
Abstract
Endocytosis is the primary mechanism by which nanoparticles are translocated over the alveolar epithelium. The purpose of this study was to elucidate the association between endocytosis and the translocation of nanoparticles at the air–blood barrier (ABB). Gold colloid particles (diameter, 20 nm) were intratracheally instilled into male ICR mice. Fifteen minutes after instillation, localized accumulation of agglomerated gold particles was observed in the cytoplasm of macrophages, on the surface of alveolar epithelial cells (AECs), and in alveoli. Electron microscopy revealed particles in the vesicles of macrophages, on the surface of AECs, and in caveolae-like vesicles in type 1 AECs. Immunohistochemistry demonstrated positive immunolabeling for caveolin-1 in the ABB of untreated lungs as well as lungs treated with gold particles. Double immunofluorescence and immunoelectron microscopy revealed the presence of caveolin-1 in AECs in the untreated lungs. These results suggest that instilled gold colloid particles are internalized into the alveolar epithelium at the ABB by caveolae-mediated endocytosis, which is regarded as a physiological function of AECs.
Keywords
Introduction
Nanoparticles, i.e., particles with a diameter of <100 nm, are generated as chemically and physically well-defined materials or occur naturally as a consequence of combustion processes. Inhalation of nanoparticles occurs on a regular basis due to air pollution and is associated with an increase in respiratory and cardiovascular morbidity and mortality. Manufactured nanoparticles may intentionally be inhaled as pharmaceuticals or may unintentionally be inhaled during production in the workplace. Hence, the interactions of nanoparticles with the respiratory tract are currently under intensive investigation (Mühlfeld, Gehr, and Rothen-Rutishauser 2008).
During the past decades, various aspects of the interaction between airborne particles and pulmonary structures have been investigated. Different mechanisms of translocation of nanoparticles over the alveolar epithelium have been discussed. Endocytosis has been shown to be the main mode of particle uptake by several authors (Furuyama et al. 2009; Kim et al. 2006; Rejman et al. 2004; Shukla et al. 2005; Singh et al. 2007; Stearns, Paulauskis, and Godleski 2001; Takenaka et al. 2006; Zhang et al. 2010). Inhaled (Takenaka et al. 2006) and instilled (Furuyama et al. 2009) gold nanoparticles were observed within membrane-bound vesicles in alveolar macrophages and alveolar type 1 cells of mice lungs. In in vitro studies, similar results were obtained for the uptake of gold nanoparticles by macrophage cells, with the particles localized in pinocytotic vesicles and lysosomes (Shukla et al. 2005), as well as for the uptake of TiO2 nanoparticles by A549 cells (adenocarcinomic human alveolar basal epithelial cells), where the nanoparticles were localized inside vesicular structures including multivesicular and lamellar bodies (Singh et al. 2007; Stearns, Paulauskis, and Godleski 2001).
Endocytosis falls into 2 broad categories: phagocytosis (the uptake of large particles) and pinocytosis (the uptake of fluid and solutes). Pinocytosis can be further classified into at least 4 basic mechanisms: macropinocytosis, clathrin-mediated endocytosis, caveolae-mediated endocytosis, and clathrin- and caveolae-independent endocytosis (Conner and Schmid 2003). However, there is little information on the types of endocytosis responsible for the translocation of nanoparticles at the air–blood barrier (ABB). We have previously reported that C60 fullerene nanoparticles were translocated at the ABB by caveolae-mediated endocytosis in mice lungs (Naota et al. 2009). In vitro caveolae-mediated endocytosis of polystyrene nanoparticles has also been reported (Xia et al. 2008). In this study, we focused on caveolae-mediated endocytosis of instilled gold colloid nanoparticles at the ABB. Caveolae are flask shaped invaginations of plasma membrane proposed to mediate the extensive transcellular shuttling of serum proteins (Conner and Schmid 2003). We used morphological approaches to investigate the distribution of gold colloid nanoparticles administered intratracheally to mice. Gold colloid nanoparticles are suitable for morphological analysis because the particles have a uniform size, are compact, spherical, chemically stable, and electron dense, and are easily detectable on transmission electron microscopy images. The purpose of this study was to elucidate the association between endocytosis and translocation of nanoparticles at the ABB.
Materials and Methods
Animals and Administration of Particles
A total of 30 male 5- or 6-week-old ICR mice were obtained from CLEA Japan Inc. (Tokyo, Japan). Animals were fed a CE-2 diet purchased from CLEA Japan, and water was given ad libitum. The mouse cages were placed in a conventional room where the temperature was maintained at around 25°C and humidity at 55 to 70%. All animal experiments were performed according to the Tottori University guidelines for animal welfare (http://www.tottori-u.ac.jp/kouhou/kisokusyuu/reiki_honbun/u0950581001.html).
The ICR mice were divided into 3 groups according to treatment: gold colloid particle–instilled group (n = 10); distilled water–instilled group (n = 8); and untreated group (n = 12). Gold colloid particles with a diameter of 20 nm (EM.GC20; mean diameter, 19.6 nm) were obtained from BB International (Cardiff, UK). For administration of the particles, the mice were anesthetized by an intraperitoneal injection of sodium pentobarbital (5 mg/100 g body weight), and then 100 µg of 20 nm gold colloid particles were intratracheally instilled. Control mice received 100 µl distilled water. The colloid gold particles were deflocculated via ultrasonic disintegration for 3 min and agitated immediately before intratracheal instillation.
At 15 min after instillation, the mice were killed by exsanguination under deep anesthesia induced by intraperitoneal injection of sodium pentobarbital. The lungs were then immediately fixed for morphological localization of the gold colloid particles.
Pathological Examination
At necropsy, the trachea was immediately exposed and the lungs gently expanded to inspiratory volume by intratracheal instillation of 0.5 ml of 10% neutral-buffered formalin (NBF) via a tracheal cannula; NBF instillation was carried out within 15 min after death. The intact lungs were immersed in 10% formalin for 24 hr to complete fixation after which the lung lobes were separated, and transverse slices were placed into embedding cassettes, routinely processed, and embedded in paraffin for histopathological examination, autometallography, immunohistochemistry, and immunofluoresece. Untreated lungs (n = 3) as well as the lungs treated with gold colloid particles (n = 4) and distilled water (n = 3) were used for pathological examination. Sections of approximately 3-µm thickness were cut and stained with hematoxylin and eosin (H&E). Histological examination was performed by 2 pathologists.
