
Abstract
Paullinia cupana is an Amazonian bush whose seeds have long been used in folk medicine. However, most of the therapeutic properties attributed to this plant are broad and nonspecific, although an antioxidant activity has been reported. On the other hand, cadmium is a heavy metal known for increasing free radicals, hence resulting in cellular oxidative damages. This study was designed to evaluate whether Paullinia cupana is able to reduce cadmium-induced morphological impairment in Wistar rat testis. Adult male Wistar rats 110 days old were ip injected with cadmium (1.15 mg/kg BW [body weight]) and subsequently treated with P. cupana during 56 days. Furthermore, groups receiving either P. cupana extract or cadmium are mentioned. After the treatment period, testis samples were subjected to histological and stereological analyses. Moderate to severe testicular impairments were shown by the animals exposed to cadmium. However, the animals supplemented with P. cupana after cadmium exposure showed a significant decrease in the proportion of damaged seminiferous tubules. Also, P. cupana supplementation was effective in maintaining the number of Leydig cells per testis in the animals exposed to cadmium. In conclusion, P. cupana supplementation was partially efficient in preventing cadmium from damaging the testis of adult Wistar rats.
Introduction
Paullinia cupana var. sorbilis Mart. (Sapindaceae), popularly called guaraná, is an Amazonian plant (Henman 1982) that has long been traditionally used as a stimulant by the Saterê-Mauê Indians (Henman 1986). Also, the folk medicine has attributed a large range of therapeutic properties to this plant, such as increased physical and sexual performance, improvement of cognitive capacity, and weight loss.
Some therapeutic properties of P. cupana seed powder have been scientifically assessed, such as an antioxidant effect associated with decreased lipid peroxidation (Mattei et al. 1998), a weight loss induction (Boozer et al. 2001), a protective effect against gastric lesions induced by ethanol (Campos et al. 2003), and a chemopreventive potential on mouse hepatocarcinogenesis (Fukumasu et al. 2005). Therefore, P. cupana therapeutic effects could be attributed, at least partially, to its antioxidant property (Mattei et al. 1998).
On the other hand, it is currently well established that one of the mechanisms through which cadmium induces cellular damage is by increasing reactive oxygen species (ROS) (Koyuturk et al. 2006; Acharya et al., 2008). Once formed, free radicals are highly reactive and oxidize lipids in membranes, amino acids in proteins, damage nucleic acids, and depolymerize hyaluronic acid (Ochsendorf 1999).
A review of the literature has demonstrated that the testis is particularly sensitive to cadmium, although the extent of testicular damages depends on a number of factors, such as the metal dose and the manner of exposure. Moreover, time-dependent effects have also been shown to play an important role in cadmium toxicity. A slight adjustment in the intraperitoneal dose of cadmium results in severe changes in testicular morphology. Predes, Diamante, and Dolder (2010) and Yang et al. (2006) showed that a single cadmium ip injection of 1.0 mg/kg BW (body weight) did not result in any marked morphological change in rodent testis 56 days following the exposure to the metal. On the other hand, ip doses ranging between 1.15 and 1.2 mg/kg BW are reported to cause drastic alterations in the testis, including seminiferous epithelium depletion, vasoconstriction, and Leydig cell numerical reduction (Leite et al. 2011; Predes, Diamante, and Dolder 2010)
Based upon the data presented, the current study investigates whether P. cupana supplementation, after cadmium exposure, has the potential to reduce the testicular impairments normally observed in Wistar rats exposed to this metal. This is our second study describing such an association (Leite et al. 2011) and the first of its kind in the literature. Therefore, it may contribute to further studies aiming to elucidate the possible therapeutic applications of P. cupana extract.
Material and methods
Chemicals
Powdered P. cupana seed was obtained from EMBRAPA Amazônia Ocidental (Brazilian Enterprise of Agriculture Research). Cadmium chloride (CdCl2) was obtained from SIGMA (St. Louis, MO). Testis samples were embedded in LEICA Historesin (Heidelberg, Germany).
Animals
Male Wistar rats were obtained from the Multidisciplinary Center for Biological Investigation (State University of Campinas, SP, Brazil). Animals were housed two per cage under controlled conditions of temperature (24 ± 4°C) with a 12h-dark/light cycle and provided with water and food ad libitum. This research was approved by the institutional Committee for Ethics in Animal Research of this University (no 2178-1).
