
Abstract
This study aims to determine the ability of laboratory animal bacteria to resist desiccation and inactivation by hydrogen peroxide vapour (HPV) on paper bedding pieces. Bedding pieces were saturated with bacterial suspensions in water or 2% (w/v) bovine serum albumin (BSA) in water, and held in a mouse facility. Viable counts showed variable survival rates over time for the bacterial species used ([Pasteurella] pneumotropica, Muribacter muris, Pseudomonas aeruginosa, Acinetobacter redioresistens, Escherichia coli, Klebsiella oxytoca, Bordetella bronchiseptica, Bordetella hinzii, Enterococcus faecalis, β-haemolytic Streptococcus spp., Staphylococcus aureus and Staphylococcus xylosus). Overall, BSA increased bacterial survival in the bedding pieces. The survival rates of Bacillus safensis were not influenced by BSA but depended on sporulation. When bedding pieces and Petri dishes inoculated with E. coli, P. aeruginosa and S. aureus were subjected to HPV disinfection, all bacterial species on the bedding pieces inoculated with bacterial suspensions in water were readily inactivated. By contrast, S. aureus and P. aeruginosa, but not E. coli cells survived HPV treatment in high numbers when inoculated on bedding pieces as a BSA suspension. Notably, all three bacterial species were readily inactivated by HPV even in the presence of BSA when smeared on smooth surfaces. In conclusion, the suspension medium and the carrier can influence the environmental survival and susceptibility of bacterial species to HPV. Our results may help to develop standard protocols that can be used to ensure the microbiological quality of experimental rodent housing.
The microbiological quality of experimental animals is a prerequisite for obtaining valid in vivo results. Room disinfection and microbiological monitoring represent two practices required to achieve and maintain the specific pathogen-free (SPF) status of laboratory animals. Microbiological monitoring of the animals housed in individually ventilated cages is usually carried out on sentinel mice exposed to soiled bedding. There are several studies that have investigated the transmission of microbes to sentinels via soiled bedding.1–4 One critical factor that may influence the transmission of bacterial agents via soiled bedding is their ability to survive for a sufficient time on bedding particles. However, there are only a few studies documenting the environmental stability of such microorganisms. 5
The successful application of hydrogen peroxide vapour (HPV) decontamination in hospital settings encouraged its use for the disinfection of laboratory animal rooms or equipment used in SPF areas as an alternative to formaldehyde fumigation. 6 Hydrogen peroxide (H2O2) is a potent oxidizing agent. Its microbicidal activity is thought to be mediated by free hydroxyl radical production, which has a deleterious effect on membrane lipids, DNA and proteins.7,8
Although microbiological monitoring by sentinel animals using soiled bedding and HPV room disinfection are routine procedures in modern animal facilities, no data are currently available on the ability of bacterial pathogens to survive on bedding particles or to resist HPV disinfection in animal room settings. For these reasons, the main goal of the present trial was to investigate the ability of selected bacteria, relevant to laboratory animals such as rodent Pasteurellaceae, Escherichia coli, Klebsiella oxytoca, Bordetella spp., Pseudomonas aeruginosa, Staphylococcus aureus and others, to survive on cage bedding pieces in the environment of an animal room. A further objective of this study was to test the ability of HPV to render inactive bacteria located on cage bedding.
Materials and methods
Bacterial isolates
Bacterial strains used in this study.
