
Abstract
The inter-α-trypsin inhibitor (ITI) family is a group of plasma proteins built up from heavy (HC1, HC2, HC3) and light (bikunin) chains synthesized in the liver. In this study we determined the distribution of ITI constitutive chains in normal and cancerous lung tissues using polyclonal antibodies. In normal lung tissue, H2, H3, and bikunin chains were found in polymorphonuclear cells, whereas H1 and bikunin proteins were found in mast cells. Bikunin was further observed in bronchoepithelial mucous cells. In lung carcinoma, similar findings were obtained on infiltrating polymorphonuclear and mast cells surrounding the tumor islets. Highly differentiated cancerous cells displayed strong intracytoplasmic staining with H1 and bikunin antiserum in both adenocarcinoma and squamous cell carcinoma. Moreover, weak but frequent H2 expression was observed in adenocarcinoma cells, whereas no H3-related protein could be detected in cancer cells. Local lung ITI expression was confirmed by RT-PCR. Although the respective role of inflammatory and tumor cells in ITI chain synthesis cannot be presently clarified, these results show that heavy chains as well as bikunin are involved in malignant transformation of lung tissue.
Keywords
T
To date, the study of ITI immunoreactivity in human tissues has been performed only with antisera directed against the whole ITI molecule or bikunin, and showed a localization of ITI-related proteins and bikunin in mast cells and connective tissues of the bladder (Odum 1989), skin, colon, and lung (Yoshida et al. 1989; Businaro et al. 1992; Odum and Nielsen 1994), kidney, testis, cerebrum, and cerebellum (Businaro et al. 1992). The presence of bikunin was also reported in some brain tumors (Yoshida et al. 1994).
Several functions have been proposed for the heavy and bikunin chains. Bikunin has potent inhibitory effects on the activities of trypsin, chymotrypsin, plasmin, and human leukocyte elastase (Wachter and Hochstrasser 1981; Albrecht et al. 1983). Recently, the potential role of ITI proteins in tumor invasion has been suspected on the basis of their interaction with extracellular matrix components and their protease inhibitory capacity, as reviewed by Bost et al. (1998). An extracellular matrix-stabilizing role (Castillo and Templeton 1993) has been reported for ITI-related proteins in the organization of the mouse cumulusoocyte complex (Camaioni et al. 1993; Chen et al. 1992,1994), in the formation of the hyaluronan-containing coat around cultured cells (Blom et al. 1995), and in the protection of synovial fluid hyaluronan against free radicals (Hutadilok et al. 1988). Moreover, bikunin displayed an in vitro inhibitory activity against urokinase-type plasminogen activators (Kikuchi et al. 1987; Kobayashi et al. 1995a), an important function in the inhibition of tumor invasion (Kobayashi et al. 1994,1995b), and showed a potent growth inhibitory activity towards cultured normal and tumor cells (Chawla et al. 1990; Perry et al. 1994).
In this study we used antibodies specific for each chain to examine the localization of ITI heavy H1, H2, H3, and bikunin chains in human specimens from normal lung and from adenocarcinoma and squamous cell lung carcinoma. Total RNAs extracted from lung tissues were also examined by RT-PCR to assess the expression of these proteins in the lung.
Materials and Methods
Tissues
Blocks of human lung tumors were retrieved from the pathology files of the Rouen University Hospital. The specimens involved adenocarcinoma (n = 3) and squamous cell carcinoma (n = 4), the corresponding normal tissues being sampled in distant histologically normal areas. The samples were fixed with 4% paraformaldehyde in PBS for 24 hr at 4C, dehydrated, and embedded in paraffin.
Antibodies
Polyclonal antiserum against bikunin was produced in rabbits by immunization with proteins purified from normal human urine (Salier et al. 1983). The ITI anti-H antisera were raised in rabbits with H1, H2, or H3 heavy chain peptides produced in a bacterial expression system. Their specificity and reactivity have previously been extensively established by immunoprecipitation and immunofluorescence on hepatoma cells in culture (Heron et al. 1994,1995a), as well as immunoprecipitation of translation products and Western blotting (Heron et al. 1995b).
