
Abstract
Depletion of Kupffer cells, known to modulate chemical-induced hepatocellular injury, has not been studied with regard to biliary epithelial injury. Here, the authors investigated the effect of Kupffer cell depletion by clodronate on the toxicity of alpha-naphthylisothiocyanate (ANIT), known to injure biliary epithelium as well as hepatocytes. Up to 99% depletion of Kupffer cells occurred in ANIT and liposome-encapsulated clodronate-treated mice. The effect of Kupffer cell depletion was most evident one day following ANIT treatment. Histologically, there was a modest increase in neutrophil infiltration of the bile ducts, hepatocytic necrosis, and microvesicular vacuolization in the ANIT and clodronate-treated mice, but differences between other groups did not persist. Clinical pathology analytes related to the biliary or hepatocellular injury were significantly elevated in ANIT and clodronate-treated mice compared to mice given clodronate only. This was also true for mice given ANIT and empty liposomes in the case of the biliary analytes. However, group means were typically higher for the ANIT and clodronate-treated group than others on the first 2 days following ANIT injection. These findings suggest that Kupffer cell reduction increases hepatobiliary damage due to ANIT treatment.
Introduction
Hepatocellular injury is one of the more common clinical manifestations of predictable and idiosyncratic drug–induced toxicity (Lee 2010). Hepatocellular injury may be the result of direct chemical-induced injury and/or indirect effects. Kupffer cells, resident hepatic macrophages, as well as hepatic macrophages derived from circulating monocytes, have been shown to play a role in hepatocellular injury produced by several therapeutic and experimental compounds (Adams et al. 2010; Stenger et al. 1969; Laskin et al. 1995; Gardner et al. 2003; Laskin et al. 1988; Ju et al. 2002; Laskin et al. 2011). However, the role of Kupffer cells in chemical-induced injury of the biliary tree has not been widely investigated. Kupffer cells are key modulators of hepatic inflammatory responses. As a source of proinflammatory mediators such as tumor necrosis factor–α, interleukin (IL) 1β, IL-6, IL-10, and nitric oxide, Kupffer cells have been characterized as contributors to drug-induced liver injury (Laskin et al. 1995; Michael et al. 1999).
To better understand the role of Kupffer cells in acute chemically induced injury of the liver, male mice were treated orally with a single dose of 50 mg/kg alpha-naphthylisothiocyanate (ANIT), a classic biliary toxin that causes acute biliary injury (Goldfarb, Singer, and Popper 1962; Plaa and Priestly 1976; Desmet, Krstulovic, and Van Damme 1968; Faiola et al. 2010). Approximately, 24 hr prior to oral administration of ANIT or vehicle (corn oil), mice were given an intravenous (IV) injection of either clodronate-containing liposomes or empty liposomes. Clodronate, a bisphosphonate molecule, selectively kills phagocytic cells such as Kupffer cells by intracellular release of the calcium-binding drug from the liposomes following phagocytosis (Bakker, Sanders, and Van Rooijen 1998). Mice were evaluated for 4 days following ANIT treatment. Assessments included clinical observations, body weights, food consumption measurements, clinical chemistry results, liver (with gallbladder) weights, macroscopic and microscopic observations, and immunohistochemical staining of liver sections for cell proliferation.
Materials and Methods
Animals
One hundred male C57BL/6J mice were obtained from The Jackson Laboratory (Bar Harbor, ME). Mice were approximately 8 weeks of age and weighed 16 to 25 g. Mice were single-housed in polycarbonate solid-bottom cages with Bed-O’Cobs® (The Andersons, Maumee, OH) in a temperature- and humidity-controlled environment with a 12-hr light/dark cycle. Mice were fed LabDiet® brand Certified Rodent Diet 5002 pellets (PMI® Nutrition International, Richmond, IN) and provided reverse osmosis-treated water ad libitum.
All procedures involving animal care and use were conducted in accordance with published guidelines (National Research Council’s Guide for the Care and Use of Laboratory Animals) and the U.S. Department of Agriculture’s Animal Welfare Act and were reviewed and approved by GlaxoSmithKline's Institutional Animal Care and Use Committee.
Mice were randomized into five groups of twenty each using Provantis NT Client Server Oracle (CSOR) while balancing body weight across groups (Table 1). Mice in Groups 1 and 3 were given vehicle (corn oil) orally once on day 1. Mice in Groups 2, 4, and 5 were given a single oral gavage dose of 50 mg/kg ANIT (Batch No. 08420DD, Sigma Aldrich Chemical, St. Louis, MO) formulated as a solution in corn oil at a dose volume of 10 mL/kg on day 1. Groups 3 and 4 were additionally administered clodronate-(dichloromethylene diphosphonic acid, disodium salt; Batch No. 124K4083) containing liposomes (100 µL of a 5 mg/ml suspension) via IV (tail vein) injection approximately 24 hr prior to treatment with vehicle or ANIT, respectively. Group 5 animals were administered empty liposomes (100 µL of a 5 mg/ml suspension) via IV (tail vein) injection approximately 24 hr prior to treatment with ANIT (50 mg/kg).
Group assignments.

ANIT = alpha-naphthylisothiocyanate.
a Given by oral gavage in corn oil vehicle; mice in Groups 1 and 3 received the vehicle at the same dose volume as the ANIT-treated mice (10 mL/kg).
b 100 µL of a 5 mg/ml suspension via intravenous (IV; tail vein) injection approximately 24 hr prior to treatment with vehicle or ANIT.
Clodronate and Liposome Preparation
Dichloromethylene diphosphonic acid, disodium salt (Clodronic acid; Batch Number 124K4083) was supplied by Sigma-Aldrich, St. Louis, MO, USA and was stored at room temperature (15–25°C). Liposomes encapsulation with concentrations of 0 (empty) and approximately 5 mg Clodronate/ml were prepared in accordance with published procedures (Van Rooijen and Sanders 1994). The starting concentration of Clodronic acid was 0.6 M or 0.87 g/5 ml.
In-life Observations and Euthanasia
Clinical observations, body weights, and food consumption measurements were recorded daily. On days 2 through 5 (approximately 24, 48, 72, and 96 hr after ANIT or vehicle administration, respectively), five animals per group were euthanized by exsanguination after administration of isoflurane anesthesia.
Serum Chemistry
Whole blood was collected from the caudal vena cava under isoflurane anesthesia at necropsy from treated and control mice. Serum was separated and analyzed on an Olympus AU640e™ chemistry analyzer (Olympus America, Melville, NY). The following parameters were measured/calculated: alanine aminotransferase (ALT), albumin, albumin/globulin ratio, alkaline phosphatase (ALP), aspartate aminotransferase (AST), total bilirubin (TBIL), direct, and indirect bilirubin when TBIL was ≥20.5 µmol/L, paraoxonase 1 (PON1), total bile acids (BILE), gamma-glutamyltransferase, glutamate dehydrogenase (GLDH), and lactate dehydrogenase (LDH).
Tissue Collection
Following blood collection, liver (with gallbladder) was collected for histologic and/or immunohistochemical examination. Samples of the left lateral and the median lobe of liver including the gall bladder were immersed in 10% neutral buffered formalin for approximately 24 hr and transferred to 70% ethanol. Following fixation, liver sections were processed routinely into paraffin. Six micron sections were stained with hematoxylin and eosin for microscopic evaluation.
