
Abstract
The Menkes copper ATPase (MNK) is a copper efflux ATPase that is involved in copper homeostasis. Little is known about the intracellular localization and cell-specific function of the MNK in human tissues. To investigate a possible role for this protein in lactation, we measured its expression in sections of tissue from nonlactating and lactating human breast. Western blot analysis showed that MNK expression was greater in lactating tissue than in nonlactating tissue. By confocal immunofluorescence, the MNK was detected in luminal epithelial cells of the alveoli and ducts but not in myoepithelial cells. In the nonlactating breast epithelial cells, the MNK had a predominantly perinuclear distribution. In lactating breast tissue, the distribution of the MNK was markedly altered. Lactating epithelial cells showed a granular, diffuse pattern, which extended beyond the perinuclear region of the cell. This pattern was similar to that observed in a previous study in which cultured CHO cells were exposed to high copper concentrations. Our results suggest that relocalization of the MNK is a physiological process, which may be mediated by copper levels in the breast or by hormones and other events taking place during lactation. A vesicular pathway for copper from the Golgi into milk, similar to that of calcium, is proposed.
The intracellular distribution of MNK in cultured cells has suggested a mechanism for copper efflux. MNK is localized to the trans-Golgi network (Petris et al. 1996; Yamaguchi et al. 1996; Dierick et al. 1997) and is also found in vesicles and, to a lesser extent, at the plasma membrane (Petris et al. 1996; Yamaguchi et al. 1996; La Fontaine et al. 1998). It is proposed that copper is transported into the trans-Golgi network by MNK and then via vesicles to the plasma membrane, where it is exocytosed. Evidence to support this hypothesis is that copper treatment of cultured CHO cells stimulated the redistribution of MNK from the Golgi to the more peripheral regions of the cell (Petris et al. 1996). These observations suggest that copper-induced trafficking may be a mechanism for maintaining cellular homeostasis in the presence of high copper concentrations (Petris et al. 1996).
It is possible that, as an efflux pump, MNK is involved in copper transport for reasons other than that of detoxification. Secretion of copper from epithelial cells of the breast during lactation is a process that requires a copper efflux mechanism. The primary function of mammary epithelial cells is to secrete a wide range of nutrients during lactation, including copper, which is essential for the growing neonate. However, little is known of the mechanisms by which copper is transported into milk. We found evidence of a role for MNK in the breast in a previous study in which the MNK gene was expressed in a human breast carcinoma line (Ackland et al. 1997). MNK was detected in a perinuclear compartment, most likely the trans-Golgi network, and was also more diffusely distributed in the cytoplasm, possibly associated with vesicles.
In this study we have used confocal microscopy to determine whether MNK is expressed in the human breast and whether there are differences in its expression pattern between nonlactating and lactating tissue, which might be suggestive of a role for MNK in secretion of copper into milk. We have also determined the intracellular location of MNK in tissues, as this has not previously been reported. All studies, apart from one report of MNK distribution in mouse (Grimes et al. 1997), were carried out using cultured cells. Our study finds that MNK expression is greater in the lactating breast compared to the nonlactating breast and that the intracellular distribution of MNK is altered in the lactating breast.
Materials and Methods
Tissue from normal nonlactating and normal lactating breast was obtained from breast biopsies performed for diagnosis of breast disease. Tissue was immediately frozen at −80C until use.
Western Blot Analysis
Breast tissue (100 mg) was disrupted in liquid nitrogen and sonicated (15 pulses, 40% power output, 30% duty cycle) in 1 ml of protease inhibitors (Boehringer Mannheim cat. no. 1836 170; Roche Diagnostics, Sydney, Australia), 20 mg/ml SDS, and 5 mM β-mercaptoethanol (Sigma-Aldrich; Sydney, Australia), then centrifuged for 5 min at 18,000 × g. The supernatant (300 μg protein) was fractionated by SDS/PAGE (7.5% polyacrylamide) using a Bio-Rad Mini Protean Gel system according to manufacturer's instructions. Proteins were transferred to nitrocellulose membranes at 4 mA/cm2 for 120 min using a BioRad Mini Trans-Blot in 25 mM Tris and 192 mM glycine, pH 8.3. After blocking with 1% casein in TBST (0.05 M Tris, 0.15 M NaCl, and 0.1% Tween-20), the membrane was exposed to the MNK antibody 1:500 dilution in blocking buffer for 2 hr or α-smooth muscle actin antibody 1:200, then washed in blocking buffer (four times for 15 min). MNK and α-smooth muscle actin were detected using a 1:1000 dilution of a horseradish peroxidase-conjugated goat anti-rabbit antibody and a chemiluminescent detection kit (Boehringer Mannheim cat. no. 1500798; Roche Diagnostics) according to the manufacturer's instructions. The membranes were placed in contact with Kodak XAR X-ray film for 30–60 sec. Extracts of cells known to express MNK protein, PMC42, a human breast carcinoma line, and the mouse fibroblast 3T3 L1 line were used as positive controls for the Western blot analysis and were prepared as previously described (Ackland et al. 1997).
