
Abstract
Among the four existing isoforms of alkaline phosphatase (AP), the present study is devoted to tissue-nonspecific alkaline phosphatase (TNAP) in mineralized dental tissues. Northern blot analysis and measurements of phosphohydrolase activity on micro-dissected epithelium and ectomesenchyme, in situ hybridization, and immunolabeling on incisors confirmed that the AP active in rodent teeth is TNAP. Whereas the developmental pattern of TNAP mRNA and protein and the previously described activity were similar in supra-ameloblastic and mesenchymal cells, they differed in enamel-secreting cells, the ameloblasts. As previously shown for other proteins involved in calcium and phosphate handling in ameloblasts, a biphasic pattern of steady-state TNAP mRNA levels was associated with additional variations in ameloblast TNAP protein levels during the cyclic modulation process. Although the association of TNAP upregulation and the initial phase of bio-mineralization appeared to be a basic feature of all mineralized tissues, ameloblasts (and to a lesser extent, odontoblasts) showed a second selectively prominent upregulation of TNAP mRNA/protein/activity during terminal growth of large enamel crystals only, i.e., the maturation stage. This differential expression/activity for TNAP in teeth vs bone may explain the striking dental phenotype vs bone reported in hypophosphatasia, a hereditary disorder related to TNAP mutation.
Keywords
Investigations of TNAP in the skeleton have been mostly performed in bone and cartilage (Weinreb et al. 1990; Rodan and Noda 1991; Anderson and Morris 1993; Aubin et al. 1993; Hsu et al. 1993) in relation to its potential role in calcium and phosphate handling during biomineralization. Many histochemical studies have been devoted to teeth from the early 1950s until the present time (see review by Kurahashi and Yoshiki 1972; Deporter and Ten Cate 1976; Linde and Granström 1980; Orams and Snibson 1982; Gomez and Boyde 1994; Linde and Lundgren 1995; Takano 1995; Wöltgens et al. 1995). More recent investigations in the bovine enamel organ by cDNA isolation (Yuan et al. 1995), in human pulp (Goseki-Sone et al. 1999), and in the early stages of murine tooth formation by immunolabeling (Hoshi et al. 1997) and by in situ hybridization (Davideau et al. 1996) suggest that the AP previously detected by histoenzymology in teeth was TNAP, as clearly established in bone (Noda et al. 1987; Thiede et al. 1988). These studies do not provide an overview of the temporospatial patterns of expression of mRNA and protein from early stages until complete enamel maturation. In this study, several methods of investigation were combined in the postnatal rat incisor. Northern blot analysis and measurements of enzymatic activity on microdissected epithelial and mesenchymal cells, in situ hybridization, and immunoperoxidase labeling were jointly performed. Our data show that TNAP mRNA and protein are effectively expressed in odontogenic cells and fluctuate differentially, supporting a prominent role for this enzyme during two critical phases, initiation and completion of dental biomineralization.
Materials and Methods
Ten-day-old (n = 12), 30-day-old (n = 18), and 56-day-old (n = 30) male Sprague-Dawley rats (Charles River; St Aubin les Elbeuf, France) were used for this study.
Tissue Preparation for RNA Investigation
The rats were decapitated after carbon dioxide asphyxiation, the mandibles were rapidly collected, and the alveolar bone was removed under a stereoscopic microscope. The incisor was extracted and the distinct epithelial and mesenchymal odontogenic cells were microdissected. The apical portion containing the odontogenic organ and the early mineralization zone of enamel and dentin was removed to ensure purity of epithelial and mesenchymal samples. Lateral cuts were made along the cemento-enamel junction and the enamel organ was scraped off the labial part of the incisor. Finally, the dental mesenchyme was collected. Dental samples and kidneys were maintained under liquid nitrogen. Epithelial and mesenchymal dissection procedures were validated by: (a) identification of enamel protein mRNA, i.e., encoding amelogenins in epithelial cells and osteocalcin mRNA in mesenchymal cells (not shown) and (b) comparative measurements of purified proteins and alkaline phosphatase activity in right and left incisors of the same animals.
Northern Blotting
Total RNA isolation was performed on 100–200 mg of microdissected tissue from 56-day-old rats with an RNA extraction kit (Euromedex; Souffelweyersheim, France). Total RNA was electrophoretically fractionated on a 1% agarose formaldehyde gel and transferred onto nylon membranes. Filters were prehybridized, hybridized with 32 P-labeled rat TNAP cDNA (G.A. Rodan; Merck Research Laboratories, West Point, PA), washed, and autoradiographed, as previously described (Berdal et al. 1993). GAPDH mRNA was also investigated to verify the amounts of mRNA in each lane.
Measurement of AP Activity
The samples from 56-day-old rats were washed twice in 0.1 M PBS (Sigma; La Verpillière, France) and rinsed in 100 mM sodium carbonate-bicarbonate solution at pH 10.2. They were homogenized at 6000 rpm with a polytron homogenizer (Ultraturax; Ika, Germany) in a solution containing 0.1% Nonidet P 40 (Sigma) 1 mM MgCl2, and sonicated at 4C. Lysates were removed by centrifugation at 3000 × g for 5 min. AP activity was determined in the supernatant. The enzymatic activity was expressed as nmol of p-nitrophenol (PNP) released per minute per mg of protein at 37C. AP phosphohydrolase activity was assessed by measuring PNP release from p-nitrophenolphosphate (PNPP) by absorbance spectrophotometry at 410 nm and compared with a PNP standard solution (Sigma). The reaction was carried out in 500 μl of buffer solution (1.5 M 2-amino-2-methyl-1 propanol, pH 10.3, with 15 mM PNPP). After 15 min at 37C with 50 μl of the tissue lysate supernatant, the reaction was stopped with 2 ml of 0.1 M NaOH. Protein concentrations were determined by Lowry's modified method (Lowry et al. 1951; Anagnostou et al. 1996) using bovine serum albumin as standard. Km values were determined by Lineweaver-Burk representations based on the use of various dilutions of the enzyme, various substrate concentrations (1, 2, 5, 10, and 15 mM), five reaction times (from 2 to 20 min), and five separate measurements.
Preparation of Incisor Samples for In Situ Hybridization and Immunolabeling
After barbital anesthesia, rats (10-, 30-, and 56-day-old) received an intracardiac infusion of 4% paraformaldehyde-15% sucrose in PBS (Sigma), pH 7.4, for 15 min. Mandibles were dissected out, fixed by immersion in the same fixative for 1 hr at 4C, and rinsed overnight in 15% sucrose-PBS at 4C. The mandibles were cut either without decalcification (left incisors) or after decalcification (right incisors). These latter were rinsed for 4 hr in PBS at 4C and decalcified for 4 weeks at 4C in PBS with 4.13% disodium ethylenediaminetetraacetic acid (Sigma) and 0.2% paraformaldehyde (Sigma) pH 7.4, dehydrated, and paraffin-embedded. Ten -μm sections of dissected left incisors were made with a cryostat at −25C (MGW Lauda Leitz; Rockleigh, NJ). Sections were deposited onto 50 mg/ml poly-L-lysine (Sigma)-coated slides and were then dehydrated in a graded ethanol series and stored at 4C. The other sections (decalcified right incisors) were made with a paraffin microtome (Leica; Rueil Malmaison, France) and were deposited on silanized slides, deparaffinized, and rehydrated before use.