Immunohistochemistry
Paraffin-embedded tissues were cut into 4-μm sections, placed on silanized slides, then treated with xylene to remove the paraffin and rehydrated through graded alcohol. After washing with PBS containing 0.05% Tween-20, the sections were placed in a humidified chamber and preincubated with normal human or goat serum for 30 min at room temperature (RT) to block nonspecific binding. The sections were then incubated with the primary antibodies (crude antisera) diluted 1:20 with Tris-buffered saline (TBS: 0.1 mol/liter, pH 8.2; Dako, Trappes, France) for 1 hr at RT, followed by a second-step incubation with alkaline phosphatase-labeled goat anti-rabbit immunoglobulins (Dako) diluted 1:40 for 30 min at RT. After washing in TBS, the alkaline phosphatase activity was detected with the Fast Red substrate system (Dako), containing naphthol phosphate, Fast Red, and levamisole in TBS. Optimal labeling was checked under the microscope during the final reaction step. Sections were finally washed under tapwater, counterstained with hematoxylin (Dako), and mounted with Faramount aqueous medium (Dako). Control experiments for staining specificity included replacement of primary antibody with preimmune rabbit serum or TBS alone. To visualize tissue mast cells, the adjacent serial sections were stained with 0.1% Alcian blue (Sigma; St Louis, MO) in acetic acid, pH 2.4, for 20 min at RT.
Microscopy
Immunostaining was studied using an Axiophot microscope (Zeiss) equipped with a 3CCD camera (Sony) and then analyzed with SAMBA 2005 (Système d'Analyse Microphoto-métrique à Balayage Automatique, Alcatel; TITN, Grenoble, France), an image processing system (Brugal et al. 1979). On final pictures, the immunostaining positivity graded from low (yellow) to high (brown-red), with background in faint pink.
PCR primer characteristics

aBase numbering or predicted size according to cited references.
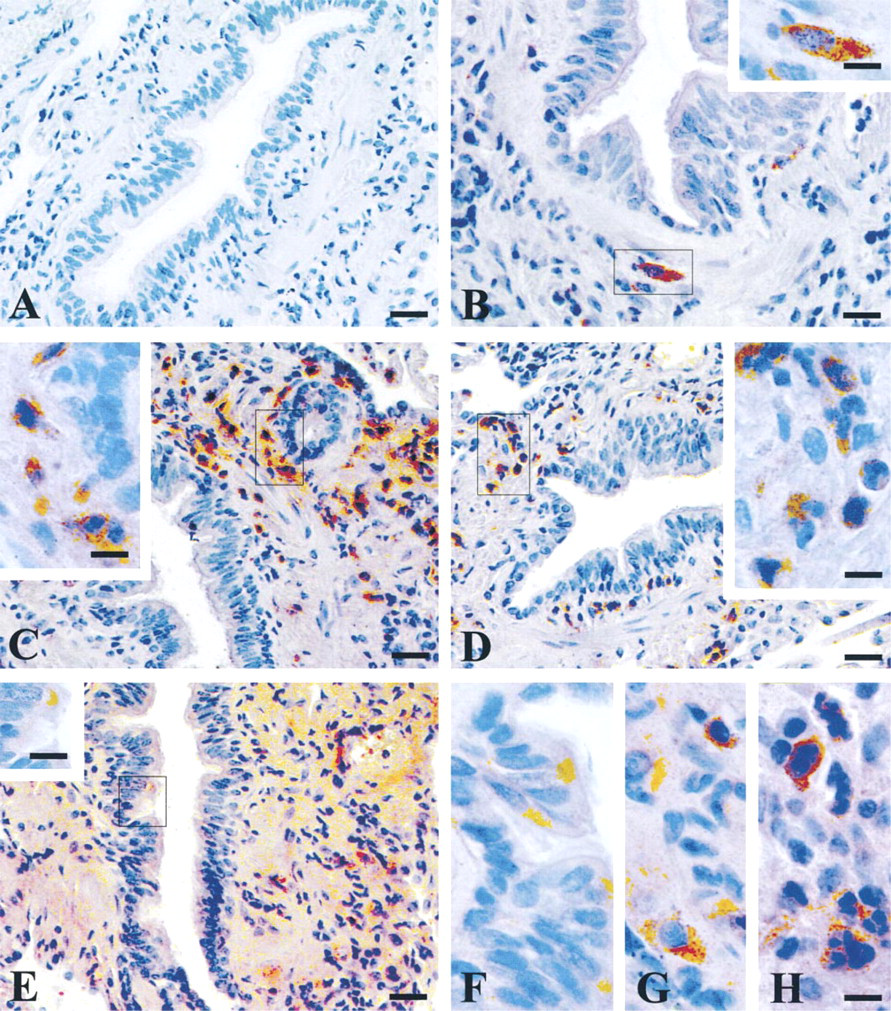
Immunohistochemical detection of ITI heavy and light chains in histologically normal human lung tissue. Tissue sections were immunostained with (A) preimmune rabbit serum, (B) H1 antiserum, (C) H2 antiserum, (D) H3 antiserum, and (E-H) bikunin antiserum. F,G, and H correspond to enlarged selected areas in bikunin-stained sections. Bars: A-E = 10 μm; F-H and insets = 3.2 μm.