Antibodies
MNK antiserum was raised against the six metal binding domains of the protein and affinity-purified as previously described (Ackland et al. 1997). The TGN38 antibody was kindly supplied by Dr K. Stanley (unpublished data). Monoclonal antibodies (MAbs) to β-casein and α-lactalbumin were purchased from Harlan Sera-Lab (Melbourne, Australia) and MAb to α-smooth muscle actin was purchased from Sigma-Aldrich.
Sectioning
Tissue blocks (1 cm3) were immersed in OCT (Tissue Tek; Melbourne, Australia) and frozen in liquid nitrogen for 3 min, then sectioned (Leica CM 1800 cryostat) at −17-20C. Sections of 8–10-μm thickness were collected on gelatin (5%)-coated slides.
Hematoxylin and Eosin Staining
Tissue sections were air-dried for 20 min, then fixed in 4% paraformaldehyde for 10 min at room temperature (RT) (Humason 1972) and hydrated through ethanol (100%, 70%, 50%, 20%) to deionized water, 1 min in each solution. Instant hematoxylin stain was applied to the sections for 5 min, followed by a deionized water rinse. Sections were blued in Scott's tapwater for 20 sec, rinsed in deionized water, and differentiated in 1% acid alcohol for 2 sec. After a deionized water rinse, yellow eosin stain was applied to the sections for 2–3 min, washed off in deionized water, and the sections were dehydrated in ethanol series (20%, 50%, 70% and 100%), each for 1 min. DPX was added and a coverslip applied.
Immunohistochemistry
Samples from nonlactating and lactating breast were processed concurrently. The sections were dried and fixed in paraformaldehyde as described above, rinsed twice for 10 min in PBS, then permeabilized with 5% Triton X-100 in PBS for 5 min and blocked with 3% BSA (bovine serum albumin) in PBS for 90 min. Primary antibodies diluted in 1% BSA in PBS (1:20 for MNK, 1:100 for β-casein, and 1:100 for α-lactalbumin, 1:50 for α-smooth muscle actin, and 1:1000 for preimmune mouse or rabbit antiserum) were applied to tissues for 2 hr at RT. After three PBS washes, second antibody DTAF (dichlorotriazinyl amino fluorescein-conjugated) AffiniPure donkey anti-rabbit IgG (Jackson Immunoresearch Laboratories; Sydney, Australia) or sheep anti-mouse FITC (Sigma-Aldrich) 1:200 dilution in 1% BSA in PBS was applied for 2 hr. Sections were washed twice for 10 min in PBS and, where indicated, ethidium bromide (1 μg/ ml) was added for 5 min to lightly stain the nuclei. A drop of Fluoroguard (BioRad; Sydney, Australia) was added to the sections and a coverslip applied. Epifluorescence was viewed with an Olympus BX50 microscope with a PlanApo × 60 1.4 oil objective. Confocal images were collected using an Optiscan F900e system in a 512 × 512 pixel format.
