
Abstract
As adaptive systems, kinship and its accompanying rules have co-evolved with elements of complex societies, including wealth inheritance, subsistence, and power relations. Here we consider an aspect of kinship evolution in the Austronesian dispersal that began from about 5500 BP in Taiwan, reaching Melanesia about 3200 BP, and dispersing into Micronesia by 1500 BP. Previous, foundational work has used phylogenetic comparative methods and ethnolinguistic information to infer matrilocal residence in proto-Austronesian societies. Here we apply Bayesian phylogenetic analyses to a data set on Austronesian societies that combines existing data on marital residence systems with a new set of ethnographic data, introduced here, on initiation rites. Transition likelihoods between cultural-trait combinations were modeled on an ensemble of 1000 possible Austronesian language trees, using Reversible Jump Markov Chain Monte Carlo (RJ-MCMC) simulations. Compared against a baseline phylogenetic model of independent evolution, a phylogenetic model of correlated evolution between female and male initiation rites is substantially more likely (log Bayes factor: 17.9). This indicates, over the generations of Austronesian dispersal, initiation rites were culturally stable when both female and male rites were in the same state (both present or both absent), yet relatively unstable for female-only rites. The results indicate correlated phylogeographic evolution of cultural initiation rites in the prehistoric dispersal of Austronesian societies across the Pacific. Once acquired, male initiation rites were more resilient than female-only rites among Austronesian societies.
Introduction
Prehistoric kinship systems, flexible and adaptive through time1–4 and integral to the organization and evolution of early complex societies,4,5–9 exhibit both ancestral and adaptive dynamics in correspondence with prehistoric human dispersals.7,10–13 As a result, certain general tendencies seem to have facilitated, though not determined, directional changes in post-marital residence system. 11 In prehistoric Indo-European and Bantu African dispersals, patriliny appears to have facilitated the origins of heritable wealth inequality.14–21 In Mainland and Island Southeast Asia, however, kinship systems, subsistence systems, and socio-economic inequality appear to have been spatially and temporally heterogeneous,3,22–26 even after demographic expansions of rice farmers from southern China.27–36 Additionally, post-marital residence systems in prehistoric Austronesian societies appear to have been more fluid, potentially transitioning back and forth between states on a time scale of centuries.4,24 Cultural phylogenetic analyses 4 indicate that patrilocality and matrilocality, respectively, are the first and second most common (likely) states in the Austronesian tree.
Recent genetic evidence also suggests that the coasts of Southeast Asia were an avenue for regional interaction. 37 In coastal societies, prolonged male absence for purposes of fishing, trade or long-distance voyaging may favor matrilocal residence and matrilineal descent, as women manage the interests of the kin group.14,23,24 This has been suggested as an underlying contributor to matrilineal systems in Island Southeast Asia. As Hage and Marck 23 point out, “In the Micronesian and larger Oceanic context, if husbands and fathers were lost (always a possibility in seafaring) they could be readily replaced by other men” (p. S123).