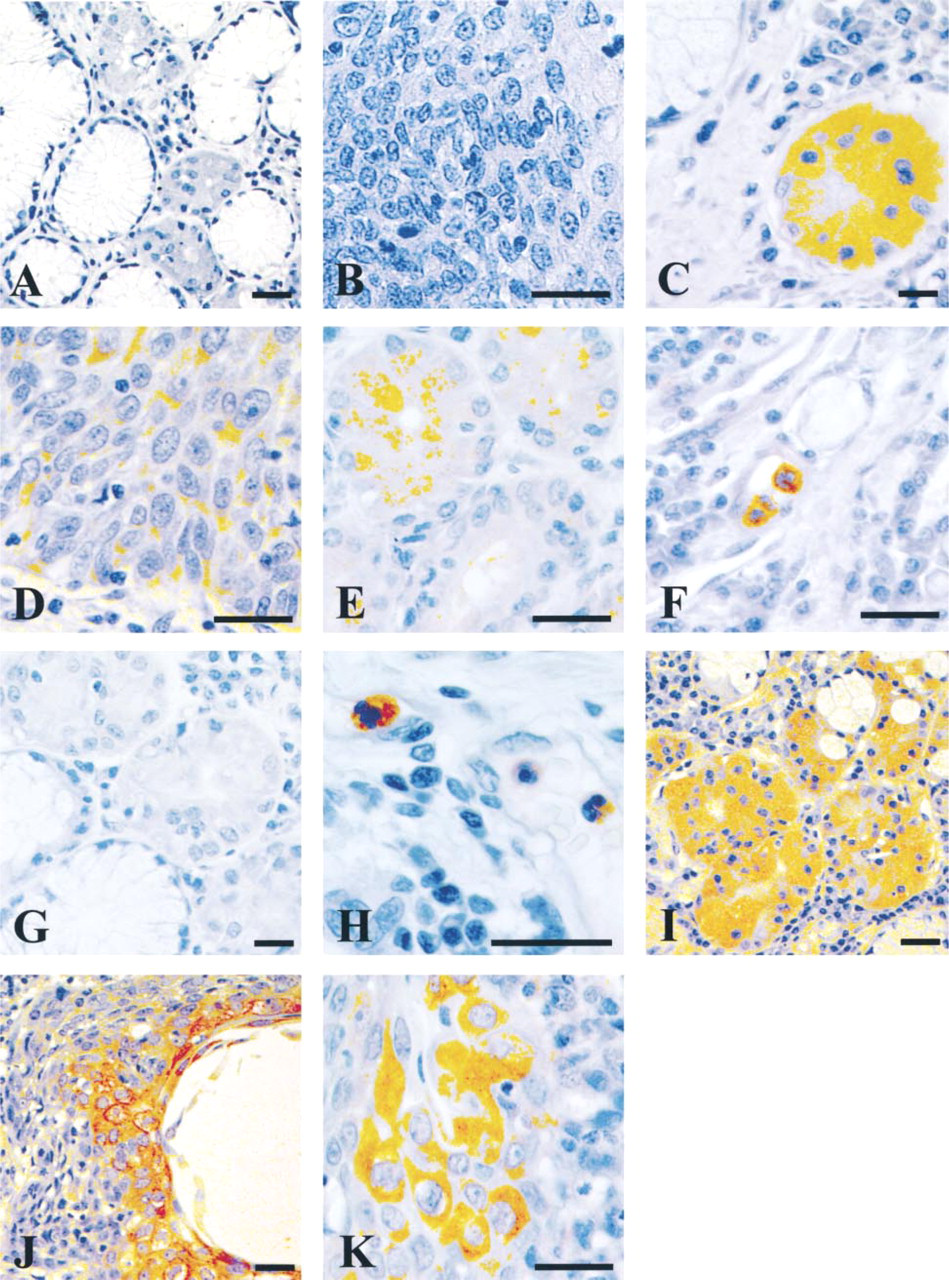
Immunohistochemical detection of ITI heavy and light chains in tumoral human lung tissue: adenocarcinomas (A,C,E,G,I,J) and squamous cell carcinoma (B,D,F,H,K). Tissue sections were immunostained with (A,B) preimmune rabbit serum, (C,D) H1 antiserum, (E,F) H2 antiserum, (G,H) H3 antiserum, and (I-K) bikunin antiserum. Bars = 10 μm.