Results
Western Blot Analysis
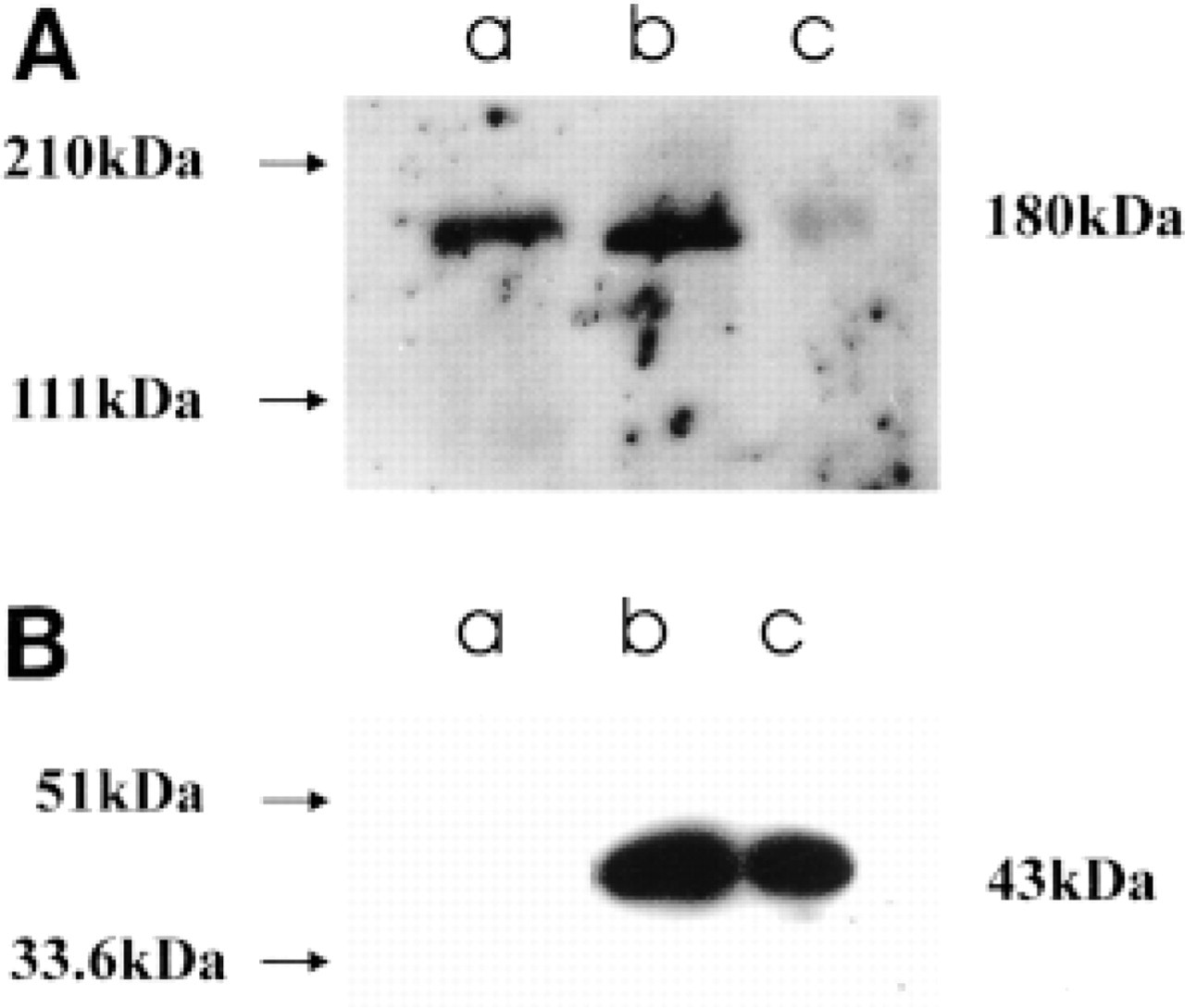
To determine whether MNK was expressed in the breast tissues and to ensure the specificity of the antiserum, tissue extracts were analyzed by Western blotting (Figure 1A). A single band of 180 kD was observed in extracts from both lactating and nonlactating tissue (Figure 1A, Lanes b and c). This size was the same as the MNK band obtained from extracts of the cultured mammary carcinoma line PMC42 (Figure 1A, Lane a) and was consistent with previous reports for human MNK (Yamaguchi et al. 1996; Ackland et al. 1997). The MNK antibody produced a much stronger signal in lactating breast tissue (Figure 1A; Lane b) than in nonlactating breast tissue (Figure 1A, Lane c). Antibody to α-smooth muscle actin was used to show the relative amounts of epithelial tissue present in each extract (Figure 1B), because α-smooth muscle actin expression is not altered with lactation. Similar amounts of α-smooth muscle actin, 43 kD, were present in lactating (Figure 1B, Lane b) and non-lactating (Figure 1B, Lane c) tissue, respectively, whereas PMC42 cells (Figure 1B, Lane a) were negative for α-smooth muscle actin.
