
Abstract
Rhesus macaques (Macaca mulatta) used in behavioural neuroscience are often required to complete cognitively complex tasks, for which a high level of motivation is essential. To induce motivation, researchers may implement fluid-restriction protocols, whereby freely available water is limited, such that fluid can be used as a reward in the laboratory. A variety of different rewards and schedules are used, but there exists a lack of data assessing their effectiveness. In this study, we aimed to quantify fluid preference in rhesus macaques and to use these preferences to compare the motivational quality of different reward schedules: the monkey's previous reward (i.e. the fluid used to reward them in past studies), their new preferred reward, a variable schedule of previous and preferred reward, and a choice between the previous and preferred rewards. We found that it may be possible to reduce the level of restriction if an adequately motivating preferred reward is identified, but that this is dependent on the animal. Each monkey responded differently to both the fluid-preference assessments and to the different reward schedules. As such, monkeys should not be subject to ‘blanket’ protocols but should be assessed individually to maintain adequate scientific data collection at the least severe level of fluid restriction.
Macaques are widely used across biomedical sciences, 1 and are often selected as a model in behavioural neuroscience studies because of their similarities to humans in brain structure and function, as well as their ability to perform complex tasks.2,3 In particularly cognitively challenging tasks, or where studies require large numbers of trials to be undertaken, a high level of motivation is required. This may lead researchers to choose to restrict the amount of freely available fluid in order to motivate a monkey to perform a sufficient number of responses. 4 Using restriction protocols has led to concerns about the impacts on the welfare of the animals,5,6 though these may be largely unfounded. 7
An NC3Rs working group 4 emphasised that researchers should carefully choose reward schedules and reward types to adequately motivate animals whilst minimising the degree of restriction required. Many aspects of reward processing have been investigated in macaques, such as reward uncertainty, 8 differing reward sizes, 9 temporally varying reinforcement schedules, 10 and the neuronal coding of reward preference. 11 However, less work has been conducted into how to use this knowledge of reward processing to refine laboratory procedures. Although there are multiple ways in which motivation could potentially be increased, we chose to investigate three aspects of reward that we thought may be effective at increasing motivation to perform in tasks and allow restriction protocols to be relaxed. The first is the use of preferred rewards, which can be more rewarding12,13 and result in more successful training of behaviour. 14 The second possibility is using a variety of rewards, since animals may perform better when their rewards are varied throughout a task, rather than using a single type of reward.15,16 Finally, giving monkeys a choice of reward may also enhance motivation.17–19 Despite these possibilities, there are currently no conclusive data to guide researchers to which method might be the best way to motivate their animals in behavioural neuroscience tasks.
This study had three main aims: (a) to explore methods to efficiently quantify fluid reward preference in rhesus macaques (Macaca mulatta); (b) to use schedules involving previous rewards and new, preferred fluids to evaluate the motivational abilities of different reward schedules; and (c) to establish if reward schedules involving preferred fluids could keep macaques sufficiently motivated to perform cognitive tasks when their fluid restriction was relaxed. We expected monkeys would be more motivated when they were rewarded with schedules that included a preferred fluid, and that increased motivation levels would enable animals to perform cognitive tasks when fluid restriction was relaxed.
Animals
Experimental procedures complied with the European Union Directive 2010 (2010/63/EU), the National Institutes of Health (Guidelines for Care and Use of Animals for Experimental Procedures), the Society for Neurosciences Policies on the Use of Animals and Humans in Neuroscience Research, and the UK Animals Scientific Procedures Act. The study was approved by the Animal Welfare Experimental Review Board (AWERB) of Newcastle University.
Four male rhesus macaques weighing 8–13 kg were used. Animal usage was opportunistic, when individuals were not partaking in neuroscience studies (the laboratory uses only male macaques). Each subject was housed with another male, in cages (either 2.1 × 3.0 × 2.4 m or 2.3 × 2.45 × 2.4 m) and had visual, olfactory and auditory contact with approximately 40 other macaques. Toys were given on a rotational basis and dry food mix (Mazuri Primate Expanded, Old World Monkey Banana Chunks, Trio Munch Rings and LP Forage Mix, Special Diet Services; Monkey Diet, LabDiet®, IPS Ltd) was added to floor shavings to encourage foraging. Cages were equipped with perches, shelves, a ‘balcony’ and hoses. The facility had a 12:12 light/dark cycle (7 a.m. to 7 p.m.) and natural light from ceiling windows. The temperature and humidity were approximately 20℃ and 24%, respectively.
During the experiment, the macaques had controlled access to fluids. The fluid-restriction protocol consisted of five days of fluid restriction (minimum daily intake given from Sunday to Thursday) with free access to water after completing work on Friday and all day on Saturday. The minimum daily fluid intake was a volume of water which sufficiently motivated the monkey to perform the laboratory task (based on being able to perform approximately 1000 trials in a session) and is expressed as a percentage of their consumption when given free access to water (Monkey 1 = 250 ml, 24% or 25 ml/kg/day; Monkey 2 = 200 ml, 25% or 13 ml/kg/day; Monkey 3 = 200 ml, 22% or 16 ml/kg/day; Monkey 4 = 385 ml, 30% or 23 ml/kg/day). Through participation in experiments, individual monkeys were permitted to work for as much fluid as they liked. On days where monkeys did not reach their daily fluid intake during the experiment, they were supplemented with additional water. All monkeys had previously performed in similar tasks in the laboratory set-up and were familiar with this type of experiment.
