 region is relatively resistant. More recently, the CA2 region has been implicated in schizophrenia (Jones and McHugh 2011; Narr et al. 2004).
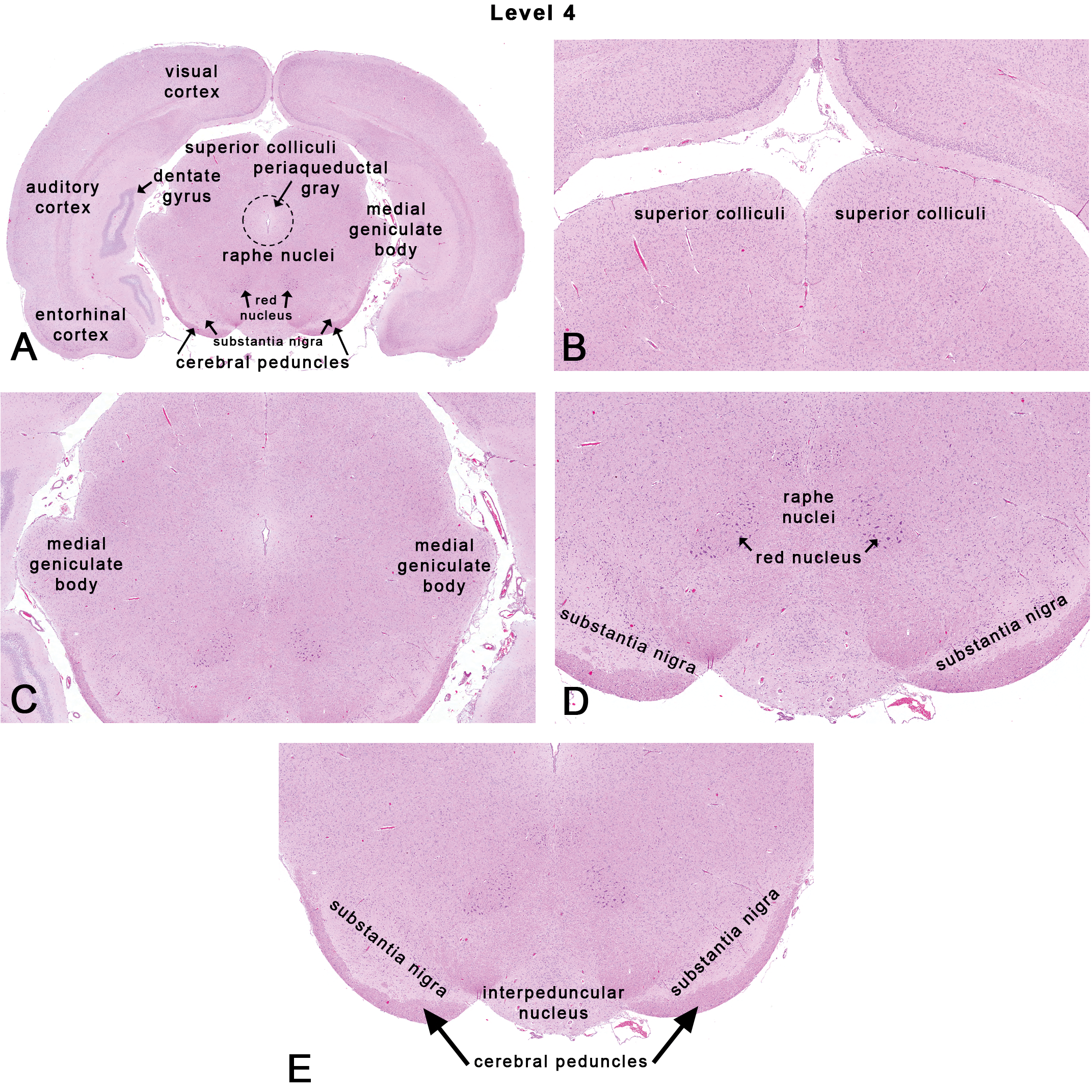
Rat: H&E. Transverse section at NTP-7, level 4 depicting subsites on a section through the superior colliculus. Dotted outlines are close approximations and an atlas should be consulted for accuracy.
Classically, the hippocampus is associated with epilepsy in humans, where “ammonal horn sclerosis” (AHS) was first described in 1880 by Sommer (Thom 2009). In the 1890s, AHS (reflected as neuronal loss and gliosis in the CA1 and CA4/end blade regions) was associated with temporal lobe epilepsy and psychomotor seizures in humans. In toxicologic pathology, examples of chemical-induced lesions in the hippocampus include trimethyltin (McPherson et al. 2003) and benzyl acetate that specifically affects the granule cell layer in the dentate gyrus (Abdo et al. 1998).
The thalamus (Figure 5C) is often referred to as the gateway to the cortex and contains multiple nuclei that relay sensory (excluding olfaction, vision, and auditory) and motor signaling to the cortex from the spinal cord, brain stem, cerebellum, and basal nuclei. The ventral caudal thalamic nuclei (both lateral and medial) play key roles in somesthesis, relaying touch and proprioceptive information from the dorsal columns of the spinal cord and trigeminal nuclei to the primary somatosensory cortex (Ohno et al. 2011). Lesions in the anterior thalamic nuclei are subsequently associated with lesions in the limbic circuit (Dumont et al. 2012) suggesting the retrograde effect and obvious interconnection between the 2 regions. In addition, the thalamus has connections with the cerebellum for controlling motor functions (Aumann et al. 1994).
Major sensory thalamic nuclei include the medial and lateral geniculate nuclei that form the metathalamus. The medial geniculate nuclei receive input from the inferior colliculi and send outputs to the primary auditory (temporal) cortex (Malmierca 2003). The lateral geniculate nuclei receive input from the retina and relay information to and from the primary visual (occipital) cortex (Woodward et al. 1990).
The hypothalamus comprises a large group of ill-defined nuclei that lie ventral to the thalamus (along the base and walls of the ventral portion of the third ventricle, contiguous with the pituitary gland; Figure 5C). The pituitary gland is profusely innervated by neurons from the hypothalamus (hypothalamic–pituitary axis). Functionally, the hypothalamus is involved in hormonal and behavioral circadian rhythms, as well as vital homeostatic mechanisms such as cardiovascular tone, eating, drinking, sleeping, thermoregulation, sexual, and emotional behavior (Thompson 1967).
The habenular nuclear complex is part of the epithalamus, located dorsally, adjacent to the dorsal portion of the third ventricle, and rostral to the pineal gland (Figure 5D). In the rat, bilaterally, the habenular complex consists of a medial and a lateral habenular nucleus. Functionally, the habenular nuclei are involved in pain processing, reproductive behavior, nutrition, sleep–wake cycles, stress responses, and learning activities (Andres, von During, and Veh 1999). There is also evidence to suggest that the lateral habenular nuclei could play a role in reward recognition (Matsumoto and Hikosaka 2007).
The amygdaloid nuclei are part of the limbic system and are composed of at least 12 nuclei located ventrally adjacent to the piriform lobe (Figure 5C). The amygdaloid nuclei are uniquely connected with other areas of the brain including the olfactory bulb and the cortex, the hypothalamus, the thalamic reticular nucleus, the trigeminal nucleus, the facial nerve nucleus, the ventral tegmental area (VTA), the locus coeruleus, the hippocampus, and the laterodorsal tegmental nucleus. Functions include processing and memory of emotional reactions (Pitkanen, Jolkkonen, and Kemppainen 2000) and playing an important role in fear (Rosen and Donley 2006), anxiety, and depression.