Structure of the Nonlactating Breast and Distribution of MNK in Epithelial Cells
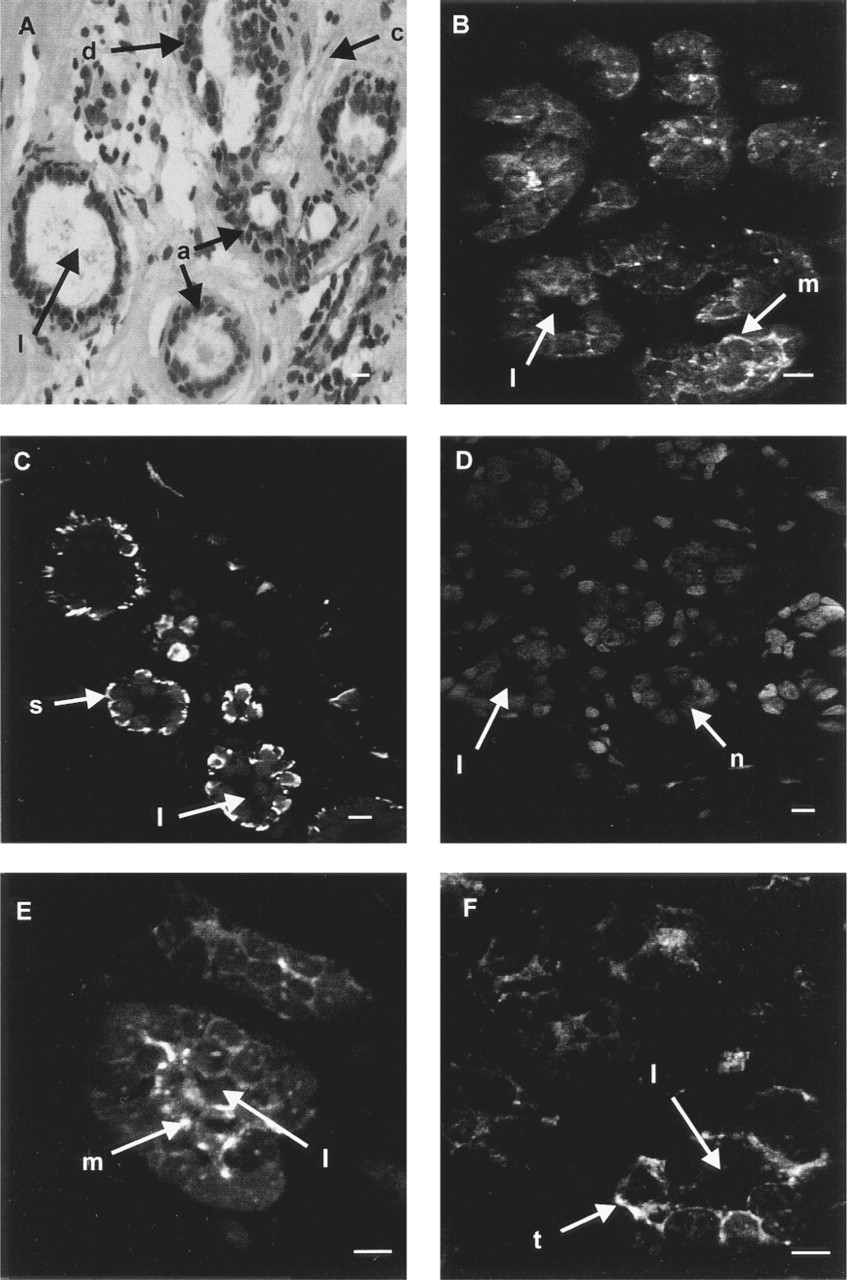
The human breast contains approximately 15–20 lobes, separated by connective tissue and fat. The lobes are composed of lobules that contain alveoli into which milk is secreted. The alveoli of each lobule are drained by a single duct that is conveyed to the exterior. Hematoxylin and eosin staining was performed to show the structural organization of the breast tissue. A section through part of a lobule from nonlactating breast tissue shows several alveoli and a duct, distinguished by its irregular shape, all of which are lined with an inner layer of luminal epithelial cells and an outer layer of myoepithelial cells (Figure 2A).