During the study, monkeys underwent daily checks by a technician or veterinarian. In case of a health or welfare concern, technicians and the veterinarian checked the animal several times per day. Fur condition, faeces, eyes, food intake and activity levels were all visually assessed. Monkeys were sedated annually to assess their general health (as per Felasa guidelines) and no microorganisms or parasites of current concern for macaques were detected. Animals were kept for future studies at the end of the current study.
Methods
Establishing fluid preferences
A fluid preference for each monkey was established by one of two methods. The first used the experimental set-up, where animals had already been trained to saccade to stimuli to access fluid rewards. We devised a simple saccade task, where different fluid rewards were delivered by looking at visually distinct stimuli presented on a screen, allowing animals to choose their reward. Fluid preferences for Monkey 1 and Monkey 2 were assessed in this way. Each monkey was seated in a primate chair and stimuli were presented on a Sony GDM F500R computer monitor (85Hz, 1280 × 1024 pixels). Stimulus presentation, reward delivery and experimental timing were controlled using Cortex (DOS-Version 5.95; IMH, http://dally.nimh.nih.gov/) running on IBM-compatible PCs (situated outside of the monkey's room).
Animals performed a saccade choice task on each experimental day, consisting of repeated trials until the animals stopped working. In each trial, they were required to fixate on a central spot for 3000 ms, after which three reward targets appeared. The monkey had to saccade to any one of the three reward targets and fixate for 250 ms to complete a trial correctly and receive an associated fluid reward (∼0.1 ml fluid). Failure to fixate on a stimulus for long enough terminated the trial and the animal was not rewarded. The three reward targets (2 degrees of visual angle, (dva)) were located at positions (x = −6.0, y = 6.0), (x = 0.0, y = −8.5) and (x = 6.0, y = 6.0), equidistant from the fixation spot (x = 0, y = 0). To control for location bias, target location was pseudo-randomised such that targets occupied the different locations for equal numbers of trials. Each reward target was associated with a different fluid reward and was distinguishable by colour for Monkey 1 (pink, red or blue) and by shape for Monkey 2 (circle, triangle and square).
We gave the monkeys the choice between water (which they had previously received as a reward) and two fruit drinks, one nutritive and the other non-nutritive. The nutritive fruit drink was Ribena (Lucozade Ribena Suntory Ltd; 40 ml of undiluted squash added to 210 ml of water), which had been successfully used by other researchers to motivate their animals. Fruit tea (a cranberry and raspberry tea bag (Twining and Company Ltd; placed in 250 ml of hot water for 5 min, before being allowed to cool) had the taste of fruit without any nutritive content.
The three fluids (water, Ribena and fruit tea) were delivered through a mouthpiece, connected to three separate bottles by plastic tubing. The bottles were calibrated to ensure that the same amount of reward was delivered from each. The fluid preference task was run for six days for Monkey 1 and eight days for Monkey 2. The additional two days for Monkey 2 were a result of a lack of consistent preference in the first six days; two more days were added to see if any preference pattern emerged (see further detail in the results section). The fluid with the highest number of choices on more than 50% of the days was taken as the monkey's preference. These preferences were then used to inform the design of the main part of the experiment, which investigated the reward value of different motivational schedules.
Establishing fluid preferences using the laboratory set-up was time-consuming, required additional apparatus and only allowed for three fluids to be tested. Given that ideally fluid-restriction refinements should be easy to implement, we decided to test fluid preferences for Monkeys 3 and 4 using a second method in the home cage to screen more fluids in a simpler and quicker task. The fruit tea was not used in the home cage as it is not as viscous as the fruit juices and the bottles used in the cage could not be calibrated to dispense equally like the bottles in the experimental set up. By using juices of similar viscosity, we hoped to control for the amount that could be consumed from the bottle.
In the home cage, the monkeys could not be fluid restricted (under the conditions of the Home Office licence), and so had free access to water during preference tests. Initial tests consisted of a range of four different juices (apple, pineapple, tropical and orange; all Tesco Stores Ltd) presented in a choice paradigm to narrow the options down to two preferred juices. Each monkey was separated from his cagemate and offered pairs of juices via 10 ml syringes. An initial, randomly selected pair of juices was presented to the monkey to sample in turn (5 ml of each) before both juices were offered simultaneously (5 ml of each). Using only 5 ml at a time meant that we did not risk the monkeys becoming satiated. The juices were offered in the same location simultaneously as they had been separately, so that the location of the syringe signalled to the monkey which juice was which. The preferred juice (noted by which syringe he chose to drink from) was then refilled and presented alongside a new, randomly selected juice. This was continued until all combinations of juice had been presented (a total of six pairs). The two juices chosen the most often were used in the experimental stage, along with the fluid with which the monkey had been previously rewarded in cognitive tasks.