Histology
Liver and gallbladders were evaluated using a subjective scale to score hepatic and biliary injury. Biliary and hepatic lesions were scored as follows: 0, no evidence of abnormality; 1, minimal; 2, mild; 3, moderate; and 4, marked manifestation of the lesion. Biliary necrosis ranged from scattered individual necrotic epithelial cells (scored as 1) to complete absence of viable biliary epithelial cells in interlobular bile ducts (scored as 4). Biliary neutrophilic infiltration varied from 1 to 3 neutrophils within or immediately adjacent to the biliary epithelium of an individual bile duct (scored as 1) to greater than 25 neutrophils within the lumen or epithelium of the bile ducts (scored as 4). Fibroblast proliferation referred to the circumferential proliferation of spindle-shaped cells (possibly myofibroblasts) around bile ducts; scoring ranged from the presence of one to two cell layers of reactive fibroblasts (scored as 1) to more than six layers of cells (scored as 4). Hepatic necrosis ranged from scattered individual necrotic cells (scored as 1) to confluent areas of hepatocellular necrosis that bridged between lobules (scored as 4). Only differences in the group mean subjective scores for each diagnosis of >0.4 units were interpreted as meaningful.
Immunohistochemistry (IHC)
Formalin fixed, paraffin embedded sections of mouse liver were cut at 5 microns and mounted on positively charged slides. The following IHC method was performed using the Bond Max Immunostainer (Leica Microsystems, Buffalo Grove, IL). Sections were deparaffinized in Dewax reagent followed by hydration in Bond Wash. Pretreatment was performed using Bond Enzyme 2 (Leica Microsystems) for 10 min. Nonspecific binding due to endogenous IgG was inhibited using Rodent Block M (Biocare Medical, Concord, CA) for 15 min. Next, a rat antimouse F4/80 antibody MCA497G (AbD Serotec, Raleigh, NC) was diluted 1/100 in Leica antibody diluent and applied to sections for 30 min. Detection of antibody was performed using a Rat-on-Mouse alkaline phosphatase polymer (Biocare Medical) for 15 min. Red staining product was developed using the Bond AP Red Chromogen (Leica Microsystems) for 20 min. Sections were counterstained in hematoxylin for 5 min.
In addition, other sections were dual labeled with rabbit polyclonal antibody specific for pan cytokeratin (Abcam Inc., Cambridge, MA) and rabbit monoclonal antibody specific for Ki-67 (Clone SP6, Lab Vision Corp., Fremont, CA) to identify proliferating biliary epithelial cells and hepatocytes. Ki-67 and pan cytokeratin-labeled slides were pretreated with ER2 (Ethylenediaminetetraacetic acid). Ki-67 slides were labeled with Biocare Rabbit on Rodent horseradish peroxidase (Biocare Medical) and developed with Leica Bond Max Refine DAB chromagen (Leica Microsystems Inc., Bannockburn, IL). Pan cytokeratin slides were labeled with Biocare Rabbit on Rodent AP (Biocare Medical) and developed with Leica Bond Max AP Red chromagen (Leica Microsystems Inc.). Sections were counterstained with 4′, 6-diamidino-2-phenylindole (DAPI; Invitrogen, Carlsbad, CA) to label the nuclei.
A quantitative assessment of immunohistochemical labeling in the liver was performed using the iCyte™ Laser Scanning Cytometer (LSC; CompuCyte Corporation, Cambridge, MA). Proliferation of biliary epithelial cells was assessed by automated quantitation of Ki-67 labeled nuclei in cells that were also labeled with the pan cytokeratin antibody (indicative of biliary origin). Hepatocyte proliferation was also assessed using Ki-67 labeling; these cells had a characteristic histologic appearance and were not labeled with cytokeratin markers. The total numbers of DAPI-stained nuclei were counted for each liver section. This population of nuclei was further restricted through the use of gating that was applied to scatter plots of nuclei area versus circularity. In addition, laser light absorbance levels around each nucleus collected using peripheral contours was taken into consideration. Hepatocyte nuclei tended to be larger, more round, and had a moderate level of scatter produced by the hepatocyte cytoplasm in contrast to nuclei of other cell types that were smaller, less round, and had little-to-no cytoplasm. Once gating was applied, region galleries were examined to ensure accurate segmentation of hepatocytes. Multiple test scans were conducted until the optimal placement regions and segmentation of hepatocyte nuclei was achieved. Hepatocytes that were Ki-67 positive were plotted and separated in a similar manner by factoring in the laser absorbance of the Ki-67 positive cells. These diaminobenzidine stained cells had higher absorbance levels than the Ki-67 negative, hematoxylin-stained hepatocytes, which were less dense.
Within the bile ducts, Ki-67 positive and negative nuclei were segmented based on the amount of cytokeratin staining in the peripheral contours. Cells stained with the pan cytokeratin antibody were counted as bile duct epithelial cells. Proliferating cells were separated from nonproliferating based on Ki-67 labeling (DAB). With this method, there was the potential for a peribiliary proliferating cell that is closely adjacent to a bile duct to be counted as a duct cell if the peripheral contour contained cytokeratin staining; however, the peripheral contours were set in close proximity to the primary event nucleus to make this a rare occurrence. Gallery images were examined of regions to confirm optimal placement of gated regions in the scatter plots. A labeling index for hepatocytes and biliary epithelium was developed using the ratios of Ki-67 stained and unstained biliary epithelium and hepatocytes at each time point.
Evaluation of cytokeratin-19 (CK-19)-labeled biliary epithelial cells could not be scored by automated quantitation using LSC due to the low numbers of this cell population; therefore, the proliferation was scored subjectively. Subjective proliferation scores of biliary epithelium dual-labeled with CK-19 and Ki-67 were determined microscopically for each animal (0 = no Ki-67+ nuclei, 1 = minimal number of Ki-67+ nuclei, 2 = mild number of Ki-67+ nuclei, 3 = moderate number of Ki-67+ nuclei, and 4 = marked number of Ki-67+ nuclei). The mean CK-19 biliary epithelial proliferation scores for each group were determined by averaging the scores of all animals in that treatment group. Only differences in the group mean subjective scores for each diagnosis of >0.4 units were interpreted as meaningful.
Statistical Analysis
The serum chemistry and IHC parameters were first analyzed via a one-way analysis of variance (ANOVA) using Proc General Linear Model in SAS (GLM Procedure of SAS/STAT 1999). For the serum chemistry parameters, the results of the Brown-Forsythe homogeneity of variance (HOV) tests and the plots of the residual versus predicted values were found to be unsatisfactory with respect to HOV across treatment groups for a majority of the parameters (Littell et al. 2002). With the exception of two serum chemistry parameters, a transformation to address the lack of HOV between the groups could not be identified. Due to the lack of HOV between the groups, all of the serum chemistry parameters and IHC parameters were then analyzed using methods for unequal variances. The pairwise comparisons were performed via a one-way Welch ANOVA using Proc GLM in SAS (GLM Procedure of SAS/STAT 1999). The Hochberg multiplicity adjustment in Proc Multitest was used (GLM Procedure of SAS/STAT 1999). The multiplicity adjustment was applied to the raw p values for each parameter by day number; adjusted p values are reported. All tests were performed at the 5% level of statistical significance. Statistical analyses were not performed on group mean body weights, food consumption measurements, or liver weights.
