
Abstract
The evaluation of neurotoxic damage involves a unique set of challenges. Vulnerable structures, such as neocortex, hippocampus, spinal cord, and peripheral nerve are complex and sharply differentiated; deficits can result from insults to one or more element(s) in the system (e.g., myelin, axon, soma, synapse, or glia). In-life assessment of neurotoxic damage is complicated by the relative inaccessibility of structures in the brain and spinal cord, and recovery is severely limited. Histopathology and electrophysiology represent two of the most commonly used and valuable techniques in this field. This review outlines the strengths and limitations of these procedures and focuses on circumstances in which findings from these measures are dissociated. Electrophysiology is noninvasive and affords a longitudinal view of onset and progression of deficits; however, measures are generally weighted to large-diameter myelinated axons and to regions of primary sensory and motor processing. Histology is a highly validated biomarker, but it is restricted by sampling issues and is insensitive to some elements of neurotoxicity (e.g., altered channel function) associated with profound functional consequences. The central tenet of the discussion is that histology and electrophysiology offer complementary views of neurotoxic damage and, whenever possible, they should be used in concert.
Introduction
The accurate assessment of neurotoxic insult is a paramount medical and societal concern (Spencer, Schaumburg, and Ludolph 2000; Genuis 2008; Sanders et al. 2009; van Valen et al. 2009). Although environmental exposure to some toxicants (e.g., heavy metals) has been reduced in the United States, this exposure has dramatically increased in other parts of the world undergoing rapid and often poorly regulated industrialization. We have little understanding of the long-term sequelae of complex exposure to toxicants such as those associated with war (possible toxic agents, prophylactic drugs, and stress), famine (increased ingestion of cycads, vitamin deficiencies), or environmental disasters (oil leaking into the Gulf of Mexico, chemical dispersants). Recently, the substantial neurotoxic hazards associated with xenobiotic accumulation of chemicals such as organochlorides have also become apparent. Perhaps the most immediate concern is the increase in iatrogenic neuropathies that result in either direct (e.g., chemotherapeutics) or indirect (e.g., antiviral compounds) effects on the structure and function of the nervous system. Finally, it is alleged that exposure to some neurotoxicants at levels subthreshold for immediate effects may nonetheless exacerbate diseases normally associated with aging, such as dementia (de la Monte et al. 2009), Parkinson’s disease (Arif and Khan 2010), and type 2 diabetic polyneuropathy (Lee, Jacobs, and Steffes 2008). These factors have led to a clear and urgent need for well-defined and sensitive biomarkers for the various manifestations of toxic neuropathies.
Assessment of Toxic Neuropathy
The evaluation of neurotoxic damage to portions of the peripheral nervous system (PNS) or central nervous system (CNS) presents a unique set of challenges. Structures such as the brainstem, neocortex, and hippocampus are highly complex and exhibit sharply differentiated regional or laminar boundaries. Deficits can result from a wide range of insults and can either be broad-based (diffuse) or highly focal. Toxic neuropathies may target myelin, axon, plasma membrane, cytoskeleton, neuronal soma, synapse, or glia, in isolation or in combination. In addition, toxin-induced “functional” deficits can involve specific channels, cell processes (e.g., exocytosis), enzymes (e.g., Na+/K+ ATPase), neurotrophic factors, specific neurotransmitters, or an imbalance in excitation and inhibition. While it is clear that functional deficits also involve altered structure, at least at the molecular or subcellular level (e.g., botulinum-induced cleavage of the presynaptic vesicle docking complex), these changes are beyond the scope of conventional histopathologic procedures. The accurate and sensitive detection of early, potentially reversible signs of toxic nerve damage is made more critical by the almost total lack of neural mitosis, which limits repair in the CNS to reorganization. The relative inaccessibility of the brain and spinal cord has been a long-standing limitation of the in-life structural assessment of neurotoxic damage, at both the macroscopic and cellular level. Increasingly, this need is being addressed by novel imaging procedures such as diffusion tensor imaging (Stebbins and Murphy 2009) or by a combination of imaging and staining procedures such as the recent detection of changes in the CNS associated with the early stages of amyloid deposition (Wong et al. 2010).
A number of procedures have been used to assess in vivo toxic neuropathy, including, but not limited to, histopathology, electrophysiology, neuroimaging, and behavioral assessments. Each of these techniques approaches the assessment of nerve damage from a different vector, and each has its strengths and limitations. A key need in the field is a better understanding of how information for the various available techniques can be fused into a coherent and accurate description of the onset and progression of toxicant-induced nerve damage. In this brief review, we will explore the relationship of histopathology to electrophysiology, and we will focus on instances in which findings from these techniques are dissociated.