Autometallography Development
Paraffin-embedded sections of the lungs treated with distilled water (n = 3) and gold colloid particles (n = 4) were used for autometallography to detect intracellular gold deposition. The sections were prepared with 1% potassium cyanide for 2 hr to eliminate nonspecific staining from silver sulfides or selenides, and then washed in distilled water for 5 min. Silver amplification was performed by placing the slides in a jar filled with an autometallography developer (50% gum arabic, citrate buffer, hydroquinone, and silver nitrate) in a water bath at 26°C. The entire setup was covered with a dark hood. During autometallography development, an electric device shook the jars gently. After 40 min, the slides were washed in running hot tap water for 30 min and in running tap water for 10 min. Autometallography development was stopped by 5% sodium thiosulfate and Fermer’s solution (20% sodium thiosulfate and 7.5% potassium ferric cyanide) to eliminate excess silver residues. The slides were then rinsed in running tap water for 20 min. The sections were counterstained with hematoxylin. The reaction product appeared as small brown to black grains of silver surrounding an invisible gold core within the tissue. All procedures and protocols were described in detail previously (Danscher and Stoltenberg 2006).
Electron Microscopy
Electron microscopy was performed using the glutaraldehyde-fixed lungs treated with gold colloid particles (n = 3) and distilled water (n = 2), and from those with untreated lungs (n = 1). Cubes of 1 to 2 mm3 were prepared from each section. The small blocks were fixed in glutaraldehyde for 3 hr at 4°C, rinsed in 0.1 M phosphate buffer, postfixed for 1 hr in 1% osmium tetroxide, dehydrated in alcohol, and embedded in epoxy resin. Semi-thin (1 µm) sections were stained with 1% toluidine blue to locate interesting areas for electron microscopic examination. Ultrathin sections stained with uranyl acetate and lead citrate were then examined under a TEM-100CX electron microscope (Japan Electron Optical Laboratory, Tokyo, Japan).
Caveolin-1 and Clathrin Heavy Chain Immunohistochemistry
Paraffin-embedded sections of untreated lungs (n = 3) and of the lungs treated with distilled water (n = 3) and gold colloid particles (n = 4) were used for immunohistochemical detection of caveolin-1 and clathrin heavy chain. For antigen retrieval, the sections were placed in citrate buffer solution (pH = 5.4) and microwaved. Endogenous peroxidase activity was quenched with 3% H2O2 at room temperature for 30 min. The slides were then blocked with 10% normal goat serum for 5 min with microwave treatment. Thereafter, the sections were reacted overnight at 4°C with primary antibodies (anti-caveolin-1, Santa Cruz Biotechnology, CA, 1:1,500 dilution), or reacted at room temperature for 30 min (clathrin heavy chain, Cambridge, UK, 1:6,000 dilution). The sections were also reacted with an equivalent amount of phosphate-buffered saline (PBS) as a negative control. After reaction with the primary antibodies, the sections were incubated with a peroxidase-labeled polymer conjugated to secondary anti-mouse and rabbit antibodies (ENVISON + kit/HRP (DAB), Dako, Glostrup, Denmark) for 30 min at room temperature. Positive reactions resulted in brown staining with 3,3′-diaminobenzidine tetrahydrochloride, and the sections were counterstained with hematoxylin.
Western Blot Determination of Caveolin Expression-1 in Lungs
To examine mouse tissues for caveolin-1 expression, approximately 100 mg of lung tissue (3 groups, n = 3 each) was harvested and processed in lysis buffer (Roche, Basel, Switzerland) to extract protein. Tissue lysates were clarified by centrifugation at 14,000 rpm for 10 min and the protein content of the supernatant was determined. Lysate supernatants were diluted 1:1 with ×2 electrophoresis sample buffer (×1 = 125 mM Tris–HCl, pH 6.8, 2% sodium dodecyl sulphate [SDS], 5% glycerol, 0.003% bromophenol blue, and 1% β-mercaptoethanol), and aliquots equivalent to 20 µg total protein per sample were resolved in a 15% SDS/polyacrylamide gel. Gels were electroblotted to a polyvinylidene difluoride (PVDF) membrane (Biorad, UK), and the membrane was probed with a 1:1,000 dilution of the primary antibody, rabbit anti-caveolin-1. After overnight incubation at 4°C, the membrane was washed and probed with the secondary antibody, an anti-rabbit horseradish-peroxidase-conjugated IgG. After incubation for 1 hr, the membrane was washed. The immunoblot procedure was performed using the enhanced chemiluminescence (ECL) kit (Thermo Fisher Scientific, IL) according to the manufacturer’s protocol.
Caveolin-1 and Cytokeratin Double Immunofluorescence
Paraffin-embedded sections of untreated lungs (n = 3) were used for double immunofluorescence to detect caveolin-1 and cytokeratin. Alexa Fluor 488-conjugated goat anti-rabbit IgG antibody (Invitrogen, CA) and Alexa Fluor 555-conjugated donkey anti-mouse IgG antibody (Invitrogen, CA) were used as secondary antibodies.
For antigen retrieval, sections were placed in citrate buffer solution (pH = 5.4) and microwaved. After preincubation with normal goat serum for 5 min in the microwave, the sections were incubated with the primary antibody for caveolin-1 (Santa Cruz Biotechnology, CA, 1:350 dilution) overnight at 4°C, and then incubated with Alexa Fluor 488-conjugated secondary antibody (1:200 dilution) for 1 hr at room temperature. Next, the sections were incubated with proteinase K for antigen retrieval for 15 min at room temperature. Thereafter, the sections were reacted with primary antibody for cytokeratin (Dako, Glostrup, Denmark, 1:400 dilution) overnight at 4°C and then incubated with Alexa Fluor 555-conjugated secondary antibody (1:300 dilution) for 30 min at room temperature. Sections were mounted in plain 80% Tris-buffered glycerol. Analyses were performed with a confocal imaging system (AX-70, Olympus Laboratory, Tokyo, Japan). Specificity control was carried out by replacing the primary antibodies with PBS.
Caveolin-1 Immunoelectron Microscopy
Untreated lungs (n = 2) were used for caveolin-1 immunoelectron microscopy. Upon dissection, the trachea was immediately exposed. A total of 0.5 ml of phosphate buffer containing 4% paraformaldehyde was instilled gently with a syringe via a tracheal cannula at low pressure. After instillation, the lungs were harvested and stored in 4% paraformaldehyde overnight at 4 °C. The pre-embedding method of immunoelectron microscopy was used for the detailed localization of caveolin-1 in the alveolar wall. In brief, pre-fixed frozen sections, 6-µm thick, were prepared on a cryostat after rinsing fixed tissues in PBS containing 10 to 20% sucrose at 4°C overnight. After performing antigen retrieval and blocking as described in the immunohistochemistry section, the sections were incubated overnight at 4°C with anti-caveolin-1 antibody (Santa Cruz Biotechnology, CA) diluted 1:40. The primary antibody was replaced with PBS in the negative controls. The sections were then incubated with goat anti-rabbit IgG conjugated to horseradish peroxidase (Thermo Scientific, IL) for 60 min at room temperature. After being washed in PBS for 30 min, the sections were immersed in 1% glutaraldehyde in PBS for 5 min. The sections were then washed again in PBS for 15 min and immersed in DAB solution without H2O2 for 30 min. The sections were then reacted with DAB solution containing H2O2 (dako liquid dab + substrate chromogen system; Dako, Glostrup, Denmark) and post-fixed for 90 min in 1% osmium tetroxide. The sections were then washed in buffer for 15 min and dehydrated in a graded alcohol series (50–100%), after which they were embedded in Epon-filled Beem capsules; 6-µm thick pre-fixed frozen sections were collected from the surface of the plastic-embedded tissues. Ultrathin sections were cut from the surface of the plastic-embedded tissues. The sections were then stained with uranyl acetate and lead citrate and examined with an electron microscope (TEM-100CX; Japan Electron Optical Laboratory, Tokyo, Japan).
