
Abstract
Diversity of living organisms and their environmental radiation exposure conditions represents a special challenge for non-human dosimetry. In order to contend with such diversity, the International Commission on Radiological Protection (ICRP) has: (a) set up points of reference by providing dose conversion coefficients (DCCs) for reference entities known as ‘Reference Animals and Plants’ (RAPs); and (b) used dosimetric models that pragmatically assume simple body shapes with uniform composition and density, homogeneous internal contamination, a limited set of idealised external radiation sources, and truncation of the radioactive decay chains. This pragmatic methodology has been further developed and extended systematically. Significant methodological changes include: a new extended approach for assessing doses of external exposure for terrestrial animals, transition to the contemporary ICRP radionuclide database, assessment-specific consideration of the contribution of radioactive progeny to dose coefficients of parent nuclides, and the use of generalised allometric relationships in the estimation of biokinetic or metabolic parameters. The new methodological developments resulted in a revision of the DCCs for RAPs. Tables of the dose coefficients have now been complemented by a web-based software tool, which can be used to calculate a user-specific DCC for an organism of arbitrary mass and shape, located at user-defined height above the ground, and for an arbitrary radionuclide and its radioactive progeny.
Keywords
1. Dosimetry for environmental radiological protection
Activities of the International Commission on Radiological Protection (ICRP) Committee 5 target topics related to environmental radiological protection, including establishment and justification of the environmental radiological protection system, its criteria, reference levels, and the basics of application practices. Justification of a methodology for assessing doses of radiation exposure of non-human biota appears in the list of topics as an important issue. Immense diversity of non-human life forms, their morphological and biological properties, behaviours, and life cycles creates a particular challenge for assessment of the environmental risks of radioactive exposures. Methods for assessing radiation doses are also challenged by the diversity of biota. This paper focusses on recent improvements in the methods, endorsed by ICRP, used to derive dose conversion coefficients (DCCs) for non-human biota, and discusses their application to practices of environmental dose assessments.
In an attempt to contend with biodiversity in assessment tasks related to dosimetry of various organisms, ICRP (2003a, 2008b) created points of reference by establishing a set of Reference Animals and Plants (RAPs), and by providing DCCs for these entities exposed to certain basic environmental radiation sources. In addition, the current approach of ICRP endorses the use of simplified, albeit plausible and robust, dosimetric models, which have been constructed to adequately reflect the most essential, from the dosimetric viewpoint, features of an organism or its environment, and appear to be the only practical means to contend with existing biodiversity.
Use of simple dosimetric models and approaches targeting absorbed dose averaged in the whole body is also compatible with the considered biological endpoints for radiological protection of non-human biota, which are defined by ICRP (2014) as ‘…those that could lead to changes in population size or structure’. Consequently, the biological endpoints can be regarded as biased towards early tissue reactions, which are generally expected at higher dose levels than those relevant to human radiological protection endpoints.
The current dosimetric approach of ICRP for non-human biota was outlined in Publication 108 (ICRP, 2008b; Ulanovsky and Pröhl, 2012), and is based on extensive experience accumulated over decades of radioecological and dosimetric studies. The approach has a strong link to, and stems from, the methodology and data developed in European-Union-supported projects (Larsson, 2004, 2008). The dosimetric data of Publication 108 were prepared using essentially the same methodology and computational tools as those incorporated in the ERICA assessment tool (Brown et al., 2008), and the radionuclide emission database of Publication 38 (ICRP, 1983).
Although extensive, the set of published DCCs for non-human biota has often been regarded as limited because of the restricted set of RAPs, or missing DCCs for certain radionuclides or particular exposure geometries. These limitations, often called ‘gaps’, and some internal inconsistencies were present in the dataset of Publication 108 (ICRP, 2008b) (e.g. few available sources for external exposure of terrestrial biota, or different ranges of accounted body masses of aquatic and terrestrial animals). Since issuance of Publication 108, new developments have provided possibilities to close some of these ‘gaps’, and to develop software tools that would enable one to go beyond the limits inherent in the printed tables of DCCs.
Over recent years, the activities of ICRP Task Group 74 have focussed on incorporating the new data and better methods into the current ICRP dosimetry system for non-human biota in order to update, extend, and harmonise the system, and to improve its self-consistency. The planned Task Group report is expected to present results of this activity systematically. The main changes in dosimetry for non-human biota resulting from this activity are described below.