Occasionally, at this NTP-7 level 3, the globus pallidus and internal capsule may also be seen (Figure 5C). The globus pallidus is a major component of the basal nuclei along with the striatum. The globus pallidus is traversed by numerous myelinated axons of the striato-pallidonigral bundle giving it a pale appearance for which it is named. The internal capsule is composed of ascending and descending axons that separate the caudate nuclei and thalamus from the putamen and the globus pallidus except in the rat where the caudate and putamen are indistinguishable and referred to as the caudate–putamen. The axons of the internal capsule connect the cerebral cortex to the pyramidal tracts of the medulla oblongata.
NTP-7, Level 4
This section is taken at the superior colliculus (also known as the rostral colliculus or most commonly, the anterior colliculus) as shown in Figures 1, 2D, and 6. On the ventral surface of the brain, this landmark corresponds to the caudal half of the cerebral peduncles (also known as crus cerebri) and the interpeduncular nucleus that are grossly visible on the ventral surface of the brain (Figures 1 and 6E). Neuroanatomical subsites in this section include the superior colliculus, the medial geniculate body (MGB), the red nucleus, the substantia nigra, the cerebral peduncle, and often one of the raphe nuclei (Figure 6A) of the reticular formation. Caudal parts of the hippocampus (particularly CA3 and dentate formation) are often present with the cerebrum. The cortical regions at this level often include the visual (occipital), the auditory (temporal), and the medial and lateral portions of the entorhinal cortices.
The superior colliculus or optic tectum is a paired structure comprising the dorsal part of the midbrain (Figure 6B). The optic tectum (dorsal part of the mesencephalon) is one of the largest brain structures in fish and birds that play an important role in directing visual attention and controlling eye movements. Experiments have demonstrated that the superior colliculus is involved in the spatial localization of a stimulus rather than its recognition (Goodale, Foreman, and Milner 1978; Heywood and Cowey 1987; Schneider 1969). Other studies have shown that lesions in the superior colliculus in the rat produce postural defects, contralateral sensory neglect, ipsilateral hypersensitivity (Krauthamer, Krol, and Grunwerg 1992), and deficits in problem-solving skills (Smith and Weldon 1976). More recently, in humans, the superior colliculus has been identified as a potential locus for attention deficit hyperactivity disorder (Overton 2008).
The medial geniculate nucleus or MGB lies on the posterolateral aspect of the thalamus as a rounded eminence (Figure 6A and C). The MGB is part of the auditory pathway (Malmierca 2003) before inputs reach the inferior colliculus. In rats, anatomically, there is a dorsal, a ventral, and a medial nucleus within the MGB. However, functional significance differentiating each nucleus remains unknown. It is speculated that the MGB influences the direction and maintenance of attention in response to auditory stimuli (Malmierca 2003).
The red nucleus (Figure 6D), as the name suggests, refers to the cluster of pinkish colored cells, which appears as a large rounded red structure in fresh slices of the human mesencephalic tegmentum (ventral part of the mesencephalon). The red nucleus is a conspicuous nucleus in all limb-using vertebrates and serves as a premotor center (ten Donkelaar 1988) receiving input from cerebellar nuclei and acting as a source of information for various precerebellar nuclei. The red nucleus gives rise to the rubrospinal and rubro-olivary tracts that also carry other motor fibers such as those of the cerebral peduncle. Studies involving induction of lesions in the red nucleus showed that injury to the fibers cause functional motor impairments (Whishaw, Pellis, and Pellis 1992).
The raphe nuclei contain neurons that provide the major serotonergic projections to the forebrain (Gaspar and Lillesaar 2012). Anatomically, the raphe nuclei are a cluster of nuclei in the midbrain (Figure 6A and D), the pons, and the medulla oblongata. The rostral raphe nuclei (midbrain and pons) are a source of divergent fibers toward the more rostral areas of the brain, while the caudal raphe nuclei (medulla oblongata) project mainly to the caudal brain stem and spinal cord. Functionally, given the increase of antidepressants such as selective serotonin receptor inhibitors into the pharmaceutical market (Cascade and Kalali 2008), the raphe nuclei are relevant sites for potential effects of toxicity (Gardier et al. 1996). Since serotonin is implicated in a wide array of physiological states such as sleep/wakefulness (Monti 2010), appetite, and anxiety, the raphe nuclei have also become targets for drugs affecting food intake (Garattini et al. 1988).
The substantia nigra (Figure 6A, D, and E) and adjacent ventral tegmental region is another widely studied area among neuroscientists. The neurons in this region are predominantly dopaminergic, a prime subanatomic area affected in neurodegenerative diseases such as Parkinson’s and Huntington’s diseases (Gibb 1991). Moreover, the VTA plays a key role in the reward and addiction behavior (Beitner-Johnson, Guitart, and Nestler 1992).
Ventrolateral to the substantia nigra are prominent white matter motor tracts called the cerebral peduncles (Figure 6A and E) that include but are not limited to the corticospinal and corticobulbar tracts. The fibers in the cerebral peduncles run from the base of the cortex along the internal capsule through the longitudinal fibers of the pons and the pyramidal tracts and carry information about movement.
At the ventral aspect of the midline, between the cerebral peduncles lies the interpeduncular nucleus that is thought to have broad inhibitory effects on many other brain regions (Nishikawa, Fage, and Scatton 1986) and is linked with decreased dopamine release and utilization from dopamine synaptic terminals.
NTP-7, Level 5
This section is taken at the inferior colliculus (also known as the caudal colliculus or more commonly, the posterior colliculus) as shown in Figures 1, 2E, and 7. On the ventral surface of the brain, this corresponds to the rostral margin of the trapezoid body. Neuroanatomical subsites at this level include the inferior colliculus, the superior olivary nucleus (also known as the dorsal nucleus of the trapezoid body), the locus coeruleus, the mesencephalic trigeminal nucleus, the spinal trigeminal tract, principal sensory nucleus of cranial nerve (CN) V, motor trigeminal nucleus, and the pyramidal tracts (Figure 7A).
