
Abstract
A significant body of evidence indicates that climate change is influencing many aspects of avian ecology. Yet, how climate change is affecting, and is expected to influence some aspects of the breeding ecology of cavity-nesting birds remains uncertain. To explore the potential linkage between timing of first clutch, and the influence of ambient temperature on hatching success, we used Eastern Bluebird (Sialia sialis) nest records over a nine-year period from Alabama, USA. We investigated changes to annual clutch initiation dates, as well as variability in hatching success associated with ambient air temperatures during the incubation period. Using a simple linear model, we observed earlier annual egg laying dates over the nine years of this study with a difference of 24 days between earliest egg-laying date of the season. Daily temperature minima increased 2 °C across the nine-year time frame of this study. These data also indicate that Eastern Bluebird hatching success was the highest when mean ambient air temperature during incubation was between 19 °C and 24 °C (78%, as opposed to 69% and 68% above and below this temperature range, respectively). Our findings of increasing maxima, earlier maxima each year, and the lower minima of temperatures within our study area could expand the breadth of temperatures experienced by nesting Eastern Bluebirds possibly exposing them to temperatures outside of what promotes nesting success. These findings with a cavity-nesting bird highlight an optimal range of ambient temperatures associated with highest hatching success, conditions likely to be affected by climate change.
Keywords
Introduction
Climate change is altering mean annual global air and ocean temperatures, increasing severe weather occurrence, making precipitation more erratic, increasing the mean global sea level, and altering many other regional natural phenomena.1–3 One of the most observable ways in which climate change affects plant and animal species is through altered phenology.4,5 Many living organisms use environmental conditions (e.g. temperature, precipitation, and photoperiod) as proximate cues to time key life history events (e.g. breeding, migration, flowering, fruiting, etc. 6 ). Moreover, we are already observing these phenological shifts in response to the changing climate in birds, invertebrates, amphibians, and plants.7–9 For birds, changes in the phenology of migration, 10 breeding dates, 11 and the distributions of species during their breeding and non-breeding seasons12–14 are all apparent.
Birds use environmental cues to time multiple parts of their life history such as breeding. 6 As such, earlier egg laying dates have been observed among species.15–17 Several studies have shown a link between avian hatching success and ambient temperatures.18–21 For insectivorous passerines, temperature can influence the availability of food during egg formation, as well as conditions for incubation. 22 For instance, Dunn et al. 23 found that the start of egg-laying in Tree Swallow (Tachycineta bicolor) correlated strongly with food abundance (flying insect biomass) but not with a peak in food supply, a peak which can occur during the nesting period.
Average annual temperatures are anticipated to increase by as much as 2.8 °C in areas of the United States by 2050. 24 These changes in ambient temperature are expected to affect avian populations as many species are not known to exhibit phenotypic plasticity in response to changing temperature. 25 Moreover, climate change can also result in fluctuations in temperature that are relatively short in duration, and breeding birds may, 26 or may not be able to adjust nest phenology in response to these erratic shifts. 27
With climate change already affecting bird species worldwide, it is important to understand what the potential impact of a warming and more erratic climate may be on avifauna. To this end, we hypothesize that the timing of avian egg-laying and hatching success will change alongside changing temperatures. Specifically, as temperatures warm, we anticipate clutches to be initiated earlier. We also expect that hatching success will be highest within a range of ambient temperatures, a range bounded by 10–15 °C on the low end. This low end corresponds with the minimum egg temperature between periods of incubation for various cavity-nesting species. 28 Conversely, the range of temperatures associated with optimal hatching success will be bounded by 24–27 °C on the higher end; the upper temperature range reflecting physiological zero, the temperature below which no embryonic development occurs. 29 Ambient temperatures approaching physiological zero may lead to increased nest attendance by adult birds with this attendance returning egg temperatures to below physiological zero, 30 as has been observed in Great Tit (Parus major). 27 Embryonic development is optimal when egg temperature is between 36 °C and 40 °C but mortality of the embryo is expected if temperatures exceed 41 °C. 31
To investigate relationships between temperature, egg laying, and hatching success, we used Eastern Bluebird (Sialia sialis) nesting records over a nine-year period to analyze changes in temperature within our study area, clutch initiation dates, as well as the hatching success of these nests over a range of temperatures. Our objective was to test if there was an observable shift in clutch initiation date, as well as any ambient temperature threshold above or below which we observe changes in hatching success.