Reverse Transcriptase Polymerase Chain Reaction
Total RNAs were extracted from liquid nitrogen-frozen normal and tumor lung tissue using the guanidinium isothiocyanate procedure followed by centrifugation through a 5.7 M CsCl cushion as described (Ullrich et al. 1977). Total RNAs (1 μg) from lung tissue or HepG2 hepatoma cells were incubated with 5 U of Tth DNA polymerase (Eurobio Laboratories; Les Ulis, France) in a final volume of 100 μl according to protocols recommended by the supplier. The cDNA was synthesized with antisense primer (Table 1) using one cycle of 58C for 15 min and 70C for 15 min. The cDNA reactions were amplified after sense primer addition using one cycle of denaturation at 95C for 2 min and 35 cycles consisting of denaturation at 95C for 1 min, primer annealing at 58C for 1 min, extension at 72C for 1 min; an additional cycle at 95C for 1 min, 60C for 1 min, 72C for 7 min was performed, and the reactions were stopped by chilling to 4C. The negative control did not contain any template. After ethanol precipitation, the whole PCR products were analyzed by 6% polyacrylamide gel electrophoresis with ethidium bromide staining.
DNA Sequence analysis
Purified PCR products were cloned into the SmaI site of pUC18 vector. DNA sequencing was performed using the dideoxy chain termination method (Sanger et al. 1977). The sequences were aligned and decoded with the Microgenie sequence analysis program (Queen and Korn 1984).
Results
Immunohistochemical Staining of Normal Lung Tissue
The distribution of ITI constitutive chains was studied on serial paraffin sections from tumor-free regions of lung tissue. The sections of lung tissue contained large segmental bronchi as well as smaller subsegmental bronchi, alveolar walls, and medium-sized pulmonary arteries. Expression of bikunin was observed in the apical pole of some mucous cells of the bronchiolar epithelium and in the underlying connective tissues (Figure 1E and 1F). Strong immunohistochemical staining for H2, H3, and weaker staining for bikunin were observed in the inflammatory granulocyte cells scattered in the interstitial connective tissue (Figures 1C-1E and 1H) as well as in the lumen of blood vessels (not shown). The negative reaction of these cells with nonimmune rabbit serum (Figure 1A) or anti-H1 antibody (Figure 1B) showed the specificity of this finding. In contrast, H1 immunoreactivity was found to be associated with mast cells (Figure 1B), identified by their positive Alcian blue staining in adjacent sections (not shown). Mast cells also stained positive with bikunin (Figure 1E and 1G). No signal could be observed with either anti-H antiserum in the bronchial epithelium or in alveolar walls.
Immunohistochemical Staining of Lung Cancer Tissues
As a first step towards determining how ITI chain expression is altered during the malignant process, we examined multiple lung tissue sections from three adenocarcinomas and four squamous cell carcinomas. The polymorphonuclear cells infiltrating the tumor stroma were strongly stained with H2 and H3 antisera (Figure 2F and 2H). No immunostaining for H2 and H3 was found in the tumor cells of squamous cell carcinomas. In contrast, highly differentiated cancer cells were weakly stained with H1 (Figure 2D). The mast cells were strongly stained with H1 antibody in one of the squamous cell carcinomas studied (not shown). The strongest staining was observed with bikunin in highly differentiated cells (Figures 2I-2K) from squamous cell carcinomas and adenocarcinomas. Detectable H2, H1, and bikunin immunostaining was observed in adenocarcinomas, with some variability in the staining intensity: strong with bikunin (Figure 2I) and H1 (Figure 2C), weak with H2 (Figure 2E). For H3, all adenocarcinoma cells were negative (Figure 2G). The staining covered the whole cytoplasm for H1 and bikunin but was predominantly apical for H2. Positive staining was not observed in a control section incubated with normal rabbit serum or TBS alone (Figure 2A and 2B).