In Island Southeast Asia, genetic and linguistic evidence suggests the initial Austronesian dispersal across the Western Pacific Ocean originated with matrilineal groups.23,24 Under the “Austric” hypothesis,38,39 Taiwan was colonized ca. 6500–6000 BP by proto-Austronesian speakers from mainland China, such that Taiwan is the probable homeland of the Austronesian dispersal into Island Southeast Asia and Oceania.36,38 A maritime dispersal began ca. 5500 BP out of Taiwan, as Austronesian-speaking peoples sailed along the Fujian coast, hunting, fishing, growing rice and millet, producing pottery, and building villages of stilt houses. 38 From the Philippines, Austronesian populations likely mixed with Australo–Melanesian populations of Melanesia about 3200 BP, before dispersing into Micronesia by 1500 BP.36,40,41
This sequence is supported by a phylogeny of 400 Austronesian languages that yields an estimated root age of 5800–4750 years BP 41 and has been the basis for a sequence of multi-trait Bayesian phylogenetic analyses. Jordan et al. 24 combined the tree with ethnographic data on marital residence to conclude that “early Austronesian societies were predominantly matrilocal, and that patrilocality was a later development in some Formosan (Taiwan) societies and the Austronesian family as a whole” (p. 1960). Combining the language phylogeny with a dataset of sociopolitical organization in 84 Austronesian societies, Currie and colleagues used similar computational phylogenetic methods to conclude that complexity rose and fell in small steps over time rather than in large jumps.42,43 These data representing political hierarchy, combined with data on human-sacrifice rituals, showed that rituals preceded, and thus facilitated, the increase in political hierarchy in Austronesian societies. 44
If initiation rites (e.g. Lutkehaus, 45 Schlegel and Barry 46 ) co-evolve with socioeconomic complexity, then they probably also co-evolve with postmarital residence systems. To explore this possibility, we compiled a new set of ethnographic data on initiation rites in Austronesian societies (see Supplemental Material) and used cultural phylogenetic analysis to infer the temporal ordering and phylogeographic structure of initiation rites in Austronesian societies.
Methods
Before phylogenetic analysis, we simply observe the geographic patterning in the data, first through simple contingency statistics and then through multiple factor analysis (MFA), looking at correlations between and among variables such as subsistence and initiation rites. The data we used for MFA (see Supplemental Material) has 84 rows, each representing a different Austronesian society. Each society is characterized by three categorical variables—subsistence, cultivation, and crop—followed by five ordinal variables—gathering, hunting, fishing, animal husbandry, and agriculture—that represent how reliant a society is on these different forms of subsistence. Next, the correspondences we observe set up “Galton’s problem” 47 as to whether the observed correlations are the result of common ancestry or convergent evolution.
Following previous studies,24,43,44,48,49 our proxy for inferring population history is the Austronesian language tree, 50 on which transitions between initiation-rite states are modeled along the phylogeny. For comparative purposes, we used the same 84 societies that Currie et al. 43 used to examine the rise and fall of political complexity across Austronesia. Data on initiations were extracted from Murdock’s Ethnographic Atlas 51 as well as from numerous ethnographies (see Supplemental Material). No initiation-rite data could be found for five societies. We coded each of the 79 remaining societies as being in one of four possible initiation states: (i) male-only initiation rites, (ii) female-only initiation rites, (iii) both male and female initiation rites, and (iv) no initiation rites.
To address this issue, we used the 79 societies for which we have data on initiation rites as phylogenetic characters in order to infer initiation rites in ancestral Austronesian societies. We used BayesTraits 52 to infer the likely transition rates among the states. First, we make a simple test of correlated, as opposed to independent, evolution between male and female initiation rites. Next, following previous studies,4,43 we used Reversible Jump Markov Chain Monte Carlo (RJ-MCMC) simulations on an ensemble of 1000 possible Austronesian language trees, with gamma hyper-priors for model parameters and exponential hyper-priors for the reverse-jump model selection.
Using RJ-MCMC allows us, first, to derive the transition likelihoods between states and, second, to determine the level of evidence for different directions of transition.4,52 To ensure that our runs of RJ-MCMC were consistent, we calculated the marginal likelihood of each run using the stepping-stone sampler, with 100 “stones” and 1000 iterations per stone. 52 By reducing over-parameterization on many iterations, RJ-MCMC makes parameter estimates more precise.43,52
Starting with a randomly drawn tree, MCMC explores the possible parameter space, landing more frequently on high-likelihood transition rates. The likelihood of a given parameter combination is evaluated by comparing the simulation output to the known initiation rites in the 79 societies. To avoid autocorrelation, we sampled only 1%, or 10,000, of the 1 million iterations. For each iteration, the reverse-jump (RJ) part of the MCMC selects a reduced-form transition model that constrains certain transition rates. For example, a reduced transition model might constrain the “female-only” to “no initiation rights” transition to be equal to zero. The reduced-form model that appears in the most MCMC samples is the most likely transition model.