Results
Morphological Examination of Gold Nanoparticles in Lungs
Light microscopy, autometallography development, and transmission electron microscopy were used to localize the gold colloid particles in the lungs.
Light microscopy showed that instilled gold colloid particles were diffused in all lobes of the lungs. Aggregates of the instilled gold colloid particles were found in the alveolar lumen of the lungs of exposed animals (Figure 1C and D). Histological sections from animals instilled with gold colloid particles showed various extents of localized accumulation of the particles in the alveoli and in the cytoplasm of macrophages (Figure 1C and D). Among the particles distributed throughout the lung lobes, some were attached to the alveoli (Figure 1C and D). Compared with untreated lungs (Figure 1A) and lungs treated with distilled water (Figure 1B), lungs treated with gold nanoparticles had a mild increase in the number of activated hypertrophic macrophages attached to the surface of the alveolar lumen (Figure 1C and D).
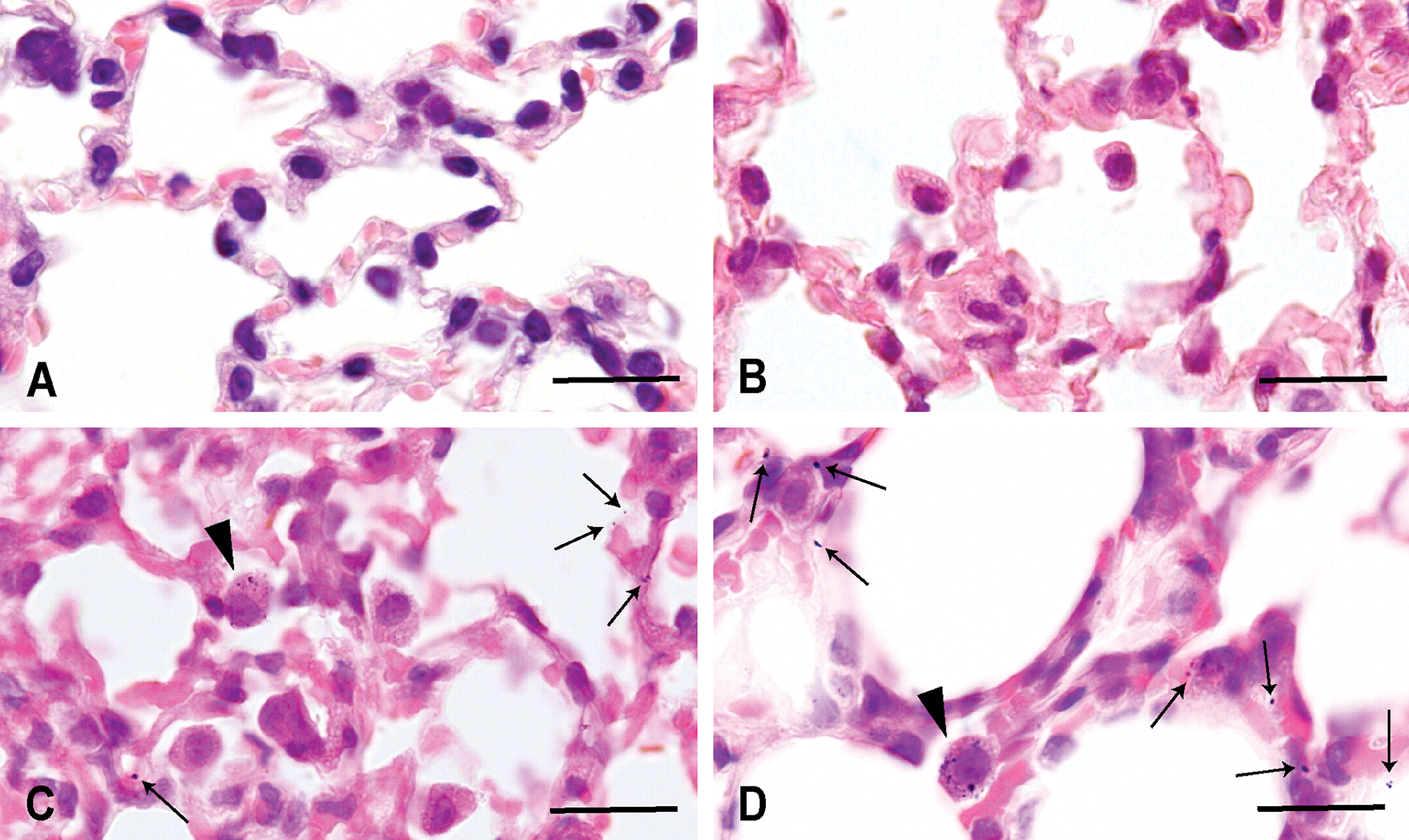
Light micrographs of intact lungs (A) and lungs treated with distilled water (B) and gold colloid particles (C, D). C: Gold colloid particles and an increased number of macrophages in the alveoli. C, D: Gold particles are attached to the surface of the alveoli (arrows) and in the cytoplasm of the macrophages (arrowheads). H&E. Bars = 20 µm.
Autometallography-enhanced gold nanoparticles were detected by light microscopy and appeared as brown to black grains. All autometallography-stained sections from distilled water–instilled lungs were void of gold particles (Figure 2A and C). Most of the instilled gold particles were present in the cytoplasm of macrophages (Figure 2B and D), both those attached to the alveolar wall and those inside the interstitium (Figure 2B). The gold particles were also located in the alveoli and interstitium (Figure 2D). A similar distribution of gold particles was confirmed in lung samples treated with autometallography and H&E staining.