Quantification of bacterial survival on paper bedding pieces within the animal room
Shepherd’s™ ALPHA-dri® bedding pieces (width/height/depth approximately 5/5/1 mm; Shepherd Speciality Papers, Kalamazoo, MI, USA) (Figure 1), made of alpha cellulose, which are used as bedding for mice housed in individually ventilated cages in our facility, were used as carrier for the bacterial cells in order to simulate natural contamination. To prepare the test bedding pieces, bacteria were cultured aerobically on Columbia blood agar plates (bioMérieux, Nürtingen, Germany) for 24 h at 37℃. Subsequently, the bacteria were suspended at OD600 = 0.5 in sterile distilled water (SDW) or SDW containing 2% (w/v) bovine serum albumin (BSA) (Sigma-Aldrich, Taufkirchen, Germany), to simulate biological soiling. Approximately 40 bedding pieces per bacterial strain were introduced into the bacterial suspensions and allowed to absorb bacteria for 5 min at room temperature. The bedding pieces were then collected with sterile forceps and placed into Petri dishes which were closed and kept during the experiment in a populated mouse room, at 22 ± 2℃, relative humidity of 45–65% and light/dark light cycles of 12:12 h, in order to test the resistance of bacteria to desiccation over time. To determine the viable bacterial counts, one bedding piece was introduced into a 1.5 mL Eppendorf tube containing 1 mL of SDW and shaken for 10 min at 1000 rpm at room temperature to allow bacterial re-suspension into water. Subsequently, serial 10-fold dilutions with 20 µL volumes were performed within a 96-well plate (Rotilabo®-microtest plates; Carl-Roth, Karlsruhe, Germany). Ten-microlitre suspensions from each dilution were then placed on Columbia blood agar and incubated for approximately 24 h at 37℃. The counts of viable bacteria on the bedding pieces were determined immediately and after 2, 6, 24 and 48 h for the Pasteurellaceae species used, whereas for the other bacterial species the amount of bacteria was recorded after 4 and 24 h, and 1, 2, 3, 4, 5 and 6 weeks. For Bacillus safensis the survival capacity was additionally tested at 12, 24, 48 and 72 weeks after sample preparation. In order to check whether the viable counts of B. safensis at later time points relied on bacterial sporulation, the 12-week-old bedding smears were stained with a malachite green stain that detects bacterial spores. For each time point and bacterial strain the numbers of colony forming units (CFUs) were determined on triplicate samples.
Form and size (cm) of the bedding pieces used in this study.
Quantification of bacterial inactivation by HPV
To test bacterial resistance to HPV, three bedding pieces inoculated 24 h before testing with E. coli, P. aeruginosa and S. aureus bacterial suspension in SDW or SDW + 2% BSA as described above, were kept for 24 h in open Petri dishes in a 59 m3 (width/height/depth 3.4/3/5.8 m) unoccupied mouse room which was then subjected to HPV decontamination. In addition, open Petri dishes, whose surfaces were smeared with OD600 = 0.5 bacterial suspension of the same strains in 2% BSA 24 h before the HPV decontamination, were placed in the room in order to evaluate bacterial survival on smooth surfaces by stamping with contact plates containing 5% sheep blood Columbia agar. All Petri dishes were kept on a small table close to the door at a height of approximately 30 cm from the floor. In parallel, replicates of the Petri dishes with the same content were kept outside the room and served as controls. The HPV decontamination experiment described above was performed three times.
Geobacillus stearothermophilus spore strips (BAG Health Care, Lich, Germany) with a certified spore population of 2.7 × 105 CFU per strip, located on the floor of the room (n = 3) or attached to the walls (n = 3), were used as indicators for the sporicidal activity of HPV in the first decontamination experiment. To test the spore survival after HPV exposure, the spore strips were aseptically transferred into tryptic soy broth and incubated at 57℃. Sporicidal activity was demonstrated by the lack of bacterial growth during an incubation time of seven days.
HPV was generated from 30% liquid H2O2 with the help of the HPV Generator Bioquell Q-10 (Bioquell, Andover, UK) which was placed in the centre of the room. The HPV generator delivered HPV into the room and included an instrumentation unit which measured the concentration of H2O2, the relative humidity and temperature of the room air. These parameters were monitored by a computer system placed outside the room. During HPV exposure, the ventilation and air-conditioning system of the room were disabled to prevent the removal and dilution of HPV. When the cycle was completed the catalytic converters R-30 (Bioquell) broke down the HPV into oxygen and water vapour. Access to the room was permitted when the HPV concentration in the room was <1 ppm, as recommended by the manufacturer. The samples were then collected and processed immediately as described above.