Paullinia cupana
P. cupana powdered seed was conserved in dry conditions at ± 4°C during the treatment. P. cupana (2 mg/g BW) was freshly diluted in 1 ml of water and administered by gavage once a day. This dose was chosen based on a previous study (Fukumasu et al. 2005).
Treatment Protocol
Twenty-two male adult Wistar rats (110 days old) were divided into four groups: control group (control, five animals), Cd group (Cadmium, six animals), CdPc group (Cadmium plus P. cupana, six animals), and Pc group (P. cupana, five animals). On the first day of the experiment, Cd and CdPc groups were ip injected with cadmium chloride (1.15 mg/kg BW). Over the following 56 days, the Cd group received daily water gavage (1 mL), whereas both CdPc and Pc groups received P. cupana gavage on a daily basis. All groups were sacrificed on the 57th day after the beginning of the experiment. The treatment period of 56 days was chosen since this interval represents the duration of spermatogenesis in rats (Russel et al. 1990).
Fixation and Processing of the Tissue
At the end of the experiment, animals were anesthetized with xylazine and ketamine (5 and 80 mg/kg BW, respectively). An incision was made in the scrotum and the left testis was exposed and dissected free. The tunica albuginea was gently fissured and the organ was immersed in a solution of glutaraldehyde 4% and paraformaldehyde 4% in 0.1 M phosphate buffer at pH 7.2. The testis fragments were fixed overnight and subsequently processed for light microscopy using routine techniques. For histological analysis, the testis fragments were embedded in Historesin, sectioned at a thickness of 3 µm, and toluidine blue stained (Monteiro et al. 2008).
Stereological Analyses
Representative areas of testicular tissue were photographed with an Olympus Bx-40 microscope and subjected to stereological analyses with an image system: Pro-Plus software version 4.5 (Media Cybernetics). A grid mask system was placed over the images and points were classified as one of the following: seminiferous tubule, intertubular space, blood vessel lumen, macrophage, and Leydig cell nucleus. The volumetric proportions between seminiferous tubule and intertubular space were assessed using a grid mask with 475 points placed over fifteen areas (7.125 points) for each animal at 400× magnification. The volumetric proportions of the intertubular space components were assessed using a grid mask of 2,028 points placed over thirty fields (60.840 points) for each animal at 1,000× magnification. The volume, expressed in ml, of each component described above was determined as the product of the testicular volume and volumetric proportions. Since the specific gravity of the testis is nearly 1.0, its volume was considered the same as its weight (Mori and Christensen 1980). To obtain a more precise liquid testis volume, 6.5% of its weight, relative to the tunica albuginea, was excluded from this organ’s weight (Russel and França 1995).
Leydig Cell Stereology
The proportion between nucleus and cytoplasm of Leydig cells was assessed using a grid mask with 475 points placed over images at 1,000× magnification. One thousand points over nuclei and cytoplasm of Leydig cells were counted per animal. The nuclear diameter of Leydig cells was obtained assessing 10 nuclei/animal. The nuclear volume was calculated using the 4/3πr 3 formula, where r was the mean nucleus radius. The individual volume of Leydig cells was obtained from the nucleus volume and the proportion between nucleus and cytoplasm. The number of Leydig cells per testis was obtained dividing the total nuclear volume of these cells by the average individual nuclear volume.
Statistical Analysis
All data were presented as the mean ± standard deviation, and analyzed via analysis of variance (ANOVA) followed by Tukey test using the program for statistical analysis (SAEG 9.0). The significance level was p < .01.
Results
Biometric Data
The data regarding the statistical analyses of body and testis weight are displayed in Table 1. Both Cd- and CdPc-treated groups showed a significant reduction in both relative and absolute weight of testis, whereas no significant change in body weight was observed for both groups with respect to the control. On the other hand, the Pc group showed a significant reduction in body weight gain.
Biometric data for the study groups in adult male Wistar rats.