Results
In-life Observations
ANIT-treated mice given no liposomes (Group 2), clodronate-containing liposomes (Group 4), or empty liposomes (Group 5) had signs of deteriorating condition which included ataxia, eyes closed, decreased activity (slight to extreme), rough coat, cold to touch, loss of skin elasticity (dehydration, slight to moderate), hunched posture, slow/irregular breathing, and/or tremors (slight; data not shown). These clinical signs were present primarily between days 2 and 5 with decreased activity and rough coat being most prevalent. Additionally, when given the clodronate-containing liposomes with (Group 4) or without (Group 3) ANIT, prostration was observed in a few mice on the day 0 (i.e., the day of liposome administration). One mouse given ANIT and clodronate-containing liposomes (Group 4) died on day 3 immediately prior to scheduled euthanasia and was therefore included in the evaluation. Data from one mouse in Group 5, euthanized on day 5 were excluded because histopathology and clinical pathology evaluation revealed that this animal was inadvertently not dosed with ANIT.
Group mean body weights for all three groups of mice given ANIT were decreased compared with the concurrent vehicle control (Group 1) values as early as day 2 (up to 17%) with the lowest values on day 4 and remained decreased through day 5 (Supplemental Table 1). The injection of clodronate-containing liposomes had no effect on body weight, as the group mean body weight for Group 3 (corn oil vehicle and clodronate-containing liposomes) and Group 1 were similar.
Frequency and severity of subjective scores of liver histopathology.
Mean subjective scores of liver histopathology.a
ANIT = alpha-naphthylisothiocyanate.
a Severity grades were assigned numeric values (not present = 0; minimal = 1; mild = 2; moderate = 3; marked = 4); scores from each animal evaluated were added and the sum was divided by the number of animals examined to obtain the mean.
b Only four animals from group 5 on day 5 were scored.
Group mean food consumption generally correlated with the decreased body weight. The group mean food consumption for the mice given ANIT alone (Group 2), ANIT with clodronate-containing liposomes (Group 4), and ANIT with empty liposomes (Group 5) decreased compared with vehicle control (Group 1) values as early as day 2 (up to 75%) and remained decreased through day 4 (no food consumed; data not shown). However, group mean food consumption was similar to the control values from days 4 to 5. The injection of clodronate-containing liposomes had no effect on food consumption, as the group mean food consumption for Group 3 was similar to that for Group 1.
Clodronate Depletion of Kupffer Cells
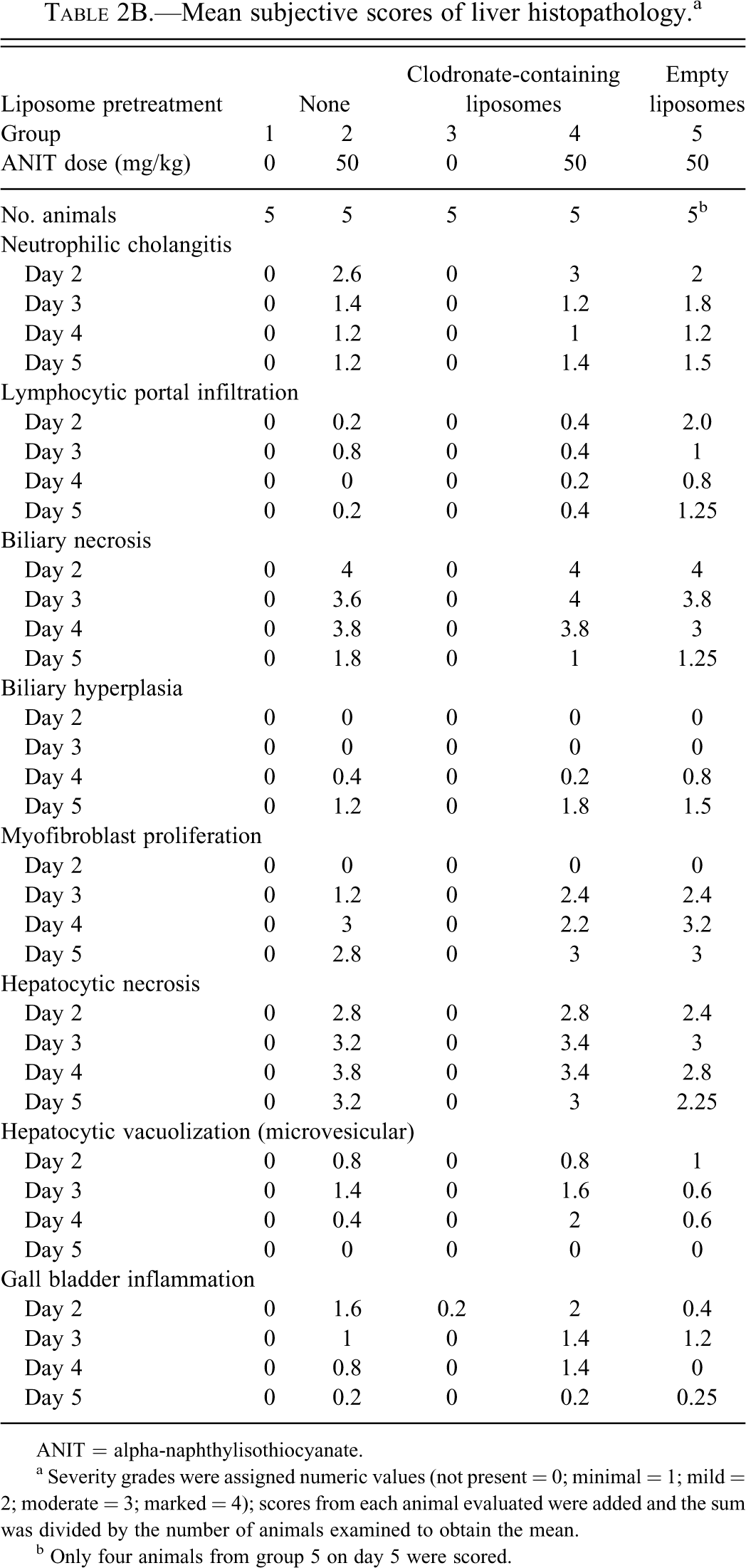
LSC revealed that treatment with clodronate-containing liposomes followed by administration of the corn oil vehicle (Group 3) resulted in a dramatic decrease (p ≤ .05) on days 2 through 4 (>95% on day 3) in the number of Kupffer cells based on F4/80 staining compared with Group 1 (Figures 1A, 2A, and 2B), demonstrating the effectiveness of this approach in depleting Kupffer cells. Likewise, F4/80 labeling intensity decreased (p ≤ .05) between Groups 3 and 1 on days 2 and 4 (Figure 1B). These decreases in number of labeled cells and labeling intensity were no longer evident by day 5. Mice given clodronate-containing liposomes followed by ANIT treatment (Group 4) had lower numbers of Kupffer cells based on F4/80 staining than Group 1 control mice at all time points, with a reduction of more than 90% of the control value on day 3. ANIT-treated mice given clodronate-containing liposomes (Group 4) showed up to 99% and 98% depletion of Kupffer cells compared with the vehicle control group (Group 1) and the group given ANIT and empty liposomes (Group 5), respectively. Groups receiving clodronate-containing liposomes with or without ANIT (Groups 3 and 4, respectively) did not statistically differ with regard to F4/80 labeling, even on day 5.
Immunohistochemical enumeration of cells stained with the mouse macrophage marker F4/80 in livers of mice treated with vehicle or alpha-naphthylisothiocyanate (ANIT) and empty or clodronate-containing liposomes. (A) Staining intensity scores of cells stained with the mouse macrophage marker F4/80 in livers of mice treated with vehicle or ANIT and empty or clodronate containing liposomes. (B).