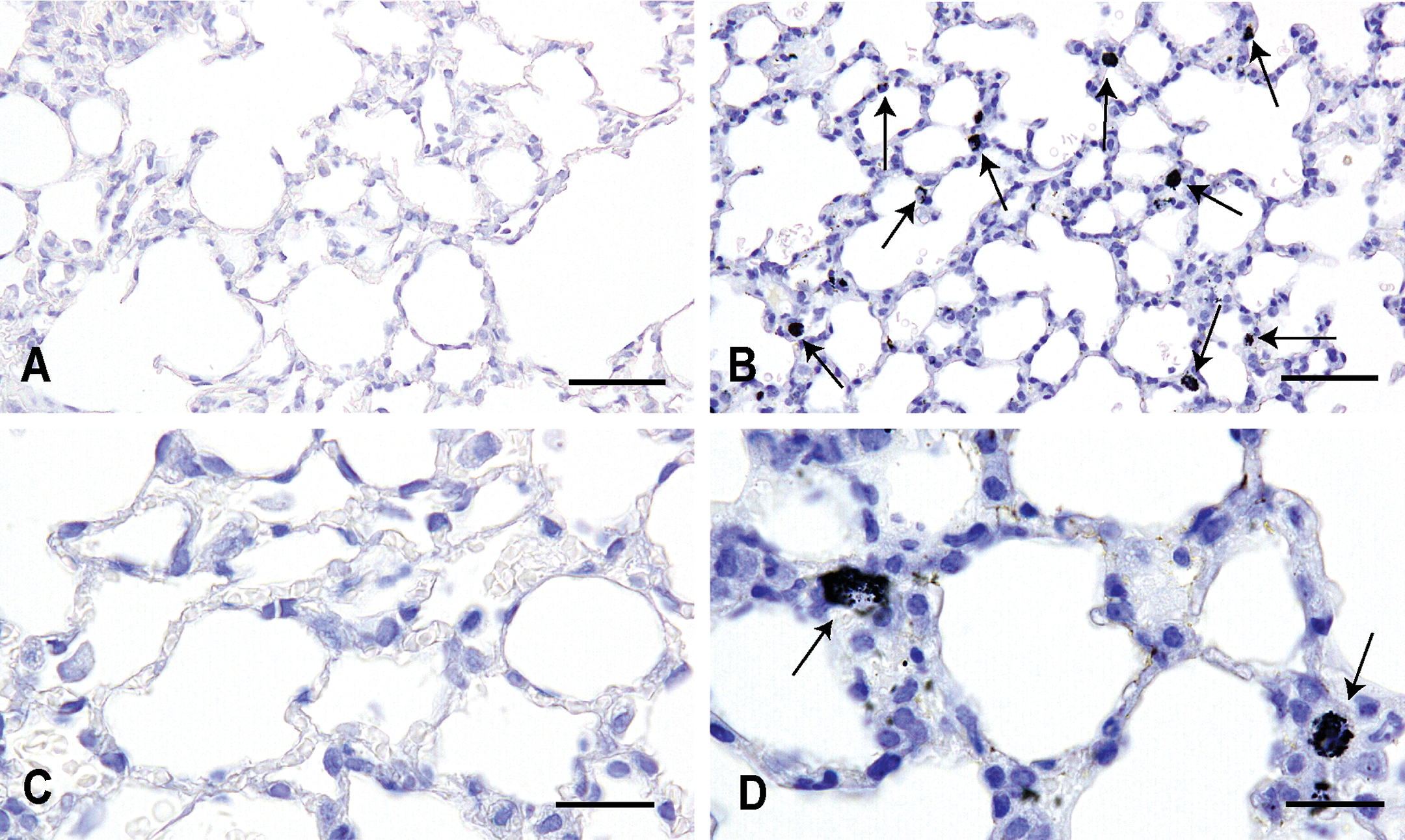
Light micrographs showing autometallography-enhanced gold colloid particles in the lungs of mice treated with distilled water (A, C) and gold colloid particles (B, D). A, C: Autometallography staining void. B: Autometallography-enhanced gold particles observed in the cytoplasm of macrophages (arrows) attached to the alveolar wall and inside the interstitium. D: Gold particles located in the cytoplasm of macrophages (arrows) and in the alveoli. Bars = 50 µm (A, B), 20 µm (C, D).
Gold colloid particles (Figures 3 and 4) appeared as electron-dense spherical particles and were easily distinguishable from other cellular structures by electron microscopy. At 15 min after instillation, gold colloid particles appeared as individual particles or as agglomerations. Particles were localized in endosome/lysosome-like vesicles and in phagosome-like vesicles of macrophages (Figure 3). In addition, particles were observed in the alveolar lumen (Figure 4A), in the endosome-like vesicles of alveolar epithelial cells (Figure 4), and on the surface of type 1 alveolar epithelial cells (AECs) (Figure 4B and C). Occasional flask-shaped invaginations morphologically indicative of caveolae were also observed in type 1 AECs (Figure 4C). Neither the mitochondria nor the nucleus contained gold particles. An increased number of endosome-like vesicles of various sizes in type 1 AECs and endothelial cells were more frequently observed in the lungs treated with gold nanoparticles and distilled water compared with those with untreated lungs.
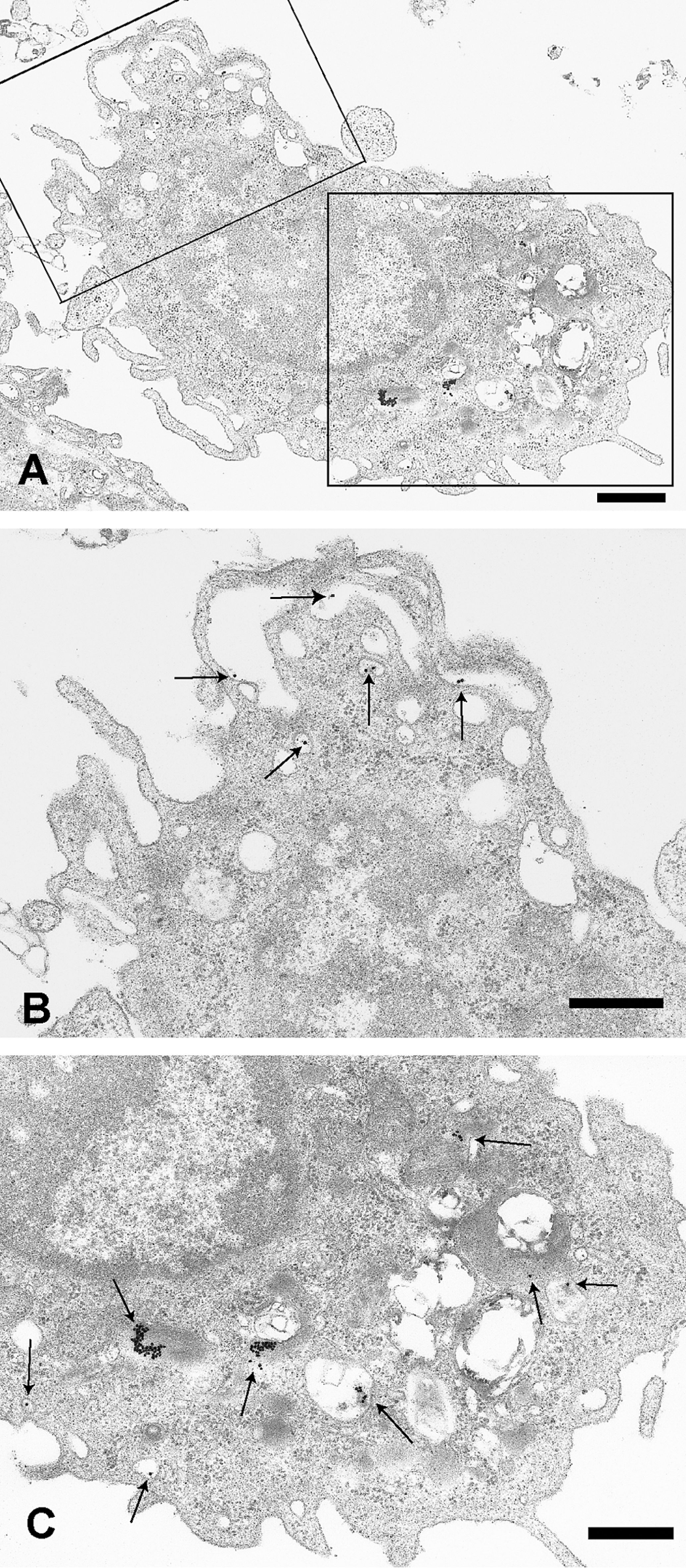
Transmission electron micrographs of gold colloid particles in a macrophage in the lungs of mice treated with gold colloid particles. A: A macrophage containing gold particles. B: Higher magnification of the left oblong box in A. Folding of the cell membrane associated with particles (arrows). C: Higher magnification of the right oblong box in A. Both individual and agglomerated particles (arrows) were localized in the endosome/lysosome- and phagosome-like vesicles. Gold colloid particles are indicated by arrows. Bars = 1 µm (A), 400 nm (B, C).