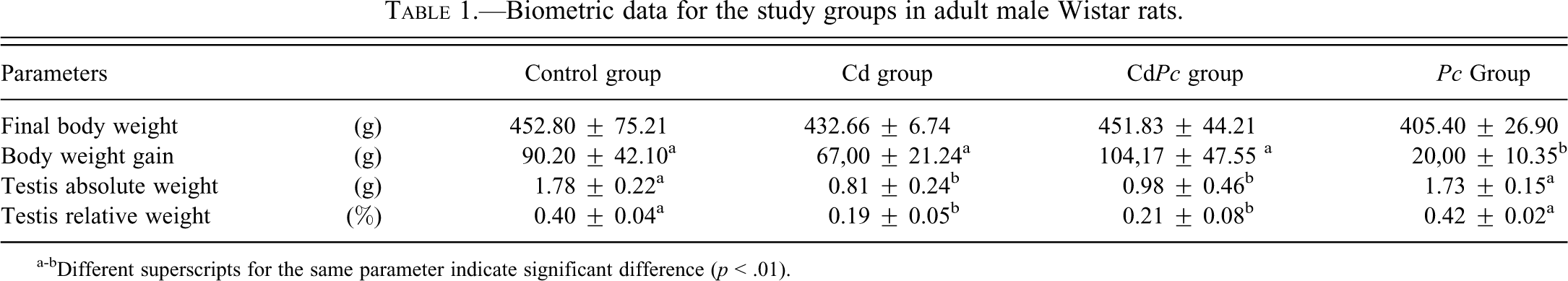
a-bDifferent superscripts for the same parameter indicate significant difference (p < .01).
Histological and Stereological Evaluations
The data regarding statistical analysis of volume (ml) and volumetric proportion (%) are displayed in Table 2. Both Cd (Fig. 2A–D) and CdPc (Fig. 3A–F) groups showed evident alterations in the testicular tissue with respect to the control group (Fig. 1A and B). The Cd group (Fig. 2A–D) had a significant reduction in the volumetric proportion of seminiferous tubules, accompanied by a significant increase of intertubular space. All animals within this group exhibited shrunken seminiferous tubules (Fig. 2A) occasionally filled with cellular debris (Fig. 2C). On the other hand, the CdPc group (Fig. 3A–F) showed no significant alteration in seminiferous tubule and intertubular space volumetric proportion with respect to the control group. Testis sections of some of the animals within the CdPc group (three of six) showed a relatively well-preserved