The two preferred fluids and the monkey's previous fluid reward were presented in 1 l bottles attached to the cage in three positions: left, middle and right. The monkey had 5-mins access to the bottles, and the volumes consumed were recorded. The 5-min test was carried out at the same time each day (09:00–10:00) for six days. Each day, the bottles were spatially arranged in a unique way that allowed every combination of fluids and positions to be presented once. The fluid chosen consistently over the six days (defined as chosen on 50% or more of the days) was used as the preferred reward when assessing the reward schedules in the laboratory. We note that each monkey drank from all three bottles on all testing days (i.e. did not simply try one bottle without sampling the others).
Assessing the motivational value of different fluid-reward schedules
Once fluid preferences were established, each monkey performed a familiar task whilst fluid restricted at their normal level (established in previous studies). Although the tasks differed between monkeys, the nature of the task was not relevant; it was only important that a monkey was familiar with a task and could consistently perform it to measure the effectiveness of the different reward schedules.
The monkeys performed their task on different days where they received one of four different reward schedules. Upon completion of a correct trial, monkeys received either the reward given to them in previous studies (previous reward), their preferred reward established from the preference tests (preferred reward), a 50% chance of receiving either the previous or preferred reward (variable schedule), or a choice between previous and preferred rewards (choice schedule). In the choice schedule, the monkeys chose their reward by fixating for 250 ms on one of two stimuli: a cross-shaped stimulus represented the previous reward and a circle represented their preferred reward (presented at (x = −6.0, y = 0.0, or x = 6.0, y = 0.0 dva). The reward schedules were carried out in four blocks of four days, with schedules randomised within blocks, such that each block lasted for four days and only one schedule was used on any given day. The number of correct trials performed was recorded on each day as a measurement of the monkeys' motivation to work.
To assess whether the monkeys' performances on their respective tasks could be maintained under less restrictive fluid-restriction conditions, the daily fluid intake of the monkeys was increased by 100 ml, and the blocks of reward schedules repeated. This increase was deemed suitable for all animals given that no monkey drank 100 ml over his minimum volume on a daily basis during the experiment.
Statistics
All data were checked for normality and equal variances and analysed using appropriate parametric or non-parametric tests in SPSS (v21, SPSS Inc, Chicago, USA). All pairwise tests were corrected for multiple comparisons using false discovery rate (FDR) post hoc tests 20 and reported using a corrected alpha value (termed the q-value). Details of individual tests are reported alongside their corresponding statistics in the Results section below.
Results
Establishing fluid preferences
To establish fluid preferences, a one-way ANOVA (Monkeys 1, 2 and 3) or Kruskal–Wallis test (Monkey 4) was used to compare the number of choices for each fluid in the laboratory (Monkeys 1 and 2), or the amount of each fluid consumed in the home cage (Monkeys 3 and 4). In the laboratory set-up, a clear fluid preference could only be established for Monkey 1. Monkey 1 differentially chose the three fluids (ANOVA, F(2,15) = 48.62, p < 0.001; Figure 1(a)), preferring Ribena to both fruit tea (t(10) = 6.78, q < 0.05) and water (t(10) = 9.64, q < 0.05) and preferring fruit tea over water (t(10) = 3.0, q < 0.05). This pattern was consistent on every day, with the highest number of choices always for Ribena, followed by fruit tea and the lowest number of choices always being for water. Whilst Monkey 2 also varied in the number of choices for each fluid (F(2,21) = 3.89, p = 0.037; Figure 1(b)), this was not consistent across days and was biased by a high intake of fruit tea in the first three days of testing (Figure 1(c)). We therefore added an additional two days of testing to examine whether any preference pattern emerged (a total of 8 days). However, the inconsistency remained, and Monkey 2 was not continued in the experiment as we could not establish a preference or be sure that the monkey understood the task.