Methods
Study area
Eastern Bluebird nesting was studied at the Wehle Forever Wild Tract (hereafter Wehle Tract) in Midway, Bullock County, Alabama, USA. This area is expected to experience an increase in average and extreme daily temperatures, as well as more erratic precipitation patterns caused by climate change. 32 The Wehle Tract is comprised of a mix of open pine uplands, mixed pine/hardwood creek bottoms, and a diverse herbaceous groundcover dominated by graminoids and forbs. The property is managed with regular use of prescribed fire on a two-year rotation during the late dormant to early-growing season (March), with fire applied to half the property alternating among years. Ninety-six nest boxes have been maintained on the property since 2011, with boxes distributed relatively evenly with equal presentation across the study area, and evenly between areas where prescribed fire is applied during rotation. All nest boxes are wooden, tall, and rectangular, with dimensions 51 × 33 × 20 cm. Boxes are attached to metal conduit ensconced in the ground with boxes affixed at ∼1.5 m above ground level. Most nest box poles are outfitted with a predator guard consisting of a metal, wrap-around disk ∼46 cm diameter.
Field methods
Contents of nest boxes were checked every 4–7 days. Nest checks were started by April 1 each year from 2012 to 2020, with the range of first checks from March 10 to April 1. Checks conducted earlier than April 1 were used to track the nest building period so that we could predict the timing of egg laying among nests. Data recorded included the approximate date of clutch initiation, the number of eggs in the nest, nest fate (if possible), and the number of eggs hatched. If a nest was found during the egg-laying process, we could obtain an exact first egg date based on one egg laid per day. 33 If the nest was not found during the egg laying process but had a full clutch and successfully hatched at least one chick, we back-dated the initiation date using an estimate of chick age (see Bluebird Nestling Day 1–21 Time Sequence Photos (sialis.org)). We assumed a 14-day incubation period, with incubation beginning on the last egg laid. From 2012 to 2020 we documented the exact day of clutch initiation for >55% of all nests (maximum = 64% nests [2014], minimum = 45% nests [2020]). We knew the day of nest initiation as Eastern Bluebird lay one egg each day. If, during our nest checks we found a nest with n = 1 + n eggs we subtracted the number of eggs from the known clutch size during last nest check. For example, if a nest was found to have 3 eggs during a check, but that nest was empty during the last check (four days prior), then we could determine the nest was initiated the day after the penultimate nest check. Nests not found during the laying process and subsequently abandoned before hatching were not included in our analysis of hatching success.
Ambient temperature collection
For temperature data we used the Climate Data Online database through the National Oceanic and Atmospheric Administration. 34 As no officially recognized weather stations were present at the study site until 2020, we obtained temperature data from four weather stations located near the Wehle Tract: Montgomery, Alabama (87 km to Wehle); Enterprise, Alabama (90 km to Wehle); Eufaula, Alabama (33 km to Wehle); and Columbus, Georgia (64 km to Wehle). Due to available weather station restrictions, only daily maximum and minimum temperatures were available from each station for our study period. We acknowledge these data may not precisely reflect fine differences in temperature across our study area. However, we believe that a more holistic measurement as afforded through these averages will enfold subtle differences typically missed in more localized measurements, such as differences due to various thermal properties associated with aspect and shading.
Statistical analyses
To evaluate changes in maximum and minimum ambient temperatures within our study area during the reproductive and incubation periods (March 1–July 31) during the years of study, we applied linear mixed-effect models. Through our analyses we used Julian date corresponding with March 1, for each year, as “1.” The dependent variable within these models was maximum, or minimum temperature while the independent variables were Julian date and year. The random effect for these models was the weather station at which the temperature data were collected. To reduce influence of temporal autocorrelation among temperatures across Julian dates, we subtracted each temperature data point from the next data point in the time series. 35 Our primary analysis of these data was performed using lmer through the lme4 package in R. 36 We then applied ANOVA with Satterthwaite approximation using the R package lmerTest to compare differences among our independent variables of Julian date and year.