Treatment with ANIT alone (Group 2) resulted in a decrease (statistically significant on day 2 only) in the number of Kupffer cells as well as in F4/80 labeling intensity compared with vehicle control (Group 1). Mice given ANIT and empty liposomes (Group 5) did not differ from the vehicle control (Group 1) in either Kupffer cell number or F4/80 staining intensity. There was a decrease in the number of F4/80-positive Kupffer cells and in labeling intensity in mice given clodronate-containing liposomes (Group 3) on day 2 (p ≤ .05) as well as a decrease in F4/80 positive Kupffer cells on day 3 compared with the mice given empty liposomes followed by ANIT (Group 5).
Mice given clodronate-containing liposomes followed by ANIT treatment (Group 4) had a greater reduction in the number of F4/80-stained Kupffer cells (p ≤ .05) than mice given empty liposomes followed by ANIT (Group 5) on day 3 only; there were no differences in F4/80 staining intensity for these two groups. No differences were found in either F4/80 cell number or staining intensity between mice given empty liposomes followed by ANIT (Group 5) and mice given ANIT without liposome treatment (Group 2).
Serum Chemistry and Histopathology
Male C57BL/6J mice treated with ANIT had lesions in the bile ducts and hepatocytes that were consistent with previous reports (Kossor et al. 1993; Faiola et al. 2010). Lesions included acute necrosis of the biliary epithelium of interlobular bile ducts with associated inflammation, as well as hepatocytic injury and necrosis and gallbladder mucosal inflammation (Tables 2A and 2B and Figure 3A–F). Treatment with clodronate-containing liposomes followed by oral gavage of corn oil vehicle (Group 3) did not produce significant histologic changes in the liver or gallbladder (Figure 2B). ANIT treatment produced a similar pattern of toxicological pathology, regardless of the pre-ANIT treatment; however, there were modest differences in the histologic scores and serum chemistry parameters among mice treated with ANIT without liposome pretreatment (Group 2), ANIT and clodronate-containing liposomes (Group 4), and ANIT and empty liposomes (Group 5) supporting an interpretation that the loss of Kupffer cells leads to a modest increase in hepatobiliary injury following ANIT treatment.
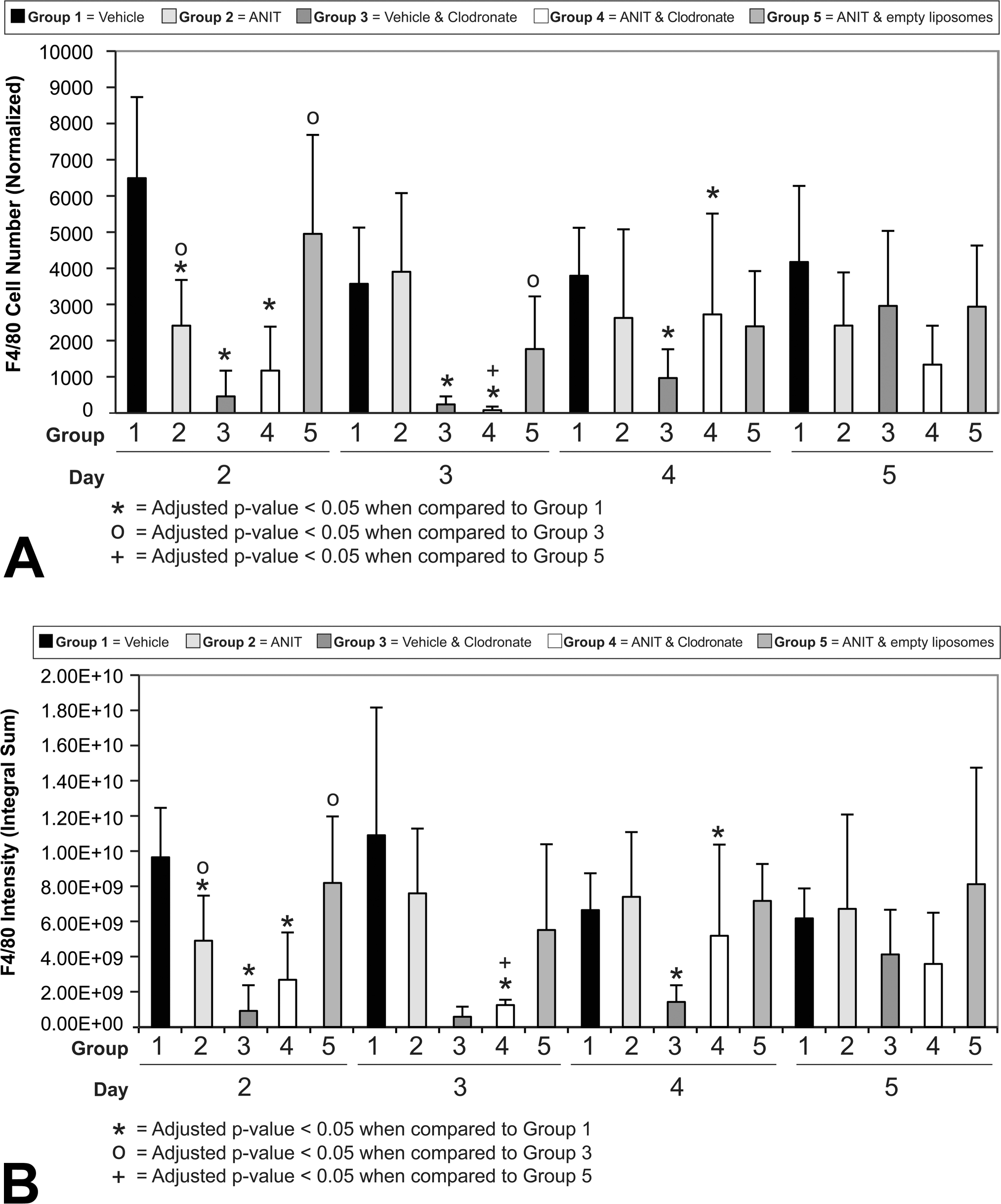
Immunohistochemical stain of livers using antibody to F4/80 from a vehicle control mouse (Group 1, day 2) and a mouse treated with clodronate-containing liposomes and alpha-naphthylisothiocyanate (ANIT; Group 4, day 2). The normal distribution of Kupffer cells is evident in panel A and the significant reduction of Kupffer cells can be seen in panel B.