Fluid preference testing in the laboratory (Monkeys 1 and 2) and in the home cage (Monkeys 3 and 4). The average number of choices for rewards in the preference test for (a) Monkey 1 and (b) Monkey 2. (c) Monkey 2 was not continued in the experiment as his preference was not stable across the 8 testing days The average consumption of each reward in 5 minutes over 6 days for (d) Monkey 3 and (e) Monkey 4. Bar charts display mean (±SEM) and box plots display medians. The previous reward of each monkey is always shown as the left-hand bar.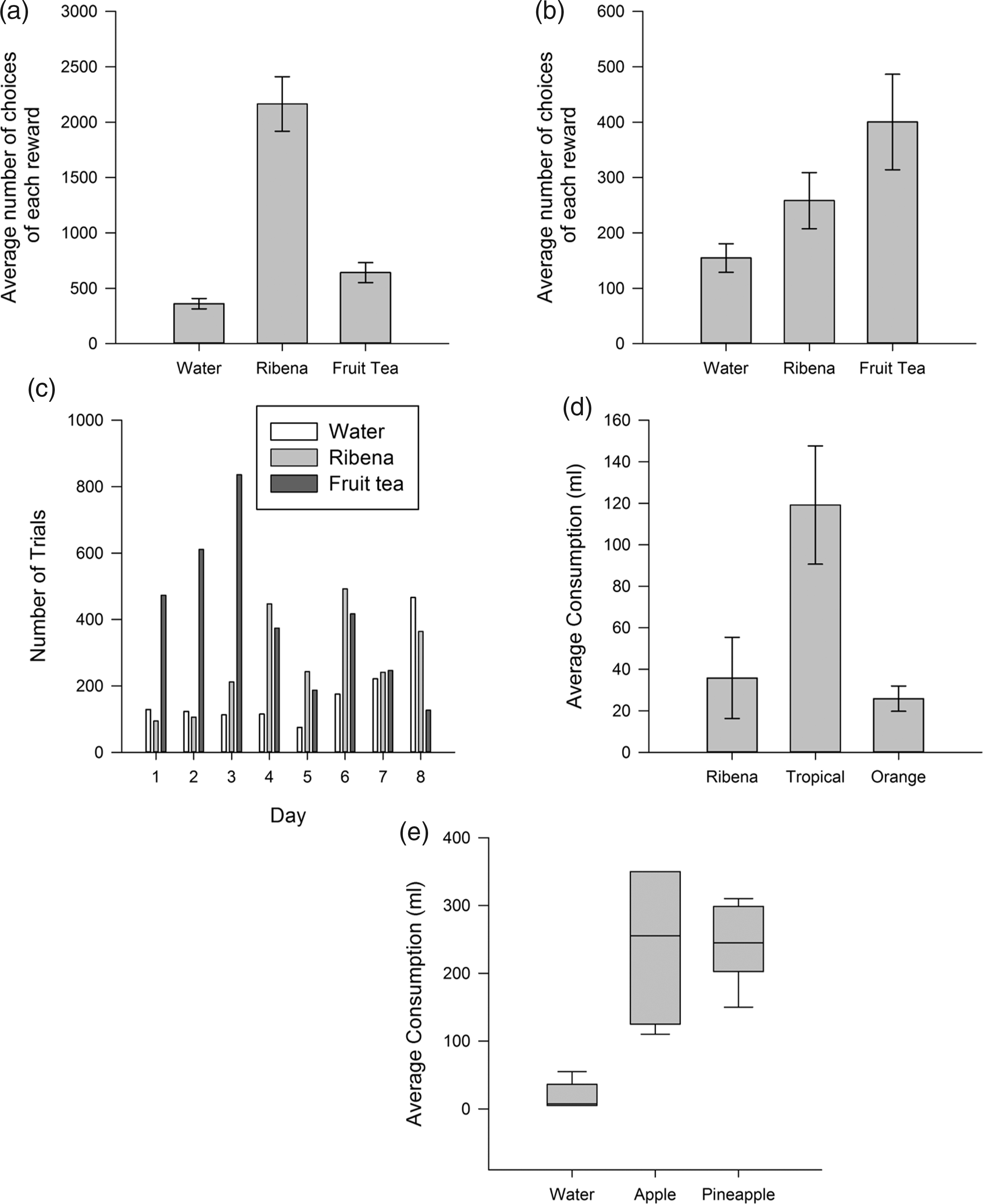
In the home cage, fluid preferences were established for both Monkey 3 (F(2,15) = 5.83, p = 0.013; Figure 1(d)) and Monkey 4 (Kruskal–Wallis, H2 = 11.43, p = 0.003; Figure 1(e)). Monkey 3 preferred tropical juice to both his previous rewards of Ribena (t(10) = 2.89, q < 0.033) and to orange juice (t(10) = 3.42, q < 0.033), with no difference between the orange juice and Ribena (t(10) = 0.091, q > 0.033). Monkey 4 preferred both new juices over his previous reward of water (apple: U = 2.93 q < 0.033; pineapple: U = 2.93, q < 0.033), with no difference between apple and pineapple juice (U = 0, q > 0.033). Apple juice was chosen to be carried forward as his preference as there was a slightly more pronounced choice for this (median consumption: Apple 255 ml, Pineapple 245 ml).
Assessing the motivational value of different fluid-reward schedules
Motivation was assessed individually for each monkey by comparing the number of trials completed at each fluid-restriction level. Monkey 1 received Ribena as his preferred reward alongside water (his previous reward). At normal fluid intake, Monkey 1's performance varied across the four reward schedules (Kruskal–Wallis, H3 = 12.40, p = 0.006; Figure 2(a)). His highest performances (defined as the number of trials completed correctly) were for his preferred reward or a variable reward schedule, for which he performed equally well (Mann–Whitney, U = 0.15, q > 0.017). His motivation was lower for the previous reward compared to both of these schedules (Ribena: U = 2.82, q < 0.017; Variable: U = 2.97, q < 0.017). Despite a trend towards decreased performance when given a choice of reward, the number of trials was not significantly different from the preferred reward (U = 1.78, q > 0.017), the variable schedule (U = 1.93, q > 0.017) or the previous reward (U = 0.30, q > 0.017). Unlike Monkey 1, Monkeys 3 and 4 did not differ in their task performance for different fluid-reward schedules (Monkey 3: H3 = 7.22; Monkey 4: F(3,12) = 1.61; p > 0.05 for both; Figure 2(b) and (c)), demonstrating that, for these two monkeys, the schedules were equally motivating at a normal restriction level.