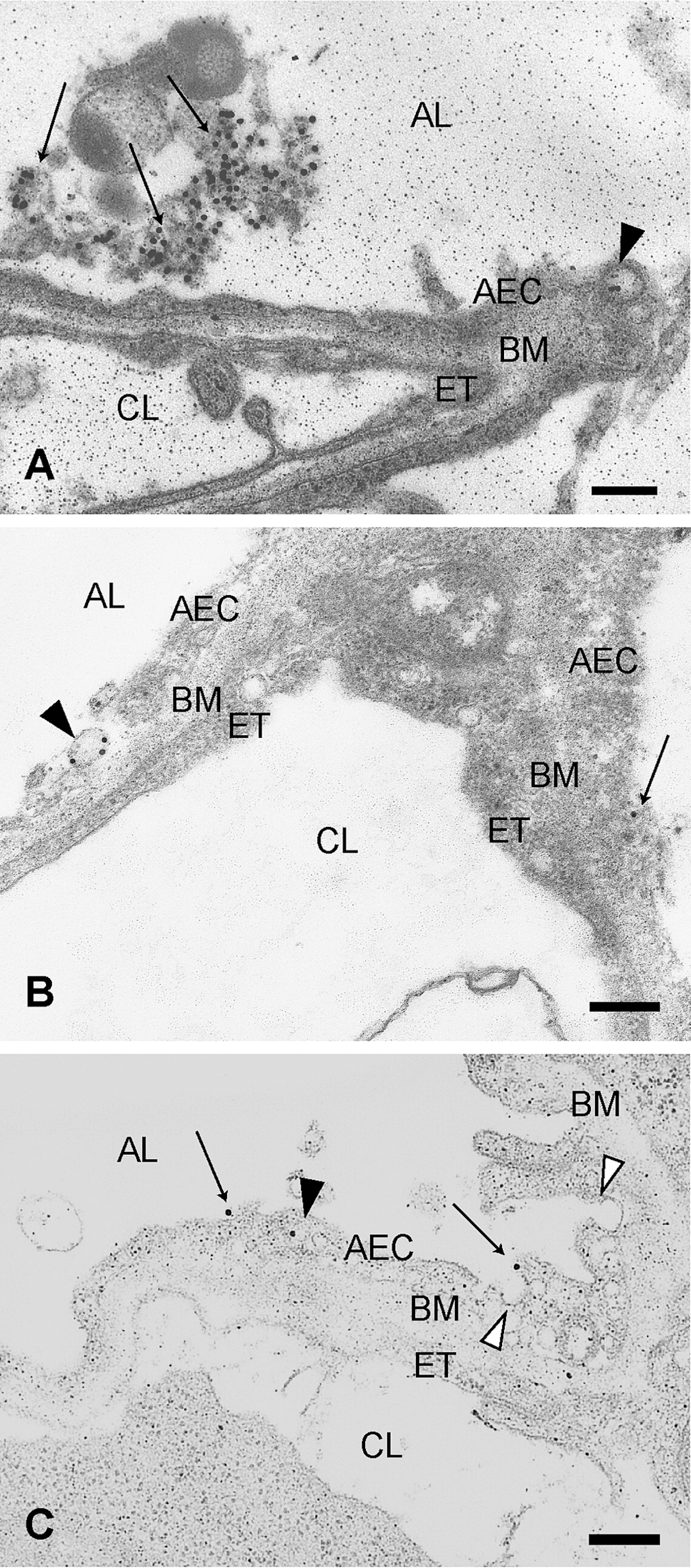
Transmission electron micrographs of the air–blood barrier in the lungs of mice treated with gold colloid particles. A: Particles were observed in the alveolar lumen (AL) (arrows) and in an endocytotic vesicle (black arrowhead) of a type 1 alveolar epithelial cell (AEC). A, B, C: Particles in the endocytotic vesicles (black arrowheads) in a type 1 AEC. B: A particle (arrow) on a type 1 AEC. C: Invagination in a type 1 AEC (white arrowheads) and particles on the surface of the cell (arrows). AEC, alveolar epithelial cell; BM, basement membrane; ET, endothelial cell; AL, alveolar lumen; CL, capillary lumen. Bars = 200 nm.
Morphological and Biochemical Examination of Endocytosis-related Proteins in the Lungs
Strong caveolin-1 immunoreactivity was seen in the cytoplasm of capillary endothelial cells in the untreated lungs as well as in lungs treated with distilled water and gold colloid particles. In addition, the cytoplasm of type 1 (Figure 5A, 5B, 5C) and type 2 AECs and some macrophages showed positive immunolabeling for caveolin-1 in all lung groups; no difference in the intensity of immunoreactivity was noted. The cytoplasm of the macrophages and type 2 AECs also showed positive immunolabeling for clathrin heavy chain in all lung groups (Figure 5D, E, and F). There was no correlation between particle deposition and positive immunolabeling for caveolin-1 and clathrin heavy chain. The specificity of the polyclonal antibodies was validated by the controls, which were consistently negative.