Strengths and Limitations of Histopathology
In many circumstances, structural neuropathology is arguably the “gold standard” for the assessment of experimental toxic neuropathy. It is of unquestionable validity and specificity, and with proper fixation and staining it can provide a remarkably detailed view of the initial stages of structural neuropathology (e.g., minimal evidence of intracytoplasmic vacuolation) as well as end-stage processes (e.g., Wallerian degeneration, apoptosis). In its more quantitative form, morphometric analysis can assess changes in isolated teased axons or alterations in the relationships between neural elements, such as those that comprise the g-ratio (i.e., ratio of the axon diameter to that of the axon and myelin). Confidence in histopathology is well earned; however, the sensitivity of these measures is often limited by both spatial and temporal sampling restrictions. A classic example is the standard practice in preclinical toxicology of sampling the peripheral nerve at a single, often proximal, site, most commonly the sciatic nerve at the mid-thigh. This practice can miss, entirely, pathology consistent with a length-dependent distal axonopathy in the more distal portions of the axon. A recent study (Schaumburg et al. 2010) compared structural changes at the distal and proximal segments of the rat caudal nerve following exposure to acrylamide. The caudal nerve was selected because it is approximately 50% longer than other nerves in the rat, and therefore more vulnerable to a length-dependent distal axonopathy. Axons in the proximal segments of the nerve were unremarkable at a time when more than 50% of the axons in the distal segments of the same nerve demonstrated severe neurotoxic damage. Structural changes were characterized by focal accumulation and whorls of 10-nm neurofilaments, axon dense and multi-vesicular bodies, and fragmented axons with macrophage invasion. Although histopathology was instrumental in identifying the distal axonopathy and schwannopathy associated with nucleoside analogs for the treatment of HIV (Anderson et al. 1992), the initial preclinical toxicology studies were negative due to the sampling limitation outlined above. It is likely that the growing use of intraepidermal nerve fiber density assessment from repeated skin biopsies (Polydefkis et al. 2004) will add greater time and space sensitivity to histopathologic techniques targeting terminal segments of axons in the PNS. However, the regions of the neuroaxis that can be sampled with this technique are limited, and the long-term value of these measures, especially in animal models, remains uncertain.
Strengths and Limitations of Electrophysiology
Table 1 lists a series of widely used electrophysiologic procedures for the assessment of neurotoxicity in the PNS, along with their sensitivities. The strengths of these measures are obvious. The techniques are noninvasive, they can be assessed at multiple time points, and they are generally sensitive to neurotoxic damage along entire vulnerable tracts or nerves. When properly performed and interpreted, a detailed analysis of a somatosensory evoked potential can outline the pattern and speed of conduction along the majority of a peripheral nerve, the dorsal roots and dorsal columns of the spinal cord, the key relay nuclei in the lower brainstem, the heavily myelinated CNS tracts of the lateral lemniscus, portions of the ventral thalamus, the thalamo-cortical radiations, and regions of primary and secondary somatosensory cortex (Arezzo, Schaumburg, and Spencer 1985). Due to the fact that electrophysiologic measures such as velocity and amplitude are parametric (i.e., continuous data where 40 is truly double 20) rather than categoric or ordinal as is the case for most histologic end points, they may be analyzed using a wide range of statistical tools, which supports the identification of subtle but significant alterations.
Electrophysiologic procedures—Peripheral nervous system.