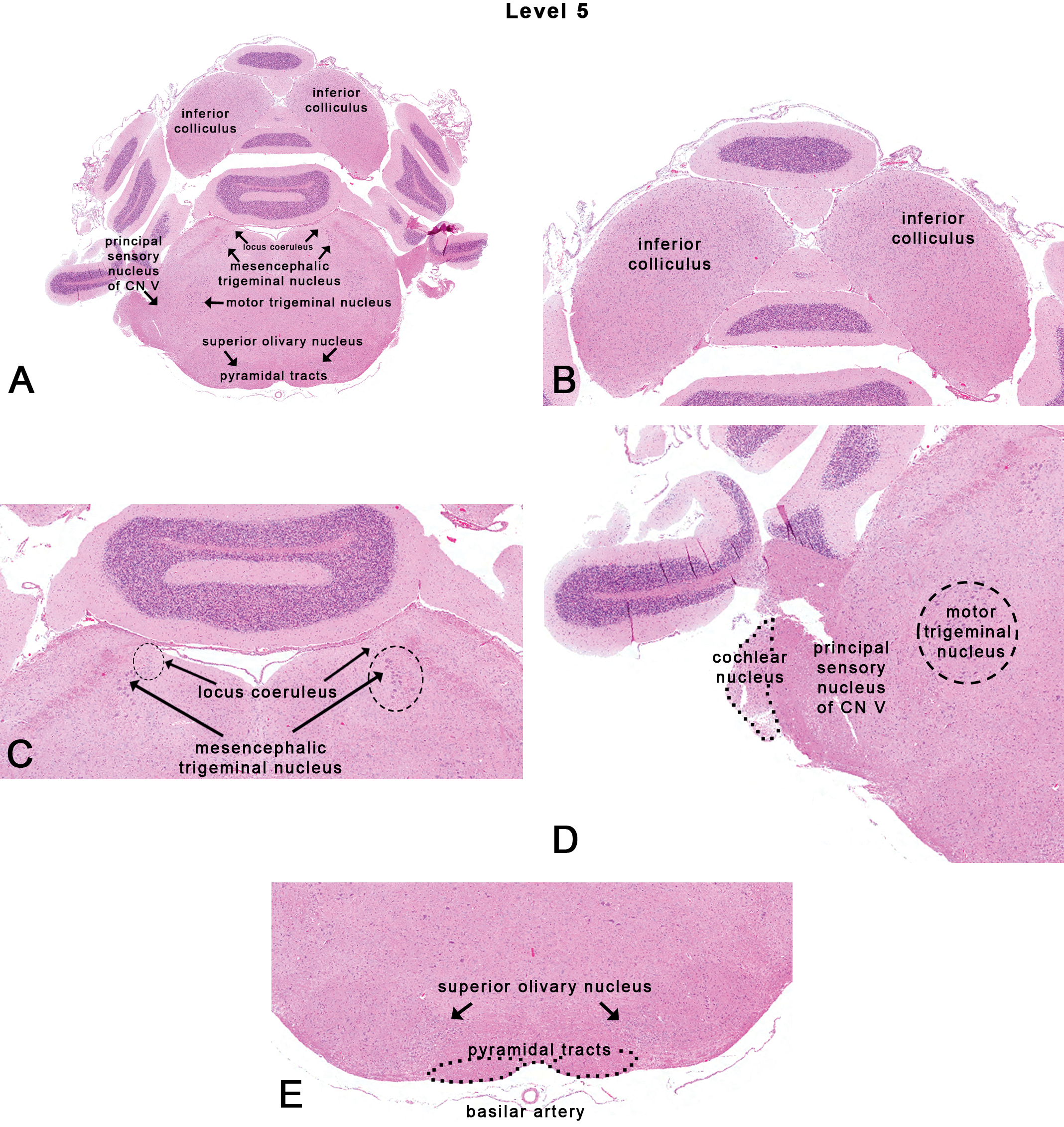
Rat: H&E. Transverse section at NTP-7, level 5 depicting subsites on a section through the inferior colliculus. Dotted outlines are close approximations and an atlas should be consulted for accuracy.
The inferior colliculus (Figure 7B) is part of the central auditory pathway (Monti 2010). Morphologically, ubiquitous laminae are seen, which may be the basis for tonotopic organization. Unilateral lesions in the inferior colliculus produce deficits in sound localization confined to the contralateral space (Jenkins and Masterton 1982). The inferior colliculus is also a site with large microvascular blood volume (Cremer, Cunningham, and Seville 1983), high rate of blood flow (Sakurada et al. 1978), and glucose metabolism (Sokoloff 1981) that makes it especially vulnerable to toxicants. Studies with carbonyl sulfide have shown that the inferior colliculus is a prominent site for neurotoxicity (Morgan et al. 2004).
Associated with the auditory pathway at this level is the superior olivary nucleus (Figure 7E) containing medial and lateral foci that receive fibers from the cochlear nuclei and contribute fibers to the lateral lemniscus. Functionally, the superior olivary nucleus is associated with processing of binaural cues for sound localization (Inbody and Feng 1981).
The locus coeruleus, which literally means “blue spot” (due to the grossly visible presence of neuromelanin in human sections), is located medial to the mesencephalic trigeminal nucleus (Figure 7C). It is the principal site for the synthesis of norepinephrine (in addition to the adrenal medulla). The locus coeruleus is not only involved in physiological responses to stress and panic but also plays a role in sleep (Jouvet 1969), attention (Robbins 1984), and memory (Sara and Devauges 1988). Interestingly, the locus coeruleus is also affected in many forms of neurodegenerative diseases. Mouse models of Alzheimer’s disease indicate accelerated neuropathologic progression after chemical destruction of the locus coeruleus (Heneka et al. 2006).
The trigeminal complex of CN V includes sensory and motor components. The motor component mainly includes a motor trigeminal nucleus (Figure 7D) associated with masticatory reflexes and control of jaw movements. The sensory nuclei include the mesencephalic trigeminal nucleus (Figure 7C), principal (or pontine) sensory nucleus of CN V (Figure 7D), and the spinal trigeminal nucleus. Neuroanatomically, the mesencephalic trigeminal nucleus is located at the lateral margins of the periaqueductal gray (surrounding the third ventricle) at the level of the superior colliculus. In deeper sections, caudal to the inferior colliculi, the mesencephalic trigeminal nucleus is located further laterally and is bordered medially by the locus coeruleus. Neurons of the mesencephalic trigeminal nucleus are cell bodies of primary afferent fibers concerned with muscles of mastication and periodontal receptors of both maxillary and mandibular teeth (Rokx, Juch, and van Willigen 1986). The principal sensory nucleus of CN V is a peanut-shaped structure that lies lateral to the pons and is visible in transverse sections (Figure 7D). The spinal trigeminal tract (Figures 7D and 8C) lies adjacent and lateral to the principal sensory nucleus of CN V and carries fibers from the ophthalmic, maxillary, and mandibular afferents. The principal sensory nucleus of CN V contains neurons that receive sensory input from the maxillary, mandibular, and ophthalmic nerve innervation areas of the skin with primary projections into the contralateral thalamus. The spinal trigeminal nucleus provides projections to the facial nucleus and the cerebellum.