2. Extending the ICRP dosimetry system for non-human biota
The current dosimetric framework for non-human biota appears to be highly compatible with the previously established framework (ICRP, 2008b; Ulanovsky and Pröhl, 2012). The existing methodology for assessing dose coefficients of internal and external exposure of aquatic organisms and internal exposure of terrestrial organisms uses a uniform isotropic model, and applies rescaling factors to obtain absorbed fractions for non-spherical bodies (Ulanovsky and Pröhl, 2006).
2.1. Revision and extension of DCCs for external exposure of terrestrial organisms
DCCs for external exposure of terrestrial organisms are based on results obtained by Monte Carlo simulation of photon radiation transport in terrestrial environments (Taranenko et al., 2004; Ulanovsky, 2014), so any contributions from α particles and electrons to the external dose of terrestrial organisms are neglected because of: (a) their short ranges in dense media; and (b) the fact that radiosensitive tissues of animals and plants are usually covered by inert layers (e.g. dead skin, fur, feather, shell or bark) and are beyond the reach of low-penetrating external radiations.
The dose coefficients for external exposure of terrestrial organisms in Publication 108 were based on the data of Taranenko et al. (2004), derived for three radiation sources in soil and for organisms on the ground surface with body mass ranging from 0.17 g to 550 kg (up to 6.6 kg for burrowing animals). External exposure of flying organisms (birds and insects) to sources in soil was modelled for organisms with body mass ranging from 35 g to 2 kg at heights not exceeding 10 m above the ground surface. The range of accounted body masses for terrestrial organisms was not in compliance with the accounted range of body masses for internal exposure and external exposure of aquatic organisms, which ranges from 1 mg to 1000 kg. Therefore, an obvious extension of the external dosimetry for terrestrial biota should improve harmonisation of the dosimetric approaches between aquatic and terrestrial species, and increase a number of radiation sources to address exposure situations of potential concern.
Extension of the dosimetric methods for external exposure of terrestrial organisms has been completed using the new approach, within which the whole-body doses arising from external exposure above the ground surface due to various photon sources in soil or in the ambient air can be estimated for arbitrary heights up to 500 m above the ground surface, and for any organisms with body masses from 1 mg to 1000 kg. Such flexibility became possible due to factorisation of the external dose into free-in-air kinetic energy released per unit mass (kerma) spectrum, and the energy-dependent ratio of whole-body absorbed dose and air kerma [see more details in Ulanovsky (2014)]. Correspondingly, each physical quantity could be calculated effectively by the Monte Carlo method using the ‘best available for selected condition’ technique, resulting in values that were computed accurately and systematically for the wide range of applicable parameters. Interpolation of the computed quantities allows derivation of dose coefficients for arbitrary masses in the range from 1 mg to 1000 kg, and for heights above the ground from 0.1 m to 500 m.
Due to the complexity and variability of the existing terrestrial life forms, all possible exposure conditions cannot be addressed. Therefore, generalised representative cases – as defined by source configuration and energy, contaminated media, organism sizes, and source/target relative locations – have been selected to model external exposure DCCs. DCCs for other exposure configurations can be inferred or deduced using interpolations between the available DCCs, or by superposition of responses from different basic sources. Currently, the following source-target scenarios are considered:
External exposure of animals and plants on and above the ground surface due to a planar radionuclide source at a depth of 0.5 g cm−2 in soil, which can be regarded as representative of radioactivity freshly deposited on the ground, accounting for surface roughness and initial migration. External exposure of animals and plants on and above the ground surface due to a uniformly contaminated volume radionuclide source with a thickness of 10 cm, which can be treated as representative of an aged contamination of soil following substantial downward migration and activity redistribution. External exposure of animals and plants on and above the ground surface due to an infinitely deep radioactive source in soil, which can be regarded as a source representative of naturally occurring radionuclides or anthropogenic contamination of the environment strongly affected by downward migration, agriculture, or decontamination practices. External exposure of animals and plants located on and above the ground surface due to immersion in air uniformly contaminated with radioactive materials. External exposure of in-soil RAPs that are situated in the middle of a uniformly contaminated volume radionuclide source with a thickness of 50 cm.
All of the above exposure scenarios, excluding the last scenario for in-soil exposure, have been systematically recalculated or newly calculated in order to allow interpolations of kerma spectra for source photon energy and height for the given source in soil, and absorbed dose per kerma for body mass and energy of photons incident on the body surface.