In Situ Hybridization
A 2.2-kb fragment of rat TNAP subcloned into Bluescript-SK+ plasmid (gift from M. Vogel and G.A Rodan; Merck Research Laboratories) was linearized with BamHI or PvuII endonucleases (Promega; Madison, WI). [35 S]-UTP-labeled single-stranded antisense and sense probes were synthesized in vitro using T7 and T3 polymerases, respectively (Promega). In situ hybridization was performed as previously described (Hotton et al. 1995). Briefly, cryostat sections were pretreated with proteinase K (Sigma), hybridized with 20 μl of labeled probes containing 60,000 cpm/μl radioactivity in a moist chamber overnight at 50C, and washed under high-stringency conditions. The slides were dipped into NTB2 autoradiographic emulsion (Kodak; Paris, France) and exposed for 4 weeks at 4C. After developing the film, sections were stained with Harris hematoxylin (Sigma), dehydrated, and mounted under a coverslip. Sections were examined and photographed with a Zeiss photomicroscope using brightand darkfield illumination.
Immunocytochemical Procedures
Monoclonal murine primary antibodies specific for rat TNAP (M. Vogel and G. Rodan) were used. Paraffin sections were treated with 0.3% hydrogen peroxide in 0.1 M Tris-HCl, pH 7.6, for 10 min to inhibit endogenous peroxidase activity. After rinsing in Tris-HCl solution, the sections were incubated overnight in Tris-HCl containing 1:30 nonimmune goat serum (Nordic; Tilburg, The Netherlands) to block nonspecific binding sites and were then incubated with serial dilutions of monoclonal rat TNAP antibodies (from 1:750 to 1:4000) for 2 hr at room temperature, rinsed in 1% Tris-HCl-bovine serum albumin (BSA), and incubated with biotinylated polyclonal rabbit anti-mouse secondary antibodies at a 1:100 dilution for 1 hr. After incubation in 1:300 diluted extravidin-peroxidase (Sigma) for 30 min, the immunoreactive sites were visualized by 3–3′-tetrachloride diaminobenzidine oxidation (Sigma), 5 mg/10 ml in 0.1 M Tris-HCl, pH 7.6, with 0.03% hydrogen peroxide. Sections were rinsed in Tris-HCl, dehydrated, and mounted in Depex (Gurr; OSI, France). Sections were lightly counterstained with Harris hematoxylin solution (Sigma). Irrelevant murine immunoglobulins (1:750-1:4000) were used as negative controls.
Results
Biochemical Investigations of Rat TNAP
The measurement of phosphohydrolase-specific activity of AP was realized in 56-day-old rat samples. This activity was consistently and significantly higher (p= 0.006) in microdissected mesenchyme (15,184 ± 911 nmol PNP/min at 37C/mg proteins) than in epithelium (7145 ± 214 nmol PNP/min at 37C/mg proteins). The reproductibility of microdissection procedures was established by the low variability between measurements (< 3%) on paired left and right incisors of the same rat (Table 1). The Km values for epithelial and mesenchymal samples were identical (2.1 mM).
Northern blot analysis was serially performed with TNAP and GAPDH probes using mRNA from two different portions of the tooth containing epithelial (EO) and mesenchymal (M) differentiated cells. The kidney (K) was used as the reference organ for rat TNAP. The transcripts (2.5 kb) expressed in dental epithelium and mesenchyme corresponded to that of the TNAP enzyme observed in the kidney (Figure 1).
Distribution of TNAP mRNA and Protein in the Mandible
The forming alveolar bone (Figure 2) and adjoining cells of the follicular sac surrounding the incisor contained immunoreactive alkaline phosphatase. Progenitor and differentiated bone cells of the mandible (Figure 3) also consistently showed AP immunostaining, with an apparent decrease from early stages of osteoblasts/recently embedded osteocytes to the stages of older osteocytes. TNAP protein was present in both the cell membrane and cytoplasmic compartments of bone cells. Immunocontrols showed no labeling (Figure 4).
Developmental Pattern of TNAP Protein and mRNA During Amelogenesis
The expression of TNAP mRNA and protein was analyzed in the incisor enamel organ (Figures 5–9 and 12). Similar data concerning the developmental patterns of AP mRNA and protein at the various ameloblast stages were obtained in 10-, 20-, and 56-day-old rat incisors. They are illustrated in the plates obtained from 56-day-old rats. The developmental pattern was followed from the presecretion stage (Figure 5) to the secretion (Figure 6) and maturation (Figures 7–9 and 12) stages, ordered on the longitudinal axis of the samples. The apical loop that contains undifferentiated epithelial and mesenchymal cells was devoid of TNAP mRNA (not shown) and protein (Figure 2). At the end of the enamel presecretion stage, the TNAP transcripts were detected in presecretory ameloblasts and stratum intermedium cells (Figure 5A and 5B). TNAP mRNA expression persisted in stratum intermedium cells during the secretion stage, whereas TNAP mRNA dramatically decreased in secretory ameloblasts (Figure 6A and 6B). This distribution was observed throughout the secretion stage. Maturation stage ameloblasts showed progressively increasing apparent concentrations of TNAP transcripts from the transition phase (Figure 7A and 7B) to the first ameloblast modulation (not shown). The highest and apparently stable steady-state mRNA levels were then observed throughout the maturation stage (Figure 8A and 8B). The TNAP mRNA and protein distribution patterns were identical, except in the ameloblasts during the maturation stage. Ruffle-ended ameloblasts showed intense labeling during the modulation process, especially adjacent to the enamel matrix inside the ruffled border, in contrast with the smooth-ended ameloblasts, which were almost devoid of staining, at least on the smooth border (Figure 9). TNAP mRNA was detected in supra-ameloblastic cells with a decreasing gradient from the transition stage (Figure 7A and 7B) throughout the successive ameloblast modulation cycles, resulting in almost complete absence of mRNA in supra-ameloblastic cells (Figure 8A and 8B). The same pattern was observed for TNAP protein in supra-ameloblastic cells (Figure 9). Finally, immunostaining was also found in the extracellular enamel matrix bordering the apical pole of the ameloblasts in the maturation stage (Figure 10).