A section through a lobule was labeled with the MNK antibody and this showed that most of the cells lining the lumen were labeled, although some only faintly (Figure 2B). MNK was not found in the myoepithelial cells, which were identified with an antibody to α-smooth muscle actin (Gugliotta et al. 1988; Rudland and Hughes 1989) and appeared as a bright band of fluorescence adjacent to the luminal cells (Figure 2C). Ethidium bromide was used to stain nuclei in Figure 2C and showed the position of the luminal epithelial cells that did not label with α-smooth muscle actin antibody. A control section labeled with preimmune serum and ethidium bromide showed no specific fluorescence (Figure 2D).
At higher magnification, the perinuclear distribution of MNK in luminal epithelial cells was more clearly shown (Figure 2E). The fluorescence was often localized to one side of the nucleus, although not necessarily the luminal side. The distribution of MNK was similar to that of the fraus-Golgi marker TGN38, which was also perinuclear (Figure 2F).
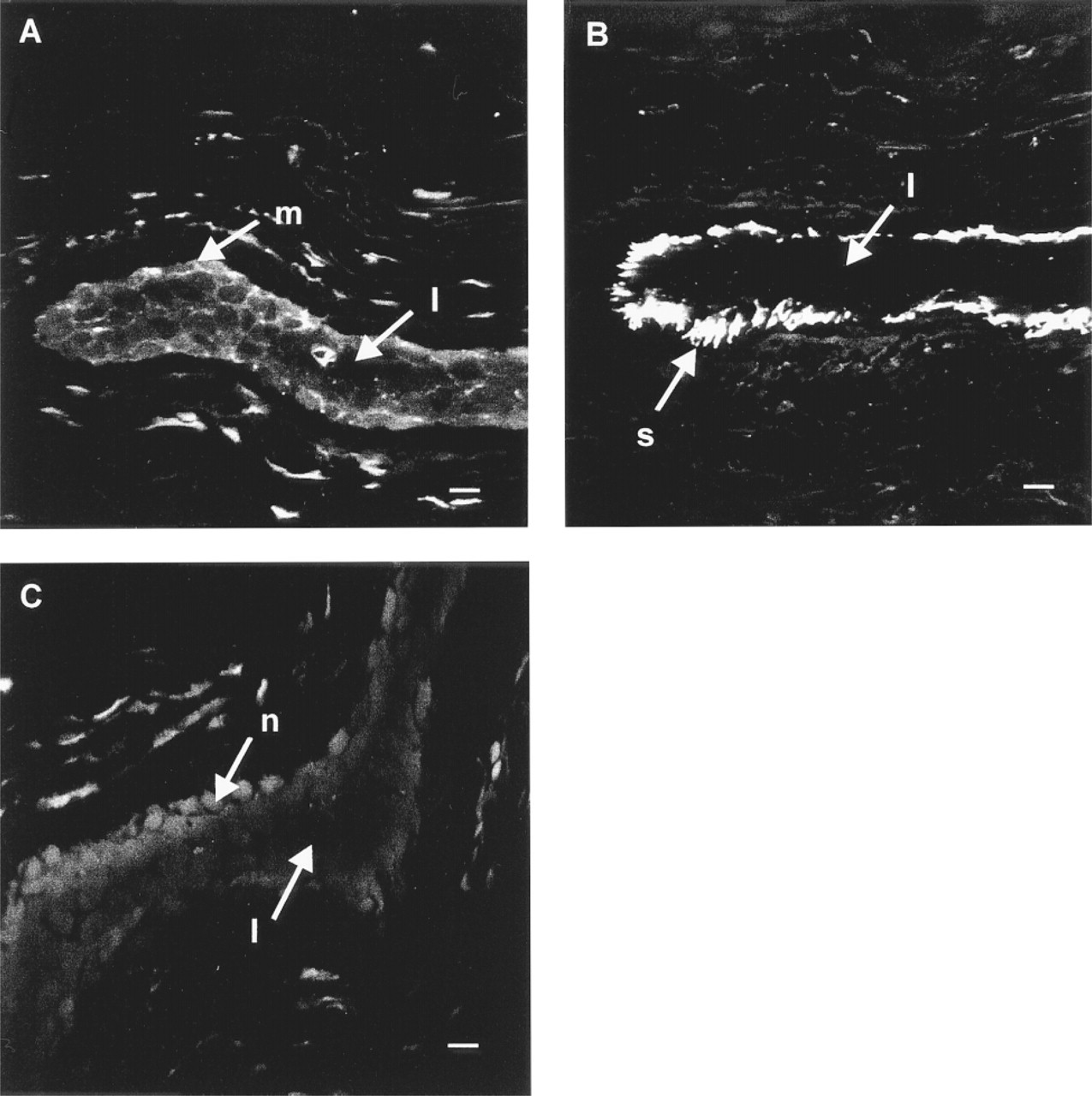
In the human breast, the arrangement of cells within ducts is similar to that found in alveoli, i.e., an inner layer of luminal secretory cells with myoepithelial cells located distally from the lumen. A longitudinal section through a duct showed MNK expression to be localized to the luminal cells (Figure 3A) but not in the myoepithelial cells. The myoepithelial cells identified with antibody to α-smooth muscle actin (Figure 3B) were negative for MNK. A section treated with preimmune serum and stained lightly with ethidium bromide to indicate structure is shown (Figure 3C). Nonspecific labeling of the connective tissue can also be seen in these images.