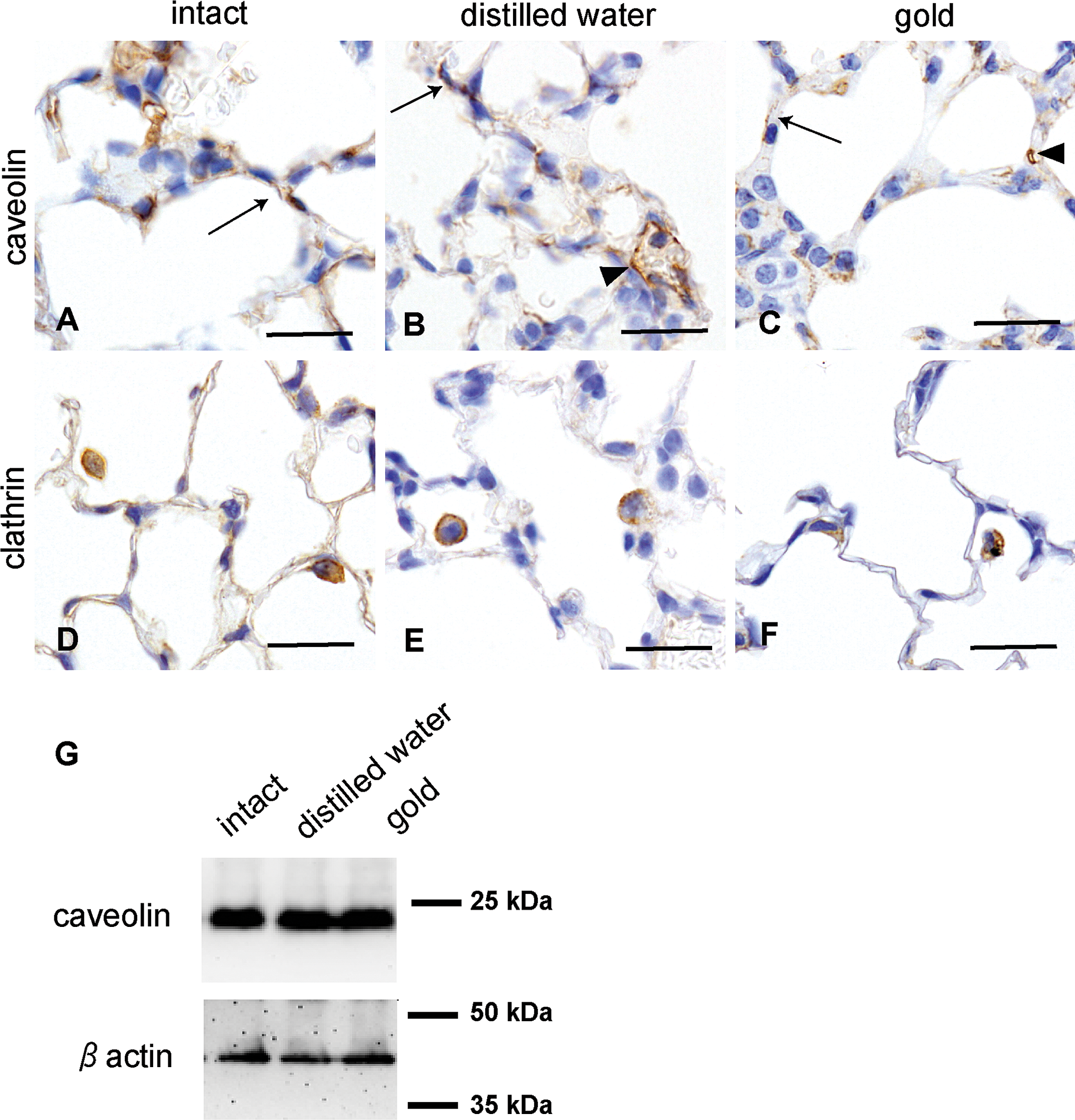
Immunohistochemical localization of caveolin (A–C) and clathrin heavy chain (D–F) in intact lungs (A, D), lungs treated with distilled water (control group) (B, E), and lungs treated with gold colloid particles (C, F). Determination of caveolin-1 by Western blotting of proteins isolated from intact lungs (left lane), lungs treated with distilled water (middle lane), and lungs treated with gold (right lane) (G). A–C: The cytoplasm of type 1 and type 2 AECs (arrows) and endothelial cells (arrowheads) and some macrophages showed positive immunolabeling for caveolin-1 in all lung groups. D–F: The cytoplasm of macrophages and type 2 AECs showed positive immunolabeling for clathrin heavy chain in all lung groups. Bars = 20 µm. G: Note that there was no difference in the amount of caveolin-1 protein between the 3 samples.
Immunoblotting employing untreated lung homogenates (n = 3), distilled water–instilled lung homogenates (n = 3) and gold colloid particle–instilled lung homogenates (n = 3) with polyclonal anti-caveolin-1 antibody revealed a 22-kDa protein. No differences were found in the amount of caveolin-1 protein between the untreated, distilled water–instilled, and gold colloid–instilled lung samples (Figure 5G).
Immunofluorescence analysis revealed strong caveolin-1 immunoreactivity in the vascular wall (Figure 6A). In addition, caveolin-1 immunoreactivity was detected as scattered fluorescent puncta in the ABB (Figure 6A). Cytokeratin immunoreactivity appeared as fluorescent puncta arranged in bright rows in the ABB (Figure 6B). Detailed localization of the caveolin-1-positive immunolabeling in the ABB was demonstrated on sections double-labeled with cytokeratin (Figure 6C). On the merged images, the overlap of the 2 markers (caveolin-1 and cytokeratin) in the ABB was confirmed by yellowish fluorescence (Figure 6C).