Alkaline phosphatase activity in 56-day-old rat incisors (mean ± SD) a
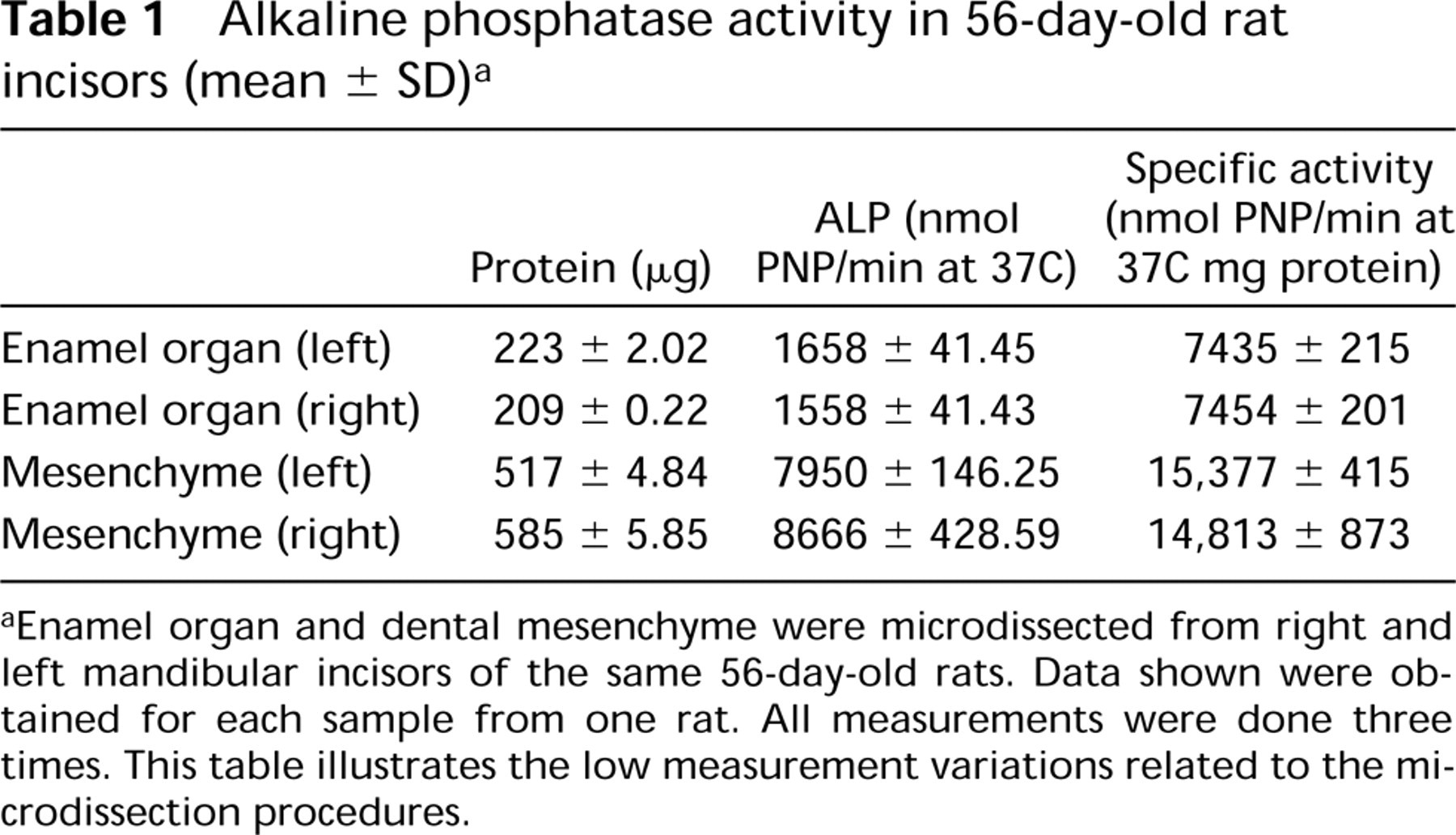
aEnamel organ and dental mesenchyme were microdissected from right and left mandibular incisors of the same 56-day-old rats. Data shown were obtained for each sample from one rat. All measurements were done three times. This table illustrates the low measurement variations related to the microdissection procedures.
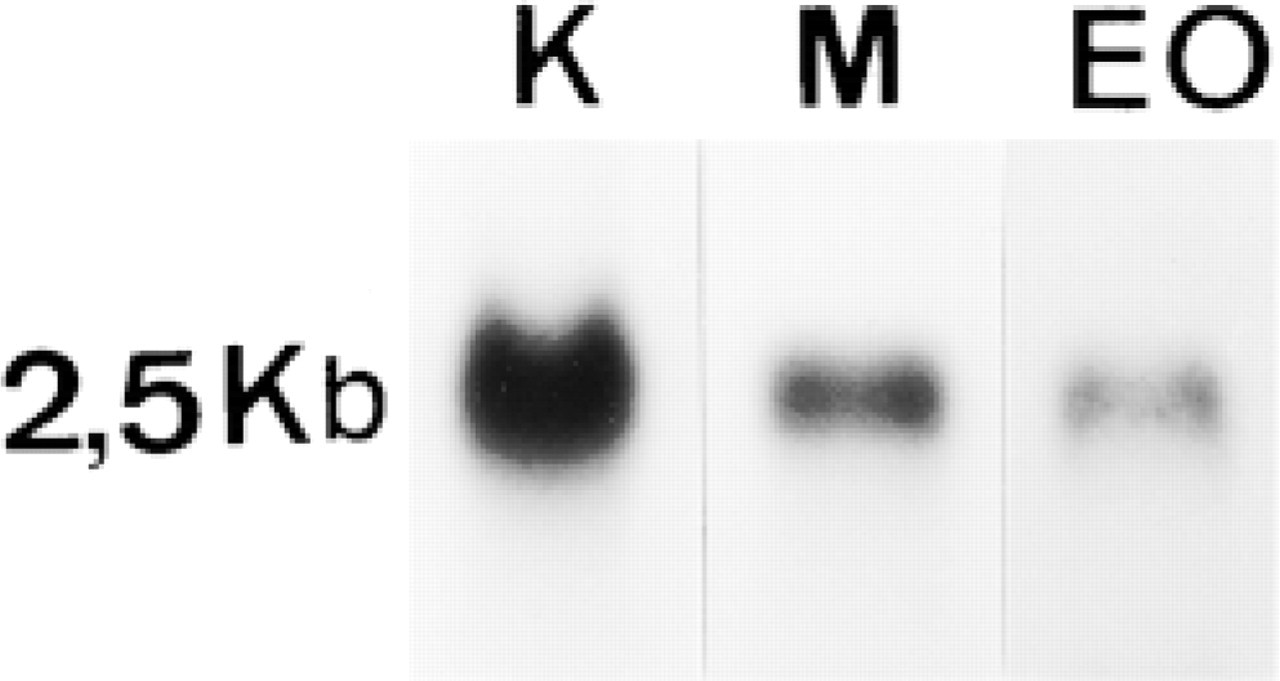
Northern blot analysis of TNAP mRNA (20 μg total mRNA) in kidney (K), microdissected dental mesenchyme (M), and enamel organ (EO). The molecular weight of TNAP mRNA was identical in all studied tissues (2.5 kb).