seminiferous epithelium, with distinctive somatic and germ cell populations (Fig. 3A and B). The remaining animals (three of six) had distorted tubules with extensive cell death (Fig. 3C), although a few seminiferous tubules showing a circular shape and filled with an unidentifiable material were also observed (Fig. 3D). In spite of the massive cell death seen in all animals of the Cd group and in half the animals of the CdPc group (three of six), a thin layer of Sertoli cells was often observed internally lining the basal membrane (Figs. 2D and 3E). Stereological analyses also showed a significant decrease in blood vessel lumen volume (ml) and volumetric proportion (%) in both Cd- and CdPc-treated groups with respect to the control (Table 2).
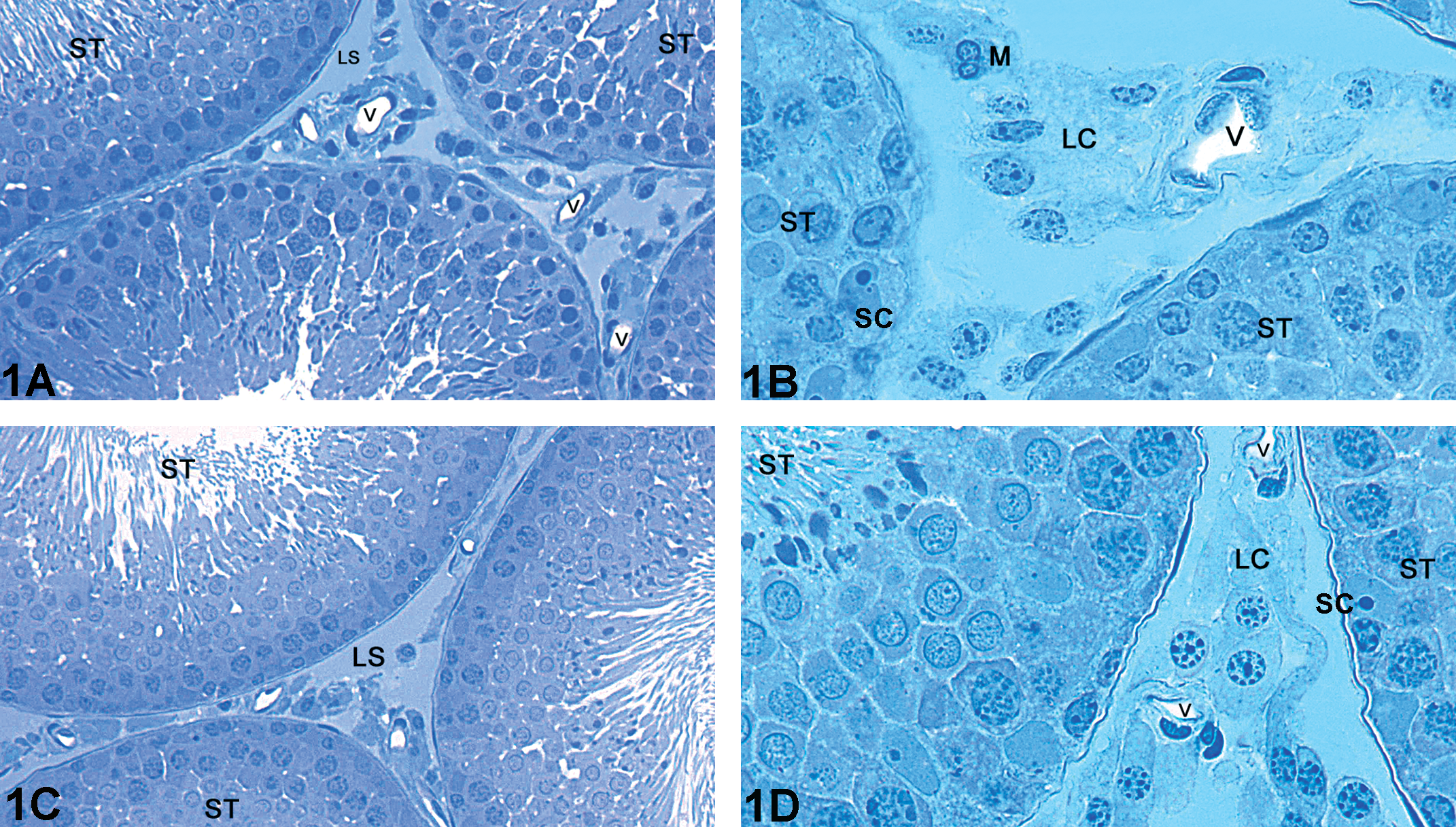
Light microscopy of testis of adult Wistar rat: (A–B) Representative areas of the control group showing Leydig cell clusters, blood vessels, and seminiferous tubules; (C–D) representative areas of the Pc group with a control-like morphology. (ST) Seminiferous tubule; (LS) Lymphatic space; (LC) Leydig cell; (V) Blood vessels; (SC) Sertoli cell. Toluidine blue stained.