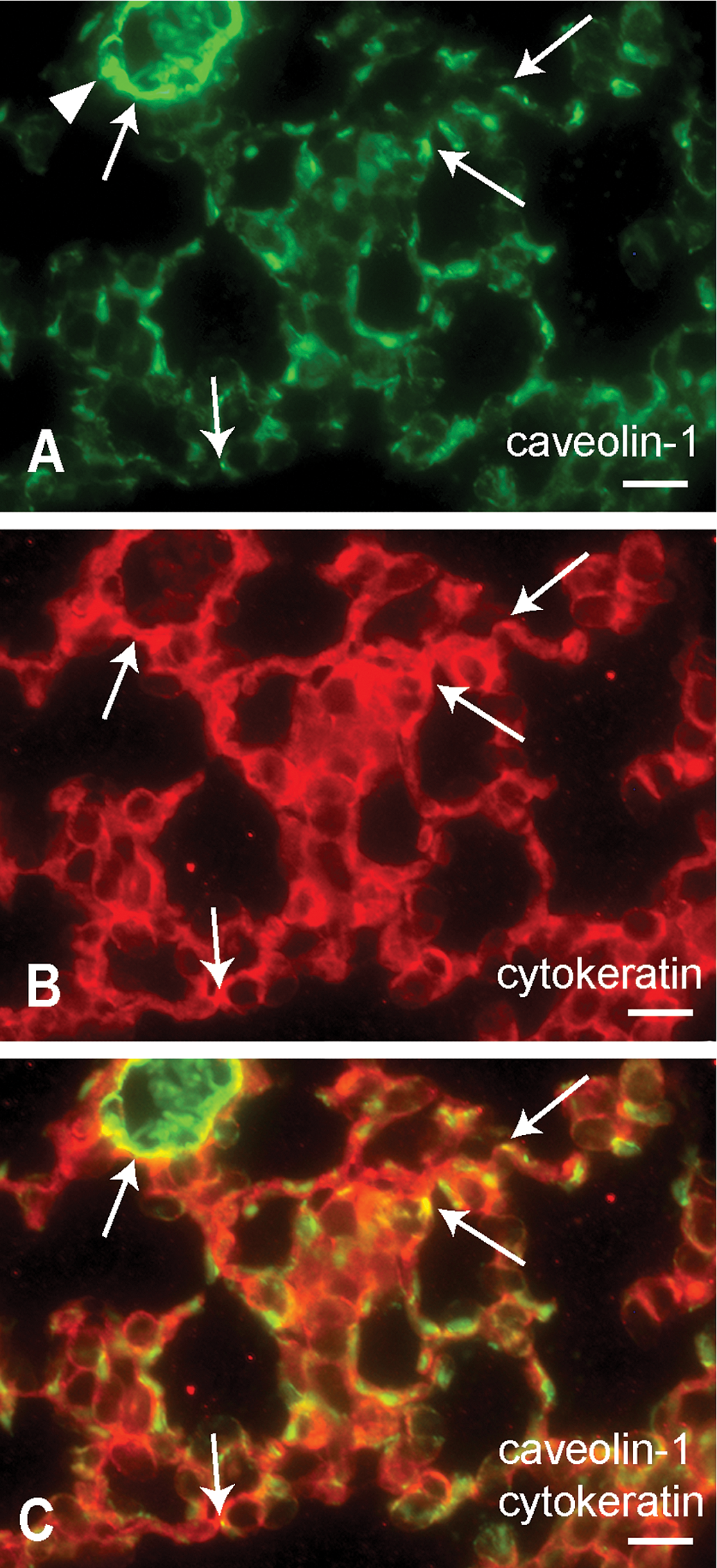
Caveolin-1 and cytokeratin double immunofluorescence in the intact lungs of mice. Double immunolabeling using antibodies against caveolin-1 as revealed by Alexa 488 (green) (A) and cytokeratin as revealed by Alexa 555 (red) (B). Merged image with the 2 fluorophores (C). A: Vascular wall strongly labeled with caveolin-1 (arrowhead). Caveolin-1 immunoreactivity appeared as scattered fluorescent puncta in the air–blood barrier. B: Cytokeratin immunoreactivity appeared as fluorescent puncta arranged in bright rows in ABB. C: Coexpression of caveolin-1 and cytokeratin (yellowish) in the ABB. Arrows in A, B, and C indicate double-stained AECs (caveolin-1-positive and cytokeratin-positive) in the air–blood barrier. Bars = 20 µm.
Immunoelectron microscopic analysis revealed that caveolin-1-positive vesicles were frequently seen in endothelial cells (Figure 7A). Vesicles in type 1 AECs showed occasional positive reactions (Figure 7B), whereas basement membranes were consistently negative (Figure 7). The intensity of immunoreactivity was stronger in the vesicles of capillary endothelial cells than in the vesicles in type 1 AECs. The localization and immunoreactivity intensity of caveolin-1 observed by immunoelectron microscopy were similar to those observed in the immunohistochemistry experiments.
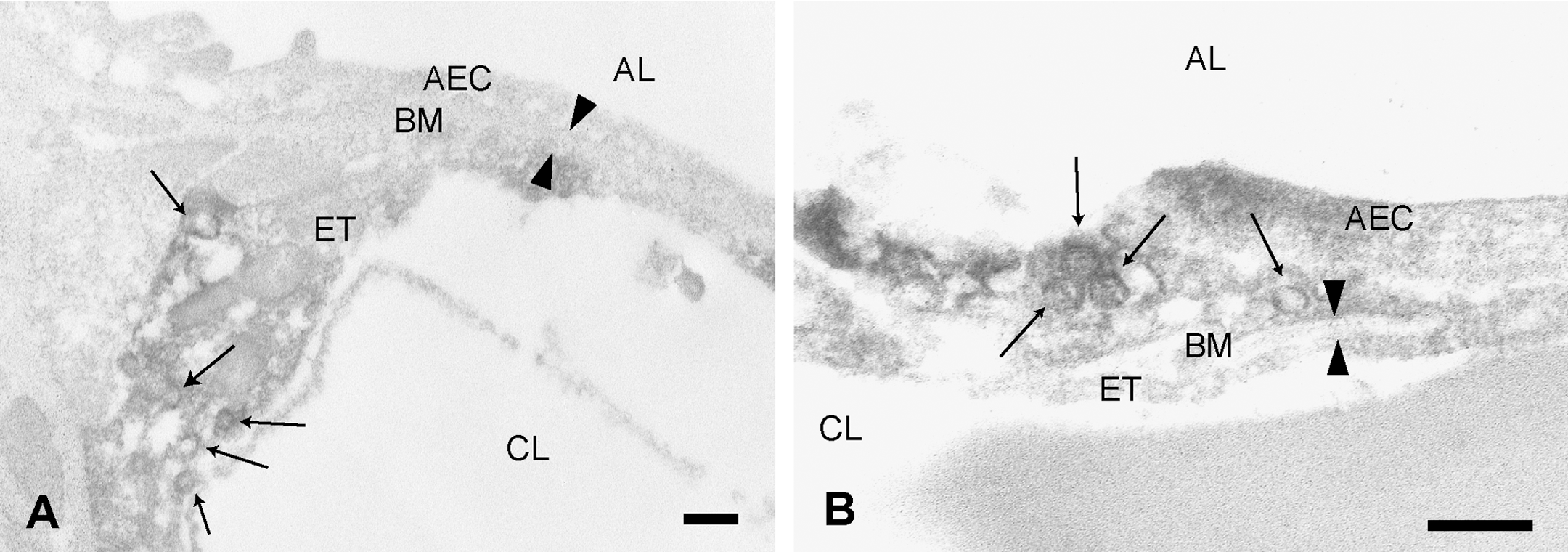
Pre-embedding immunoelectron microscopy for caveolin-1 in the air–blood barrier of intact lungs. A: Vesicles in endothelial cells (ET) showed intense positivity (arrows), whereas the basement membrane (BM) showed no positivity (arrowheads). B: Vesicles in type 1 AECs showed caveolin-1 immunoreactivity (arrows), whereas the BM showed no reactivity (arrowheads). AEC, alveolar epithelial cell; BM, basement membrane; ET, endothelial cell; AL, alveolar lumen; CL, capillary lumen. Bars = 200 nm.