The average number of correct trials performed by (a) Monkey 1, (b) Monkey 3 and (c) Monkey 4 when rewarded with their previous reward, preferred reward, a variable schedule or a choice schedule at both their normal and increased fluid intakes. Bar charts display mean (±SEM) and box plots display medians.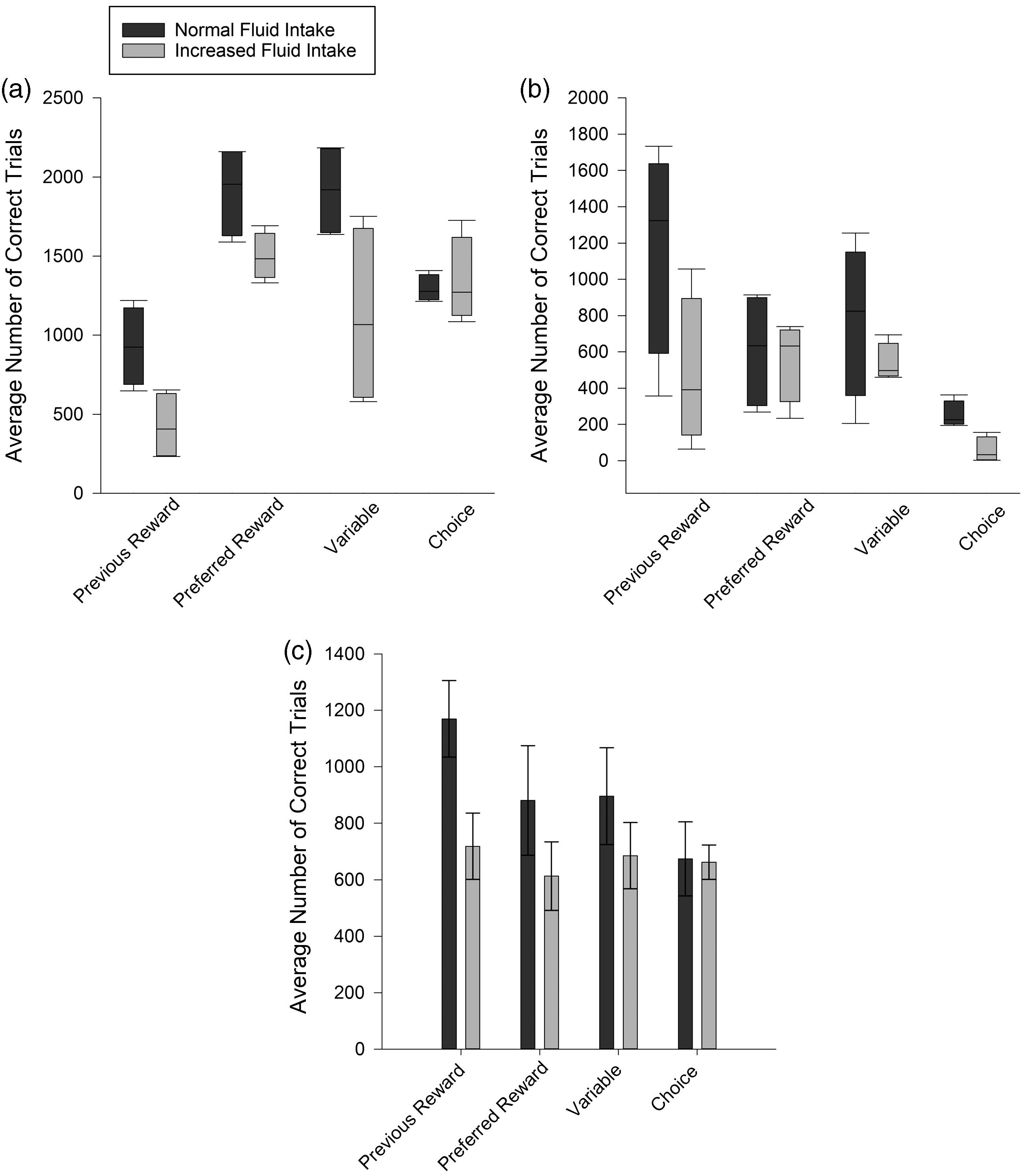
The minimum level of task performance in the laboratory is considered to be 1000 daily trials, given the recording requirements of our experimental approaches. When rewarded with his previous reward, Monkey 1's performance did not differ from 1000 trials (one sample t test: t(3) = 0.57, q > 0.0375), and all other reward schedules elicited performance of over 1000 trials (t(3) < 6.89, q < 0.0375 for all), showing all reward schedules to be sufficiently motivating. These results suggest that the inclusion of a preferred reward increased motivation to beyond that of water. Monkey 3's performance was no different from 1000 trials when rewarded with his previous reward, preferred reward or a variable schedule (t(3) < 2.41, q > 0.0125 for all) but dropped to lower than 1000 trials when he was given a choice (t(3) = 19.84 q < 0.0125). Monkey 4's performance did not differ from 1000 trials for any schedule (t(3) < 2.49, q > 0.0125 for all), suggesting that all schedules were sufficiently motivating at the normal fluid-restriction level.
Monkey 1: pairwise comparisons between the numbers of trials performed after the daily fluid allowance had been increased by 100 ml. Results are controlled for multiple comparisons using False Discovery Rate (FDR) tests.