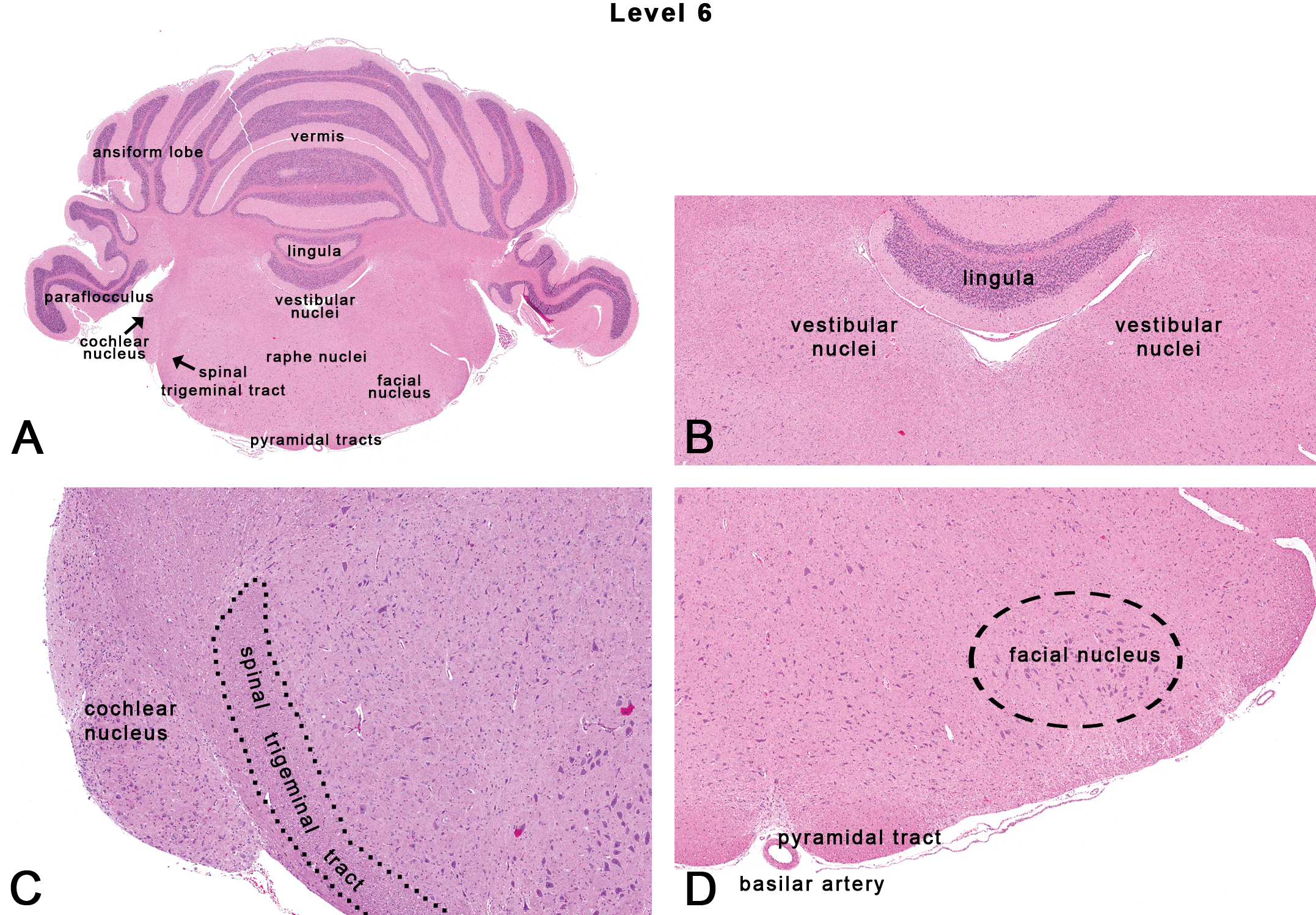
Rat: H&E. Transverse section at level 6 depicting subsites on a section through the mid-cerebellum. Dotted outlines are close approximations and an atlas should be consulted for accuracy.
The pyramids (Figure 7A and E) are paired motor tracts that extend along the length of the medulla oblongata and are bordered medially by the ventral median fissure. The pyramidal motor tracts contain fibers that comprise the corticobulbar and corticospinal tracts carrying information regarding voluntary motor activity and are continuations of the internal capsule and cerebral peduncle/longitudinal fibers of the pons. In comparison, the extrapyramidal motor tracts, primarily those originating in the basal nuclei and rubrospinal tracts, are those that do not follow the medullary pyramids and are involved with involuntary motor activity.
NTP-7, Level 6
This section is taken at the level of the mid-cerebellum corresponding to the location of CN VIII (Figures 1, 2F, and 8). Neuroanatomic subsites at this level include various parts of the cerebellum (such as the vermis, ansiform lobe, paraflocculus, lingula or the 1st cerebellar lobule), the vestibular nuclei, the cochlear nucleus, the facial nucleus, and the spinal trigeminal tract (Figures 8A, 8C, and 9A).
The cerebellum is phylogenetically an old structure overlying the pons. Histologically, the cerebellar cortex contains three layers: the superficial pale molecular layer (O’Halloran, Kinsella, and Storey 2011), the middle Purkinje cell layer, and the deep granular cell layer (containing granule cells). Functionally, the cerebellum receives most of its input from extracerebellar sources including the vestibular nerve, as well as optic and auditory inputs (Glickstein 1997). The primary output (via the Purkinje neurons) is to the deep cerebellar roof nuclei. Although the cerebellum has traditionally been considered as a center for motor control and coordination, more recently the cerebellum is also recognized as a site for cognitive functions (O’Halloran, Kinsella, and Storey 2011). For example, attention-deficit hyperactivity disorders in humans have been associated with changes in the cortex, basal nuclei, and cerebellum (Curatolo, D’Agati, and Moavero 2010). Changes in proportions of granule cells in the rat cerebellum have also been associated with hyperactivity behaviors (Altman 1987). Therefore, as research elucidates other cognitive pathways, it is important to be aware of lesions within the cerebellum even in the absence of motor symptoms.
The vestibular nuclei (Figure 8B) contain 4 major nuclei and a number of smaller groups collectively known as the vestibular nuclear complex. Functionally, the vestibular nuclei are important for the maintenance of balance and equilibrium (Rubertone, Mehler, and Voogd 1995).
The cochlear nucleus (Figures 7D and 8C) is the first relay station along the central auditory pathway and is located at the dorsolateral border of the junction of the pons and medulla oblongata. At the cochlear nucleus, signals of the cochlear nerve diverge into a number of parallel ascending tracts, each with its own particular course and destination as well as conduction velocities and relays, along the central auditory pathway (Malmierca 2003). In general, most ototoxicants affect the peripheral auditory system (cochlea of the inner ear). However, since the cochlea is rarely evaluated in routine screening toxicity studies, lesions noted in the central auditory pathway warrant further investigation to delineate toxicological effects (Morgan et al. 2004).
The facial nucleus (Figure 8D) is composed of motor neurons that control the superficial muscles of the head and neck (Moller and Sen 1990). Occasionally, a slightly rostral representation of this section will reveal the genu of the facial nerve, which is a curve that the fibers of the root of the facial nerve follows around the abducens nuclei (of CN VI) in the pontine tegmentum.
The spinal trigeminal tract (Figure 8C) lies adjacent and lateral to the nucleus of the spinal trigeminal tract. The spinal trigeminal tract carries afferents from the ophthalmic, maxillary, and mandibular regions.
NTP-7, Level 7
This section was taken through the caudal medulla oblongata, approximately 2 to 3 mm rostral to the caudal termination of the cerebellum as seen from the ventral surface of the brain (Figures 1, 2G, and 9). Neuroanatomical subsites include the area postrema, the vagal nucleus or the dorsal motor nucleus of vagus, the hypoglossal nucleus, the reticular formation, raphe nuclei, and the inferior olivary nucleus (Figure 9).
The area postrema is 1 of the 7 circumventricular organs (Garman 2011). In the rat, the area postrema is a quadrant of tissue located in the fourth ventricle, dorsal to the nucleus of the solitary tract (Figure 9B). The area postrema is richly vascular and is comprised of fenestrated endothelial cells (i.e., it lacks a BBB). The relevance of this site in routine toxicity screening studies lies in identifying lesions that may result from large polar molecules that circumvent the BBB to enter the brain. Neurons in the area postrema participate in baroreflexes and cardiovascular control (Papas and Ferguson 1991; Skoog and Mangiapane 1988). The area postrema has also been implicated as a chemoreceptor trigger zone for emesis. Lesions in the area postrema prevent vomiting following exposure to most emetic drugs (Miller and Leslie 1994).
The vagal nucleus or dorsal motor nucleus of vagus and solitary tract nucleus are located in the dorsomedial portion of the medulla oblongata ventral to the area postrema (Figure 7B). The vagal nucleus is the source of general visceral efferent (parasympathetic autonomic) fibers in vagus CN X. The solitary tract nucleus is associated with visceral sensation and taste from the facial (CN VII), glossopharyngeal (CN IX), and vagus (CN X) CNs.