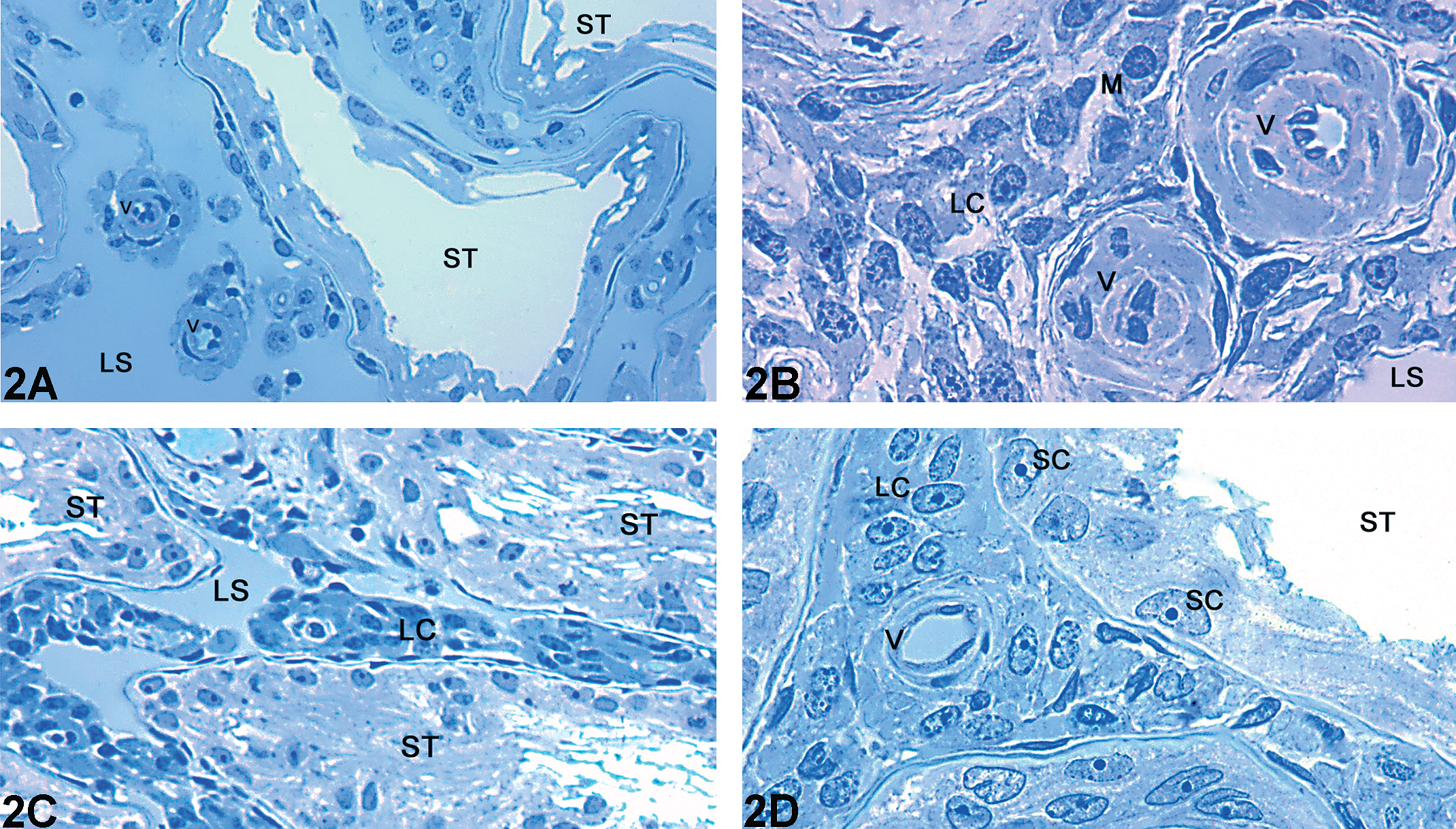
Light microscopy of testis of adult Wistar rat: Representative areas of the Cd group. (A–B) Seminiferous tubules exhibiting extensive damage in the seminiferous epithelium. The Intertubular space is enlarged and contains many Leydig cell clusters; (C) dense clusters of Leydig cells surrounding constricted blood vessels; (D) a thin basal lining of Sertoli cells and absence of germs cells in the seminiferous epithelium. The Intertubular space shows clusters of Leydig cells and a blood vessel with a normal morphology. (ST) Seminiferous tubule; (LS) Lymphatic space; (LC) Leydig cell; (V) Blood vessels; (M) Macrophage; (SC) Sertoli cell. Toluidine blue stained.
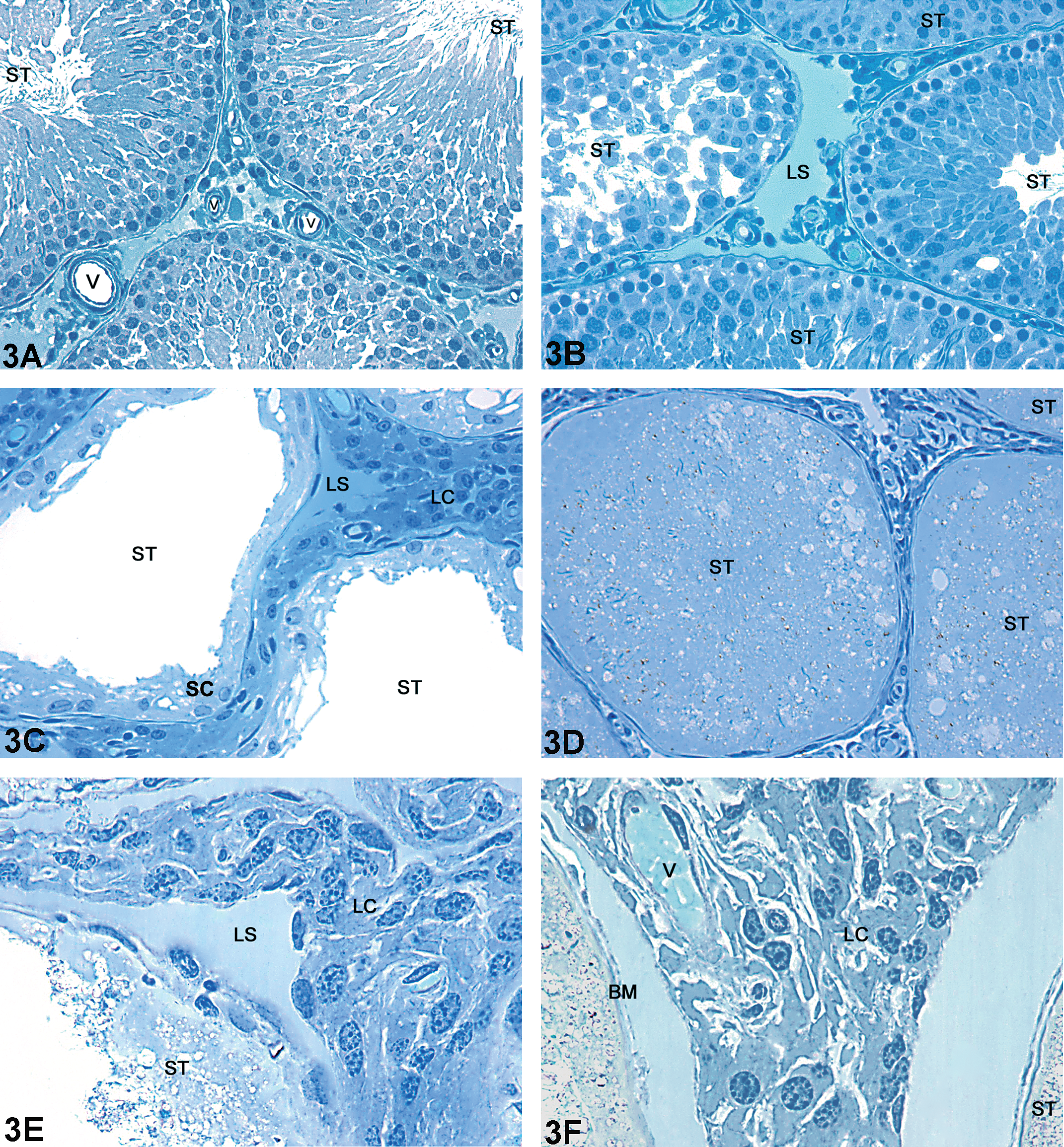
Light microscopy of testis of adult Wistar rat: Representative areas of the CdPc group. (A) Seminiferous tubules and intertubular space exhibiting a control-like