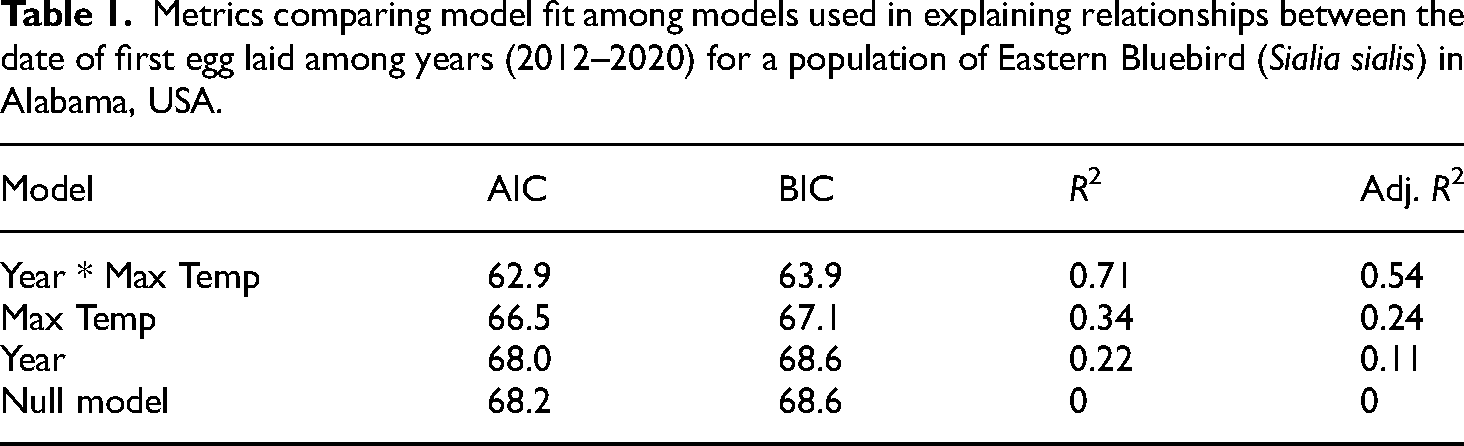
To investigate how date of clutch initiation has changed over a nine-year period, we selected the earliest Eastern Bluebird clutch initiation date per year. We chose to use the earliest clutch initiation date for each year as Eastern Bluebird are multi-brooded and delineation among nesting attempts can be difficult when considering the population as a whole. Also, the first egg date provides a clear demarcation we could tie to for our analysis. We performed a simple linear regression in the statistical software program R 36 using the clutch initiation date as the dependent variable and the year as the independent variable. We also tested relationships between the ambient temperature within the 24-h period when the first egg was determined to have been laid (given the temperature relationships detailed in Simmonds et al. 26 ), as well as an interaction between this ambient temperature and year. We compared among these models using a variety of metrics (eg, AIC, AICc, R2) to identify the best-fit model. This process was carried out using the function compare.performance in the performance package in R. 36
Through post-analysis, we tested the distribution of our response variable (clutch initiation date) for adherence to normality, using q-q plots of the residuals. We visually inspected each plot and found no clear deviation from normality. For these analyses we applied an alpha of 0.05 as a cutoff in identifying statistical significance.
We ran two separate models, each assessing changes in temperature relative to Julian date and year. The response variable in each model was temperature (°C). It is expected that temperature would increase from Mar 1 to Jul 1 within our study area each year. Since we were testing for a potential increase in maximum and minimum temperatures across our study period (Mar 1–Jul 31, 2012–2020), not simply Julian dates within years, we included a multiplicative relationship between Julian date and year. As temperature data were collected at four weather stations, we treated weather station as a random effect using the package lme4 37 in R. 36 We transformed data using Z-scores as values for the dependent variables in our models. We checked for the potentials of overdispersion and heteroscedasticity using q-q plots of model residuals. We calculated 95% confidence intervals around parameter estimates and considered results for our fixed effects that did not include zero as evidence of non-trivial association.