The hypoglossal nucleus is located ventral to the area postrema (Figure 9A and B). This nucleus contains mainly motor neurons that innervate the lingual muscles.
Historically, the reticular formation (Figure 9C) was considered a radiating network of fibers intermingled with dispersed neurons and glial cells that extend through the medulla oblongata, pons, and mesencephalon of the brain stem. Although investigators have described reticular nuclei, functional maps have often demonstrated indistinct cytoarchitectural boundaries (Andrezik and Beitz 1985). In general, the reticular formation may be considered as a network for massive integration of polymodal input and output, representing “the coordinating apparatus” for sensory-motor functions. In the rat, afferent connections of the reticular formation suggest involvement of pathways centered in the sleep–waking cycle (Shammah-Lagnado et al. 1983). Destruction of the midbrain reticular formation in cats results in a sleeping or stuporous state (Lindsley, Bowden, and Magoun 1949).
The inferior olivary nucleus is located ventromedially in the caudal part of the medulla oblongata, just dorsolateral to the pyramidal tracts (Figure 9D). The inferior olivary nucleus receives afferent connections from many different sources in the brain stem and spinal cord, while efferent fibers from the inferior olivary nucleus are directed to the cerebellum and are associated with motor functions.
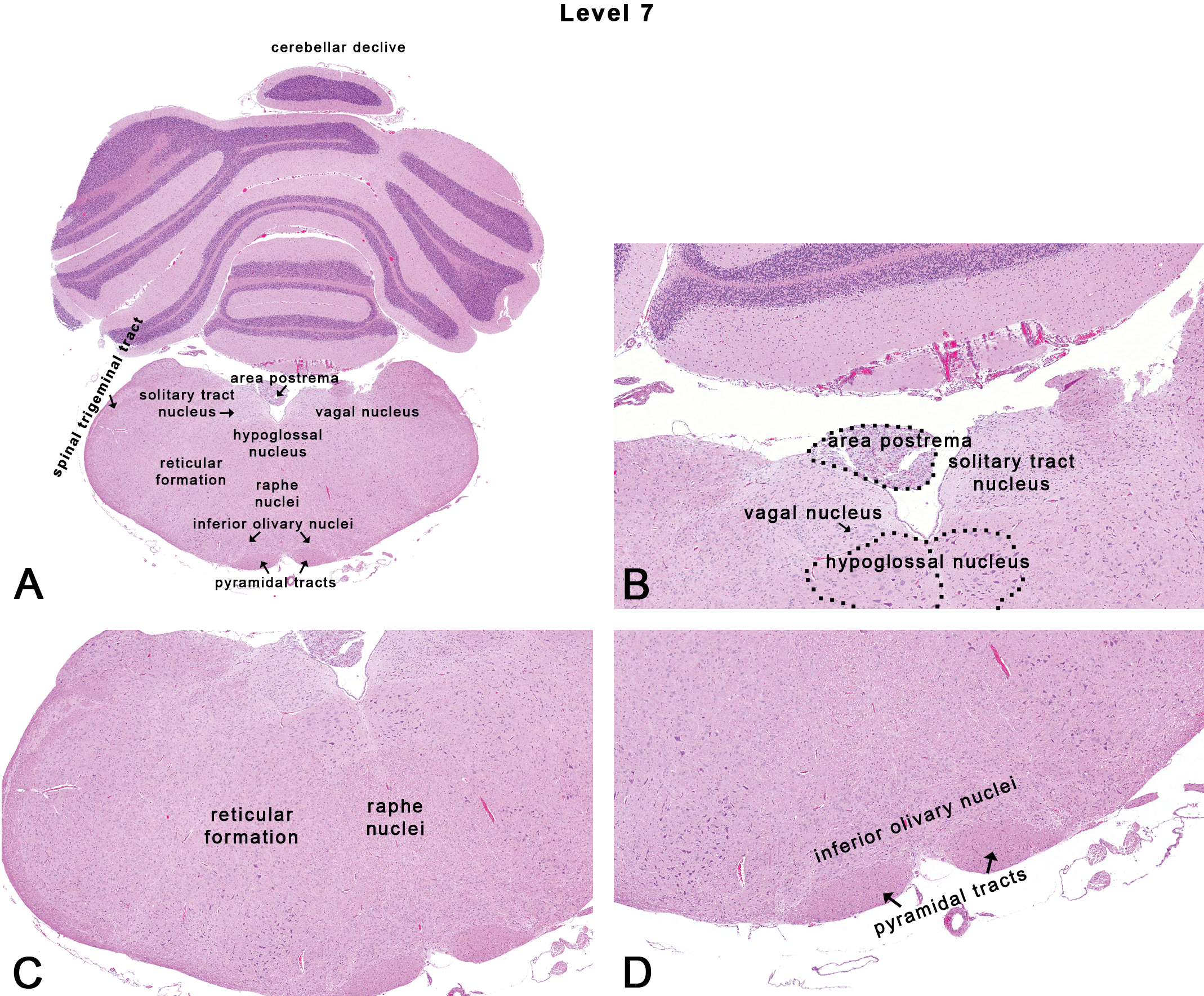
Rat: H&E. Transverse section at level 7 depicting subsites on a section through the caudal medulla oblongata near the caudal termination of the cerebellum. Dotted outlines are close approximations and an atlas should be consulted for accuracy.
Spinal Cord
The neuroanatomy of the spinal cord predominantly consists of a central, butterfly- or H-shaped zone of gray matter and an outer rim of white matter. The gray matter is divided into the dorsal horn and the ventral horn, which are separated by the intermediate gray matter. Neurons in the dorsal horn receive incoming or afferent sensory signals from dorsal spinal radicular nerves, whereas the large neurons of the ventral horn distribute outgoing or efferent motor impulses to various effector organs by ventral spinal radicular nerves. The white matter contains several symmetrical ascending tracts (with the prefix “spino”) and descending tracts (with the suffix “spinal”) that are divided into the dorsal, lateral, and ventral columns (funiculi). Major ascending pathways include the spinothalamic and spinocerebellar tracts. Major descending motor tracts include the corticospinal and rubrospinal tracts. Unlike in humans where the lateral corticospinal tract plays a major role in motor control, mice depend mainly on the rubrospinal tract for their motor functions (Hagan, Bolon, and Keene 2012). Rostral regions of ascending tracts such as the dorsospinocerebellar, gracile, and cuneate, as well as the caudal end of descending tracts such as the lateral columns, ventrolateral, and ventromedial tracts are especially vulnerable to many toxicants such as n-hexane, methyl n-butyl ketone, 2,5-hexanedione, acrylamide (Spencer and Schaumburg 1977). Examples of agents showing preferential susceptibility to different parts of the spinal cord include carbon disulfide, dibromoacetic acid, and aniline. Carbon disulfide was shown to affect the fasciculus gracilis in the cervical spinal cord, and lateral and ventral funiculus nerve tracts in the lumbar spinal cord (Sills et al. 1998); aniline most severely affected the lateral and ventral columns of the thoracic spinal cord in rats (Okazaki et al. 2001); and lesions with dibromoacetic acid were more severe in the lumbar region compared with the cervical region (Moser et al. 2004).