morphology; (B) well-organized seminiferous tubules adjacent to one with severely impaired seminiferous epithelium; (C) shrunken seminiferous tubules lacking germ cells and intertubular space with dense clusters of Leydig cells; (D) seminiferous tubule with a circular profile and filled with cellular debris. The intertubular space has dense clusters of Leydig cells; (E) cluster of Leydig cells in the intertubular space; (F) absence of germ cells in the seminiferous tubule, which presents only a thin line of Sertoli cells. The intertubular space shows dense clusters of Leydig cells surrounding a blood vessel. (ST) Seminiferous tubule; (L) Lymphatic space; (Lc) Leydig cell; (V) Blood vessels; (M) macrophage; (BM) Basal membrane; (SC) Sertoli cell. Toluidine blue stain.
Volume (ml) and volumetric proportion (%) of the testis components for the study groups in adult male Wistar rats.
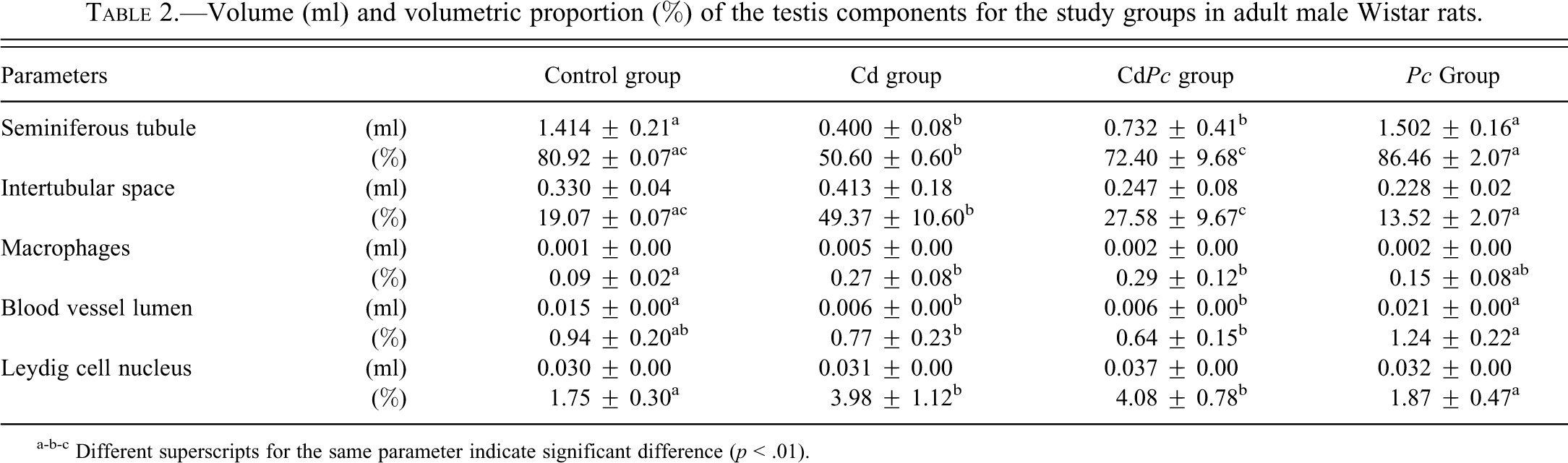
a-b-c Different superscripts for the same parameter indicate significant difference (p < .01).