2.2. Effect of radioactive progeny on DCCs
Decay products of many radionuclides are themselves radioactive and may contribute to radiation exposure. Contribution of the radioactive progeny to radiation exposure of non-human biota is commonly attributed to the parent radionuclide, subject to various assumptions. The most common is a simple and pragmatic method when only a part of the full decay chain is taken into account under the assumption of full equilibrium between the parent radionuclide and its daughter radionuclides. Criteria for truncation of decay chains are commonly based on the selection of an upper limit for the half-life of a daughter to be accounted for in the given chain. Selection of the upper limit varies in the literature, and it can be found as 1 d (Amiro, 1997), 30 d (Jones, 2000; DOE, 2002), 180 d (Yu et al., 1993), and 100 y (Higley et al., 2003). An approach, adopted in Publication 108, used the value of 10 d as the chain cut-off criterion, following an approach introduced in the FASSET and ERICA projects (Larsson, 2004, 2008) and used in the ERICA tool (Brown et al., 2008). The ERICA tool and the current ICRP approach additionally truncate the chain at a daughter nuclide if its half-life is longer than that of the parent, because, in such case, the parent and the daughter would never reach equilibrium and, correspondingly, equilibrium activity ratios for such nuclides are indefinite.
Truncation of decay chains based on the criteria of maximum allowed half-life of a daughter is a simple and pragmatic solution when dealing with a limited set of radionuclides of immediate practical importance for a specific assessment. However, conditional on the assessment task, the chain truncation criteria may vary and need to be modified appropriately. Moreover, the development of software tools for DCC calculation, which use a comprehensive database of radionuclides, also requires the introduction of a uniform, accurate, and flexible method to account for the effect of radioactive progeny. Even more important, the method of cutting decay chains and assigning equilibrium activity ratios to accounted chain members implicitly assumes a single-thread linear decay chain, so this method may lead to ambiguous results when applied to complex, branching and merging, decay chains.
A more robust and flexible approach, capable of addressing various exposure scenarios (emergency, existing, or planned exposure situations), can be formulated using weighting factors for members of the decay chain based on their time-integrated activities. If accounting solely for radioactive decay (i.e. implying full absorption of radioactive substances in the organism in the case of internal exposure, or no migration of radioactivity deposited in the environment), one can express the dose as follows:
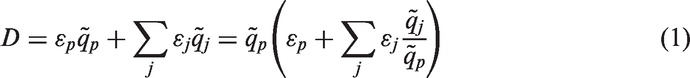
The implementation of this approach within software tools for DCC calculation can account for the effect of radioactive progeny for any type of radioactive decay chain by numerical integration of the differential equations describing the decay chain. Correspondingly, the integration time can be selected to be pertinent to the specific assessment task (so called ‘fit-for-purpose’); for example, based on life time and behaviour of the studied organisms, exposure conditions in their habitats, and existing temporal changes in radioactive contamination of the environment. An example of such a coherent approach can be found in Ulanovsky (2014), where DCCs for external exposure of terrestrial biota are computed for different averaging times conditional on the environmental source: 15 d for freshly deposited (planar) source in soil, 1 y for aged (10-cm-thick volume) source in soil, and infinite time (secular equilibrium ratios) for natural radionuclides uniformly distributed in the soil depth.
2.3. Transition to the contemporary database of radionuclides
Publication 107 (ICRP, 2008a) replaced Publication 38 (ICRP, 1983) with an updated and substantially extended database of radiation emitted due to decay of 1252 radioactive isotopes of 97 elements. Therefore, transition to the new radiation emission database appeared among the priority objectives of the Task Group. The new ICRP dataset of DCCs for non-human biota has been recalculated completely using this contemporary radionuclide emission database. The transition to the new database has also required extension of the set of photon and electron absorbed fractions up to 10 MeV to address maximum emitted energy for some newly included radionuclides. As with the previous data from Publication 38, numerical integration of continuous spectra of β electrons is used in the calculation of DCCs, thus accounting for the different range of electrons of various energies in the same emission spectrum. As a result of the transition, the updated internal DCCs are supplemented by fractions of dose due to four types of radiation, unlike the three types used in Publication 108, because of separation of high-linear-energy-transfer (LET) radiation into two groups: (a) fission fragments and α-recoil nuclei, and (b) α particles (see also Section 3.2).