Typically, three transverse sections of the spinal cord are taken in NTP studies (Figure 10). These include the first cervical spinal cord level, a mid-thoracic spinal cord level, and the mid-lumbar spinal cord level. Sections of the first cervical spinal cord often contain the gracile and cuneate nuclei located dorsally which are involved in proprioception from most parts of the body; the first cervical spinal cord sections also harbor the majority of the white matter motor tracts. The lumbar spinal cord sections taken through the lumbar intumescence contain cell bodies of motor nerves supplying hind limbs. In addition, some sections of the spinal cord may often contain partial areas of the dorsal and ventral nerve roots (Figure 10C), and occasionally, dorsal root (spinal) ganglia containing cell bodies of sensory nerve fibers. The cell bodies of the somatic nervous system are located in the ventral regions of the gray matter, while the cell bodies of the autonomic nervous system are located in the lateral regions of the gray matter of certain areas of the spinal cord in the thoracolumbar region. For identification of specific tracts within different levels of the spinal cord, rat and mouse spinal cord atlases are available online: http://www.christopherreeve.org/site/c.ddJFKRNoFiG/b.4819993/k.4888/Spinal_Cord_Atlas__Rat_Spinal_Cord.htm (accessed August 10, 2012); http://www.christopherreeve.org/site/c.ddJFKRNoFiG/b.4427053/k.F400/Spinal_Cord_Atlas__Introduction_and_Mouse_Spinal_Cord.htm (accessed August 10, 2012).
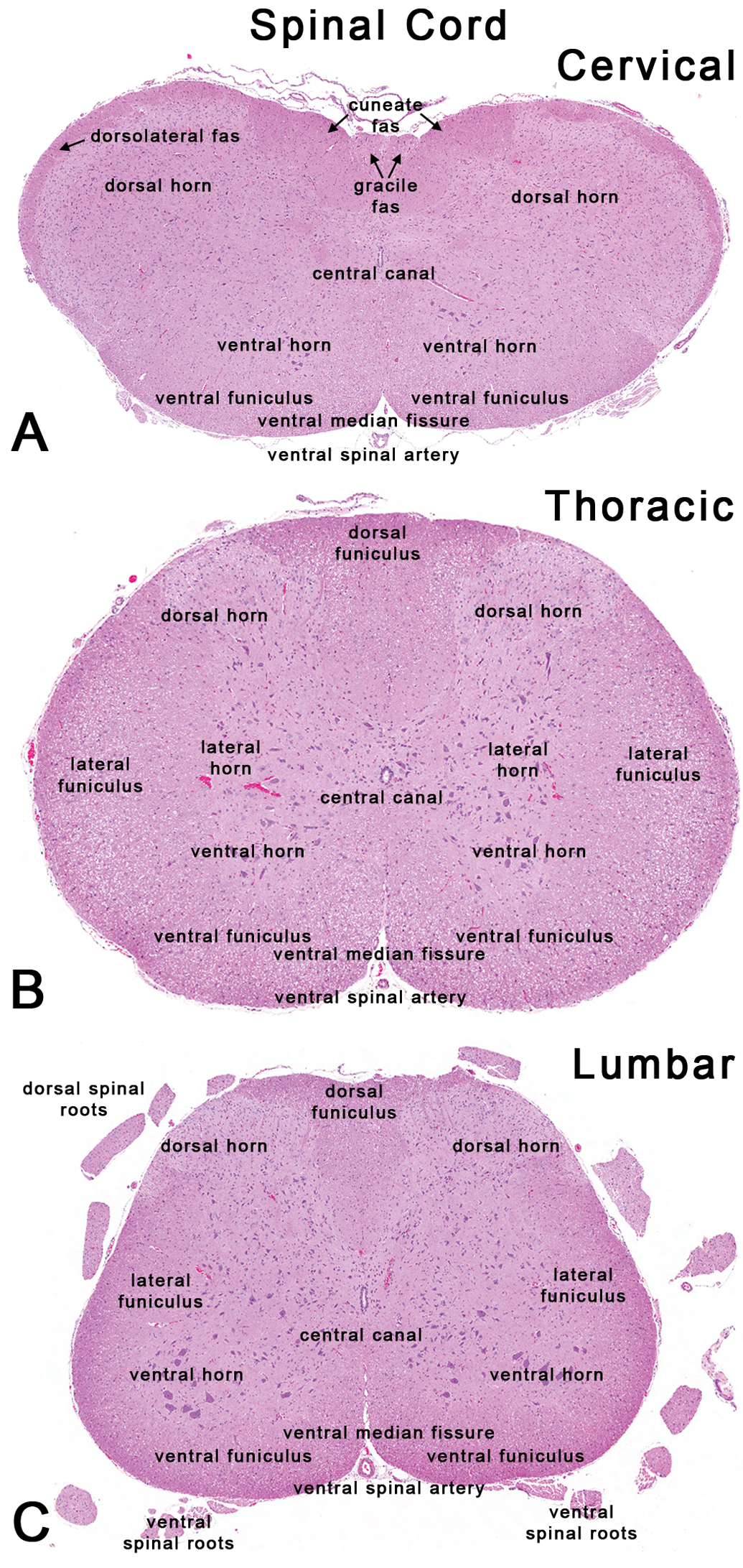
Rat: H&E. Transverse sections of the spinal cord through the first cervical, mid-thoracic, and mid-lumbar levels. cun fas = cuneate fasciculus; dorsolateral fas = dorsolateral fasciculus; gr fas = gracile fasciculus.
Peripheral Nerves
Transverse or cross (Figure 11A) and longitudinal sections (Figure 11B) of the sciatic and/or tibial nerves are usually evaluated as representative samples of the peripheral nervous system in NTP studies. Typically, these are mixed nerves containing both sensory (afferent) fibers and motor (efferent) fibers. Classic examples of peripheral neuropathy due to toxicants include acrylamide (Smith and Oehme 1991), hexacarbons (Spencer and Schaumburg 1977); (Couri and Milks 1982), organophosphates (Jortner et al. 2005), iminodipropionitrile (Carden et al. 1987), and carbon disulfide (Gottfried et al. 1985; Sills et al. 1998). Other examples include drugs withdrawn from the market due to peripheral neuropathy. These include an antimicrobial agent (clioquinol), a diuretic (muzolimine), and a vasodilator drug (perhexiline; Myers and Ronthal 1978; Pohlmann-Eden et al. 1991; Wadia 1984). More recently, emerging toxic neuropathies such as those seen with cancer chemotherapeutics, immunomodulator drugs (tumor necrosis factor-α antagonists), and aerosolized porcine neural tissue in swine abattoirs (Kushlaf 2011) further underscore the importance of including peripheral nerves during toxicity evaluation studies.