Statistical analysis
Two-tailed Student’s t-test was used to determine significance in the differences between the bacterial survival rates in SDW versus 2% BSA. Differences were considered significant when confidence levels of >95% were achieved.
Results
Survival of the bacterial clinical isolates on bedding sheets within the animal room
Relative survival rates of the bacteria on bedding pieces under controlled environmental conditions of the animal room varied considerably depending on the bacterial species and the presence/absence of BSA. Generally, for all bacterial species tested the presence of 2% BSA resulted in a significant increase in the survival rate on the bedding pieces, compared with the water-suspended bacteria. The least resistant bacterial species proved to be the three rodent Pasteurellaceae species tested which died in under 24 h when absorbed onto the bedding pieces in SDW, whereas the presence of 2% BSA increased their survival for several hours (Figure 2). The other non-spore producing bacteria tested were able to survive longer in the animal room compared with the species from the Pasteurellaceae family (Figure 3). P. aeruginosa and Acinetobacter radioresistens survived in high numbers over the six-week testing period (Figure 3A). The Enterobacteriaceae species tested consistently survived over the six-week test period (Figure 3B), whereas the Bordetella species included were able to survive longer than two weeks only in the presence of 2% BSA (Figure 3C). The Gram-positive cocci displayed different survival rates. The most sensitive of them were the β-haemolytic streptococci which survived less than one week on SDW-impregnated bedding and approximately three weeks on BSA-saturated bedding. By contrast, enterococci and staphylococci were able to survive for at least six weeks in the bedding inoculated with BSA-suspended bacteria (Figures 3D and E). The very long survival times recorded for B. safensis were the result of sporulation as demonstrated by the malachite green staining at 12 weeks after bedding inoculation, when only spores and no vegetative bacteria were observed. The bacterial spores were still viable 72 weeks after the start of the experiment. There were no significant differences in survival times of B. safensis in relation to the presence of BSA (Figure 4).
Survival of rodent Pasteurellaceae species on paper bedding pieces in an animal room. Continuous lines indicate that the bacterial inoculum was prepared in 2% bovine serum albumin (BSA), whereas disrupted lines denote inoculum prepared in sterile distilled water. Each datum point represents the mean of the replicates of three bedding pieces for one strain. Error bars represent the standard deviation. Strains designation: Muribacter muris ( Survival patterns of Pseudomonas aeruginosa (♦)/Acinetobacter radioresistens ( Survival kinetics of Bacillus safensis on paper bedding pieces in an animal room. Continuous lines denote survival of bacteria prepared in 2% bovine serum albumin (BSA), whereas disrupted lines indicate preparation in sterile distilled water. Mean and standard deviation of three bedding pieces are shown for each time point. ), [Pasteurella] pneumotropica biotype Jawetz (
), [Pasteurella] pneumotropica biotype Jawetz ( ), [Pasteurella] pneumotropica biotype Heyl (▴).
), [Pasteurella] pneumotropica biotype Heyl (▴). ) (A), Escherichia coli (▪)/Klebsiella oxytoca (
) (A), Escherichia coli (▪)/Klebsiella oxytoca ( ) (B), Bordetella bronchiseptica (
) (B), Bordetella bronchiseptica ( )/Bordetella hinzii (▪) (C), Enterococcus faecalis (♦)/β-haem streptococci () (D), and Staphylococcus aureus ()/Staphylococcus xylosus (▴) (E) on paper bedding pieces in an animal room. Continuous lines indicate survival of bacteria prepared in 2% bovine serum albumin (BSA) on the bedding pieces, whereas disrupted lines show preparation of the bacterial inoculum in sterile distilled water. Each datum point represents the mean of the replicates of three bedding pieces for one strain and the corresponding standard deviation.