Leydig Cell Stereology
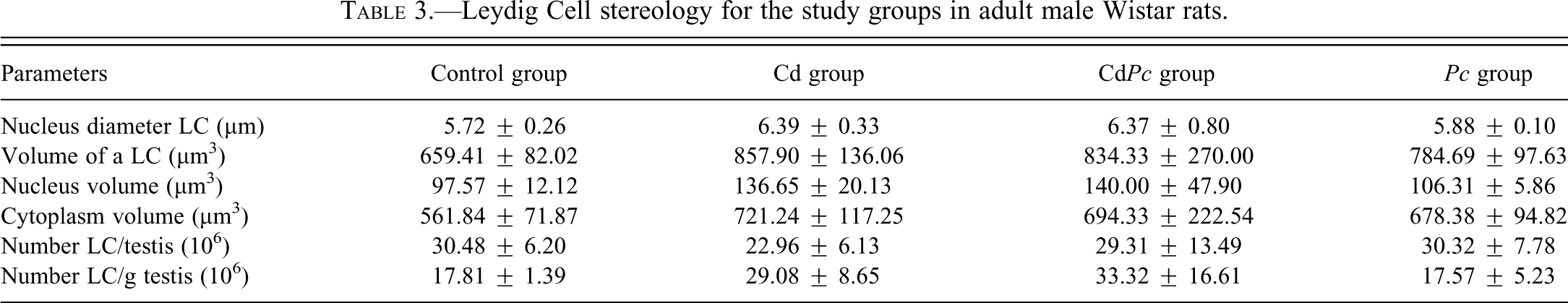
The data regarding Leydig cell stereology is displayed in Table 3. There is no significant alteration in Leydig cell morphology between control group and the remaining groups. Nonetheless, the Cd group clearly exhibited a reduced number of Leydig cells per testis with respect to the control group, although this result is not statistically significant.
Leydig Cell stereology for the study groups in adult male Wistar rats.
Discussion
A vast number of studies have reported the deleterious effects of cadmium on mammalian testis. However, the majority of such investigations have been performed to evaluate cadmium toxicity within a short period after the exposure to the metal. Furthermore, most studies regarding cadmium toxicity in testis have focused on evaluating biochemical and molecular changes, rather than morphological changes.
A previous study showed that rats ip injected with cadmium chloride in a single dose of 1.0 mg/kg BW did not show any marked changes in testicular morphology one week afterwards (Yang et al. 2006). These results are in agreement with a more recent study (Predes, Diamante, and Dolder 2010), in which no evident changes were observed in rat testis 56 days after cadmium ip injection (1 mg/kg BW). In the present research, a cadmium dose of 1.15 mg/kg BW was chosen and the stereological and histopathological analyses were performed after 56 days. Several of our results are in accordance with a previous study (Predes, Diamante, and Dolder 2010), which reported similar morphological changes in rat testis 56 days after being exposed to cadmium (1.2 mg/kg BW).