The expected transition rates among the states (12 possible) are the mean of the RJ-MCMC samples. Here we report the probability that each of our four states was ancestral in Austronesia. In addition, we use the RJ-MCMC output to report which is the most likely constrained transition model—the one that appears most frequently in the MCMC samples—as well as the most likely ancestral state. From these likelihoods, Bayes factors (where the likelihoods are significantly different) were calculated by comparing the fit of the original rate matrix with rate matrices estimated from all of the other datasets in pairwise fashion. The logarithm of Bayes Factor is simply twice the difference between logarithms of the marginal likelihoods of complex versus simple models, respectively. 52
Results
Before addressing ancestral phylogenetic relationships, it is instructive first to observe contemporary patterning in the data. Figure 1 maps the Austronesian ethnolinguistic groups, 43 color-coded by their type of initiation rite. The most striking aspect of the distribution is that the 11 societies of Taiwan and the Philippines (numbered 9–19 in Figure 1) are all without initiation rites. Given that Taiwan is the probable ancestral homeland of the Austronesian dispersal,36,38,40 a reasonable hypothesis is that proto-Austronesian society lacked initiation rites. We test this hypothesis with the phylogenetic analysis, below.
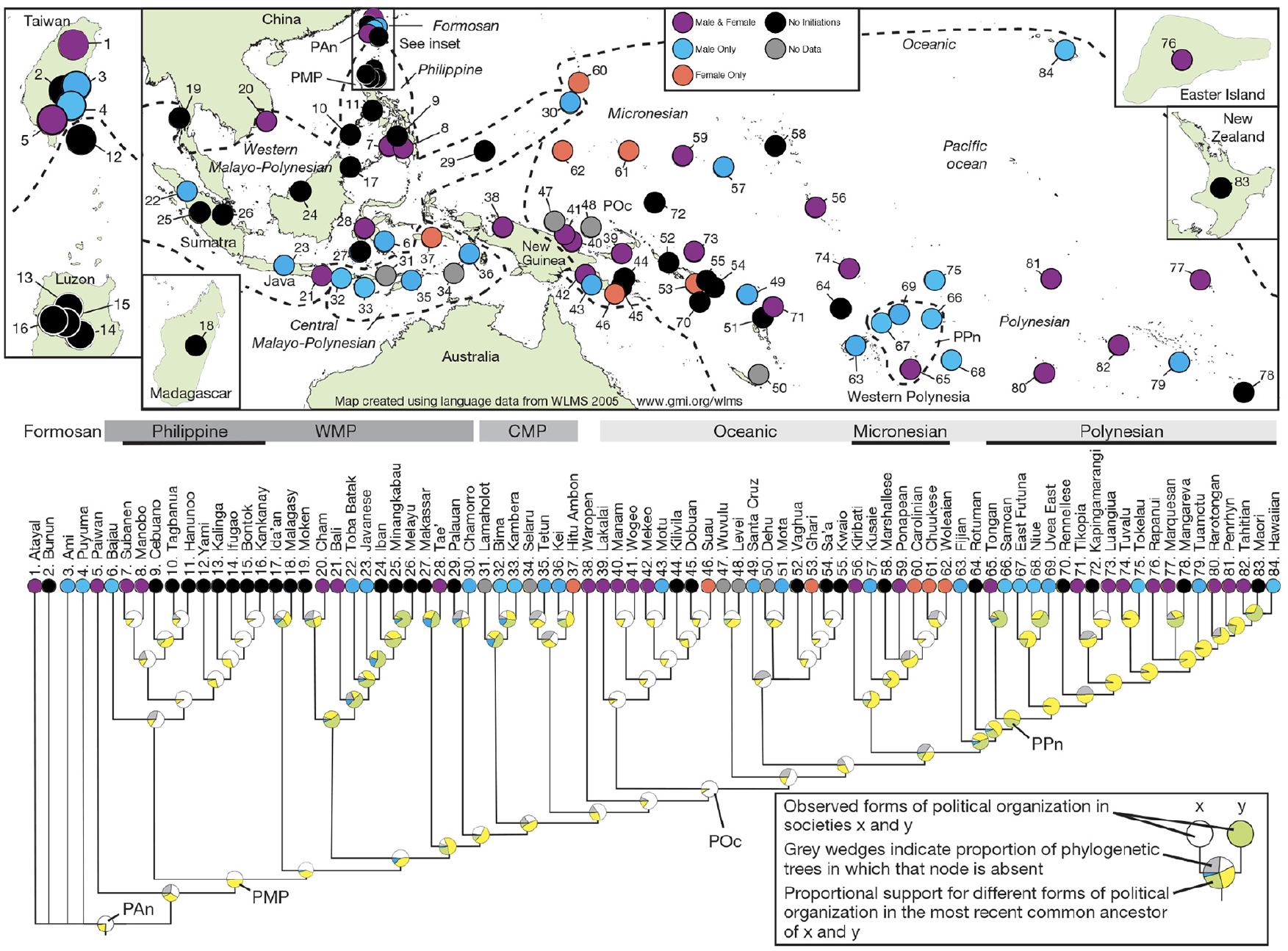
Map of Austronesian ethnolinguistic groups (after Currie et al. 43 ), color-coded by type of initiation rites: blue, male only; red, female only; purple, both female and male; black, no initiation rites; gray, no data.
Another notable, initial observation is the relative rarity of female-only initiation rites: there are only six, five of which occur in Western Micronesia (one of which is missing data on marital residence). Further, when these are placed on the Austronesian language tree (Figure 1), there is but one instance where female initiation rites occupy adjacent branch tips: the three Micronesian societies Carolinian, Chuukese, and Woleaian.