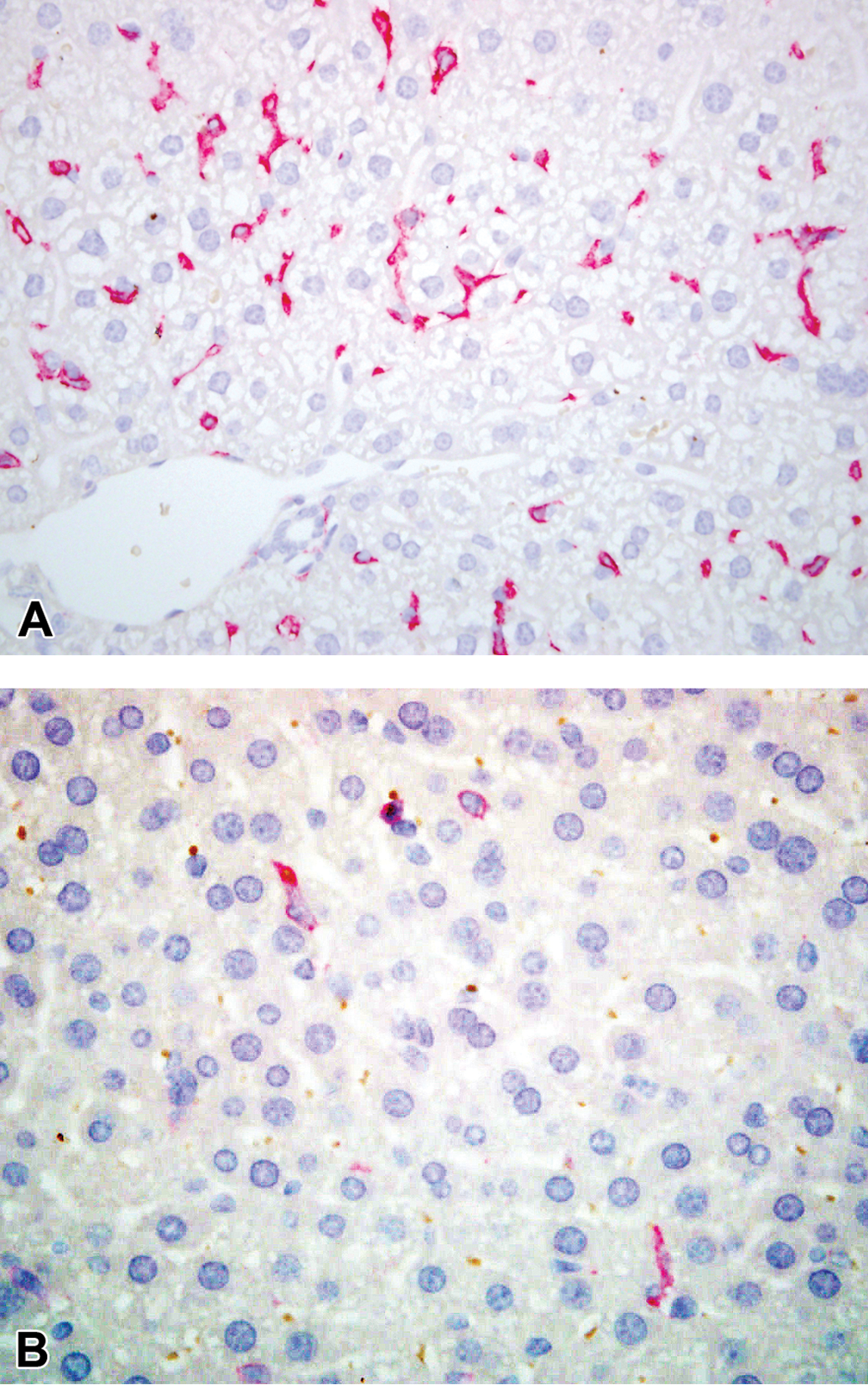
Hematoxylin and eosin (H&E)-stained sections demonstrating the appearance of the vehicle control mice (Group 1, day 2; A) and lesions observed following acute alpha-naphthylisothiocyanate (ANIT) treatment, including neutrophilic cholangitis (Group 4, day 2; B), biliary hyperplasia, (Group 5, day 4; C), prominent circumferential myofibroblast proliferation surrounding bile ducts (Group 4, day 4; D), gall bladder inflammation and focal ulceration (Group 4, day 4; E) and acute hepatocellular necrosis (Group 4, day 3, F).
Biliary necrosis was equally severe in all ANIT-treated groups on day 2. The majority of the intralobular and larger bile ducts contained only eosinophilic debris and a small number of residual biliary epithelial cells that retained their outline, typical of coagulative necrosis. Biliary injury tended to be more severe in mice given ANIT and clodronate-containing liposomes (Group 4) than in mice given ANIT and empty liposomes (Group 5) on days 3 and 4 and in mice given ANIT only (Group 2) on days 3 and 5. By day 5, biliary necrosis had diminished in all ANIT-treated groups but was most evident in mice given ANIT only (Group 2).
Neutrophil infiltration of the bile ducts was more prominent in mice given ANIT and clodronate-containing liposomes (Group 4) than in the other two ANIT-treated groups (ANIT alone [Group 2] and ANIT and empty liposomes [Group 5]) on day 2. By day 3, inflammation was more pronounced in mice given ANIT and empty liposomes (Group 5); but on days 4 and 5, all ANIT-treated groups were similar with respect to inflammation. Neutrophils were found within the lumen of affected bile ducts, between surviving biliary epithelial cells and surrounding affected bile ducts, often in association with significant edema (Figure 3B). Lymphocytic infiltration of the portal tract was more prominent in mice administered ANIT and empty liposomes (Group 5) than the other ANIT-treated groups at all time points.
Biliary hyperplasia was first evident in all ANIT-treated groups on day 4 and was most prominent in mice administered ANIT and empty liposomes (Group 5). Biliary hyperplasia was characterized by an increase in the number of profiles of intralobular bile ducts and their tributaries within the connective tissue of the portal tracts and typically the ducts were evident abutting the interface of the limiting plate and the portal tract. Biliary epithelial cells tended to be more basophilic and robust than normal biliary epithelial cells (Figure 3C). Rare mitotic figures were evident. This response increased in all groups by day 5, at which point, it was most pronounced in mice administered ANIT and clodronate-containing liposomes (Group 4). Circumferential pale eosinophilic, spindle-shaped cells, interpreted as fibroblasts, although it is possible that they were myofibroblasts, were evident around damaged bile ducts on day 3 forming cuffs of cells several cells thick (Figure 3D). These cuffs were more prominent in ANIT-treated groups given liposomes (Groups 4 and 5) compared with mice given ANIT alone (Group 2). On day 4, this response was accentuated in mice given ANIT alone (Group 2) and mice given ANIT and empty liposomes (Group 5), and by day 5, all ANIT-treated groups had a similar degree of fibroblast proliferation.
Gallbladder inflammation, characterized by mucosal and subserosal edema and infiltrating neutrophils and smaller numbers of lymphocytes, was more pronounced in mice given ANIT and clodronate-containing liposomes (Group 4) on days 2 through 4 compared with mice given ANIT only (Group 2). In more severely affected individuals, the lining epithelium of the gall bladder was focally necrotic, leading to ulceration and fibrin exudation into the lumen of the gall bladder (Figure 3E). However, mice given ANIT and empty liposomes (Group 5) had less gallbladder inflammation on days 2 and 4 than mice given ANIT only (Group 2).
Treatment of mice with ANIT (Group 2) resulted in elevations in ALT, AST, BILE, and TBIL as early as day 3; ALP as early as day 4; and GLDH on day 5 (data not shown) compared with the mice treated with vehicle (Figures 4 and 5; p ≤ .05). Differences were also evident between the vehicle-treated mice (Group 1) and those given empty liposomes and vehicle (Group 3) after day 2 (Figures 4 and 5; p ≤ .05), but the magnitude of increase in Group 3 was modest.