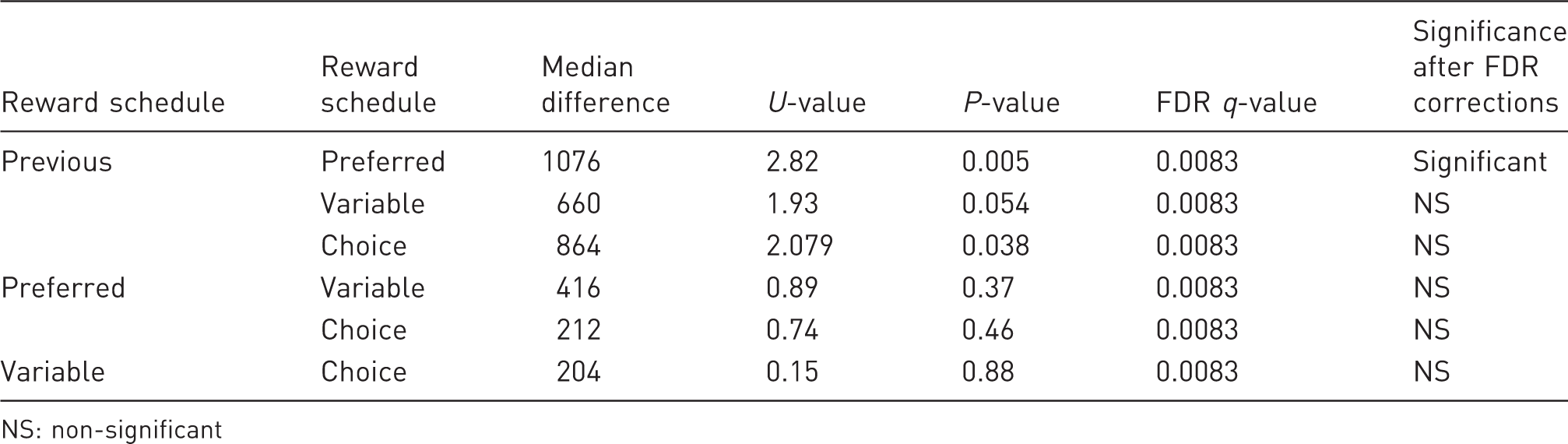
NS: non-significant
Monkey 3: pairwise comparisons between the numbers of trials performed after the daily fluid allowance had been increased by 100 ml. Results are controlled for multiple comparisons using False Discovery Rate (FDR) tests.
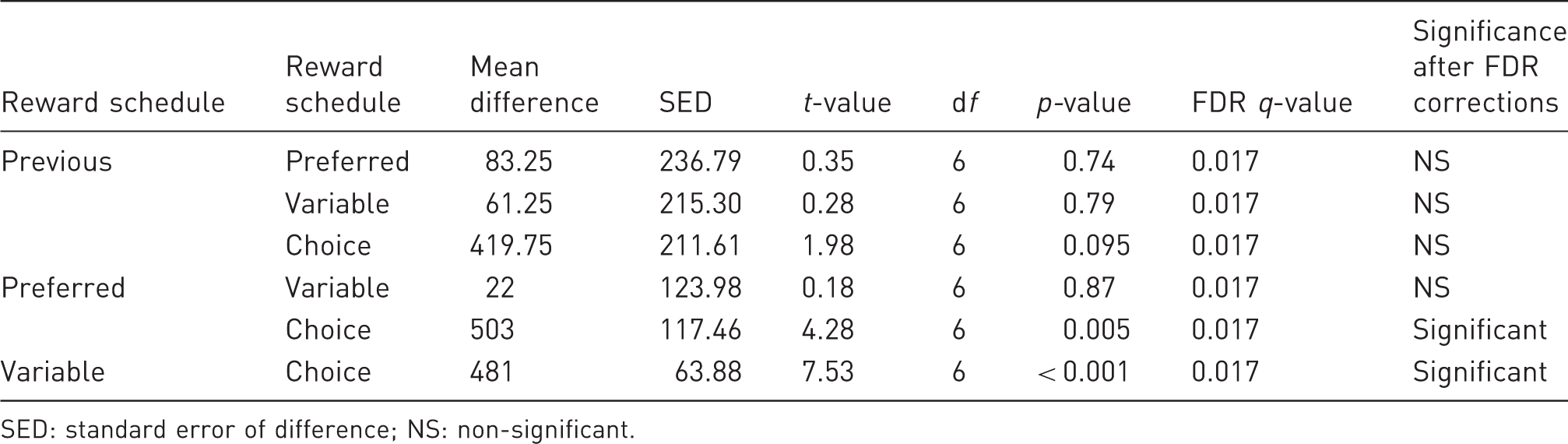
SED: standard error of difference; NS: non-significant.