Next, we simply compare how initiation rites correspond to kinship systems, using available data.24,50 Treating Table 1 as a contingency table, the relation between kinship system and initiation rite yields p = 0.084 (χ2 = 11.1, d.f. = 6). Regardless of how one interprets this p value and the small numbers in the contingency table, we note that 55% of matrilocal societies have no initiation rite, compared to 30% of patrilocal or ambilocal societies.
Ancestral initiation rites versus kinship systems for the 76 Austronesian societies with data on both.

This pattern of matrilocal societies having more female and/or no initiation rituals at all is more pronounced in the eastern part of Austronesian dispersal. Using the geography of Currie and colleagues, 43 we divided the societies into two parts: Western (Formosan, Philippine, Western Malayo–Polynesian, and Central Malayo–Polynesian) and Eastern (Oceanic, Micronesian, and Polynesian). In the corresponding contingency tables (Tables 2 and 3), there is a co-occurrence pattern between marital residence and initiation rite in the Eastern group (p < 0.03, χ2 = 13.77) but not in the Western group (p = 0.57, χ2 = 2.96).
Ancestral initiation rites versus kinship systems in Western Austronesian societies.

Ancestral initiation rites versus kinship systems in Eastern Austronesian societies.

The MFA results (Figure 2) indicate that dimension 1, which explains almost 20% of the variance in the data (full data in Supplemental Material), is loaded primarily on the subsistence variables (Figure 2, upper right). Because marine subsistence increases in importance moving from Taiwan out toward Oceania, dimension 1 also correlates with geography (Figure 2, upper left). Dimension 2, which accounts for almost 14% of the variance, has loadings that separate kinship systems (Figure 2, lower left) but not subsistence systems. Note that, consistent with previous studies, 24 societies with matrilocal residence plot close to the center of Western Malayo–Polynesia, Formosa (Taiwan), and the Philippines (Figure 2, lower right).