Expression of TNAP mRNA and Protein in Dentinogenic Cells
Consistent with the data obtained with TNAP antibodies, TNAP mRNA appeared to be present in the odontoblasts and subodontoblastic cells (Figure 5A and 5B), but the relative abundance of TNAP mRNA decreased in odontoblasts with the progressive deposition and biomineralization of mantle dentin followed by orthodentin. TNAP protein immunolabeling was present in the cell membrane of odontoblasts and subodontoblastic cells (Figure 11). Immunostaining was distributed in the cytoplasmic compartment and plasma membrane, from the supranuclear area to the secretory pole of odontoblasts. As observed in osteoblasts and epithelial cells, the nuclei of odontoblasts and subodontoblastic cells were devoid of labeling (Figure 11). The extracellular signal evidenced by serial dilutions of TNAP antibodies in the predentin-dentin border (1:1000 dilution, Figure 12 vs 1:750 dilution, Figure 11) became progressively more obvious, suggesting enrichment of TNAP epitopes in this extracellular zone compared to the intracellular compartments. Such an effect of TNAP antibody dilution was also observed for the extracellular labeling of enamel matrix (Figure 10).
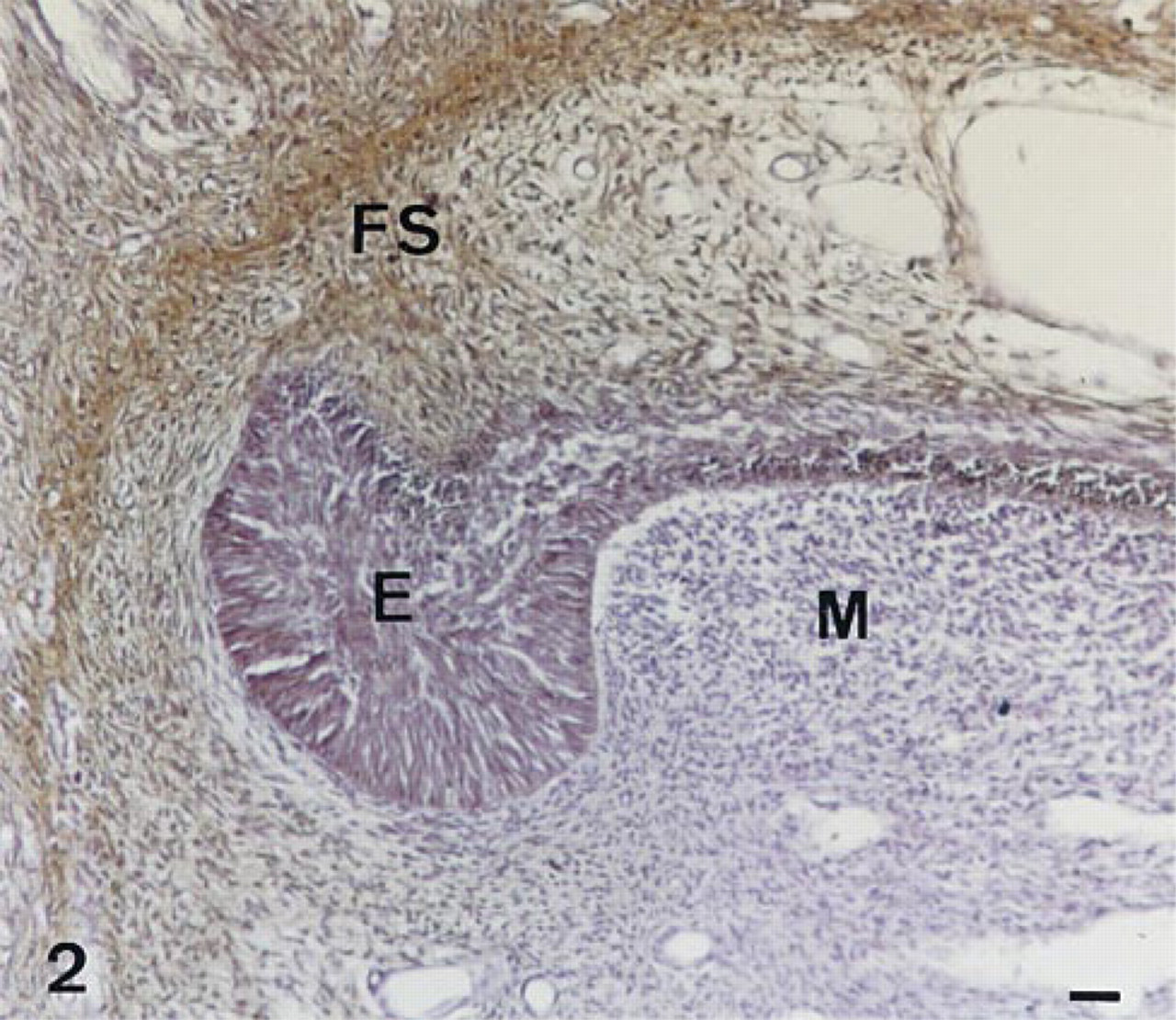
Immunolocalization of AP in the apical loop of the enamel forming part of a 56-day-old decalcified rat incisor. Undifferentiated epithelial (E) and ectomesenchymal (M) cells of the rat incisor do not appear to contain immunoreactive AP. In contrast, the cells of the follicular sac surrounding the forming rat incisor react with the TNAP antibodies (FS). Bar = 12.2 μm.
Discussion
This study was devoted to the comparative investigation of AP protein and mRNA in three different mineralized tissues: the best characterized bone, used as a reference system, as well as enamel and dentin, which have previously been mainly investigated separately by enzymatic methods, as many studies have been devoted to the fine analysis of AP activity by light and electron microscopy of bone and teeth (for reviews see Kurahashi and Yoshiki 1972; Deporter and Ten Cate 1976; Linde and Granström 1980; Orams and Snibson 1982; De Bernard et al. 1986; Takano et al. 1986; Bonucci et al. 1992; Gomez and Boyde 1994; Linde and Lundgren 1995; Wöltgens et al. 1995). TNAP immunolabeling has been previously performed during the first steps of amelogenesis and dentinogenesis (Hoshi et al. 1997), whose data are consistent with our study. The findings of present experiment, exploring the overall distribution pattern of TNAP protein during dentinogenesis and the sequence of presecretion/secretion/maturation stages of amelogenesis, are also consistent with published distributions of AP activity. The respective distributions of AP protein and activity in enamel and dentin were identical, in contrast with cartilage, where the protein detected was either enzymatically inactive or active in the extracellular compartment, depending on the stage of biomineralization (De Bernard et al. 1986). In bone, TNAP protein was not associated with a distinct extracellular structure, as described previously (Pinero et al. 1995). The further results obtained here by comparative enzymology and Northern blot analysis established that the AP expressed in rat teeth is the tissue-nonspecific alkaline phosphatase (TNAP), as previously suggested (Hoshi et al. 1997) and demonstrated in bovine (Yuan et al. 1995) and human species (Goseki-Sone et al. 1999). Quantitative methods also showed that the enzyme activity was related to the steady-state level of mRNAs: higher in mesenchyme than in the enamel organ. Similarly, in primary cell culture systems, the AP activity (Collin et al. 1992) and mRNA steady-state level (Stein et al. 1998) varied in parallel during the stages of osteoblast proliferation, differentiation, and bone mineralization. The observed patterns for mRNA and protein in bone tissue in vivo shown here were consistent with the concept established in vitro that AP is intensely expressed during terminal differentiation of osteoblasts and initial biomineralization and decreases when bone mineralization has been achieved.