To evaluate the presence of discrete temperature changes outside of which hatching success differs, we ran a change point analysis using non-parametric binary segmentation from package changepoint. 38 We used daily maximum temperatures specific to the time of each nest used in our analysis for this evaluation. We excluded those records for nests deemed subject to alternate causes of nest failure (n, x% of all nests), such as: predation (n = 304, 19%), abandonment (n = 163, 10%), and unknown fate (n = 28, 2%). We calculated the hatching success of each remaining nest as the percentage of eggs laid that successfully hatched. Using the binary segmentation option, we defined the number of changepoints desired as two, allowing us to identify a range of ambient temperatures in which hatching success was significantly higher. Once change points were identified, we separated the observations into three bins: before the first change point (cooler temperatures), in-between the first and second change point (moderate temperatures), and after the second change point (warmer temperatures). We then ran a Kruskal–Wallis test of these data (Kruskal and Wallis 39 ) to test whether the differences in median hatching success between the change points were significantly different.
Results
Correlations in maximum and minimum temperatures among the four sites from which we used data revealed consistency among these sites (R2 = 0.98, for correlation among maximums and for correlations among minimums). Maximum ambient temperatures measured for our study area (2012–2020) showed an increase with Julian date within years (βyear 0.01, std. error < 0.01; F1, 93,425 = 6795.18, P < 0.01) but no change in the maximum temperature among years (F1, 18 = 1.31, P = 0.25). Minimum temperature measurements also increased with Julian date within years (t = 99.54, P < 0.01) and across the years of our study (βyear = 0.09, std. error = 0.04; t = 3.09, df = 1, P < 0.01) reflecting a trend of warming from 2012 to 2020 (F1, 129 = 9.62, P < 0.01).
The linear regression model of clutch initiation date among years showed a significant (R2 = 0.73, P = 0.01) negative relationship between the earliest clutch initiation date and year. The earliest clutch initiation date in 2013 was March 27th; in 2020 it was March 3rd, a difference of 24 days (Figure 1). The best-supported model, the model predicting the date of nest initiation among years (2012–2020), determined as the model with the lowest residual sums of squares, included the maximum temperature during the 24-h period when the first egg was laid, year, and an interaction between maximum temperature and year (Table 1). Results from this model showed that the mean Julian date of nesting for our study population became earlier across the span 2012–2020 (P < 0.01). Results from this model showed that the first eggs laid for the population became earlier across the years of study (t = −2.56, df = 1, P = 0.05), the maximum temperature within the 24-h period in which the first egg was laid decreased among the years of this study (t = −2.51, df = 1, P = 0.05) but showed a positive interaction between maximum temperature and year (t = 2.51, df = 1, P = 0.05).

Date of earliest egg found among a population of Eastern Bluebird (Sialia sialis) by year in Alabama, USA.
Metrics comparing model fit among models used in explaining relationships between the date of first egg laid among years (2012–2020) for a population of Eastern Bluebird (Sialia sialis) in Alabama, USA.
Mean clutch size in our study population that successfully fledged young was 4.45 eggs/nest. For hatching success, we used a total of 896 successful nests from the years 2012–2020. Two change points in hatching success were identified (Figure 2); the first occurred at approximately 19 °C and separated a mean hatching success of 69% at temperatures below the change point, and 78% at temperatures above the change point. The second change point occurred at approximately 24 °C and separated a mean hatching success of 78% at temperatures below the change point and 68% at temperatures above the change point. Hatching success exhibited a significant difference between temperature bins (separated by the change points) indicated by the change points during the incubation period (P < 0.01, df = 2). The moderate temperature bin showed significantly higher hatching success than both the cooler temperature (P < 0.01, df = 1) and the warmer temperature bins (P < 0.01, df = 1); however, there was no significant difference in hatching success (P = 0.7, df = 1) between the cooler temperature and warmer temperature bins.

Change point analysis plot showing the proportion of nests successfully hatched by maximum ambient temperature during the nesting period. The vertical black lines represent the change points identified in the analysis. The numbers represent the hatching success on either side of the change point, with the hatching success being highest during average incubation temperatures between the two change points. Gray points reflect individual nests used in this analysis.