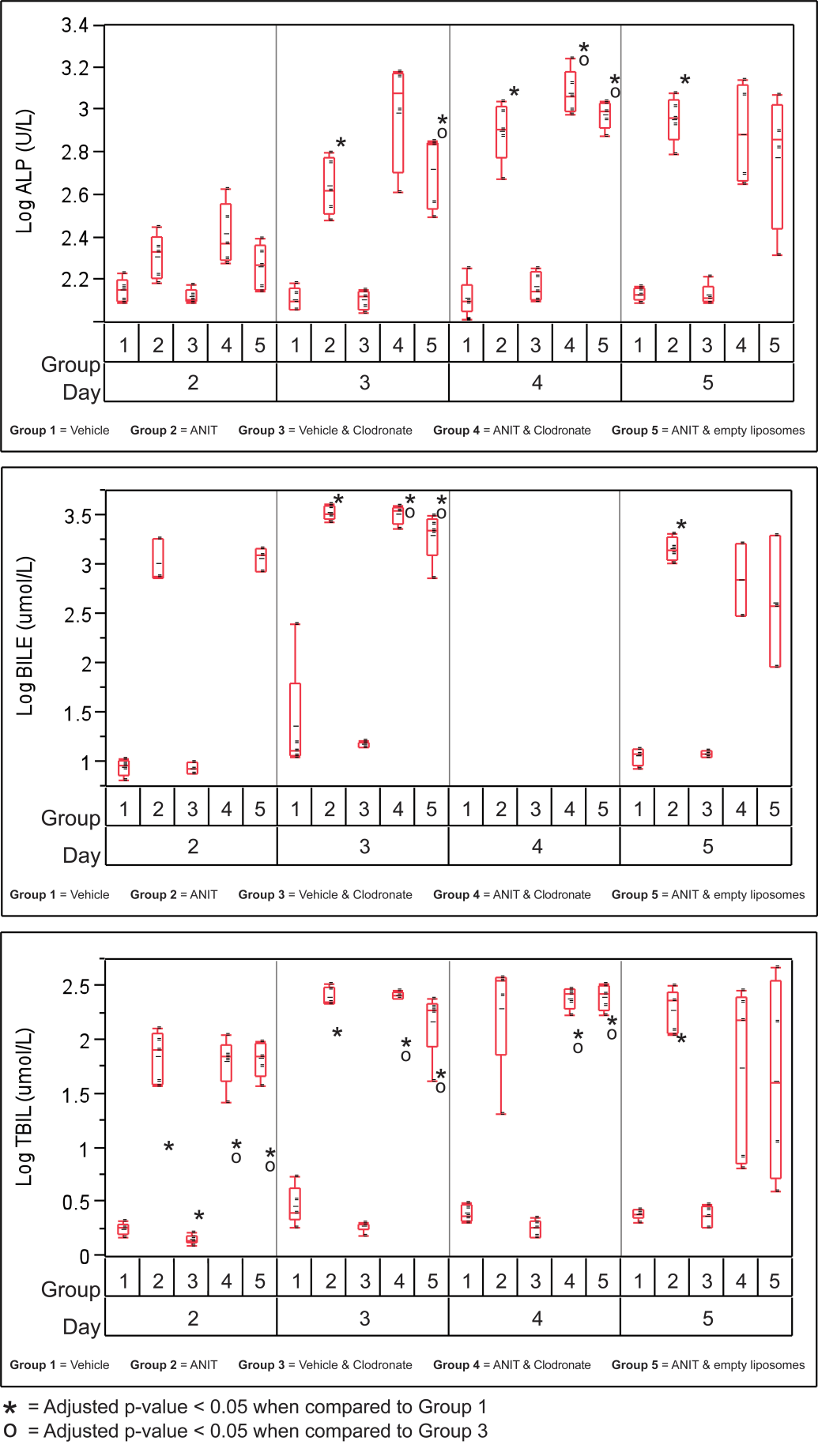
Variability plots for biliary-related serum chemistry analytes from mice treated with vehicle or alpha-naphthylisothiocyanate (ANIT) and empty or clodronate-containing liposomes.
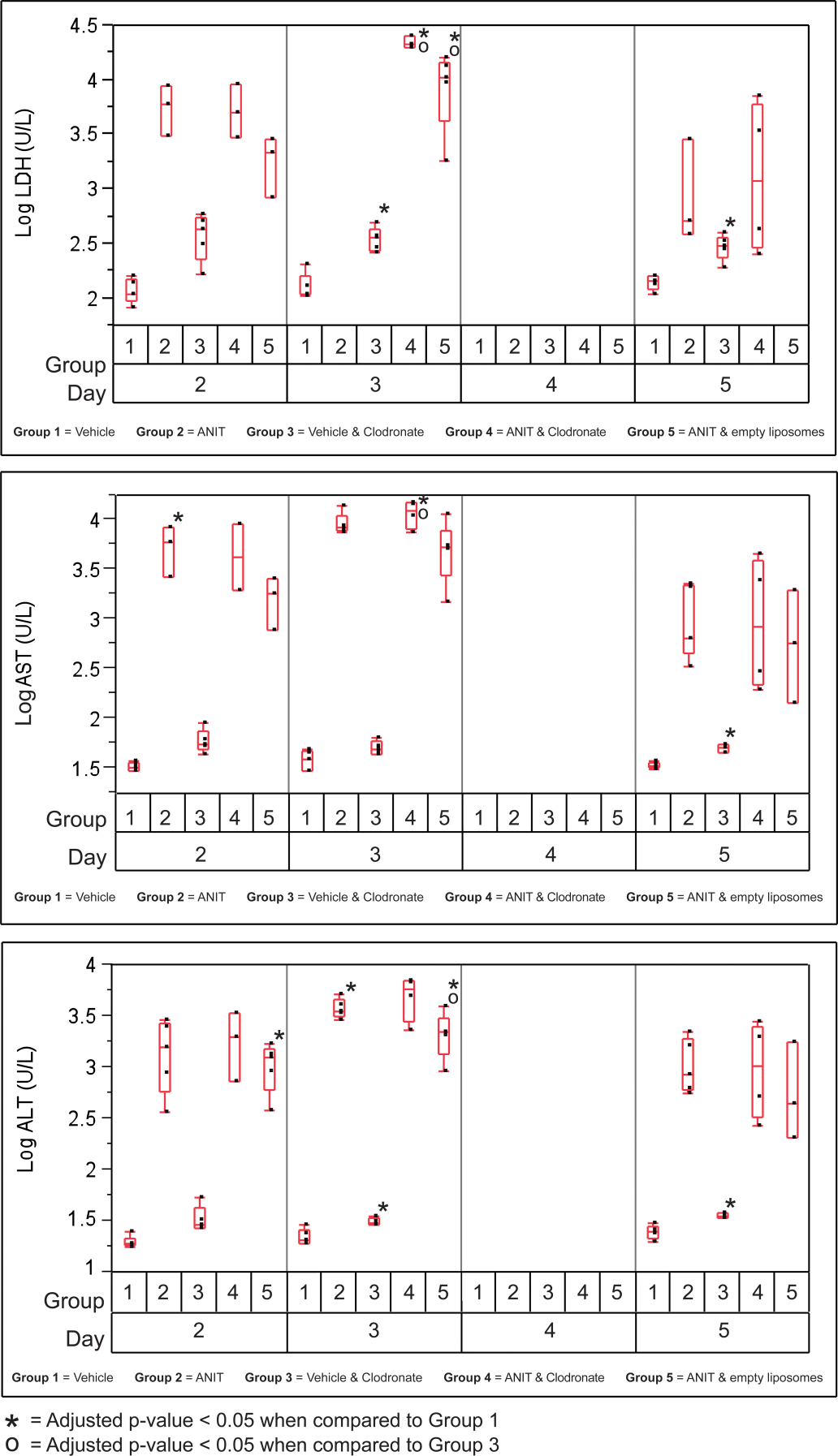
Variability plots for selected hepatocellular-related analytes from mice treated with vehicle or alpha-naphthylisothiocyanate (ANIT) and empty or clodronate-containing liposomes.