(
(
(
Structure of the Lactating Breast and Distribution of MNK in Epithelial Cells
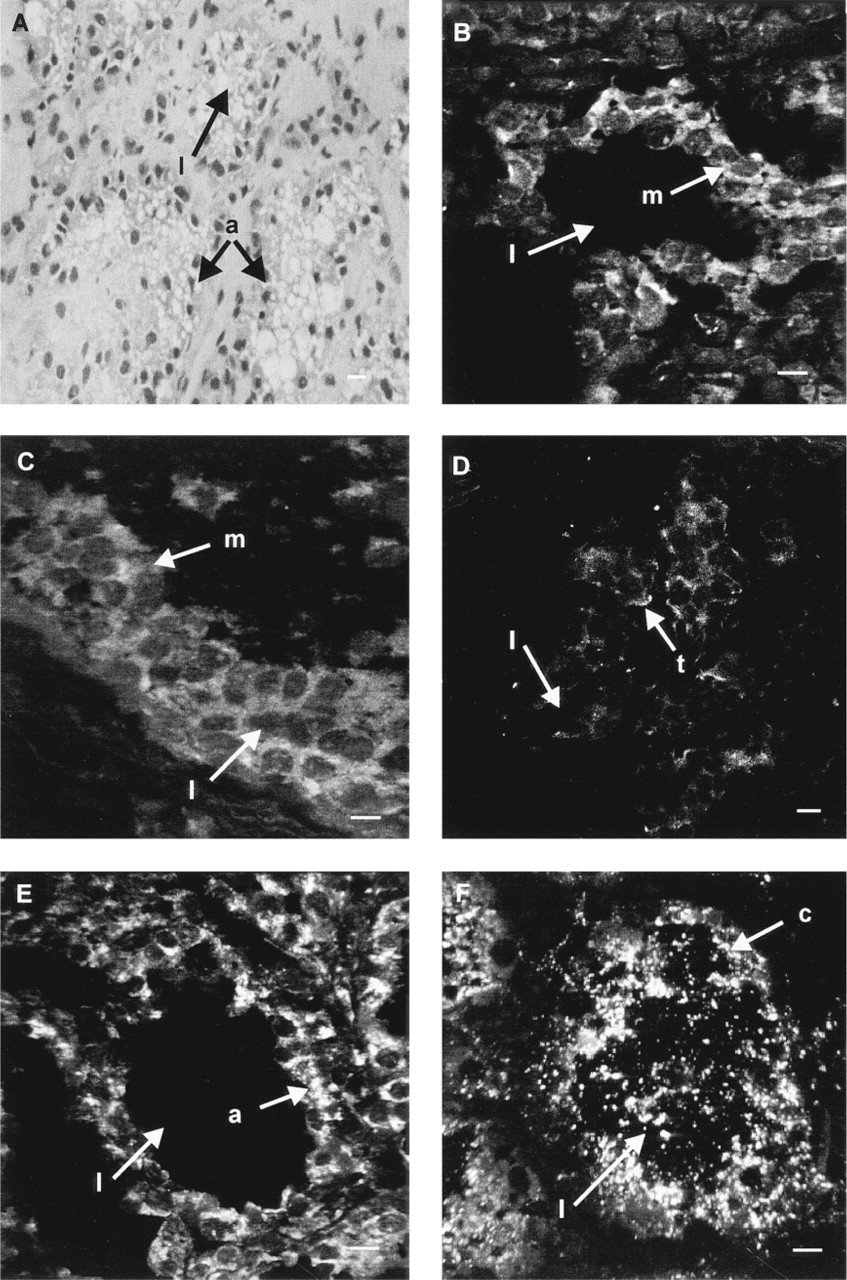
Sections of lactating breast tissue were stained with hematoxylin and eosin to demonstrate the extended alveoli and ducts and the expanded lumina that are characteristic of the lactating breast (Figure 4A). The intracellular distribution of MNK was markedly different in lactating tissues. In place of the compact perinuclear labeling seen in nonlactating tissue, MNK in lactating cells appeared as a granular, diffuse pattern that extended beyond the perinuclear region of the cell in both alveoli (Figure 4B) and ducts (Figure 4C). This pattern was reminiscent of the distribution of MNK in cultured CHO cells exposed to high copper concentrations (Petris et al. 1996). In addition, the intensity of fluorescence was increased in lactating tissue relative to nonlactating tissue. In contrast to the dispersed label of MNK, TGN38 remained localized to the perinuclear region (Figure 4D), showing that the change in MNK distribution was not simply due to altered TGN distribution associated with lactation.