)/Bordetella hinzii (▪) (C), Enterococcus faecalis (♦)/β-haem streptococci () (D), and Staphylococcus aureus ()/Staphylococcus xylosus (▴) (E) on paper bedding pieces in an animal room. Continuous lines indicate survival of bacteria prepared in 2% bovine serum albumin (BSA) on the bedding pieces, whereas disrupted lines show preparation of the bacterial inoculum in sterile distilled water. Each datum point represents the mean of the replicates of three bedding pieces for one strain and the corresponding standard deviation.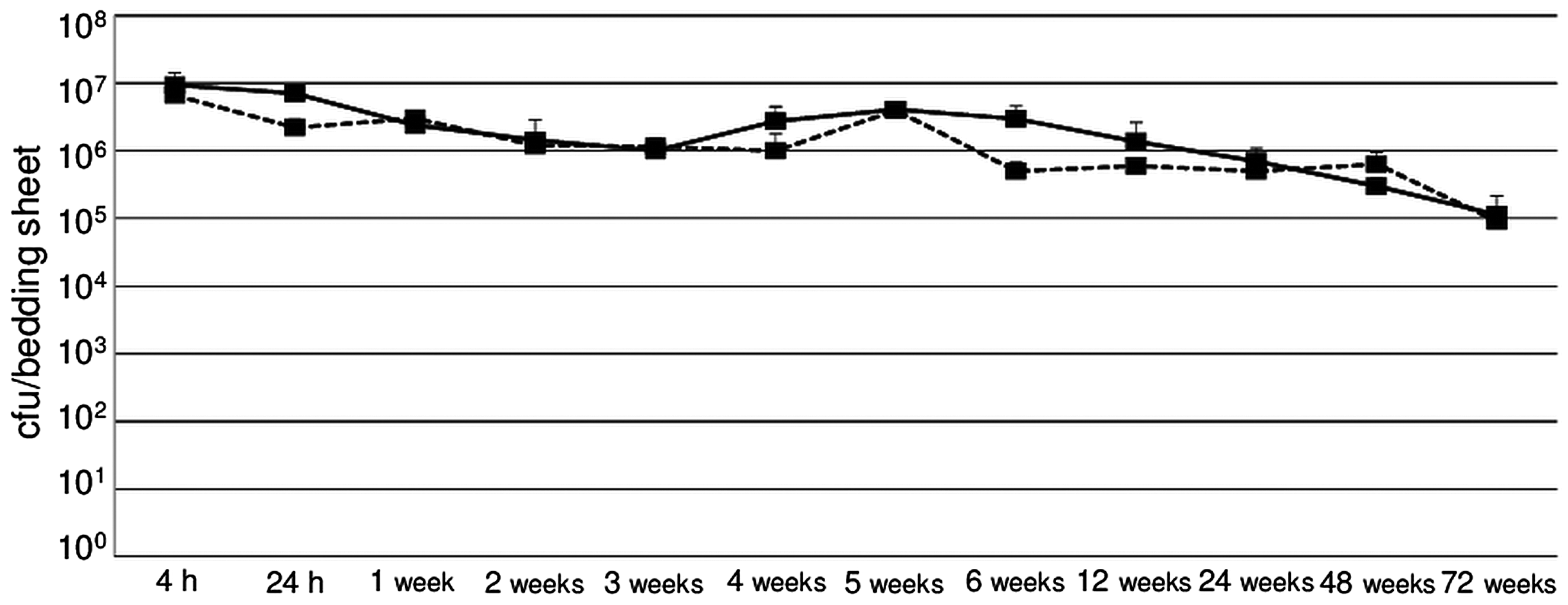
Survival of the bacterial clinical isolates on bedding pieces after exposure to HPV
The cycles used for the decontamination of the room with HPV consisted of an injection phase followed by a dwell phase of 20 min each. Subsequently, the HPV was converted to oxygen and water vapour during the aeration phase which lasted until the HPV concentration dropped to below 1 ppm. The HPV decontamination was also accompanied by changes in temperature and relative humidity. The parameters recorded were similar among the three cycles performed. The data from a representative HPV treatment cycle are shown in Figure 5.