2.4. Revised tables of dose coefficients and the software tool
Although the new updated DCC tables for RAPs are prepared for the same list of radionuclides and for the same decay chain criterion as in Publication 108, they were recalculated completely using the updated methods, as well as the new radiation emission dataset from Publication 107 (ICRP, 2008a). Other distinctive features of the new tables are their organisation and layout, which differ substantially from those used in Publication 108. The present tables are organised on a nuclide basis, making it easier to reveal interspecies and intersource variability of dose coefficients for the given radionuclide. In the majority of cases, such variability can be regarded as low or insignificant, indicating that DCCs are not among the main sources of uncertainty for an exposure scenario, and that an assessor has to pay attention to other, probably more significant, sources of uncertainty in the estimated doses.
In Publication 108, DCCs were given for adult forms of RAPs and for RAPs at various stages of development (eggs, mass, larvae, etc). The new tabulation only shows DCCs for adult forms of RAPs. The reasons for this decision are two-fold. First, variability of the dose coefficients for many radionuclides can be regarded as insignificant and ignored, or can be caught relatively easily by simple interpolation of the values shown for other RAPs. Second, the special purpose software tool BiotaDCC can be used to generate DCCs ‘on demand’ by means of a fully flexible user-friendly interface, accounting for non-standard (user-defined) organisms of arbitrary shapes and body masses, and under various exposure conditions, including different methods of accounting for radioactive progeny contribution. The software tool is planned to complement the printed tables of dose coefficients.
Some radionuclides are known to concentrate in certain organs of an organism (e.g. iodine isotopes in thyroids, bone-seeking actinides in skeleton, etc.), thus resulting in highly inhomogeneous dose distribution within the body. Under such circumstances, the average absorbed dose in the whole body may not reflect the actual risk of radiation exposure (e.g. Gómez-Ros et al., 2008; Caffrey et al., 2015). Still, internal doses in organs can be evaluated using known radionuclide activity in the organ and its mass, and simply replacing the body of the whole organism with another artificial ‘body’ with the size and mass of the organ of interest.
3. Environmental dose assessment
3.1. Assessment of internal environmental exposures
Within the current ICRP dosimetric methodology for non-human biota, doses of internal exposure are expressed as a product of the DCC and of average activity concentration of a radionuclide in the whole body. If the latter is unknown, it can be estimated from the known activity concentration in the environment using concentration ratios (CRs), which are the lumped factors describing transfer of radionuclides from the environment to the organism. Current estimates of CRs (ICRP, 2009) bear significant uncertainties because they are defined for chemical elements for stationary equilibrium conditions, thus ignoring non-stationarity of intake and retention of radioactivity in the body. Correspondingly, reduction of the uncertainties related to the lumped CRs has stronger potential to improve dose estimates than further improving the accuracy of the DCC alone.
Reduction of uncertainty related to transfer of radioactivity from the environment to the organism can be achieved via the use of radionuclide-specific CRs, which account for nuclide-specific radioactive decay properties, particularly for those radionuclides with a physical half-life comparable to their biological half-life. Additionally, allometric relationships can be used to interpolate vital morphological or biological properties between various types of species to quantify pathway-specific intake, and to describe retention of radioactivity in the body, thus resulting in more plausible estimates of CRs derived from simple (e.g. single-compartment) biokinetic models. As an example of biologically plausible interpolations, generalised allometric relationships are introduced and shown below for metabolic and inhalation rates of terrestrial mammals.
Allometric relationships are traditionally expressed (Rubner, 1883; Kleiber, 1947) as a power function of body mass:

Biological data, accumulated and analysed over recent decades (e.g. Marquet et al., 2005; Nagy, 2005; West and Brown, 2005; White and Seymour, 2005), provided sufficient information to demonstrate more complex relationships than those represented by Eq. (2) (e.g. Kolokotrones et al., 2010). Correspondingly, a generalisation of Eq. (2) can be suggested as follows:

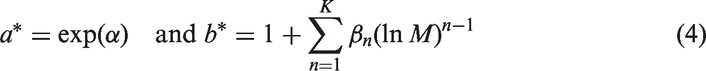
Examples are shown in Figs. 1 and 2, where the generalised Eq. (3) is used to approximate a specific basal metabolic rate of mammals derived from the data of Jones et al. (2009), and a specific ventilation rate of mammals based on the data of Bide et al. (2000).