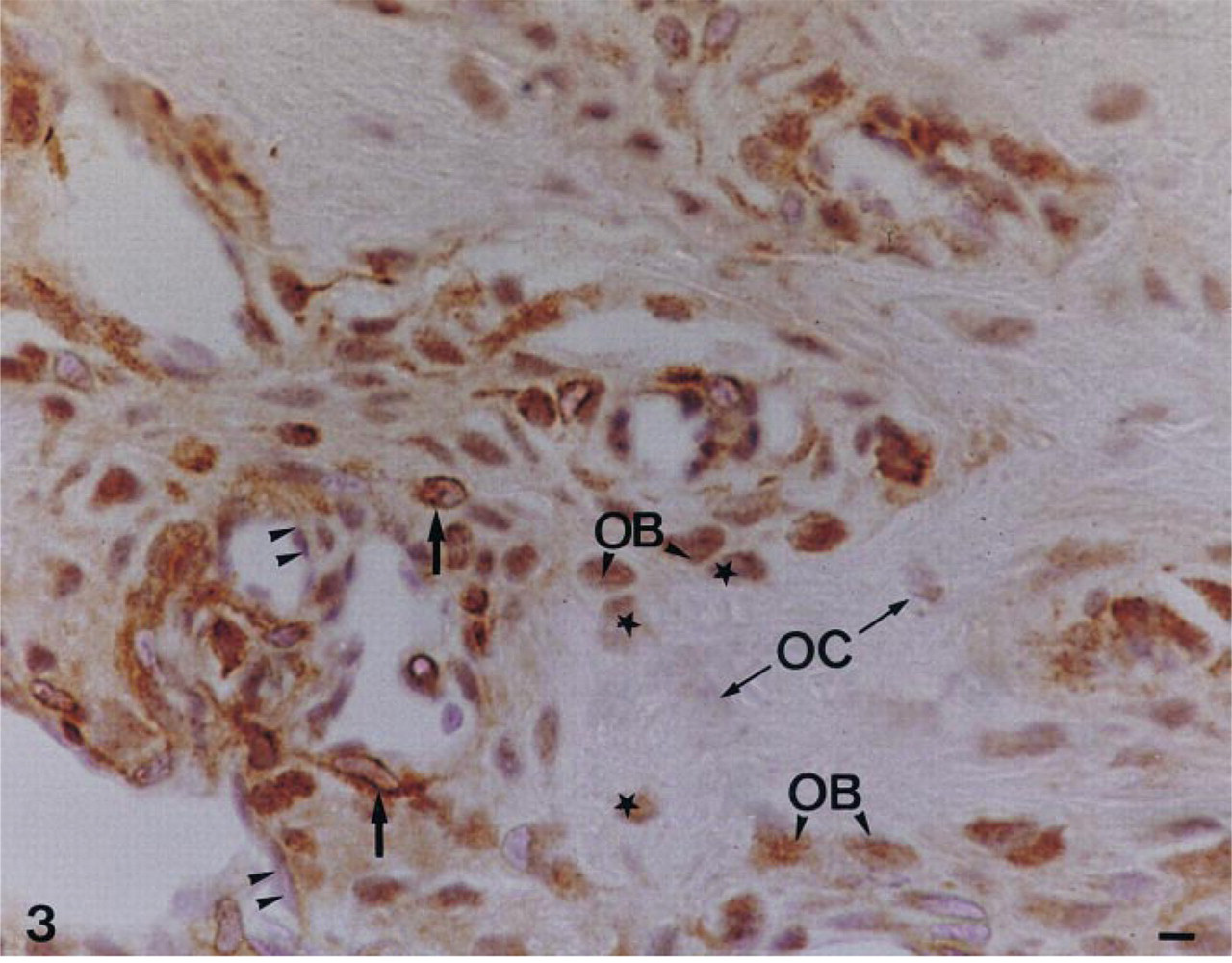
Immunolocalization of TNAP in a decalcified woven bone located underneath the anterior third of the incisor of a 56-day-old rat mandible. Forming endosteum contains mixed immunonegative (double arrowheads) and immunopositive cells (black arrow). Alkaline phosphatase is localized in the differentiating osteoblasts (OB). Young osteocytes (black star), recently embedded in the woven bone, are immunoreactive, whereas older osteocytes in the central part of bone matrix (OC) do not contain a significant amount of immunoreactive alkaline phosphatase. Bar = 1.2 μm.
Control with irrelevant antibodies in the forming mandible on a serial section of the 56-day-old decalcified rat incisor. Bar = 1.2 μm.