The literature has shown the potential of antioxidants to decrease cadmium-induced cellular impairments in rat testis. Koyuturk et al. (2006) reported that the treatment with an antioxidant blend prior to cadmium ip injection reduced both oxidative stress and necrotic cell death in rat testis. Moreover, Yang et al. (2006) reported that prior α-tocopherol treatment reduced the degree and number of necrotic seminiferous tubules in rodents ip injected with cadmium. In view of the fact that P. cupana seed reportedly has antioxidant properties (Mattei et al. 1998), this extract is likely to have contributed to reduce epithelial cell death in half the animals from the CdPc group, hence maintaining the seminiferous tubules morphological proportions.
In spite of the absence of germ cells in all animals of the Cd group and in half the animals from the CdPc group (three of six), a thin layer of Sertoli cells was often detected lining the basal membrane within the seminiferous tubules. In a previous study, the exposure to cadmium destroyed various types of testicular cells, including both germ cells and Sertoli cells (Parizek 1960). Nonetheless, Predes, Diamante, and Dolder (2010) reported the presence of seminiferous tubules occupied only by Sertoli cells 56 days after a cadmium injection. According to Hascheck and Rousseaux (1998), Sertoli cells are remarkably resistant to xenobiotics, and in most cases of testicular toxicity, germ cell death or depletion is followed either by regeneration of the germ cell population or by the persistence of shrunken tubules lined only by Sertoli cells. Amara et al. (2008) correlated an increase in ROS in testis of cadmium-exposed rats with damage to DNA integrity. Since Sertoli cells do not undergo meiosis, these cells are less likely to be affected by the adverse effects of xenobiotics, such as cadmium-induced DNA damages. This indirect resistance may account for the permanence of Sertoli cells in seminiferous tubules after cadmium-induced seminiferous epithelium degeneration.
Although the literature lacks information on the effects of cadmium on Leydig cells, reports have suggested that this cellular population might be a target of such metal (Yang et al. 2003). A previous experiment accomplished in our laboratory showed that a single cadmium ip injection (1.15 mg/kg BW) resulted in a slight decrease in Leydig cell number 48 hr after the administration of cadmium. This evidence is supported by the fact that we observed a large amount of cells undergoing apoptosis in the intertubular space whereas few recognizable Leydig cells could be seen. However, the treatment with P. cupana during 56 days prior to cadmium exposure helped maintain the number of Leydig cells approximately the same as the one observed for the control group (Leite et al. 2011). In the current study, a similar pattern of results was found, i.e., the number of Leydig cells per testis is slightly smaller (approximately 25%) in the Cd group, coinciding with the data found in our previous experiment (Leite et al. 2011). On the other hand, the animals that received P. cupana (CdPc group) after cadmium exposure showed nearly the same number of Leydig cells as that observed for the control group. The consistency of these data indicates that P. cupana supplementation either before or immediately after cadmium exposure helps maintain the number of Leydig cells per testis. Such results are coherent with the histopathological analysis of seminiferous epithelium, which showed less morphological alterations in the animals supplemented with P. cupana after cadmium exposure.
The vascular endothelium is also an important target of cadmium and has been related to earlier events of histological damages caused by this metal. A previous experiment of ours (Leite et al. 2011) showed that animals exposed to cadmium at a dose of 1.15 mg/kg BW exhibited a slight decrease in blood vessel lumen 48 hr after the metal administration. In addition, the current study showed a sharp reduction in blood vessel lumen volume in both Cd and CdPc groups 56 days after the animals were exposed to the same dose of cadmium as used in our previous experiment. This result shows that the deleterious effects of cadmium in blood vessels are time-dependent and progressive. Vasoconstriction in animals exposed to cadmium was also reported in preceding literature (Skoczynska and Martynowicz 2005; Tzotzes et al. 2007), and could be explained, at least partially, by the observation that this metal induces intracellular oxidative stress, resulting in decreased nitric oxide synthesis (Skoczynska and Martynowicz 2005).
Conclusions
The present research showed that both Cd and CdPc groups have evident testicular histopathological alterations due to cadmium exposure, mainly in seminiferous epithelium and blood vessels. However, the animals treated with P. cupana during 56 days after cadmium exposure (CdPc group) exhibited a significant reduction in the proportion of damaged seminiferous tubules. Also, P. cupana supplementation during 56 days after the exposure to the metal helped maintain the number of Leydig cells approximately the same as that observed for the control group animals. The mechanisms by which P. cupana protects the testis remain unexplained. We suggest that P. cupana seed powder may act as an antioxidant, either chelating the metal or neutralizing ROS. Therefore, further assays are necessary in order to elucidate this question.
Footnotes
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Supported by the Conselho de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) Scholarship.