Serum chemistry analyte values related to the biliary tree (i.e., ALP, BILE, and TBIL) did not differ statistically between all three ANIT-treated groups (Groups 2, 4, and 5); however, ALP was slightly elevated in mice given ANIT and clodronate-containing liposomes (Group 4) at all time points compared with those given ANIT and empty liposomes (Group 5) as well as at all time points except day 5 compared with those given ANIT only (Group 2; Figure 4). BILE and TBIL were also increased in mice treated with ANIT and clodronate-containing liposomes (Group 4) or ANIT and empty liposomes (Group 5) compared to mice treated with ANIT only (Group 2). The difference was only significant on day 3, however. Mice treated with ANIT and clodronate generally tended to have modestly higher means for biliary related analytes, ALK PHOS, BILE and TBILI than mice treated with ANIT and empty liposomes.
Generally, hepatocellular necrosis was more severe in mice given ANIT and clodronate-containing liposomes (Group 4) than in mice given ANIT and empty liposomes (Group 5) but was similar to the mice given ANIT alone (Group 2; Figure 3F). The pattern of hepatocellular necrosis was similar in all affected groups but varied by intensity. There was multifocal, lytic necrosis of small clusters of hepatocytes, leaving wispy eosinophilic remnants. Most often, groups of 10 to 15 hepatocytes were affected and these foci were arranged at the periphery of portal tracts in the majority of cases. Significant inflammatory infiltrates were not evident. Hepatocellular microvesicular vacuolization that can be caused by accumulation of microvesicular lipid and associated with hepatocellular injury, possibly caused by mitochondrial injury, was similar in all ANIT-treated groups on day 2 but tended to be increased in mice given ANIT and clodronate-containing liposomes (Group 4) on days 3 and 4. Compared with ANIT-treated mice given empty liposomes (Group 5) as well as mice given ANIT alone (Group 2), ANIT-treated mice given clodronate-containing liposomes (Group 4) generally demonstrated greater increases in hepatocyte-related serum chemistry analytes (ALT, AST, and LDH) at all time points but no statistically significant differences were identified (Figure 5).
Proliferation Assay
Biliary epithelial cell Ki-67 labeling indices in mice given clodronate-containing liposomes and corn oil vehicle (Group 3) were not significantly different than in mice given corn oil vehicle only (Group 1) on any day measured (Figure 6A). On days 2 through 5, mice given ANIT and clodronate-containing liposomes (Group 4) had an increase in biliary epithelial cell labeling compared with mice given vehicle and clodronate-containing liposomes (Group 3; 4.6-, 3.5-, 2.8-, and 4.6-fold, respectively). The biliary epithelial Ki-67 labeling indices of mice given ANIT and empty liposomes (Group 5) were increased compared with mice given vehicle and clodronate-containing liposomes (Group 3) on days 2 through 5 (7.1-, 7.7-, 3.7-, and 4.5-fold, respectively). All three groups that received ANIT (Groups 2, 4, and 5) had the largest increase in Ki-67 labeling on day 5 of the study. Given the variability of the data and the lack of statistically significant differences between any of the groups, the effect of clodronate-containing liposome treatment on ANIT-driven biliary proliferation was not clear.
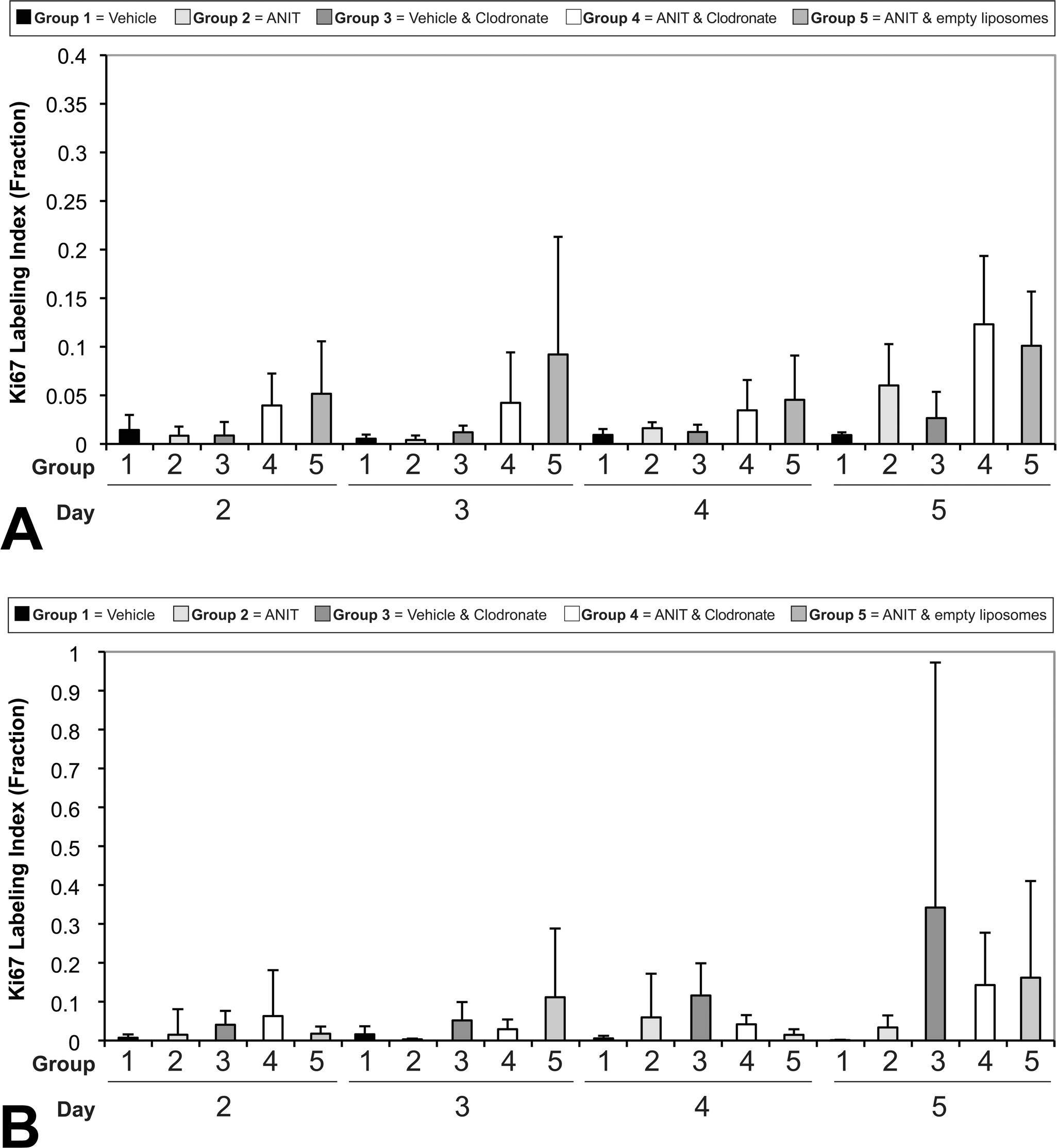
Labeling index for Ki-67 stained biliary nuclei from mice treated with vehicle or alpha-naphthylisothiocyanate (ANIT) and empty or clodronate-containing liposomes (A). Labeling index for Ki-67 stained hepatocellular nuclei from mice treated with vehicle or ANIT and empty or clodronate-containing liposomes (B).
Proliferation of CK-19-labeled biliary epithelial cells as determined by subjective scoring progressively increased in mice treated with ANIT alone (Group 2; first evident on day 4), ANIT and clodronate-containing liposomes (Group 4; first evident on day 3), and those given ANIT and empty liposomes (Group 5; first evident on day 2; data not shown). Mean subjective Ki-67 proliferation scores for biliary epithelium reached a peak on day 5 in the ANIT-treated groups (group mean subjective score for Group 2 = 1.5, Group 4 = 2.4, and Group 5 = 2.25). There was negligible labeling for CK-19 in mice given vehicle (Group 1) and clodronate-containing liposomes followed by vehicle (Group 3) on any day.