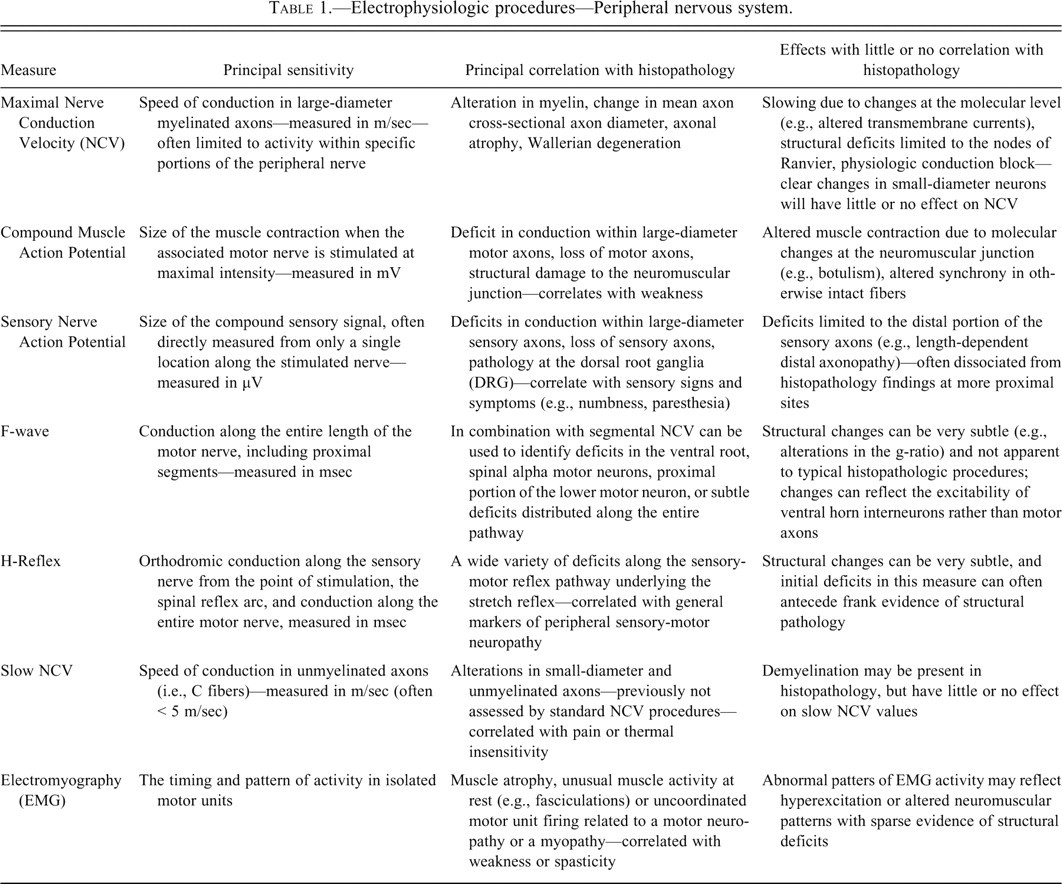
The limitations of electrophysiology are also obvious as these measures are generally sensitive, but not specific. A reduction in the amplitude of an evoked response recorded in Brodmann’s area 17 (i.e., primary visual cortex) may be due to structural damage in one or multiple portions of the visual system or related to transient functional changes with no structural correlate. Further, since most electrophysiologic measures reflect ensemble activity, damage in a small number of neurons can remain undetected in the averaged response. For example, a toxin that alters myelin and slows conduction in 5-10% of axons may be clearly evident in a histopathologic analysis, but undetected by standard electrophysiologic measures (Arezzo and Zotova 2002). If a few axons with maximal cross-sectional diameter and myelin remain intact, velocity can be within normal limits, although response amplitude, duration, and area may be affected.
The insensitivity of electrophysiologic measures to some forms of neurotoxic insult is also due to the fact that whole classes of neurons and entire regions of the brain are not adequately measured by standard electrophysiological techniques. Electrophysiology is not synonymous with nerve function; most measures reflect activity in only a subset of neurons, and then only for a restricted set of functions. When considering toxic neuropathies targeting sensory function in the PNS, it is imperative to differentiate the large-diameter myelinated fibers associated with mechanoreceptors and proprioception from the small-diameter, largely unmyelinated axons processing pressure, pain, and temperature. Large- and small-diameter fibers differ in their anatomy, spinal pathways, neurogenesis, energy utilization, ion distributions, relation to supporting glia cells, sensitivity to differing neurotrophic factors, modalities that they convey, and the signs and symptoms associated with their dysfunction. Standard whole-nerve electrophysiology is only sensitive to the integrity of large-diameter, myelinated axons, leaving even intense functional pathology in small-diameter Aδ and C-fibers undocumented (Arezzo and Zotova 2002). To rectify this situation, recent studies have applied electrophysiologic techniques originally designed to assess transmission within the neocortex (i.e., multiple unit activity) to the assessment of small-fiber conduction in peripheral somatic and autonomic nerves (Zotova et al. 2008). These procedures are capable of measuring nerve conduction velocity to levels of <5 m/sec and are thus sensitive to activity in pain pathways. In parallel, new clinical techniques have emerged (i.e., laser-evoked potentials) that examine cortical activity evoked by rapidly pulsed thermal signals (Pazzaglia and Valeriani 2009). In combination with epidermal fiber density, we now have the tools to examine longitudinal changes in both structural and functional measures of small-fiber neuropathies (Atherton et al. 2007).