Kinetics of data recoded during a representative hydrogen peroxide vapour (HPV) cycle. Hydrogen peroxide (H2O2) concentration, relative humidity and temperature were registered by the instrumentation unit of the HPV generator and displayed on the control computer system placed outside the room.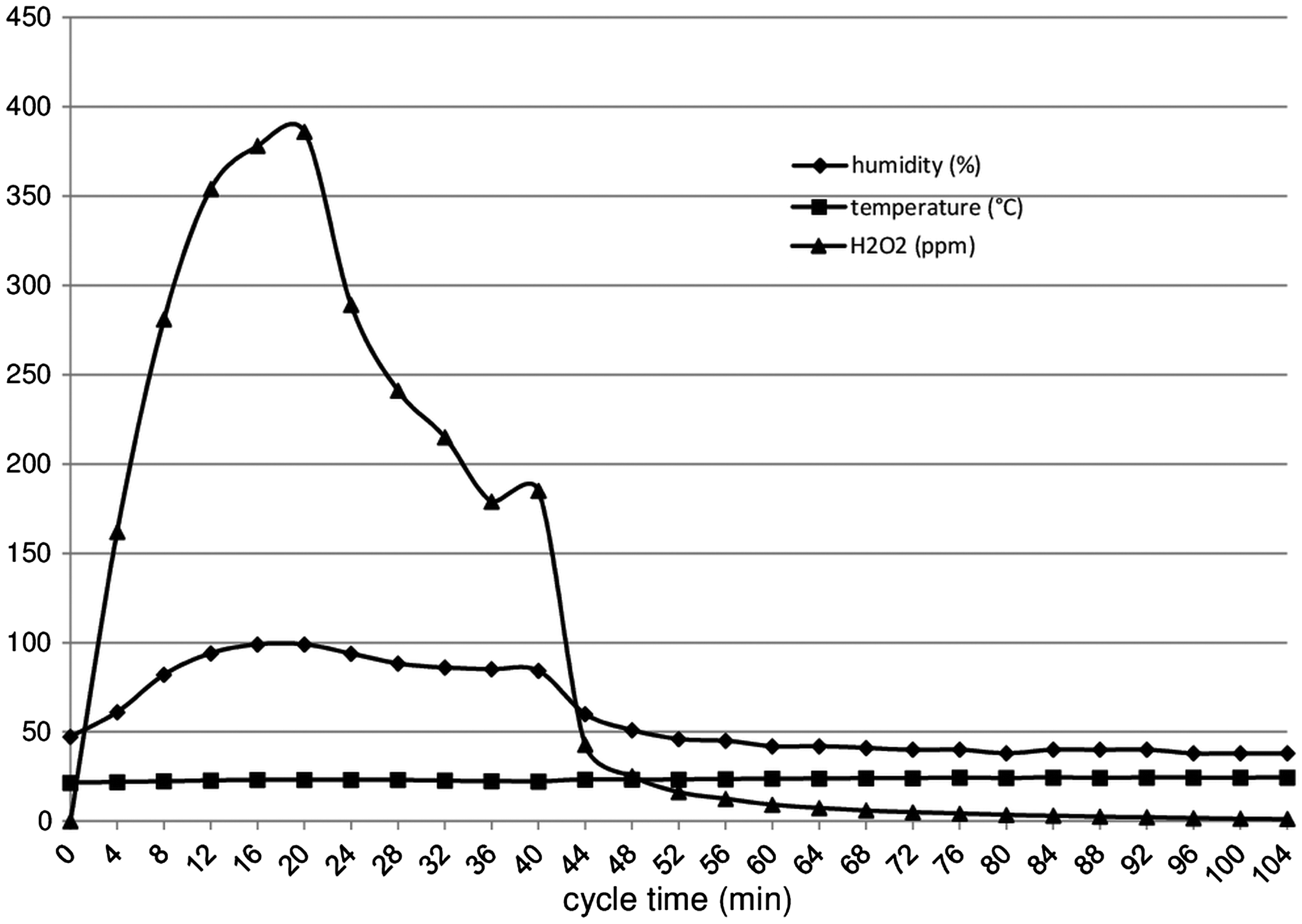
Survival of Staphylococcus aureus, Pseudomonas aeruginosa and Escherichia coli exposed to hydrogen peroxide vapour.