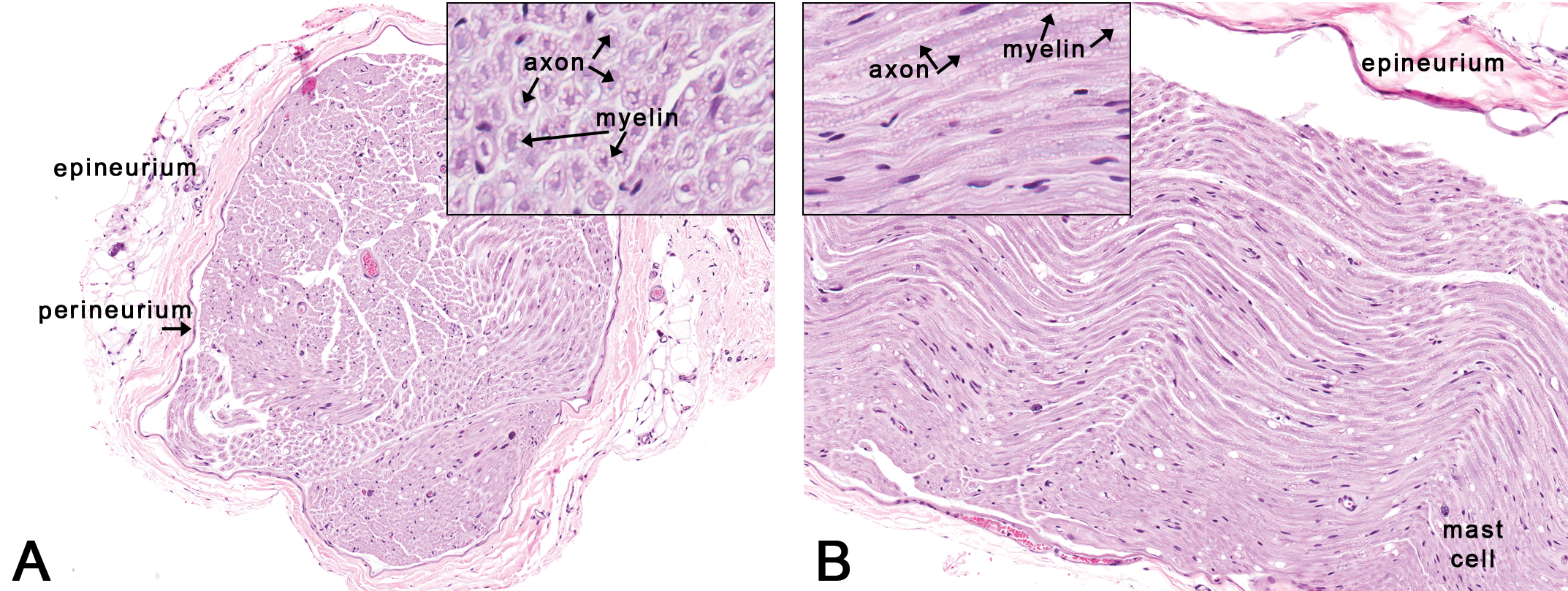
Rat: H&E. Sections through the sciatic nerve. Figure 11A is a transverse section through the sciatic nerve depicting the outer connective tissue layers (epineurium and perineurium) around a nerve fiber bundle. Inset reveals individual nerve fibers in transverse section showing central axons and surrounding myelin. Figure 11B is a longitudinal section through the sciatic nerve showing myelin around the axon. A few scattered mast cells are not uncommon in the peripheral nerves in rats.
Trigeminal Nerve and Ganglion
Where appropriate, longitudinal and transverse or cross sections of the trigeminal nerve and ganglion are evaluated. The trigeminal nerve (CN V) is the largest of the CNs that contain both sensory and motor components. The trigeminal nerve leaves the cranium and passes through the trigeminal canal in the rostral part of the petrous portion of the temporal bone. Here, the trigeminal ganglion rests in the cavum trigeminale of the dura mater (Figure 12A). A stereotaxic atlas of the trigeminal ganglion has been documented by Schneider et al. (1981). The trigeminal ganglion contains sensory nerve cell bodies (Figure 12B and C) of the general somatic afferent axons of all three branches (ophthalmic, maxillary, and mandibular) of the trigeminal nerve. Evaluation of the trigeminal nerve and ganglion is relevant in neurodegenerative diseases such as amyotrophic lateral sclerosis (Sandyk 2006) where neuronal loss is investigated using a mouse model (Yang et al. 2011). Other toxicants affecting the trigeminal nerve and ganglion include pyridoxine (vitamin B6) neurotoxicity (Levine and Saltzman 2004), cadmium (Habeebu et al. 2001), and lithium toxicity (Levine, Saltzman, and Kumar 2004).
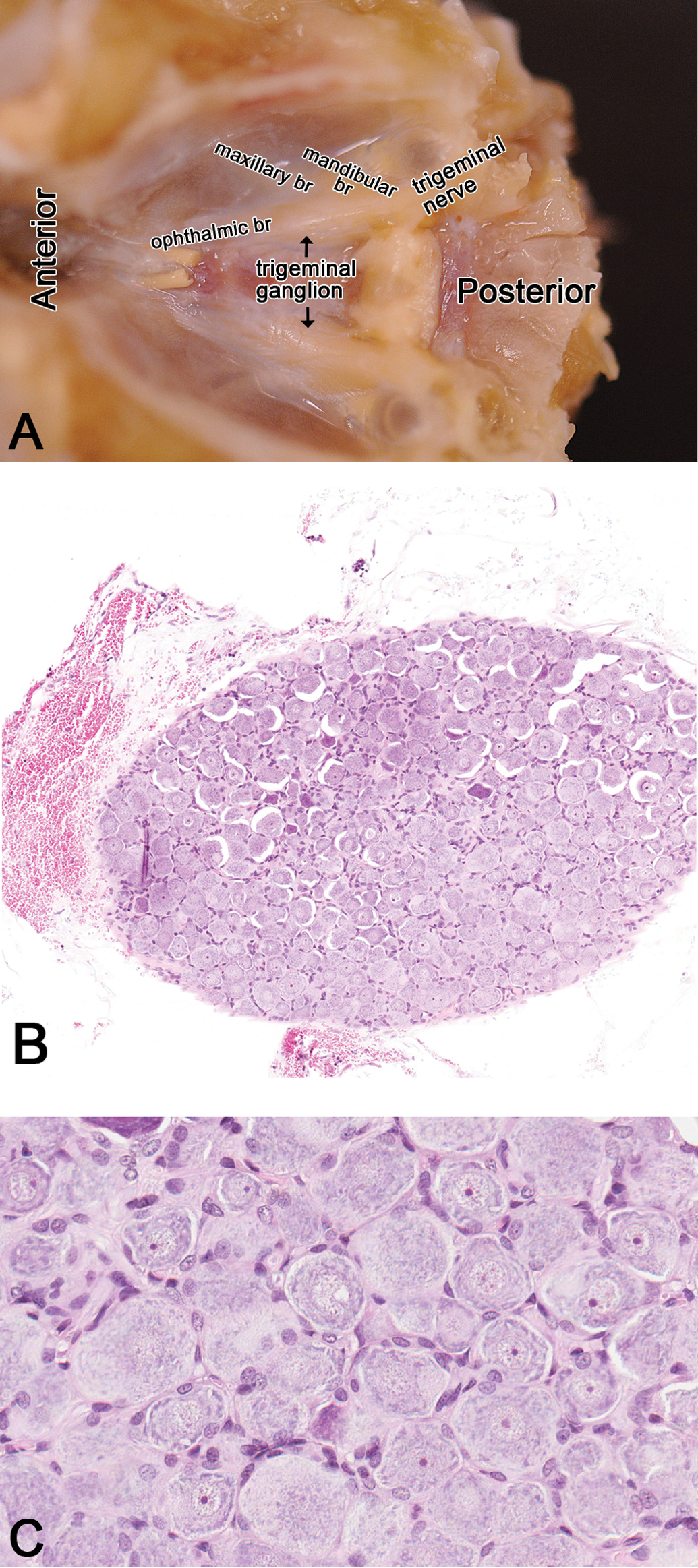
Rat: H&E. Trigeminal nerve and the ganglion. Figure 12A: dorsal view of the cranial cavity (brain removed) revealing the course of the trigeminal nerve and its branches (ophthalmic, maxillary, and mandibular branches) and its ganglion, rostrolateral to the pituitary gland. Figure 12B: a section of the trigeminal ganglion. Ganglion cells at higher magnification are shown in Figure 12C. Mandibular br = mandibular branch; maxillary br = maxillary branch; ophthalmic br = ophthalmic branch.