In the CNS, the principal limitation of electrophysiology is the fact that almost all available measures rely on computer enhancement of a repetitive event such as sensory evoked potentials (see Table 2 ). This is an effective means of monitoring the integrity of ascending and descending fiber tracts and of primary and secondary regions of sensory neocortex, but when using these techniques, the majority of brain structures are not adequately evaluated. Toxicant-induced damage limited to regions such as association cortex, cerebellum, anterolateral thalamus, hypothalamus, or to nonsensory fiber tracts would be detected by histopathology (if those regions were sampled), but not by standard electrophysiologic measures. Novel electrophysiological techniques have been designed, in part, to increase sensitivity of electrodiagnostic tests to nonsensory regions of the CNS. These techniques include endogenous evoked potentials, alterations in spectral content of electroencephalography (EEG) and the examination of non-phase locked changes in the patterns of brain activity.
Noninvasive electrophysiologic procedures—Central nervous system

Dissociation of Histopathologic and Electrophysiologic Findings
A recent study (Cartwright et al. 2009) clearly illustrates the potential dissociation of histologic and electrophysiologic findings in neurotoxicology studies. The study examined the effects of posaconazole, a potent antifungal compound that causes phospholipidosis in neurons of the brain, dorsal root ganglia (DRG), and spinal cord in dogs. The histologic findings included intracytoplasmic lysosomal vacuoles containing electron-dense laminated whorls of material from portions of the cell membrane. The underlying cause of the observed phospholipidosis is not fully understood, but it is thought to reflect drug-induced interference with the breakdown and turnover of the cell membranes and the possible sequestering of compound by the blood-brain barrier (Cartwright et al. 2009). Phospholipidosis is a relatively common finding in certain tissues, but it is rare in the CNS. This study utilized an extensive battery of validated electrophysiologic measures, including auditory, visual, and somatosensory evoked potentials for the CNS and maximal nerve conduction velocities, compound muscle action potentials, and F-waves for the PNS (see Tables 1 and 2). Although phospholipidosis was present in multiple regions of the CNS, including structures involved in sensory transmission (e.g., DRG, dorsal column nuclei, and thalamus), all electrophysiologic measures were unremarkable, even after a period of 12 months of dosing. One possible interpretation is that the electrophysiologic measures were simply insensitive to the structural changes caused by the phospholipidosis. An alternative explanation, favored by the authors, was that although present at the dose levels and for the time period examined, the induced phospholipidosis was insufficient to interfere with normal function. This conclusion is supported by the lack of evidence of neural degeneration, apoptosis, or necrosis in any CNS or PNS tissue examined and is consistent with the reported lack of functional deficits in some instances of drug-induced phospholipidosis in other organ systems (Reasor and Kacew 2001).
As stated earlier, another important way that the findings from histopathology and electrophysiology can be dissociated is the fact that electrophysiologic responses can be altered by nonstructural or very subtle structural changes that are below the resolution of even the most sophisticated histopathologic procedures. Table 1 highlights several PNS electrophysiologic measures and their possible dissociation from structural evidence of neuropathology. Some of the most deadly environmental toxins (e.g., scorpion alpha-toxins, tetrodotoxin) are not associated with a structural alteration in neurons and have no measurable histopathologic effects. Generally, these toxins cause channelopathies, affecting the movement of Na+, K+, or Cl– ions across the neuronal membrane. The effects are often rapid and profound, and are easily measured by electrophysiologic end points such as conduction block or evidence of hyperexcitability. Channel-specific toxins have numerous mechanisms of action, including interference with the normal conformational changes required to close activated transmembrane channels with consequent out-of-control excitation, massive neurotransmitter release, spasm, and death (Spencer, Schaumburg, and Ludolph 2000). Toxin-induced alterations in channels may also include more subtle and chronic deficits such as alterations in the normal anchoring of intraaxonal ions at the region of the nodes of Ranvier (Waxman and Ritchie 1985). As the distribution of specific ions within the axon becomes less segregated, the efficiency of saltatory conduction is diminished, resulting in slowed nerve conduction velocity. Once again, these changes, which can clearly impact behavior and electrophysiology, are not detected by standard histopathologic procedures. Finally, some toxins have both structural and functional consequences that can be dissociated by time and exposure. For instance, the chemotherapeutic agent oxaliplatin has two forms of neurotoxic insult; initial effects include peri-oral paresthesia linked to acute hyperexcitability related to Na+ channels, while later effects are a sensory polyneuropathy due to the gradual accumulation of the heavy metal platinum in the dorsal root ganglia (Park et al. 2008). Histopathology is sensitive to the chronic, but not the acute, manifestation, whereas electrophysiology provides a sensitive measure of both forms of insult.