Bi-variate plot of the first two dimensions of multiple factor analysis. Three categorical variables (subsistence, cultivation, and crop) and five ordinal variables (gathering, hunting, fishing, animal husbandry, and agriculture) were used to determine how reliant a society is on different forms of subsistence, from low to high. In the four panels of the figure, the results are then plotted with each society grouped according to region, subsistence, initiation rite, kinship system.
Without yet addressing ancestral relationships, the MFA suggests that female-only initiation rites are distant (in variable space rather than phylogenetic space) from the other kinds of initiation rites. In addition, female initiation rites tend to be in Micronesia, whereas a lack of initiation rites tends to predominate in the Formosa/Philippines region. It appears that over the course of the Austronesian dispersal, the trend was from intensified-agriculture, no initiations, and matrilocal residence in the Austronesian homeland to more fishing and higher incidence of female initiation rites out in Micronesia and Oceania.
Although residence pattern and initiation rites both load on the same MFA component, this correlation does not necessarily mean they co-evolved. To test this, we used the correlated-evolution test. Through the Bayesian MCMC procedure, we inferred the most likely evolutionary transition rates among the four possible initiation rites, as well as the probability of each initiation rite being the root ancestral state.
We start with a simple phylogenetic test 52 of (in)dependence in evolution of female versus male initiation rites. The independent model assumes that female and male initiations appeared and disappeared independently of each other. The dependent model—with four possible combinations (0,0; F,0; F,M; 0,M) and eight possible single-state transitions—assumes that the rate of change in female and male initiations were dependent on each other. From 50 runs of the independent discrete model (Table 4), we obtained a marginal likelihood of −125.11. From 50 runs of the dependent discrete model (Table 5), the marginal likelihood is −116.15. The log Bayes factor (=2×(–116.15 – 125.11) between the dependent and independent models is 17.9, enough to justify the more complicated model 52 of correlated evolution between female and male initiation rites. The two highest rates in this model are transitions away from female-only rites (F,0), whereas transitions from both rites (F,M) or from no rites (0,0) have the lowest rates (Table 5). In other words, the dependent model in Table 5, which is justified by the Bayes factor, indicates that the initiation rites prefer to be in the same state, that is, relatively stable when both female and male rites are in the same state (both present or both absent) but relatively unstable for female-only rites. The dependent model was ambivalent about the initiation rites for the root, that is, proto-Austronesian society (identical 25% probability for female, male, both, or none).
Average of 50 runs of the independent, discrete model. The average log likelihood of these runs was −125.11 ± 0.08.
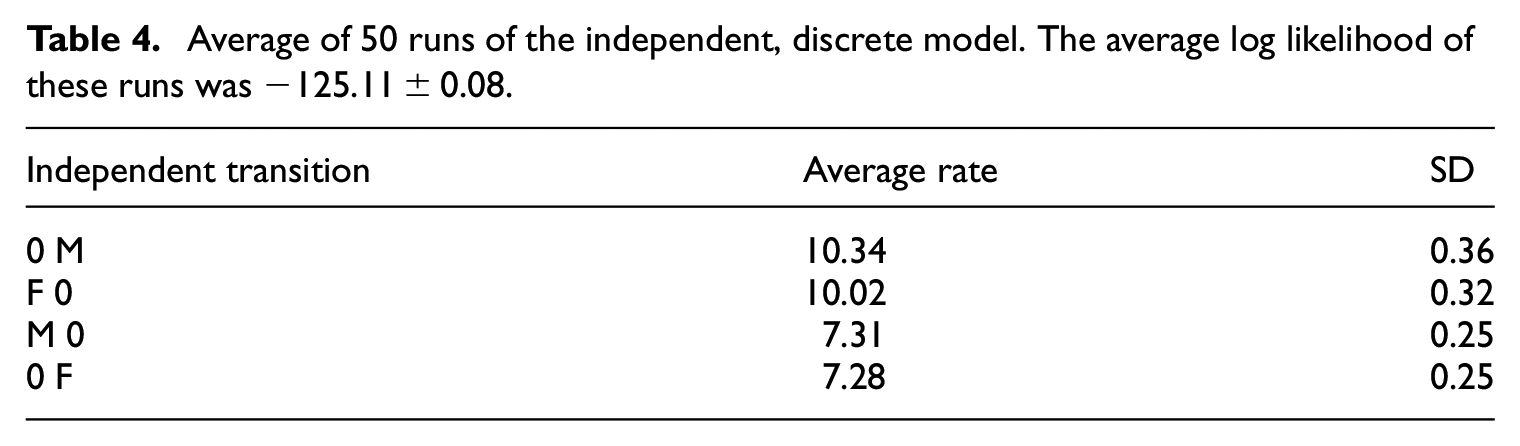
Average of 50 runs of the dependent, discrete model of correlated evolution. The combined states are represented\newline by presence/absence female rite first, male rite second. The average log likelihood of these runs was −116.15 ± 0.09.