Specific basal metabolic rate of mammals derived from the data of Jones et al. (2009) (dots), fit using generalised allometric Eq. (3), fit and prediction confidence intervals (CI) (lines), and reference data for humans (ICRP, 2002a). Specific ventilation rate of mammals (Bide et al., 2000) (dots), fit using generalised allometric Eq. (3), fit and prediction confidence intervals (CI) (lines), and reference data for humans (ICRP, 2002b).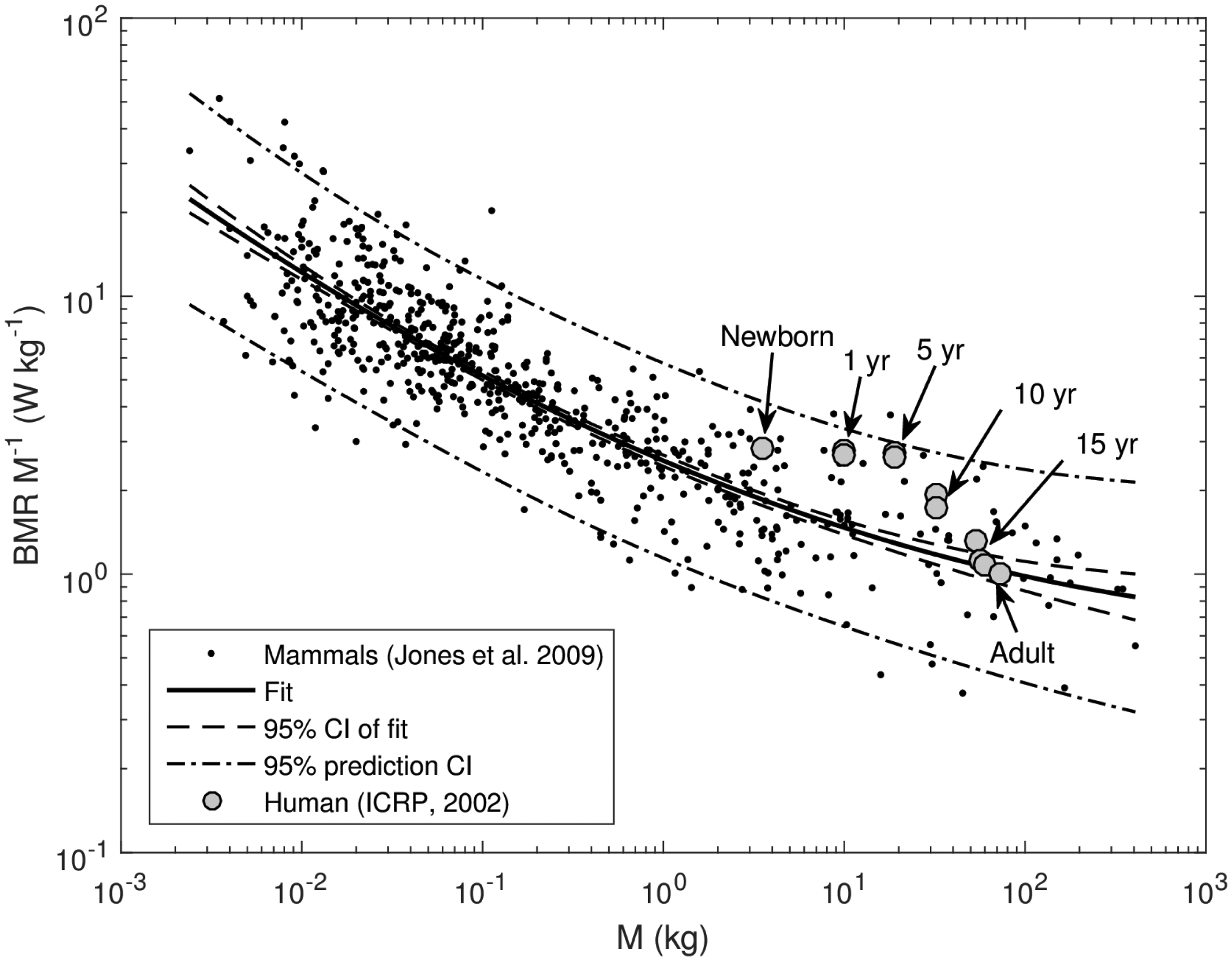
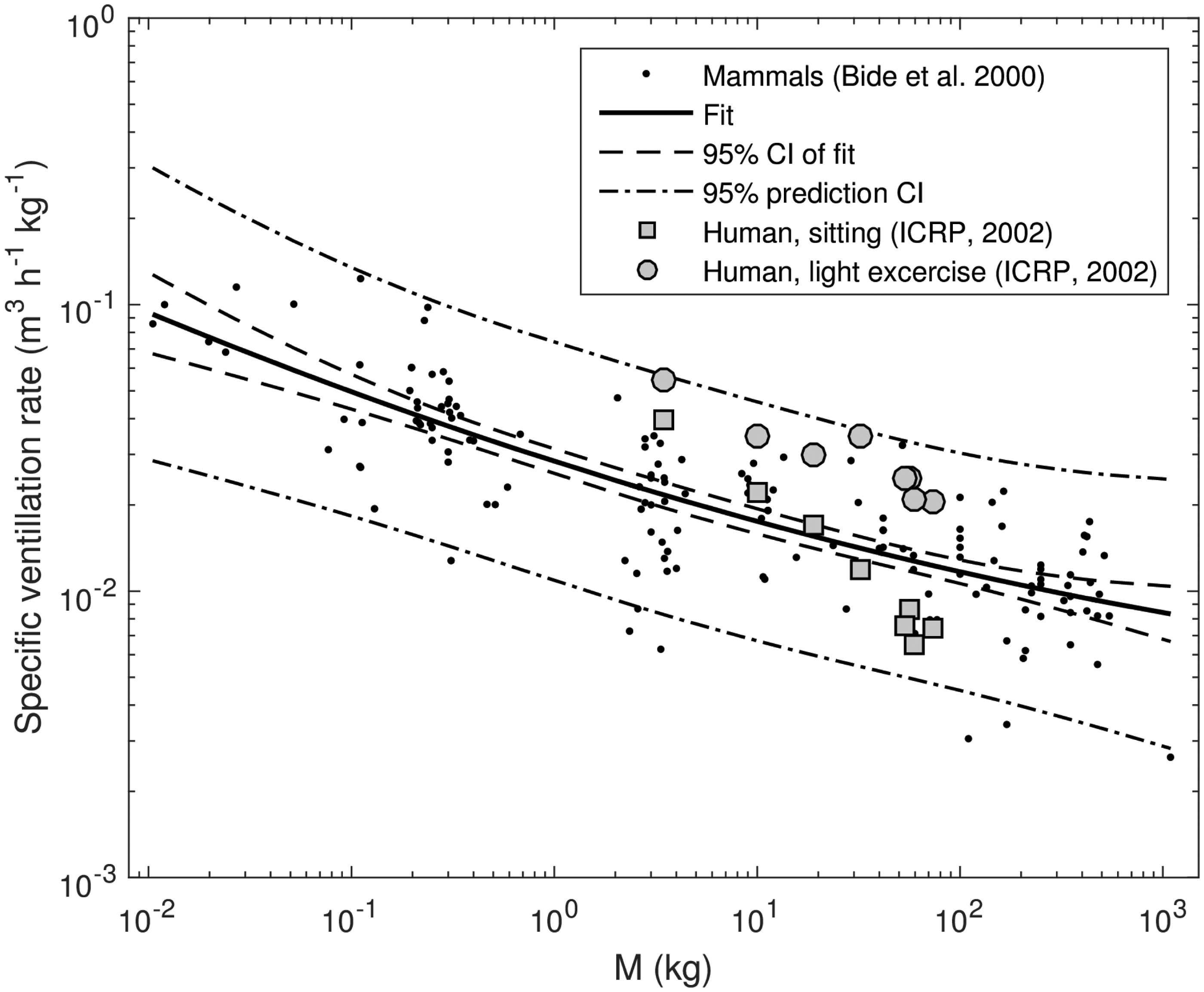
Figs. 1 and 2 show that deviations from the simple power law [Eq. (2)] are significant, and the data can be viewed as realisations of random processes with systematic parts described by the fit (solid lines) and stochastic components, which can be attributed to interspecies variability and are characterised by a geometric standard deviation of 1.47 (specific basal metabolic rate, Fig. 1) or 1.55 (specific ventilation rate, Fig. 2).
Also shown in Figs. 1 and 2 are corresponding age-dependent reference data for humans (ICRP, 2002a,b), which demonstrate good compliance with other mammals. The human data appear to show age dependence of basal metabolic and ventilation rates; however, as seen from the figures, this dependence is within uncertainty bands imposed by interspecies variability and, correspondingly, age effects can be treated as secondary and statistically insignificant when applied to allometric scaling for animals of different types.