The effects of botulinum toxin illustrate another example of toxin-induced structural change at the molecular level which dramatically alters electrophysiology with little or no histopathology signal. Botulinum toxin is synthesized by bacteria of the genus Clostridium that, when activated, consists of a disulfide-linked heavy and light chain. When the heavy chain binds to an acceptor site on a neuron, the light chain enters the cell and cleaves one or more elements of the presynaptic vesicle docking complex, thus preventing the fusion of synaptic vesicles to the neuronal membrane (Dolly et al. 1984). Following exposure to even minute amounts of the toxin, neuromuscular transmission can be blocked, resulting in flaccid paralysis of the affected muscle. Changes in electrophysiology are rapid and severe (Arezzo 2009). The compound muscle action potential can be reduced to unmeasurable amplitudes within hours of exposure, with no change in histopathology findings. Eventually structural changes occur, but they represent indirect responses to the toxin-induced damage such as collateral sprouting.
Histopathology and electrophysiology findings are also dissociated for compound-induced seizures. Although some seizures have an underlying structural component, such as focal necrosis, most reflect a nonstructural imbalance of excitatory and inhibitory processes due to altered release of specific neurotransmitters or faulty neurocircuits (Niedermeyer 2005). When structural changes are detected, it is often not clear if the neuronal damage is the cause of the seizures or a result of the seizures. In vivo electrophysiology in the form of monitoring the pattern of EEG provides a sensitive biomarker for frank seizures that may occur without an associated convulsion (e.g., absence seizure) or for a compound-induced pattern of changes in brain waves that is consistent with lowered seizure thresholds (i.e., organized bursts of repetitive sharp waves). This nonstructural event is the dose-limiting toxicity for many drugs that target CNS function.
The recovery process is a final area in which histopathology and electrophysiology provide different views of neurotoxic insult. Most toxicology studies incorporate assessments at a fixed period following termination of exposure to the possible toxicant. There are numerous circumstances where both structural and functional findings continue to worsen for a period after the exposure to a neurotoxicant is terminated. This phenomenon, termed “coasting,” is most likely due to either damage set in motion during treatment or to time-sensitive insults in the elongated processes of the PNS. For instance, compounds that disrupt microtubule function (e.g., taxol) alter axonal transport in PNS axons, but the full consequences of depriving the distal most portions of the affected axons of needed materials is manifest over a protracted period (Swain and Arezzo 2008). In many studies, the lack of overt histologic evidence of damage at a posttreatment time point is heralded as evidence of “recovery.” In fact, this finding may simply mean that the active degenerative process is no longer ongoing and that the cleanup of neuronal and myelin debris has been completed. True “recovery” requires evidence of partial or complete return to normal function. This can best be documented by tracing longitudinal changes associated with the progression of the toxic damage during and immediately following exposure and then the reversal of the deficits during the recovery period. For example, the intramyelinic edema and vacuolation induced by a gamma-aminobutyric acid (GABA) transaminase inhibitor were associated with an approximate 20% slowing of conduction in the optic tracts during a 12-week treatment period in dogs. However, conduction returned to normal in the same cohort of animals following a 4-week period off compound (Schroeder et al. 1992).
Conclusions
In this review, we have focused on the circumstances in which findings from histopathology and electrophysiology can be dissociated. While it is important to recognize this possibility, in most circumstances there is excellent agreement between these measures. Histopathology is highly specific and can document toxicant-induced abnormalities in both cellular and subcellular structures (e.g., organelles). It can also provide insight into the relationship between neurons, glia, blood vessels, and inflammatory processes, which often holds the key to toxic insult. Electrophysiology provides an option for tracking longitudinal change in individual subjects, is sensitive to many forms of structural (i.e., axonopathies, myelinopathies, neuronopathies) and nonstructural (i.e., biochemical, molecular, or metabolic) deficits, and can explore activity at multiple levels of the elongated pathways forming the neuroaxis. The central tenet of this review is that neuropathology and electrophysiology are complementary procedures and that a complete view of neurotoxic insult often requires information from both procedures.
Footnotes
Acknowledgments
We want to thank Linda O’Donnell and Shirley Seto for their assistance in the preparation of this article.
This work was partially supported by NIH 1R21RR025896-01.