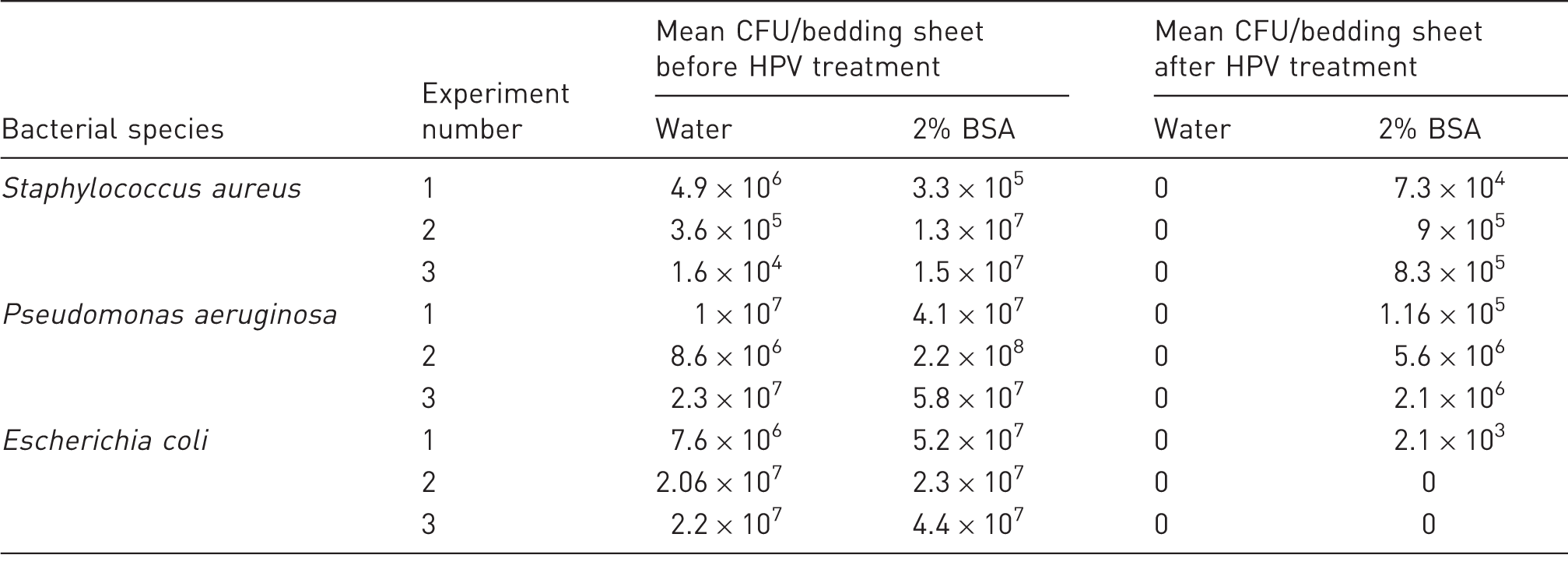
CFU: colony forming units; HPV: hydrogen peroxide vapour.