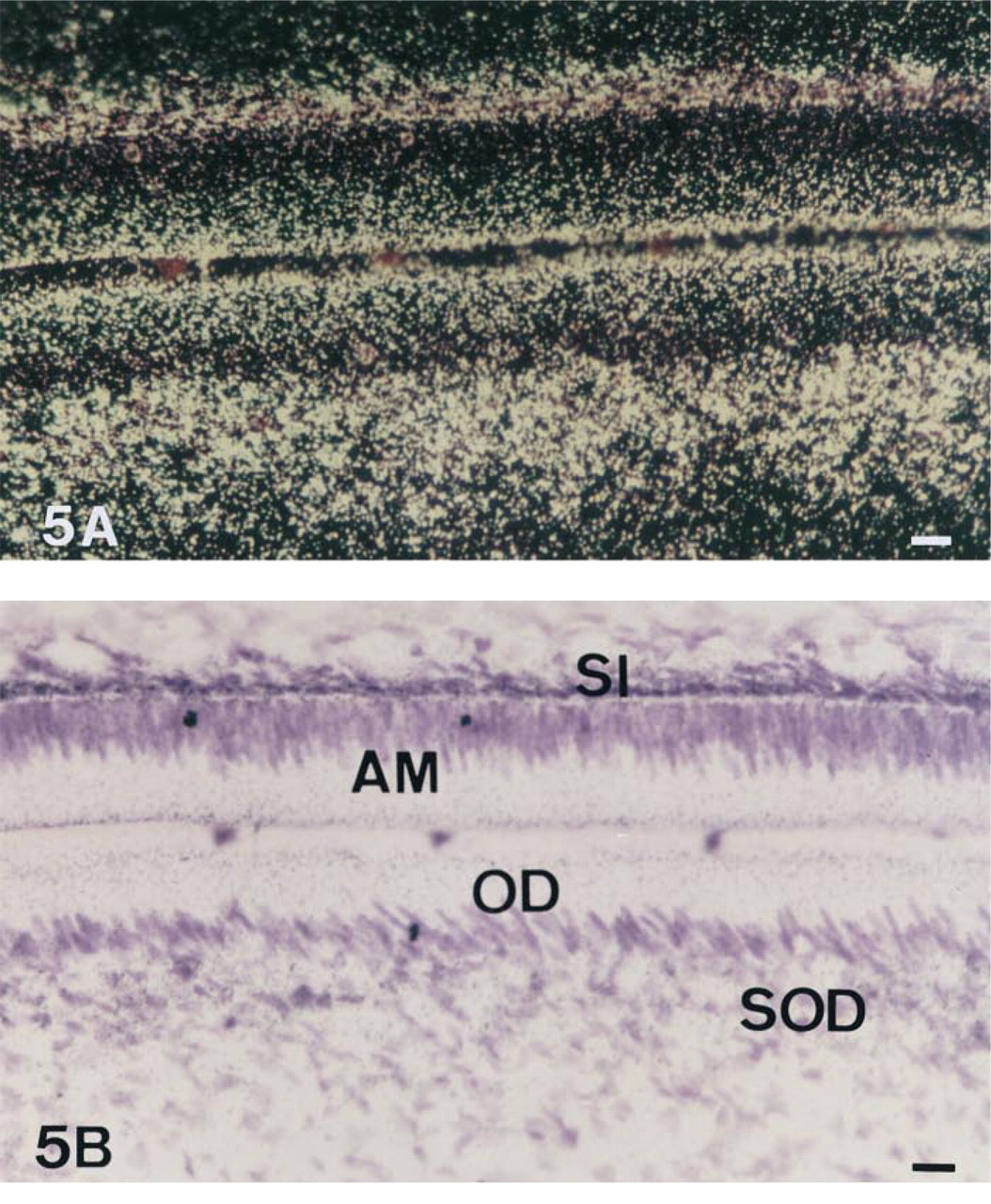
In situ hybridization with AP antisense riboprobe of a 56-day-old undecalcified rat incisor, dark- (A) and brightfield (B) views. In the region of the presecretion stage of enamel formation, ameloblasts (AM) show many transcripts for AP but less than supra-ameloblastic cells of the stratum intermedium (SI). In the region of the formation of the mantle dentin, AP transcripts are present at high levels in odontoblasts (OD) and subodontoblastic cells (SOD). Bar = 6.1 μm.
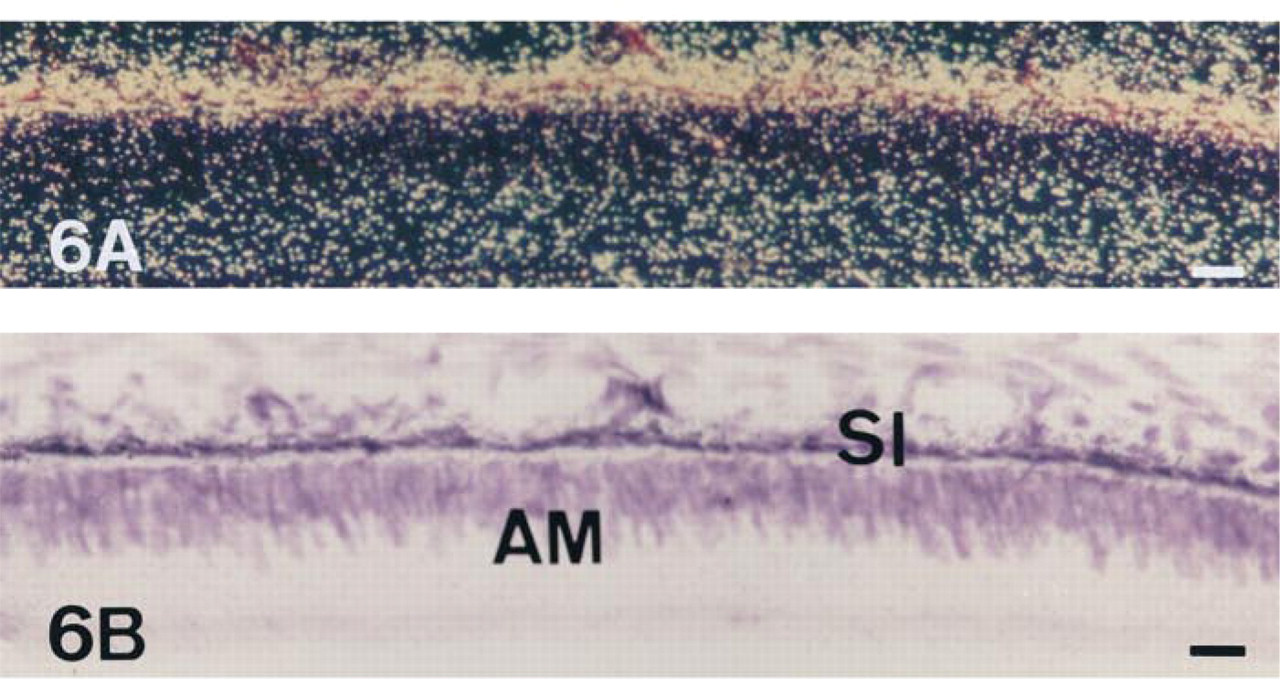
In situ hybridization with AP antisense riboprobe of a 56-day-old undecalcified rat incisor in the region of the secretion stage of enamel formation, dark- (A) and brightfield (B) views. Ameloblasts (AM) are almost devoid of transcripts, in contrast to a specific subpopulation of supra-ameloblastic cells, the stratum intermedium (SI). Bar = 6.1 μm.