Discussion
Evaluating changes in the date of first Eastern Bluebird nests within our study area, as well as nesting success for the period 2012–2020 showed a pattern of earlier date of first clutch initiated for the population during this nine-year period. The average maximum temperature measured in our study area did not differ among years. The minimum temperature during the period of our study however, was found to increase. The stationary maximum temperature but increasing minimum temperature measured likely decreases the breadth of temperatures that may be experienced by nesting Eastern Bluebirds possibly exposing these birds to a more narrow span of ambient temperatures outside of what promotes highest nesting success. We acknowledge that the varying distances (∼33–90 km) between our study site and the four weather stations where we obtained temperature data may be slightly more coarse in precision to evaluate specific change points affecting hatching success, but our findings of specific change points at 19 °C and 24 °C do align well with what has been published about thermal optima for avian incubation among other cavity-nesting species.28,29
The results of the linear regression show that Eastern Bluebirds on the Wehle Tract are exhibiting a trend towards earlier date of first clutch initiated annually which is consistent with previous research showing earlier nesting dates for many bird species in association with climate change.10,16,40,41 Herein it is important to distinguish that these studies evaluated shifts in nesting dates for the mean of populations, whereas we focused on the first clutch identified by year for our study population. Simmonds et al. 25 found earlier clutch initiation timing correlated with increased mean daily temperatures immediately prior to laying, which may explain earlier clutch initiation dates in Eastern Bluebirds in conjunction with increasing daily temperature. The date of first egg among nests was considerably earlier in 2012 and considerably later in 2019, relative to the trend among other years (Figure 1). After accounting for temporal autocorrelation among temperature data we did not find differences in maximum temperatures to suggest that 2012 and 2019 differed from other years within our dataset. Why the timing of first egg among Eastern Bluebird within our population differed from the general pattern in 2012 and 2019 provides basis for exploration beyond the current study.
One area of exploration that may be fruitful would be to evaluate how thermal conditions at relatively small spatial scales influence the timing of breeding within populations. While the temperature data used in our study showed strong correlation among the sites where these data were collected our approach does not account for differences that can occur at smaller spatial scales such as among nest boxes within the population. We also did not account for extremes in extreme temperatures that can occur over short periods, conditions that may become more commonplace within the scope of predictions in climate change. 42 Small spatial and temporal scale differences may influence the timing of breeding within populations as well as within-species responses to climate change. 43 Conversely, conditions such as the North Atlantic Oscillation (NAO) can influence climate over broad areas across the globe. 44 The NAO Index indicated negative pressure conditions during 2012, 45 a year in which the first Eastern Bluebird egg laid within our study population was earlier than expected (Figure 1). The NAO Index was high in 2019, a year in which the first egg in our population was later than anticipated. How small- and large-scale drivers relate to differences in the timing of reproduction and the ultimate success of that reproduction differ relative to the ecological processes considered. 46
Our change point analysis provides evidence that hatching success in Eastern Bluebirds is affected by ambient temperatures during incubation, with decreased hatching success during more extreme hot and cold temperatures. This finding parallels others in showing that environmental temperatures during nesting may have different influences depending on the species of focus,47,48 with Halupka et al. 49 providing evidence that rising temperatures will mainly affect migratory and large-bodied birds.
Microclimates within nest cavities can provide a stable environment for altricial birds during their development. 50 The thermal tolerance of avian embryos is estimated at 41 °C, as a maximum, for periods less than a few hours. 31 This tolerance is much greater than the high-end optima (24 °C) we’ve identified in our study. Nord and Nilsson 51 found that increasing ambient temperatures by as little as 1.5 °C during incubation can hasten embryonic development. Results from our study did not provide evidence that maximum ambient temperatures increased among the years of study (2012–2020). We did find, however, that the minimum ambient temperature increased. The static maximum ambient temperature among years, but increasing minimum ambient temperature reduces the range that nesting Eastern Bluebirds, and their nests are exposed. Our finding of a range of optima for ambient temperature 19–24 °C does not account for various factors such as parental involvement in regulating temperature within the nest cavity, which can likely modulate temperatures within nest boxes in most cases.52–54
Our analyses reveal that nesting success is tied to ambient temperatures. Consequently, we might expect to see a shift in the timing of the nesting period to coincide with those temperatures that provide the highest hatching rates, which is reflected in the nesting dates becoming earlier each year. If Eastern Bluebirds are unable to shift the timing of their egg production accordingly, then hatching success may decrease for at least some portion of the population. Harrod and Rolland 55 found the variability in maximum temperatures during incubation negatively affected nest success in Eastern Bluebirds. While we did not evaluate ambient temperature fluctuations as a factor affecting hatching success in our population, temperature variability could have been influential to reproductive success within this population and others.