Again, we compared the monkeys' performances to a 1000-trial threshold. Monkey 1's daily performance remained around 1000 trials when rewarded with the choice and the variable schedules (t3 < 2.47, q > 0.025), and he performed over 1000 trials when rewarded with his preference (t3 = 6.67, q < 0.025). However, his performance was not sufficient (<1000 trials) when rewarded with his previous reward (t3 = 5.36, q < 0.025), suggesting that his previous reward had now decreased in value whilst the preferred reward continued to be motivating. In contrast, Monkey 3's performance with his previous reward remained at around 1000 trials (t3 = 2.51, q > 0.0375) whereas performance dropped below 1000 when he was given his preferred reward, the variable schedule or a choice of rewards (t3<26.67, q < 0.0375 for all), indicating that the previous reward was the only motivating fluid at this restriction level. For Monkey 4, trials completed for his previous reward, preferred reward and variable schedule did not differ from 1000 (t3 < 3.19, q > 0.0125 for all) but did fall below 1000 for the choice schedule (t3 = 5.52, q < 0.0125), demonstrating the lack of value this had at an increased fluid intake.
Difference in trial performance when the fluid allowance was increased. Results are controlled for multiple comparisons using False Discovery Rate (FDR) tests.
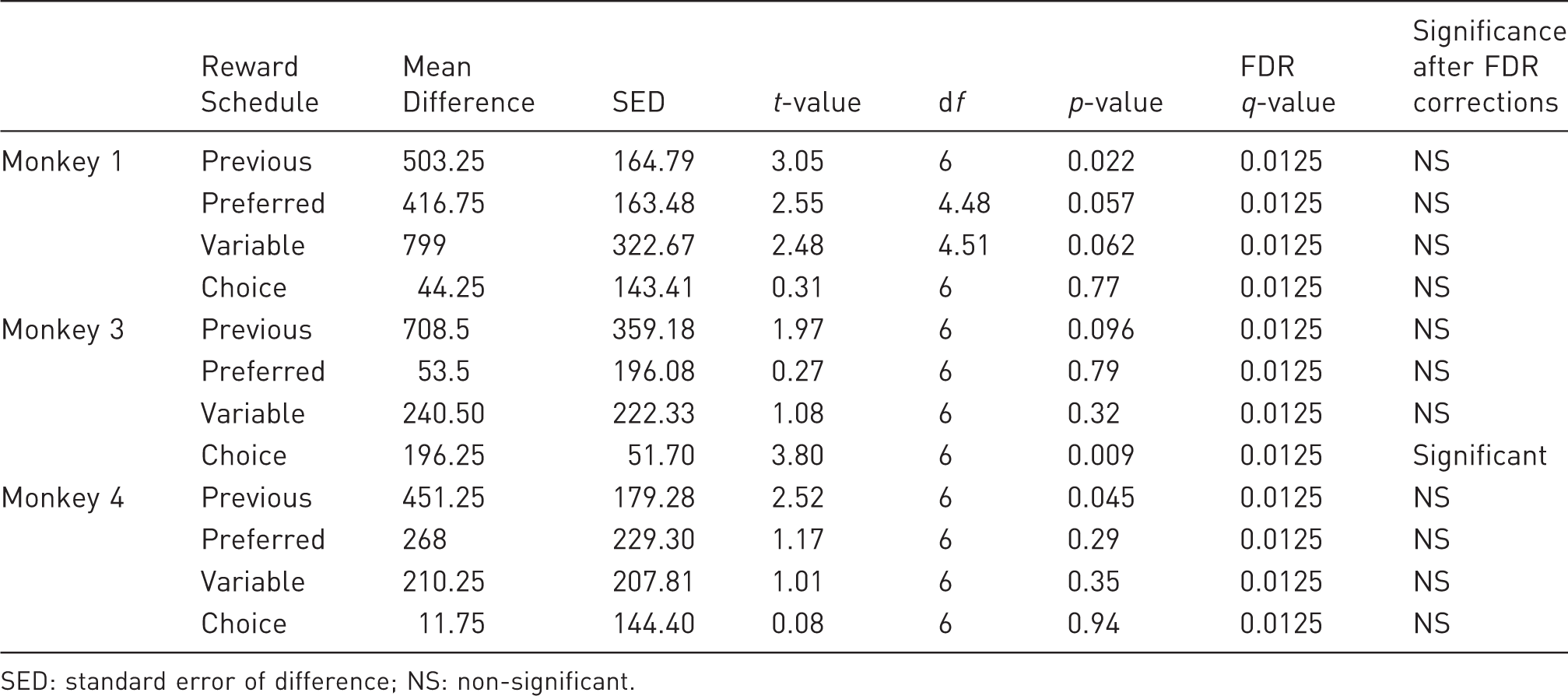
SED: standard error of difference; NS: non-significant.
Discussion
Our study shows that using a preferred fluid can help to increase motivation in cognitive tasks and allow restriction protocols to be relaxed to achieve adequate performance in the task. However, we only found the use of preferred fluids to be effective when the fluid preference was established under fluid control in the laboratory, and not when it was established in animals that were under free access to fluids in the home cage. In addition, schedules where an animal could choose his reward (Choice), or where the preferred reward arrived only 50% of the time (Variable), did not appear to increase motivation beyond simply being rewarded all the time with the preferred fluid. We discuss how our results can potentially contribute towards establishing methodology to enhance motivation in laboratory macaques performing behavioural neuroscience tasks, and enabling researchers to relax fluid control protocols to address welfare concerns.