3.2. Accounting for radiobiological effectiveness of radiation
Radiation effects on biota are known to depend not only on the absorbed dose, but also on the ‘quality’ of radiation, which in turn depends on the type and energy of the specific radiation. For example, α particles and other densely ionising radiations, such as heavy ions, are known to cause a stronger radiological effect on living tissue than that produced by β or γ radiation. This is known as the relative biological effectiveness (RBE) phenomenon, which can be expressed quantitatively via radiation quality factors based on LET in the tissue (ICRP, 2003b, 2007). For the purpose of human radiological protection, the concept of radiation weighting factors,
3.3. Uncertainties of external environmental exposures
Similar to internal exposures, in the case of non-human biota exposed to external radiation sources, uncertainties about DCCs due to interspecies variability can often be regarded as insignificant when compared with the uncertainties resulting from variability of environmental contamination and ‘source-target’ geometry for the biota populations of dosimetric interests. Even for aquatic organisms, for which exposure conditions can be approximated easily by simple exposure geometries within infinite homogeneous media, variability of radiation source strength in the environment can result in large variability of external doses among species of the studied population. For terrestrial organisms, due to diversity of landscapes, variability of radiation sources and exposure conditions can be much higher, thus making the external dose assessment for terrestrial species even more complicated. Doses of external exposure strongly depend on the organism’s life cycle, migration, and seasonal variations of behaviour. Currently adopted practices make use of a superposition principle by splitting a complicated external exposure scenario into a series of simple scenarios and summing their contributions, taking into account the time quota (occupancy factors) spent by the organism in the various locations.
Dealing with the uncertainties pertinent to external exposure of both aquatic and terrestrial biota can be achieved using probabilistic models, within which the uncertainties can be quantified and handled. However, use of probabilistic approaches requires collection of relevant data and statistical modelling. Modern developments in the field of global positioning system and remote tracking equipment open new possibilities to test and validate existing dosimetric approaches for free-ranging animals in highly heterogeneously contaminated environments (e.g. Hinton et al., 2015). Such studies can provide data to be used as a ‘benchmark’ for testing existing dosimetric models and dose assessment techniques; thus, the practical and scientific importance of such studies cannot be overestimated.
4. Conclusions
The recent activity of Task Group 74 regarding ‘more realistic dosimetry on non-human species’ resulted in substantially revised and updated dosimetric methodology for non-human biota, including tables and software to derive ‘fit-for-purpose’ dose coefficients for animals and plants, not limited by the ‘family’ of ICRP RAPs alone. The current dosimetry system for non-human biota was built assuming uniform distribution of radioactivity in a homogeneous skeleton-less body. Although in many practically relevant cases, these assumptions are plausible, and corresponding uncertainties in DCCs are much less than those for other parameters important for dose assessment, there may be situations when the non-uniformity of activity distribution in the body becomes essential for dose calculation. In such situations, some simple scaling techniques can still be used, as suggested in Publication 108.
Compared with Publication 108 and the ERICA tool (Brown et al., 2008), the current methodology for assessment of external exposure DCCs for terrestrial animals and plants has been systematically expanded and improved, and appears to be more harmonised and self-consistent. Use of the contemporary radionuclide database puts the new dosimetric methodology in line with other ICRP reports.
The new, radionuclide-based layout of the DCC tables facilitates quick interpolation of DCCs between species and sources of exposure. Additionally, ‘fit-for-purpose’ DCCs can be produced using the new software tool, which complements the report of Task Group 74 and is expected to appear as an open-access web-based application.
Besides DCCs, there are a number of other open issues that are influential to plausibility of environmental dose assessments. These issues include: (a) quantification of the radionuclide-specific CRs; (b) systematic consideration of uncertainties caused by variability (either of organisms or environmental contamination and transfer); (c) introducing and applying techniques of probabilistic modelling and assessments; (d) studying and accounting for dosimetric effects due to metabolism and biokinetic; (e) studying the biological effects of radiation on non-human biota in order to justify radiological protection endpoints and to quantify pertinent radiation weighting factors; and (f) development of appropriate statistical and information processing techniques to allow for effective analysis of large biological and environmental datasets for highly diverse biota and heterogeneous environments.