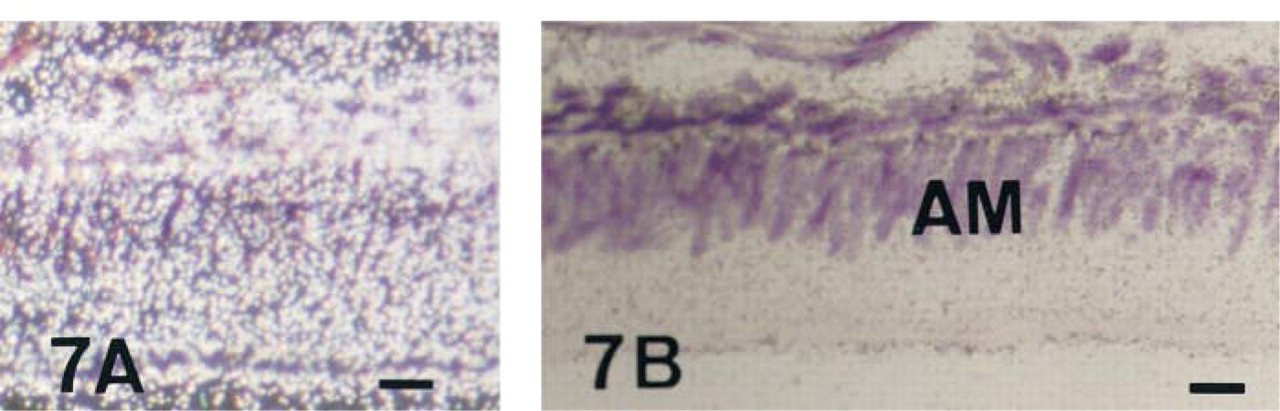
In situ hybridization with AP antisense riboprobe of a 56-day-old undecalcified rat incisor in the region of the transition stage of enamel formation, dark- (A) and brightfield (B) views. Labeling in ameloblasts (AM) increases during the transition phase and stays homogeneous throughout the maturation process. Supra-ameloblastic cells still show a significant signal. Bar = 3 μm.
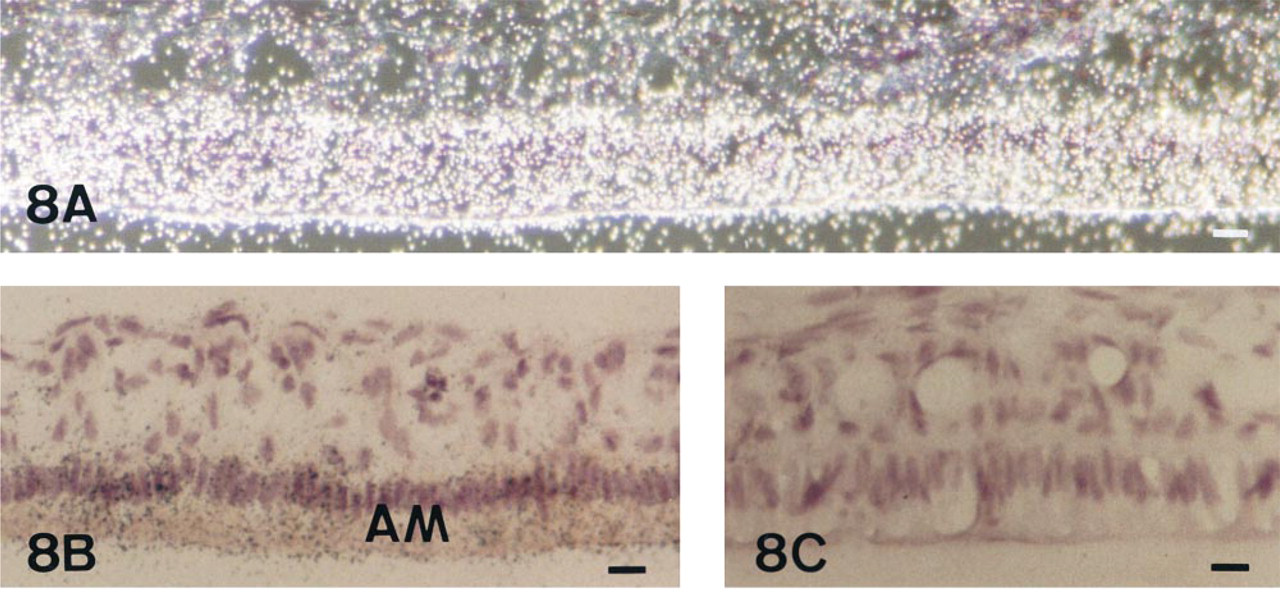
In situ hybridization with AP antisense riboprobe of a 56-day-old undecalcified rat incisor in the region of the late maturation stage of enamel formation, dark- (A) and brightfield (B,C) views. Labeling inside ameloblasts (AM) is still homogeneous and significant throughout maturation until the latest stages shown here, but no positive cells are detected in the supra-ameloblastic cells at these stages. In situ hybridization with AP sense riboprobe on the serial section is devoid of signal (C). Bars = 3 μm.
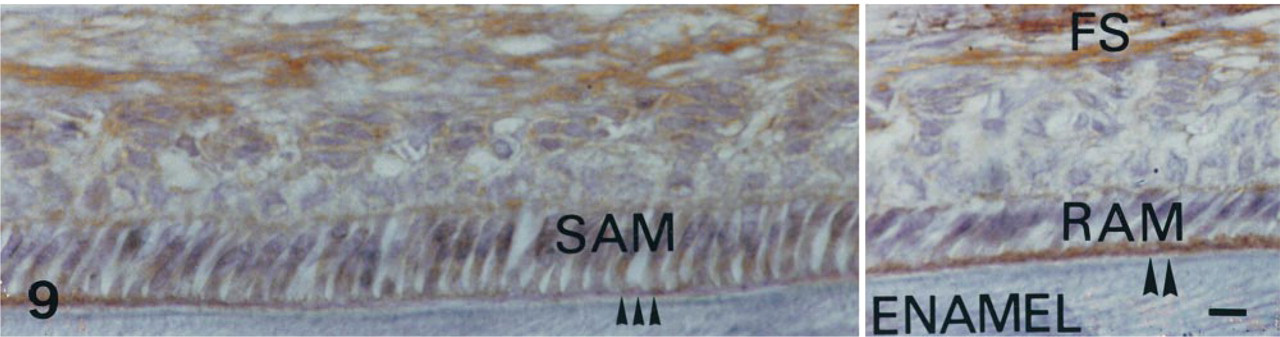
AP immunolabeling in a 56-day-old decalcified rat incisor in the region of the late maturation stage of enamel formation. The follicular sac (FS) is heavily labeled. The maturation stage ameloblasts show various morphologies. The smooth-ended ameloblasts (SAM) are characterized by a smooth border and wide intercellular spaces (triple arrows). The ruffle-ended ameloblasts (RAM) show a wide apical border which corresponds to the ruffled border adjacent to the enamel matrix (double arrowheads). The same TNAP pattern is observed throughout the modulation cycles. Immunostaining is shown during the third modulation cycle. It appears strikingly different, depending on ameloblast morphology. Very significant labeling is mainly associated with the ruffled border in RAM. The labeling is weak in SAM. Bar = 3 μm.