Two of the major milk proteins of the lactating breast, β-casein and α-lactalbumin, were also detected by immunofluorescence (Figure 4E and 4F). As with MNK, the luminal epithelial cells were labeled. The intensity of fluorescence seen with the milk protein antibodies in tissues from the lactating breast was much greater than that of MNK and extended throughout the cytoplasm of the cells. Milk proteins were also detected in the lumen of most alveoli, although sometimes present in small amounts that did not show up in the images, as for α-lactalbumin in Figure 4F. In contrast, MNK was located only within cells. It was not possible to determine whether MNK was co-localized with either of the milk proteins immunohistochemically because of the intense fluorescence throughout the cytoplasm in sections labeled with the antibodies to β-casein and α-lactalbumin.
Discussion
We have investigated the expression and subcellular localization of MNK in sections from nonlactating and lactating human breast. This is the first report describing the intracellular localization of the MNK protein in any tissues derived from humans. Previous studies have used cultured cells for investigations of the expression and localization of MNK (Yamaguchi et al. 1996; Ackland et al. 1997). The Western blot analysis of breast tissue confirmed the immunohistochemical data, showing that the MNK protein was expressed in the human breast and that its expression was significantly increased in tissue from the lactating breast compared with the nonlactating breast. The size of the protein in both lactating and nonlactating tissue (180 kD) was the same as that obtained from extracts of a human mammary carcinoma cell line, PMC42, found here and in a previous study (Ackland et al. 1997).
In nonlactating human breast tissue, MNK was localized primarily in the perinuclear region of the luminal epithelial cells and its distribution was similar to that of TGN38. This is consistent with previous immunocytochemical studies obtained from a variety of cells, including cultured human breast carcinoma cells (Ackland et al. 1997), CHO cells (Petris et al. 1996), human HeLa cells (Yamaguchi et al. 1996), human fibroblasts and CHO cells (Dierick et al. 1997), in which MNK was localized to the trans-Golgi network.
A major finding of our study was that the distribution of MNK protein was altered in the lactating breast tissue compared to the nonlactating breast tissue. In the lactating breast epithelial cells, the perinuclear fluorescence was still detected but there was an additional diffuse, granular fluorescence that extended into the cytoplasm of the cell. Overall, the fluorescence in the lactating tissue was considerably more intense than in the nonlactating tissue. The relocalization of MNK to the cytoplasm seen here in lactating epithelial cells is analagous to that seen in cultured CHO cells in the presence of high concentrations of copper acetate and which is associated with cellular copper efflux (Petris et al. 1996). On the basis of the CHO studies, it was proposed that the MNK protein relocalizes to the plasma membrane and facilitates the efflux of copper from the cell (Petris et al. 1996). Because of difficulties in accurately determining the in-tracellular location of the proteins in solid tissue compared with cultured cells, we could not conclusively demonstrate that the MNK protein relocalizes to the plasma membrane, although this cannot be ruled out. It is also possible that the pathway followed by MNK in luminal epithelial cells is similar to that found in the liver for the Wilson disease protein, which does not locate to the plasma membrane in the presence of elevated copper but which redistributes to a cytoplasmic vesicular compartment near the hepatocyte canalicular membrane (Schaefer et al. 1999). Overall, the results of our analysis in lactating and nonlactating breast suggest that MNK is redistributing in lactating cells, and this might be consistent with an involvement of this protein in secretion of copper from epithelial cells during lactation. If this is the case, it is the first demonstration of MNK relocalization in a physiological context.