Discussion
Several of the bacterial species are important in the quality assurance of the SPF status of laboratory rodents. With an increasing number of genetically-engineered animals available, even agents which have recently been regarded as non-pathogenic for wild-type mouse strains have become problematic with some animal strains. 13 Detection and environmental sanitation of such microbes are essential in obtaining and maintaining microbiologically standardized laboratory rodents. In the present investigation we analysed the ability of bacteria relevant to laboratory rodents to survive on bedding particles in the animal room environment, which is essential for microbiological monitoring using soiled bedding sentinels. As expected, we noticed considerable differences in the survival rates of the bacterial strains tested (Figures 2–4). Overall, the presence of 2% BSA, used to simulate biological soiling, led to increased survival rates on bedding pieces for all vegetative bacterial species tested compared with their corresponding water-suspended samples. Our findings are in agreement with those of Coughenour et al. who have found that the presence of BSA enabled MRSA to survive significantly longer on five different environmental surfaces. 14 Rodent Pasteurellaceae species displayed a very short survival time on the bedding particles, suggesting that their transmission to sentinels via the soiled bedding was improbable. This is in agreement with a previous study that emphasized the fragility of [Pasteurella] pneumotropica in the environment and the absence of transmission to sentinels via soiled bedding. 5 The other bacterial test strains were able to survive for at least three weeks on BSA-soiled bedding sheets (Figure 3), thus allowing enough time for a probable transmission to the sentinels. Notably, B. safensis easily survived over one year on the bedding pieces, independent of the presence of BSA, due to its sporulation behaviour (Figure 4). Most of the species tested in the present study are of potential importance for both humans and animals. The environmental stability of some of the bacterial species used in the present study has previously been tested in hospital settings.15,16 However, their stability under animal room conditions on bedding particles was not tested previously.
Numerous studies tested the efficiency of HPV to inactivate relevant human bacterial pathogens on hard surfaces such as stainless steel.15,17 Nevertheless, only a few studies have investigated porous surfaces, such as cotton or carpet, as carriers for the bacteria.18,19 From this perspective, our study on the efficacy of HPV to inactivate bacteria on cellulose bedding is new and has relevance for both laboratory animals and hospital settings, where cellulose/paper materials can be contaminated by such bacteria. The resistance of bacteria to HPV can also vary with the strain, as has previously been shown, 15 where a reference strain of Klebsiella pneumoniae was more sensitive than a clinical isolate. To replicate the natural conditions, our study used clinical isolates recovered exclusively from the same areas that were subjected to HPV disinfection.
HPV readily inactivated S. aureus, P. aeruginosa and E. coli cells on bedding pieces which were inoculated with SDW bacterial suspensions. However, soiling, as simulated by 2% BSA, resulted in increased survival rates of S. aureus and P. aeruginosa, but not of E. coli, when exposed to HPV (Table 2). However, a reduction of approximately 2 log10 CFU/bedding sheet compared with the non-treated controls was observed after HPV treatment (Table 2). Interestingly, the BSA-suspended bacteria which were smeared onto the Petri dish plastic surfaces but not the bedding pieces were readily inactivated by HPV, suggesting that a porous and thicker carrier is necessary to promote bacterial survival even when bacteria are BSA-soiled. Our results concur with those of Otter et al. who have shown that the presence of BSA led to an increased survival rate of S. aureus to HPV inactivation. 20 The present study extends our knowledge regarding the increased survival rate in the presence of BSA also for P. aeruginosa. Similarly, Pitten et al. have shown that the presence of albumin may also contribute to increased survival rates of nosocomial bacteria exposed to liquid disinfectants with H2O2 as the active agent. 21
HPV disinfection should be used after rooms are cleaned. However if the cleaning is suboptimal, microbes embedded in animal secretions and excretions may contribute to the failure of HPV disinfection. From this perspective, future experimental HPV disinfection studies should consider more natural aspects regarding bacterial soiling and carriers, and should not be limited to artificial conditions of water-suspended bacteria that had dried on hard surfaces such as stainless steel.
In summary, our findings show that bacterial species relevant to laboratory animals can survive for different periods of time on bedding pieces under animal room conditions, and that BSA soiling favour bacterial survival in the environment and their resistance to HPV when placed on bedding. Our results indicate that an extensive room cleaning is a pre-requisite for a successful HPV disinfection. Overall, such aspects must be considered when developing standard operating procedures for laboratory rodent colonies.
Footnotes
Acknowledgements
We gratefully acknowledge Sonja Green, Andrea Grunwald, Theresa Ohly and Manuela Stockhausen for their excellent technical assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.