RT-PCR
To determine whether the immunohistochemical staining observed in lung tissue sections corresponded to simple adsorption of proteins or to local synthesis by lung tissues, total lung RNAs extracted from the corresponding normal and tumor tissues were analyzed using RT-PCR. The PCR products obtained with tumor (Figure 3, Lanes 3) and normal human lung tissue (Figure 3, Lanes 4) displayed sizes similar to those observed with HepG2 RNAs (Figure 3, Lanes 1), corresponding to the expected values, i.e., 138 bp (H1), 127 bp (H2), 318 bp (H3), and 294 bp (bikunin). Sequence analysis of the amplified products confirmed that they consisted of authentic H1, H2, H3, and bikunin cDNAs. A semiquantitative comparison of ITI expression in HepG2 cells (Figure 3, Lanes 1) and nontumor lung (Figure 3, Lanes 4) or lung cancer tissues (Figure 3, Lanes 3) showed equivalent levels of H1 and H3 mRNA transcripts but a lower level of H2 and bikunin in lung compared to hepatoma cells.
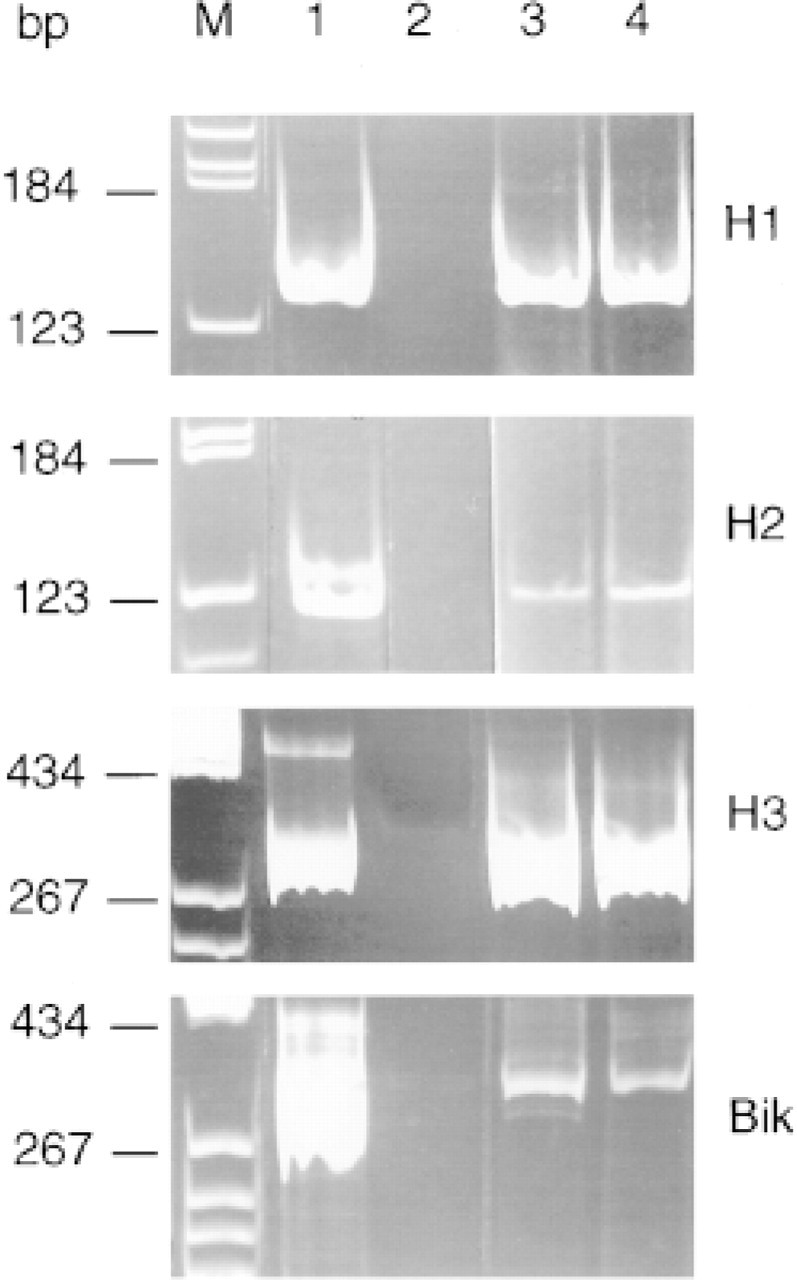
Amplification products obtained from RT-PCR as described in Materials and Methods. Lane M, DNA molecular weight markers; Lane 1, HepG2 RNAs; Lane 2, no RNA (negative control); Lane 3, human lung adenocarcinoma RNAs; Lane 4, nontumorous lung RNAs.