(
We detected MNK in luminal ductal and alveolar cells, both of which were positive for the major milk proteins β-casein and α-lactalbumin. Nonsecretory myoepithelial cells, which were identified by the presence of α-smooth muscle actin, were negative for MNK and for milk proteins. The presence of MNK in luminal secretory cells but not in myoepithelial cells is also evidence to supporting a role for MNK in secretory cells.
Studies in rats have shown that during lactation there is increased uptake of copper into the mammary gland. Injected 67 Cu was taken up rapidly by the mammary gland such that the rate of 67 Cu uptake by the mammary gland over 4 hr exceeded that of the liver, with two thirds of the total injected dose found in the mammary gland (Donley et al. unpublished observations). It is possible that the relocalization of MNK seen in lactating human breast cells may be the result of an increase in copper flux through luminal secretory cells of the breast, analagous to the effect of copper in cultured cells. Therefore, trafficking of MNK may be controlled by the copper levels in the mammary gland, which in turn may regulate secretion of copper into milk during lactation.
In lactation, changes in the breast include epithelial cell proliferation and differentiation, and the synthesis and secretion of milk. These are mediated by the hormones estrogen, progesterone, and prolactin. These lactational hormones may also regulate the movement of MNK from the trans-Golgi network. In previous work we investigated the effect of hormones on expression and localization of MNK using the cultured mammary carcinoma line PMC42. MNK was located in a cytoplasmic region in addition to the perinuclear distribution. In these cells, exposure to lactational hormones did not alter the distribution of MNK (Ackland et al. 1997). Our data, however, showed that the PMC42 cells were not polarized, and this may account for the lack of effect of lactational hormones because nonpolarized cells may not be responsive to lactational hormones. Further studies are required to establish the relationship between hormones and the localization of MNK.
The MNK protein may reside in the membrane of vesicles containing milk proteins. It was not possible, however, to determine this with immunofluorescence because the labeling with β-casein and α-lactalbumin was much stronger and more extensive than that of MNK. It was interesting to note that these milk proteins were not found predominantly on the apical side of the cells, as might be expected if they were being secreted into the lumen during lactation.
If the MNK protein functions to export copper into milk, the mechanism for this may be similar to that of other ions that are secreted during lactation. A vesicular pathway from the Golgi is taken by the major milk proteins (Linzell and Peaker 1971) and also by calcium (Baumrucker and Keenan 1975; Neville and Peaker 1979). Free calcium diffuses into the mammary cell via the basolateral membrane and is pumped by an ATPase into the Golgi lumina, and is then transported in vesicles to the plasma membrane, where the vesicle contents are released into the ductal and alveolar lumina. The pathways taken by sodium and potassium ions, and also by lipids, do not involve the Golgi (Linzell and Peaker 1971; Patton and Keenan 1975). It therefore is unlikely that MNK is associated with this route. Our results suggest that the proposed vesicular pathway of copper from the Golgi into milk is similar to that of calcium.
The toxic milk mouse has a defect that results in the production of copper-deficient milk by lactating dams (Rouch 1983). The gene underlying this disease is homologous to the Wilson disease gene, which is required for copper transport from the liver into the bile and for incorporation of copper into ceruloplasmin (Theophilos et al. 1996). It is not yet clear whether the toxic milk defect directly affects the mammary gland and impairs the transfer of copper into milk or whether copper-deficient milk is a consequence of low levels of plasma copper that lead to reduced mammary copper uptake from the blood.
In conclusion, our results are consistent with the possibility that the Menkes protein transports copper to breast milk via a vesicular pathway. Lactation is associated with an increase in expression of MNK. Furthermore, the changes induced in the lactating breast cause redistribution of the Menkes protein resembling that observed in cultured cells exposed to copper, suggesting that this relocalization is a physiological process.
Footnotes
Acknowledgements
We are grateful to Mr John Collins and Dr Robert Brown of Melbourne Pathology for providing tissue samples.