Shifts in egg laying dates may alter nestling survival in accordance with the “mismatch hypothesis.” 56 Visser et al. 56 concluded that climate change may affect the hatching synchrony between birds and their food resources, resulting in hatching no longer coinciding with periods of peak food abundance. This shift in the earliest clutch initiation date for Eastern Bluebirds may be a response to earlier periods of peak food abundance, or it may be independent of food resource timing, which would ultimately lead to a mismatch in the period of nesting and the most abundant food resources.
Given our findings that Eastern Bluebird nesting ecology is influenced by ambient temperature, we expect other cavity nesting birds may also be affected by increasing average temperatures.
Conclusion
In summary, we observed Eastern Bluebird within our study population in Alabama, USA experienced earlier annual egg laying dates over the nine years of this study, a difference of 24 days between earliest egg-laying date of the season. The eggs of these birds had the highest hatching success when mean ambient air temperature during incubation was between 19 and 24 °C. These differences will likely influence the reproductive timing and success of these birds, as well as other cavity-nesting passerines as changes in climate occur.
Footnotes
Acknowledgments
Appreciation is extended to numerous field technicians and volunteers who routinely performed nest box monitoring and data entry at the Wehle Tract and include A. Arnold, W. Price, T. Waldon, T. Rush, D. Duffie, C. Wood Megelsh, H. Wright, H. Holiman, A. Patterson, R. Paiva, and N. Edwards. Equipment and funding for this study was provided by the Wehle Land Conservation Center of the Alabama State Lands Division and in part by the Robert G. Wehle Charitable Trust and McIntire-Stennis project under accession number MISZ-082100.
Author contributions
Spencer Weitzel: conceptualization (supporting); investigation (supporting); visualization (supporting); writing-original draft (equal); writing-review & editing (equal). Scott Rush: conceptualization (lead); methodology (lead); supervision (supporting); investigation (lead), visualization (lead); writing-review & editing (lead). John Trent: data curation (lead); investigation (supporting); resources (supporting); supervision (supporting). Eric Soehren: resources (lead); supervision (lead); conceptualization (supporting); investigation (supporting); visualization (supporting).
Data availability
Data collected and used in supporting the findings of this study are available on request from the corresponding author.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
All elements of this study adhered to ethical guidelines approved by Mississippi State University Animal Care and Use Committee. Permits were provided by U.S. Geological Survey, Bird Banding Laboratory (Permit 23424) and Alabama Department of Conservation and Natural Resources.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Author biographies
Scott A Rush is an Associate Professor in the Department of Wildlife, Fisheries and Aquaculture at Mississippi State University. Through his current work Scott strives to understand the function and resiliency of ecological communities especially within the context of human-wildlife conflicts. Scott received his PhD from the University of Georgia (2009). His post-doctoral training, completed at The University of Windsor's (Windsor, Ontario) Great Lakes' Institute for Environmental Research, addressed changes in energy mobility through Great Lakes ecosystems. Scott and his students continue to work within an expanding research framework addressing landscape and trophic ecology relative to wildlife populations.
Spencer L Weitzel holds a BS in Environmental Science from the University of Nevada, Reno and a MSc in Wildlife, Fisheries and Aquaculture Science from Mississippi State University. He currently works for the US Fish and Wildlife Service in Arkansas working to restore ecosystem function via habitat restoration with a focus on conserving rare habitat types and benefitting threatened and endangered species.
John A Trent is a Wildlife Biologist with the Alabama Department of Conservation and Natural Resources State Lands Division. He received his BS from the University of Tennessee at Knoxville in Wildlife and Fisheries Science. Since 2007 he has worked throughout the state of Alabama on a variety of wildlife inventory and monitoring projects designed to better our understanding of the state's rich biodiversity.
Eric C Soehren is an ecologist with the Alabama Department of Conservation and Natural Resources (ADCNR), State Lands Division and currently manages the Wehle Land Conservation Center in Midway, Alabama, a state-owned facility that provides public outdoor recreational opportunities, coordinated educational programs, and fosters wildlife monitoring and research initiatives on the surrounding Wehle Forever Wild Land Trust lands. Eric received his BS and MSc degrees in Biology from Jacksonville State University and has been employed by ADCNR since 1997. Eric's research and management interests are broad and varied, but those that have applied outcomes benefiting and enhancing the conservation and function of natural communities and their associated biodiversity are most important to him.