There were increases (on days 2–5) in hepatocyte Ki-67 labeling in vehicle control mice given clodronate-containing liposomes (Group 3; Figure 6B). Data from mice given ANIT and clodronate-containing liposomes (Group 4) and ANIT and empty liposomes (Group 5) were more variable but tended to show increased proliferation compared with vehicle control mice (Group 1) and the increases were most evident on day 5. In general, mice in Groups 4 and 5 had lower hepatocellular Ki-67 labeling compared with Group 3 on days 3 though 5 although there were no statistical differences identified in any of the groups at any time.
Discussion
In this study, Kupffer cells were depleted up to 99% in ANIT-treated and vehicle-treated C57BL/6J male mice (Groups 3 and 4, respectively) by intravenously injecting liposome-encapsulated clodronate, confirming that this is an effective approach to deplete Kupffer cells. When mice given ANIT and clodronate-containing liposomes (Group 4) were compared with mice given ANIT and empty liposomes (Group 5), it was apparent that loss of Kupffer cells led to a short-term, mild to modest increase in biliary and hepatocytic injury based on subjective histologic assessment. Relevant lesions included increases in neutrophilic cholangitis and gallbladder inflammation on day 2, hepatocytic necrosis on all days, microvesicular vacuolization on days 3 and 4, and biliary necrosis on day 4. The effects of clodronate-mediated depletion of Kupffer cells were similar, but less pronounced, when mice given ANIT and clodronate-containing liposomes (Group 4) were compared with mice given ANIT without liposomes. Neutrophilic cholangitis was increased in Group 4 compared with Group 2 on day 2, with biliary necrosis increased on day 3 and biliary hyperplasia increased on day 5, as well as an increase in hepatocytic necrosis and microvesicular vacuolization on day 4. In contrast, lymphocytic infiltration of the portal tracts was increased at all time points in mice given ANIT and empty liposomes (Group 5) and generally decreased in mice given ANIT alone (Group 2) compared with mice given ANIT and clodronate-containing liposomes (Group 4). This difference in lymphocytic infiltration might reflect differences in Kupffer cell signaling. In Group 5, Kupffer cells may have been activated following phagocytosis of empty liposomes causing them to release cytokines that recruited lymphocytes. Little Kupffer cell signaling would be likely in Group 4 because phagocytosis of clodronate killed the majority of the Kupffer cells or in Group 2, since these animals did not receive liposomes. Serum chemistry data.
Serum chemistry data also supported the greater injury to the biliary tree and hepatocytes in mice treated with ANIT and clodronate-containing liposomes (Group 4) than mice treated with ANIT and empty liposomes (Group 5) or mice given ANIT alone (Group 2), although the differences were not statistically significant. Biliary proliferation in ANIT-treated mice given clodronate-containing liposomes (Group 4) was also modestly increased in a time-dependent manner, while in mice given ANIT and empty liposomes (Group 5) there was a nontime-dependent increase in biliary proliferation, although there was prominent intra-animal variation in this end point. These findings show that reducing the number of Kupffer cells with clodronate-containing liposomes modestly increased hepatobiliary damage due to ANIT treatment and suggest that Kupffer cells play a role in protecting the liver from acute ANIT-induced biliary and hepatocellular damage. Treatment with empty liposomes was also shown to increase hepatocyte-related enzymes and although the mechanism is not clear, it is possible that the increased burden of phagocytosis led to Kupffer cell activation and release of inflammatory mediators that damaged hepatocytes.
Experimental inhibition of Kupffer cells with dextran sulfate or gadolinium chloride has been reported to inhibit acute acetaminophen-induced hepatic toxicity in rats (Laskin et al. 1995) and mice (Michael et al. 1999). However, Kupffer cells can also serve a protective role in chemical-induced liver injury, as they produce IL-10, IL-6, and IL-18-binding protein (Ju et al. 2002). Elimination of Kupffer cells by intravascular injection of liposomes containing clodronate, a bisphosphonate molecule that selectively kills phagocytes when taken into the cell (Bakker, Sanders, and Van Rooijen 1998), leads to a prominent increase in acetaminophen-induced hepatocellular necrosis in mice (Ju et al. 2002). Kupffer cell depletion in mice has been shown to increase cholestasis-driven liver necrosis following bile duct ligation, an effect attributed to IL-6 release by the Kupffer cells (Gehring et al. 2006). This result suggests that cytokines including IL-10 and IL-6 likely play a significant role in acetaminophen- and cholestasis-induced hepatocellular toxicity, such that in the absence of IL-10 or IL-6, hepatocellular injury is exacerbated. Acetaminophen-induced hepatocellular injury is accentuated in IL-10 knockout mice (Bourdi et al. 2007), providing further evidence supporting this hypothesis.
The contradictory views on the role of hepatic macrophages have been attributed to at least two possible issues. First, it has been argued that the different methods used to eliminate Kupffer cells may not eliminate all Kupffer cell functions (Ju et al. 2002). More recently, different populations of macrophages have been identified within the liver with resident Kupffer cells responding differently than infiltrating macrophages that arrive in response to toxic injury (Holt, Cheng, and Ju 2008).
A similar study conducted in mice treated with clodronate prior to receiving a toxic dose of the hepatocyte toxicant, acetaminophen, showed a much more dramatic increase in hepatocellular injury compared with mice receiving empty liposomes (Ju et al. 2002). One explanation for the differences between the study using a hepatocyte toxicant and this study using a biliary epithelium toxicant may be that Kupffer cell interactions with hepatocytes may have greater impact due their close physical relationship and the close association of hepatocytes to the sinusoids where inflammatory cell-mediated injury is likely to be more intense. The biliary epithelium is sheathed in the connective tissue of the portal tract and may be more isolated from direct and indirect immune-mediated injury that can be modulated by the Kupffer cells and their products. In a related study, IL-10 knockout mice showed only modest differences in biliary injury induced by ANIT compared with wild-type C57BL/6J mice (Faiola et al. 2010). Similar studies on acetaminophen hepatotoxicity in mice with a variety of inflammatory modulators have demonstrated a more pronounced effect than what was observed in the biliary tree in this study (Bourdi et al. 2007; Bourdi et al. 2002; Yee et al. 2007). It is also possible that the dose of ANIT administered in this study was sufficiently high that the effect of modulating the Kupffer cell population was obscured by direct chemical toxicity.
In this study, hepatocytic microvesicular vacuolization, biliary necrosis, hepatocytic necrosis, neutrophilic cholangitis, and gallbladder inflammation were increased in ANIT-treated mice given clodronate-containing liposomes (Group 4) compared with ANIT-treated mice given empty liposomes (Group 5) and mice given ANIT only (Group 2) at one or more time points. These findings suggest that reducing the number of Kupffer cells with clodronate-containing liposomes increased hepatobiliary damage due to ANIT treatment and, that Kupffer cells may play a role in protecting the liver from acute ANIT-induced biliary and hepatocellular damage. Overall, this study supports the view that modulation of the innate immune system by depleting the liver of Kupffer cells can enhance the extent of a chemically induced injury of the biliary tree. Additional studies to further examine the role of Kupffer cells and other aspects of innate immunity in chemically induced biliary injury are needed to further enhance our understanding of these biologic processes.
Footnotes
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors received no financial support for the research, authorship, and/or publication of this article.
Abbreviations
Supplemental Table 1: Intergroup comparisons of body weight (grams).