Autonomic Ganglia
Autonomic ganglia are involved in the regulation of involuntary visceral motor activity of the cardiovascular, respiratory, gastrointestinal, and genitourinary systems. In NTP studies, autonomic ganglia are evaluated from routine sections of the intestine. Enteric autonomic ganglia can be affected by excitotoxic mechanisms such as hypoxia or ischemia or excitotoxins in the food (Kirchgessner, Liu, and Alcantara 1997), immunosuppressants such as FK506 used in transplant surgeries (Higuchi et al. 2006), and drugs such as amiodarone used to treat cardiac arrhythmias (Costa-Jussa and Jacobs 1985). More recently, the role of intestinal autonomic ganglia in a mouse model of Parkinson’s disease is also being investigated (Kuo et al. 2010; Wakabayashi et al. 2010). In the intestine, the Meissner’s plexus is located in the submucosal layer, while the Auerbach’s or Myenteric plexus is located in the muscularis propria between the circular and longitudinal muscle layers (Figure 13). Although sections of the colon are probably most suited for assessment of the autonomic ganglia in rats and mice, autonomic ganglia are also consistently present in sections of the duodenum and rectum in an internal evaluation (unpublished internal evaluation).
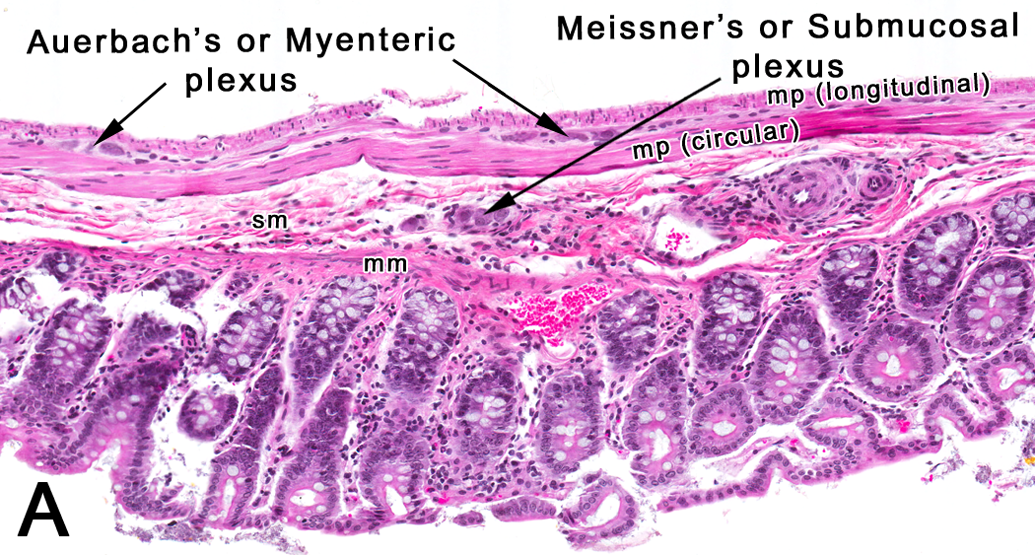
Rat: H&E. Colon. A transverse section of the colon depicting the location of intestinal autonomic ganglia. mm = muscularis mucosa; mp = muscularis propria; sm = submucosa.
Conclusion
The objective of this article is to provide an introductory and practical guide for pathologists to navigate a relatively small number of highly relevant neuroanatomical subsites within 7 sections of the brain. A brief functional relevance has been provided as a template for potential evaluation of neuronal circuits and pathways that may be involved in the pathogenesis of toxicant-induced injuries as well as neurodegenerative disorders. Minimal pathologic lesions confined to small anatomic subsites warrant a more detailed evaluation since it is likely to be amplified via expansive neuronal circuitry in the brain. In fact, it has clearly been demonstrated that neuropathology progresses along stereotypical anatomic trajectories (Tiedeken, Muha, and Ramsdell 2013) such as those seen with neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease (Braak et al. 2004), and amyotrophic lateral sclerosis (Kanouchi, Ohkubo, and Yokota 2012). Moreover, the clinical signs seen in animals with lesions at specific neuroanatomic sites generally parallel lesions and clinical signs observed in humans. For example, lesions in the ventral hypothalamus in rats are associated with aggressiveness and hyperphagia, while lesions at the same site are associated with rage and hyperphagia in humans (Reeves and Plum 1969). Similarly, rats with lesions in the nigrostriatal complex showed deficits in maze tests, while humans with lesions in the nigrostriatal complex demonstrated deficits in route-walking tests (Bowen, Hoehn, and Yahr 1972). Hence, an awareness of functional relevance of anatomical subsites and pathways are important for pathologists during histopathological evaluation to better characterize neurotoxicity. In addition, it is important for the pathologist to note during routine histopathological evaluation that chemically induced lesions at a specific neuroanatomic subsite often impact other neuroanatomic subsites that are often linked via existing neuronal circuitry or pathways (Dumont et al. 2012; Morgan et al. 2004).
In conclusion, this review of neuroanatomic subsites is brief and serves as a guide to navigate across 7 sections of the brain. It should be used in conjunction with excellent reference atlases that are available for both the rat and the mouse brains (Franklin and Paxinos 2008; Kruger, Saporta, and Swanson 1995; Paxinos and Watson 2009; Paxinos et al. 2008). The brain atlases are also available on CD-ROM that allows easy digital navigation of brain sections that are arranged stereotaxically and allow easy retrieval and viewing of any of the neuroanatomic subsites. For the spinal cord (Watson, Paxinos, and Kayalioglu 2009), rat and mouse atlases are available online from the Christopher and Dana Reeve Foundation at: http://www.christopherreeve.org/site/c.ddJFKRNoFiG/b.4819993/k.942A/Dougs_Rat_Page.htm and http://www.christopherreeve.org/site/c.ddJFKRNoFiG/b.4427053/k.F400/Spinal_Cord_Atlas__Introduction_and_Mouse_Spinal_Cord.htm.
Footnotes
Acknowledgments
The authors are grateful to the reviewers who took the time to provide comments on the article in-preparation. These include veterinary pathologists with expertise in neuropathology—Drs. Robert H. Garman (Consultants in Veterinary Pathology, Inc.), Brad Bolon (Ohio State University), and James P. Morrison (Charles River Laboratories, Pathology Associates); veterinary pathologists at the NTP—Drs. David E. Malarkey and Ronald A. Herbert, Dr. Gordon Flake (MD pathologist—NTP), Dr. Danielle Bury (MD Resident Pathologist—Northwestern Deanery, Manchester, UK), Dr. Thimmasettapp Thippeswamy (Veterinary Anatomist and Experimental Neurobiologist, Iowa State University), Dr. Lola Hudson (Veterinary Anatomist, North Carolina State University), and Dr. Kristen Ryan (Neurotoxicologist—NTP).
Authors’ Note
The contents of this article were presented by Dr. Rao at the NTP Neuropathology Symposium at NIEHS, Research Triangle Park, North Carolina in April 23–24, 2012. Drs. Little and Sills pioneered the compilation of the list of neuroanatomical subsites within the NTP-7 sections, which is available in a CD format at the NTP. Brain sections were obtained from NTP Contract Laboratories; and the images and figures involved exceptional attention to detail given by Ms. Eli Ney (NIEHS) and Mr. David Sabio of Experimental Pathology Laboratories, Inc.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported (in part) by the Intramural Research Program of the NIH, National Institute of Environmental Health Sciences.