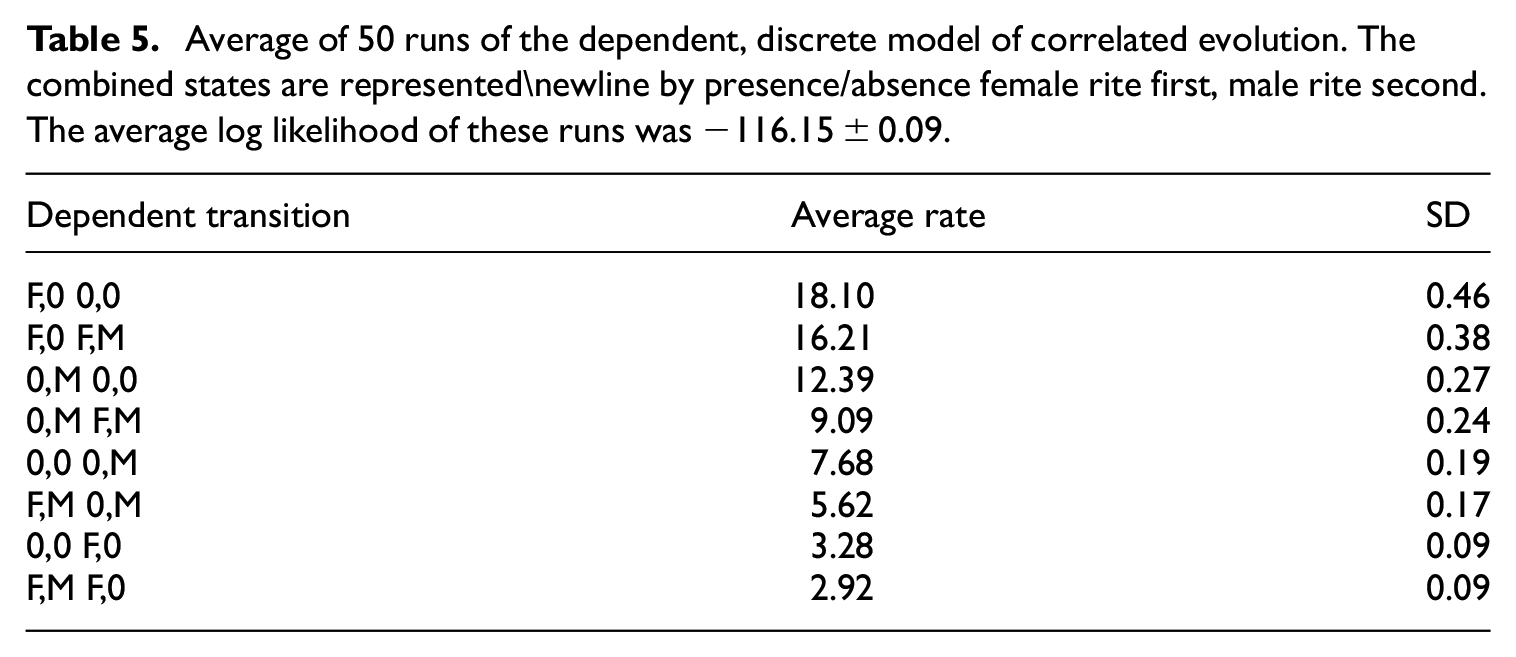
Next, we move to the RJ-MCMC approach, which provides a means of selecting constrained-transition models from the space of possible models, without over-parameterization. Figure 3(a) shows the average transition rates sampled over all selected models. It shows that transitions toward female-only initiation rites are the least likely. We find a similar pattern from the RJ-MCMC, which samples constrained-transition models from the space of possible models. In the most likely constrained-transition model (about 5% of the RJ-MCMC samples), transitions toward female initiation rites never occur (Figure 3(b)). Male-only rites appear stable, arising from the male-and-female-rites state (Figure 3(b)).