Discussion
Gold colloid particles were used in this study to assess the translocation of nanoparticles at the ABB because spherical gold colloid particles can be easily identified on transmission electron microscopy images. Particles 20 nm in diameter were selected as representative of nanoparticles small enough to be internalized by endocytotic vesicles. The time point at which the evaluations were performed after instillation was chosen from the results of a preliminary experiment in which there was no difference in the localization of gold colloid particles in the lungs at 0, 5, and 15 min after instillation. According to a previous report by Furuyama et al. (2009), most of the instilled 20 nm gold colloid particles in the alveoli were agglomerated at 1 hr after instillation. In addition, particle uptake on the alveolar surface was observed at 15 min after instillation. Therefore, the time point of 15 min after instillation was chosen for examining the translocation of nanoparticles by endocytosis at the ABB in this study.
Phagocytosis is conducted primarily by specialized cells, including macrophages, monocytes, and neutrophils (Conner and Schmid 2003). Pinocytosis is effective for liquids and must be considered (Hastings, Folkesson, and Matthay 2004; Nichols 2003; Ivanov et al. 2005) as a pathway for the translocation of nanoparticles at the ABB. Clathrin-mediated endocytosis is part of this process, but no clathrin-coated vesicles or particles in clathrin-coated vesicles were observed in this study. Furthermore, positive immunolabeling for clathrin heavy chain in the ABB was exclusively observed in type 2 AECs and not in type 1 AECs. These findings suggest that clathrin-mediated endocytosis is a minor mechanism of gold nanoparticle internalization. Another mechanism, caveolae-mediated endocytosis, may also play a role in the internalization of gold particles.
The function of alveolar caveolae is not fully understood. Alveolar caveolae may play a role in mediating transportation of macromolecules (Newman et al. 1999) from the airspace to the blood (Gumbleton 2001; Kim and Malik 2003). At the electron microscopic level, caveolae are most frequently observed as “smooth coated” or “noncoated” omega-shaped invaginations connected to the plasmalemma or plasma membrane by a neck-like structure (Rothberg et al. 1992). The majority of the vesicles present in type 1 AECs are considered caveolae (Gumbleton 2001). A principal component of the striated coat of the caveolae that is also a critical structural and functional element is the cytoplasmically orientated integral membrane protein caveolin (Rothberg et al. 1992). As a biochemical marker, caveolin enables identification of caveolae by a means other than morphological identification alone (Gumbleton 2001; Newman et al. 1999). In this study, smooth-coated vesicles and invaginations, which are suggestive of caveolae, were observed in endothelial cells and type 1 AECs in all lung groups examined by electron microscopy. These findings are consistent with those of our previous study on lungs treated with fullerene nanoparticles (Naota et al. 2009).
The following nanoparticles have been reported to be translocated at the ABB in association with endocytosis in vivo: gold particles (Furuyama et al. 2009; Takenaka et al. 2006); carbon particles (Shimada et al. 2006); and C60 fullerene particles (Naota et al. 2009). In in vitro experiments, internalization of 500 nm-sized latex breads (Rejman et al. 2004) and 60 nm-sized polystyrene particles (Xia et al. 2008) by caveolae-mediated endocytosis has been reported. However, to date, no study has reported the translocation of nanoparticles by caveolae-mediated endocytosis in vivo. The rapidity of the translocation process and the limited extent of translocation of nanoparticles by endocytosis at the ABB may be responsible for the difficulties associated with morphological detection. In this study, electron microscopy revealed gold particles in the caveolae-like vesicles in the cytoplasm of type1 AECs. Immunohistochemistry showed positive immunolabeling for caveolin-1 in the ABB. Both double immunofluorescence and immunoelectron microscopy techniques were used to characterize the antibody reactions and provide unambiguous cellular and subcellular localization of the caveolin immunomarker. Double immunofluorescence revealed colocalization of cytokeratin and caveolin-1. Immunoelectron microscopy showed positive immunolabeling for caveolin-1 in the vesicles of type 1 AECs. These results suggest that 20 nm gold colloid particles are internalized in the alveolar epithelium by caveolae-mediated endocytosis.
In this study, vesicles containing particles were infrequently observed by electron microscopy. There was no correlation between particle deposition and intensity of positive immunolabeling for caveolin-1. There was no difference in the intensity and distribution of positive immunolabeling for caveolin-1 among all lung groups, including untreated lungs in immunohistochemistry; however, in electron microscopy, an increased number of caveolae-like vesicles was observed in capillary endothelial cells and type 1 AECs in both the gold colloid particle–instilled and distilled water–instilled groups. However, in Western blot analysis, no difference in the amount of caveolin-1 protein was found in all groups. Caveolae-mediated endocytosis of macromolecules is one of the physiological functions of the lung (Kim and Malik 2003). The gold particles to which the mice in this study were exposed may have been internalized by this physiological transportation system.
No pathological findings were observed in the lungs treated with gold colloid particles at 15 min after instillation; these findings agree with the results of previous reports, both in vivo (Huang et al. 2008; Lasagna-Reeves et al. 2010) and in vitro (Connor et al. 2005; Brandenberger et al. 2010; Shukla et al. 2005; Wang et al. 2008; Zhang et al. 2010). Gold nanoparticles might thus have a great advantage in terms of their biocompatibility and biological safety and may have potential as a drug delivery system (Huang et al. 2008). Pulmonary drug delivery is gaining increasing acceptance as a convenient, reproducible, and noninvasive technique (Fiegel, Fu, and Hanes 2004; Fu et al. 2002). In the present study, a limited number of particles were internalized by ABB by endocytosis. However, there are various mechanisms of nanocarrier cell internalization that are significantly influenced by the physicochemical properties of the nanoparticles (Hillaireau and Couvreur 2009). Depending on cellular uptake and intracellular trafficking, different surface modifications are possible. A better understanding of the translocation of nanoparticles by endocytosis at the ABB will help resolve the function of these vesicular compartments and their toxicity. Furthermore, detailed information on the translocation of nanoparticles by endocytosis may encourage the development of more specific drug targeting approaches for promoting pulmonary drug delivery.
In conclusion, the results of our study indicate that instilled gold colloid nanoparticles can be internalized by AECs at the ABB by caveolae-mediated endocytosis, which is regarded as a physiological function of AECs.
Footnotes
Abbreviations
Acknowledgments
This work was supported in part by Research Fellowships of the Japan Society for the Promotion of Science for Young Scientists. The authors thank Ms. E. Kawahara at Tottori University for her excellent assistance in electron microscopy and K. Rattanapinyopituk and T. Mukaiyama for their technical assistance.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors received no financial support for the research, authorship, and/or publication of this article.