Our study provides the first empirical support for the idea that using preferred fluids can enhance motivation and allow relaxation of fluid control for macaques used in behavioural neuroscience tasks. Reward preference is evident at both neuronal11,21 and behavioural levels in macaques,22,23 however, using preferences to refine fluid control may be dependent upon the method by which fluid preferences are established, or it may be dependent on the individual. We established preferences using one of two methods and were only able to establish a fluid preference that enhanced motivation and task performance in one animal (Monkey 1), where preference was established under fluid control in the laboratory. Interestingly, this method was not always successful, as Monkey 2 did not show a consistent preference using this method. It is unknown whether this was because Monkey 2 could not learn to associate a symbol to a corresponding fluid to make a choice or due purely to a lack of preference. Although we could establish fluid preferences for the two animals tested unrestricted in their home cage (Monkeys 3 and 4), these preferences did not translate to improved performance in the laboratory.
One possible reason for this difference is because the preferences were evaluated under different fluid control protocols: Monkey 1's preference testing was conducted under fluid control, whilst that of Monkeys 3 and 4 was done whilst the animals had free access to water. Perhaps the preferences that monkeys have when satiated simply do not transfer to preferences under fluid control, and consequently preferred fluids established under free access were less effective as rewards when the monkeys were fluid restricted later in the study. An alternative explanation, which we currently cannot rule out, is that inter-individual differences were responsible for the results.
A further possibility is that the previous reward of each monkey impacted on the efficacy of the new, preferred reward. Monkeys 1 and 4 had previously been rewarded with water, and Monkey 3 with Ribena. Monkey 3 continued to perform well for his previous reward at the increased fluid allowance, whereas Monkeys 1 and 4 decreased their performance to below 1000 trials. The monkeys are supplemented with water if they have not reached their daily intake allowance via task performance and thus for Monkey 3, it may be that Ribena remained motivating when he had learned he would receive only water afterwards. Conversely, for Monkeys 1 and 4, it was probably less motivating to be rewarded with water, as it could be received for ‘free’ after work, especially when motivation was lower, and they could afford to wait for their water. This would be an interesting aspect of reward motivation to be investigated in the future, as it may provide a potential method to reduce fluid restriction, by rewarding animals solely with fluids which they cannot receive away from the experimental set-up.
We had predicted that other reward schedules containing a preferred fluid (Variable and Choice) would enhance motivation, but this was not necessarily the case. Although the variable schedule produced performance similar to that of the preferred reward at the normal fluid intake, performance dropped for two of the monkeys when the fluid restriction was relaxed. It is perhaps surprising that the variable schedule was not more motivating given previous evidence: variability in reward size, and the consequential unpredictability, results in dopamine release in macaques, particularly when the chance of receiving a reward is at 50%. 24 In addition, for rats, using two varied rewards throughout a task has been shown to reduce habituation and enhance performance above that of a single reward alone.15,16,25 However, we found no evidence for enhanced performance using these schedules, and there would be no advantage to their use beyond providing a preferred fluid alone. Our findings are more in line with Bowman et al., 26 who showed that variable schedules are less motivating to primates than they are to other species.
When considering the choice schedule, we expected that free choice may enhance motivation,12,17,18 but we found a decrease in motivation. We speculate that low levels of performance were likely due to the additional effort that was required. The monkeys first had to perform the initial task correctly, before being offered a choice. This meant that a trial took longer and required additional cognitive steps. While choice is often seen as potentially rewarding,17–19 our findings suggest that the costs and benefits may balance out under these laboratory conditions.
Although we tested three aspects of reward, there are further avenues yet to be fully explored. First, occasional larger rewards (‘jackpots’) have been advocated as potentially rewarding for animals. 27 However, there is currently no evidence they prove motivating to macaques performing in cognitive neuroscience studies. In addition, recent work by Fischer and Wegener used a non-binary positive reinforcement approach. 28 They provided different volumes of fluid dependent on how optimally the monkey had performed a task (e.g. larger volumes for successfully performing a new step of the task and lower volumes for reverting to a previous version of the task). This training technique warrants further implementation to assess effectiveness in other laboratories.
Conclusions
Our study demonstrates that if an animal has a preferred fluid reward, it can be possible to use it in cognitive tasks to relax their fluid restriction whilst keeping their performance at a level to enable sufficient high-quality scientific data collection. This has laboratory-welfare implications, since relaxing fluid-restriction protocols addresses one of the key concerns around the use of primates in behavioural neuroscience. 4 Whilst we advocate establishing and using preferred fluids for primates, more research may be required to find a more efficient method to identify preferred fluids that increase motivation under laboratory conditions. The only method we found to be effective was conducting preference tests in the laboratory, which can be time intensive. It thus increases the time animals have to engage in licenced protocols. Whether this is justified in light of the limited impact of fluid control, 7 remains to be determined. Our data do not provide support for home cage preference testing, but we would encourage further exploration of this issue.
Footnotes
Acknowledgements
We would like to thank the staff of the Comparative Biology Centre (Newcastle University) for excellent technical support. With thanks also to Dr Michael Boyd for his computer programming skills.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this news article: The work was supported by the NC3Rs (NC/K500410/1) and the Wellcome Trust.