Transition rates between initiation rites (weighted black arrows) and percent probabilities for each rite being the ancestral root (red shade): (a) expected values over all MCMC samples and (b) expected values for the most likely transition model (Bayes factor of 17,149) determined by reverse-jump MCMC.
The female model in Figure 3(b) has a high Bayes factor of over 17,000, which is strong justification for the model being the best of the models we have explored. The calculation is 43 : posterior odds = 30/(960 − 30); prior odds = 52/(2,76,44,436 − 52); Bayes factor = (posterior odds)/(prior odds) = 0.32/0.00000188 = 17,149. The transition rates from this model are shown in Figure 3.
The arrows leading away from female-only rites in the model in Figure 3(b) are consistent with the Dependent model in Table 5, suggesting that female-only rites were unstable without also male initiation rites being present. As shown by the probability of each state being the root in Figure 3, this model also supports the hypothesis that female-only initiation rites were practiced during Austronesian origins.
These results, and the resulting phylogeny (Figure 4), all support transitions to female-only initiation rites being rare in Austronesian history. We found support for this in additional classes of Bayesian phylogenetic models, one in which the presence or absence of female initiation and male initiation rites were independent and another in which they were correlated but where double transitions (from none to both or vice-versa) were not allowed. In these models, adopting female initiation rites was rare relative to losing female rites, whereas adopting male rites was much more common than losing male rites (see Supplemental Material). In summary, the results indicate that the most likely ancestral state is female-only rite, but that subsequent transitions tended to be away from female-only initiation rites.

Language fan phylogeny of Austronesian ethnolinguistic groups, 50 color coded by type of initiation rites: blue, male only; red, female-only; purple, both female and male; black, no initiation rites. The societies are ordered by the “ape” package in R to enhance visual clarity.
Discussion
The prehistory of Oceania, relatively less affected by external contacts compared with other areas of the world, is an excellent place to attempt cultural reconstruction based on the comparison of related cultures. 53 Austronesian cultures have been described as “a natural laboratory for cross-cultural research due to the diversity of environments they inhabit and cultural features they have evolved.” 44
One facilitator of political complexity may be initiation rites—ceremonies that transfer an individual’s status from child in the domestic domain to citizen in the sociopolitical domain.45,46 It has been suggested that male peer-bonding during initiation facilitated complex socioeconomic organization. 46 Data summarized from Murdock’s Ethnographic Atlas 51 have been used to suggest that adolescent initiation rites co-evolved with socioeconomic complexity, with initiations for girls predominating in hunting-gathering and/or horticultural societies, and ceremonies for boys becoming more frequent in more complex societies. 46
Building on these results, here we find three notable patterns for initiation rites among early Austronesian societies: (1) female-only initiations were likely present during Austronesian origins; (2) female-only rites were not a stable state; and (3) initiation rites exclusively for one sex appear less stable than rites for both or neither sex.