Discussion
The presence of bikunin was previously reported in a variety of human and rodent tissues, i.e., brain, liver, kidney, intestine, and pancreas (Wakayama et al. 1996; Itoh et al. 1996; Odum and Nielsen 1994; Yoshida et al. 1994), in which the positive immunoreactivity was attributed to a reabsorption of plasma ITI. In the present work, we used antibodies specific for each heavy and light chain of the ITI family to localize H1, H2, H3, and bikunin in human normal and tumor lung tissues. In normal lung, we found intense immunoreactivity for bikunin in scattered goblet cells of the bronchial epithelium and in polymorphonuclear and mast cells of the underlying connective tissue. These results are in line with those of Yoshida et al. (1989), who described the presence of a bikuninrelated molecule, the acid-stable trypsin inhibitor (ASTI), in bronchial epithelial cells; they further provide evidence that bikunin is not restricted to mucus, as proposed by Businaro et al. (1992).
Taking advantage of specially designed antisera, we could clarify the ITI immunoreactivity of mast cells described by Odum and Nielsen (1994). Strong immunostaining was observed for bikunin but also for H1. The latter component could not be originally demonstrated because of the immunoabsorption procedure used and/or the low reactivity of the antiserum towards H1 chains. Mast cells may therefore be able to synthesize an HC1/bikunin species, although such an association has not been reported thus far.
In the present study we demonstrate for the first time a strong immunostaining of polymorphonuclear cells for H2, H3 and, to a lesser degree, for bikunin. The question of whether this immunostaining is a widespread phenomenon or results from local lung activation of polymorphonuclear cells cannot be clarified at this stage. However, preliminary experiments from our laboratory using peripheral blood cell smears also showed immunostaining with H2, H3, and bikunin antisera in some leukocytes, suggesting a constitutional expression, whereas no labeling could be observed with H1 antiserum (unpublished data).
In non-small-cell lung cancer tissue, we found positive immunostaining for bikunin in stromal cells, as shown by Yoshida et al. (1989), and also clear cytoplasmic immunostaining in more differentiated adenocarcinoma and squamous cancer cells. Such a positive cytoplasmic immunostaining was also found for H1 and H2 in adenocarcinoma cells and for H1 in squamous cancer cells.
In a previous work, Okumichi et al. (1984) were able to extract bikunin from lung cancer tissue and suggested that it corresponded to a production by cancer tissue rather than to a reabsorption from plasma. We have demonstrated here by RT-PCR analysis a local transcription of all ITI components in lung tissue. These mRNAs could be produced by infiltrating blood cells or lung epithelial or stromal cells. When each ITI chain is considered, the transcription levels observed in cancer tissue did not differ from those seen in non-tumor lung tissue. However, the levels of H2 and bikunin transcripts observed in lung tissue by RT-PCR were lower than those of H1 and H3. The discrepancy between the low bikunin mRNA levels and the strong immunohistochemical staining observed in bronchial epithelium and inflammatory cells could be explained by a low mRNA stability. In a separate study, we observed that H2 and bikunin transcripts obtained by RT-PCR from human normal spleen were mainly unstable non-poly (A) mRNAs, while H1 transcripts were present as poly (A) mRNAs (unpublished observation). We believe that ITI family genes are constitutively expressed in the liver as stable poly (A) RNAs, whereas their transcription in other tissues could respond to an inflammatory or a tumor process and result in unstable non-poly (A) RNAs. Such a difference in mRNA stability is likely to explain our previous results (Salier et al. 1987), in which Northern blot analysis could find ITI mRNAs only in the liver, whereas those from Itoh et al. (1996) showed detectable bikunin mRNA in lung by RT-PCR analysis.
The present study has established a local synthesis of bikunin, H1, H2, and H3 chains in human normal and tumorous lung. It should be noted that in plasma, ITI is only present as HC1/HC2/ bikunin or Pαd (HC3/bikunin). The presence of H2, H3, and bikunin chains in polymorphonuclear cells suggests that a local synthesis of ITI-like protein (HC2/bikunin) may take place in the lung. Such a tissue-specific expression of HC2/bikunin has been described in hepatoma cells (Heron et al. 1994). Further studies are needed to assess the potential prognostic value of ITI-related protein immunoreactivity in lung cancer, as well as their role in the early stages of bronchial transformation and/or invasion.
Footnotes
Acknowledgements
Supported by INSERM grant PARMERCA9611.
We wish to thank Ms E. Abdelouhab for preparation of tissue sections, Mr M. Fraterno for SAMBA analysis, Ms N. Porchet for secretarial assistance, and Ms A. Brickwood for careful reading of the manuscript.