Although female initiation rites tend to be far from the Austronesian homeland, where we are more likely to find matrilocal residence, multiple factor analysis and contingency tables indicate that subsistence was relatively uncorrelated with (orthogonal to) social-organizing phenomena of kinship system and initiation rites. Moreover, transition rates from the phylogenetic analysis indicate that female-only initiation rites were not a stable or likely state in proto-Austronesian societies. Figure 3(b) shows that the heaviest arrows (transition rates) point away from female initiation rites and toward either male-only initiation rites or both male and female initiation rites together.
Whereas there may have been a relationship among Eastern Austronesian societies between kinship system and initiation rite, there appears little relationship in the Western Malayo–Polynesian region, with its cluster of societies with no initiation rights (Figure 4) that spans multiple marital-residence states. Notably, the large cluster of societies with no initiation rites, in the Western Malayo–Polynesian region, is where cultural phylogenetic evidence for matrilocality is strongest . 24
Compared to patrilocal or ambilocal societies, matrilocal societies were observed twice as often to have no initiation rites (Table 3). If proto-Austronesian society was matrilocal, 24 a parsimonious inference is that proto-Austronesians had either female-only initiations or no initiations at all.
This difficulty inferring causality between kinship and initiation rite does not rule out a significant, potentially highly variable, state of tension between kinship system and initiation rite. While not an explanation, ethnographic detail may help us understand how this could work. Consider the Amis of Taiwan, potentially descendants of a proto-Austronesian homeland society, who embody matrifocality of household organization but with fraternal/paternal village community governance. 54 In the 19th century, the Austronesian-speaking Amis “lived on subsistence agriculture and hunting, practiced headhunting and initiatory discipline, tended to marry endogamously within the same village, [and] resided matri-uxorilocally” 54 (p. 188). About once every 3 years, when a new set of late-adolescent boys was initiated into this paternal/fraternal organization, the boys left their matrifocal households. This initiation set proceeded through the paternalistic hierarchy with age, from “sons” to “elder brothers” and eventually to “fathers.” Headquartered in a house in the village center, the hierarchical paternal system served to resolve community disputes, make collective decisions about land, and coordinate activities among the matrifocal households. 54 The initiation ceremony of males—feasting, dancing, ritual sacrifice of a pig, thrashings by senior initiation brothers—united members of the initiation set under paternal-based social norms (p. 196). 54
In any case, we expect fluidity during Austronesian dispersal from Taiwan in terms of household kinship, collective governance, and sacrificial ritual.24,43,44 By adding initiation rites into this cultural story, we have found evidence that all these cultural practices were correlated in their co-evolution during the prehistoric dispersal of Austronesian societies across the Pacific. Once acquired, male initiation rites appear to have been more resilient than female-only rites among Austronesian societies. This observation says little about causality, however, and though we have offered some hypotheses, it ultimately opens new questions future research.
Supplemental Material
sj-pdf-1-sci-10.1177_00368504211031364 – Supplemental material for Evolution of initiation rites during the Austronesian dispersal
Supplemental material, sj-pdf-1-sci-10.1177_00368504211031364 for Evolution of initiation rites during the Austronesian dispersal by R Alexander Bentley, William R Moritz, Damian J Ruck and Michael J O’Brien in Science Progress
Footnotes
Acknowledgements
We thank Helen Bass for her year-plus of ethnographic research, Tom Currie for comments and permission to use the map in Figure 1, and Luke Matthews for comments. DJR is supported by grants from the College of Arts and Sciences and Office of Research and Engagement, University of Tennessee–Knoxville, as well as by a grant from the Office of the Provost, Texas A&M University–San Antonio. RAB and DJR thank their students in ANTH 493 at the University of Tennessee, Knoxville for exploring some of these data in class. Data and relevant code for this research work are stored in GitHub repository: ![]() .
.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: DJR was supported by grants from the College of Arts and Sciences and Office of Research and Engagement, University of Tennessee–Knoxville.
Supplemental material
Supplemental material for this article is available online.
Author biographies
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.