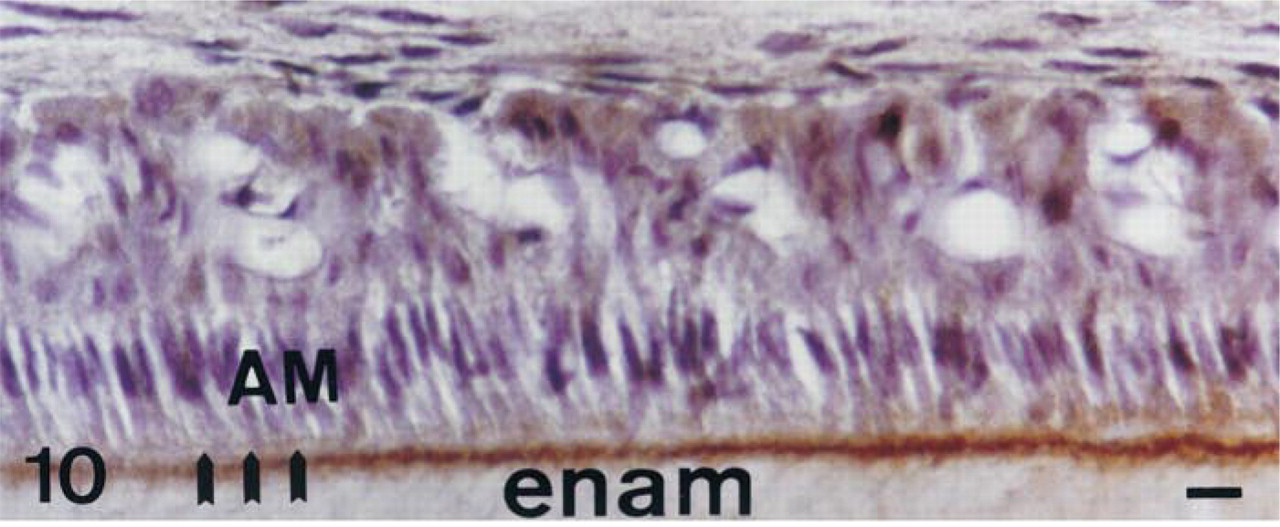
AP immunolabeling in a 56-day-old decalcified rat incisor in the region of the maturation stage of enamel formation; 1:1000 dilution of primary antibodies. During the first modulation, at this dilution the extracellular compartment contains an immunolabeled border facing the ruffle-ended ameloblasts. Triple arrows, ruffled borders; AM, ameloblasts; enam, enamel. Bar = 3 μm.
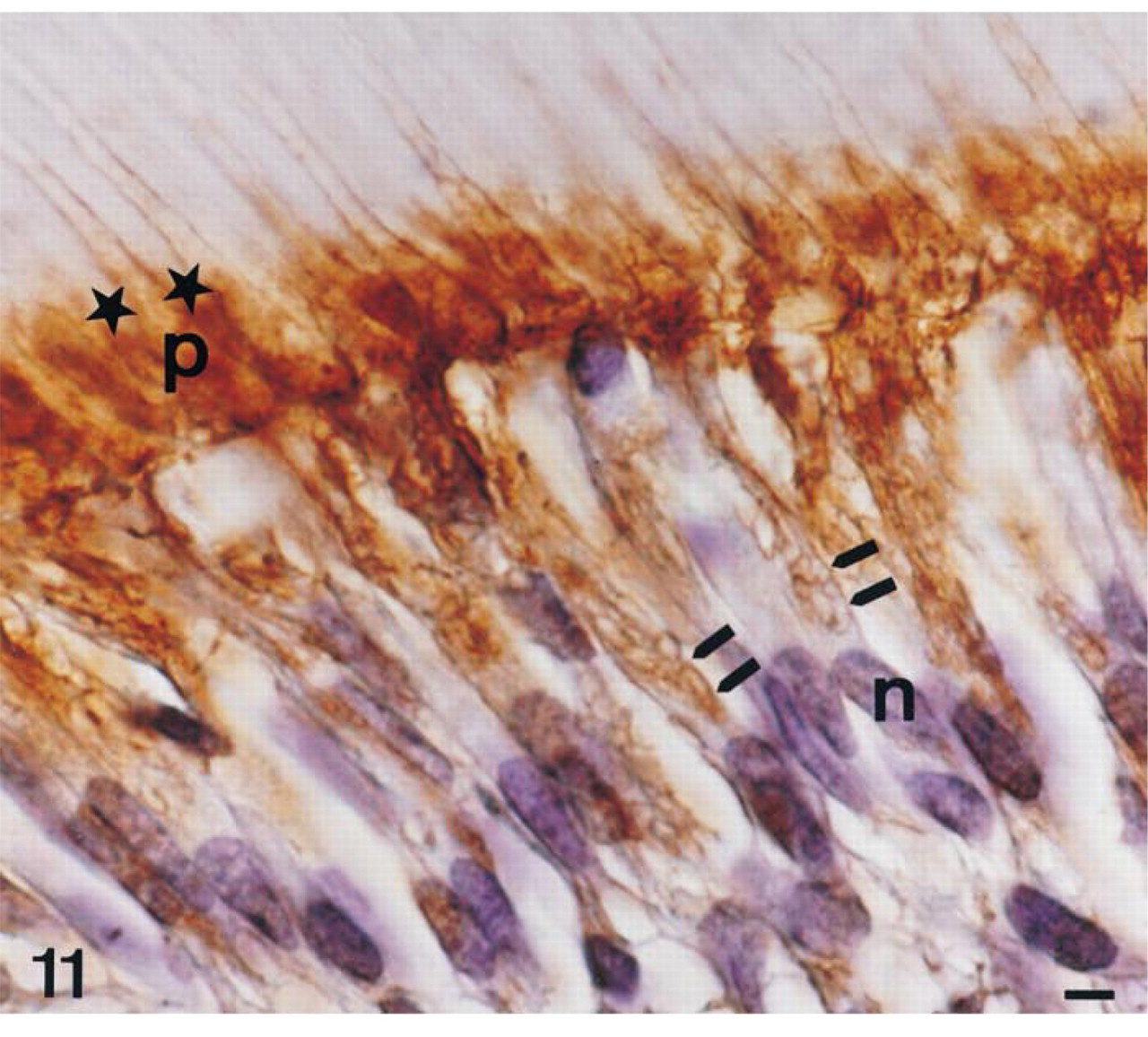
AP immunolabeling in a 56-day-old decalcified rat incisor in the region of mineralization of intercanalicular dentin in the area corresponding to enamel maturation stage; 1:750 dilution of primary antibodies. Immunolabeling is detected in the cytoplasm and the plasma membrane (double arrows) of odontoblasts (OD) and subodontoblastic cells (SOD). In the extracellular compartment, the predentin-dentin junction is also immunolabeled (star). Surrounding cellular process, p; noyau, n. Bar = 1.2 μm.
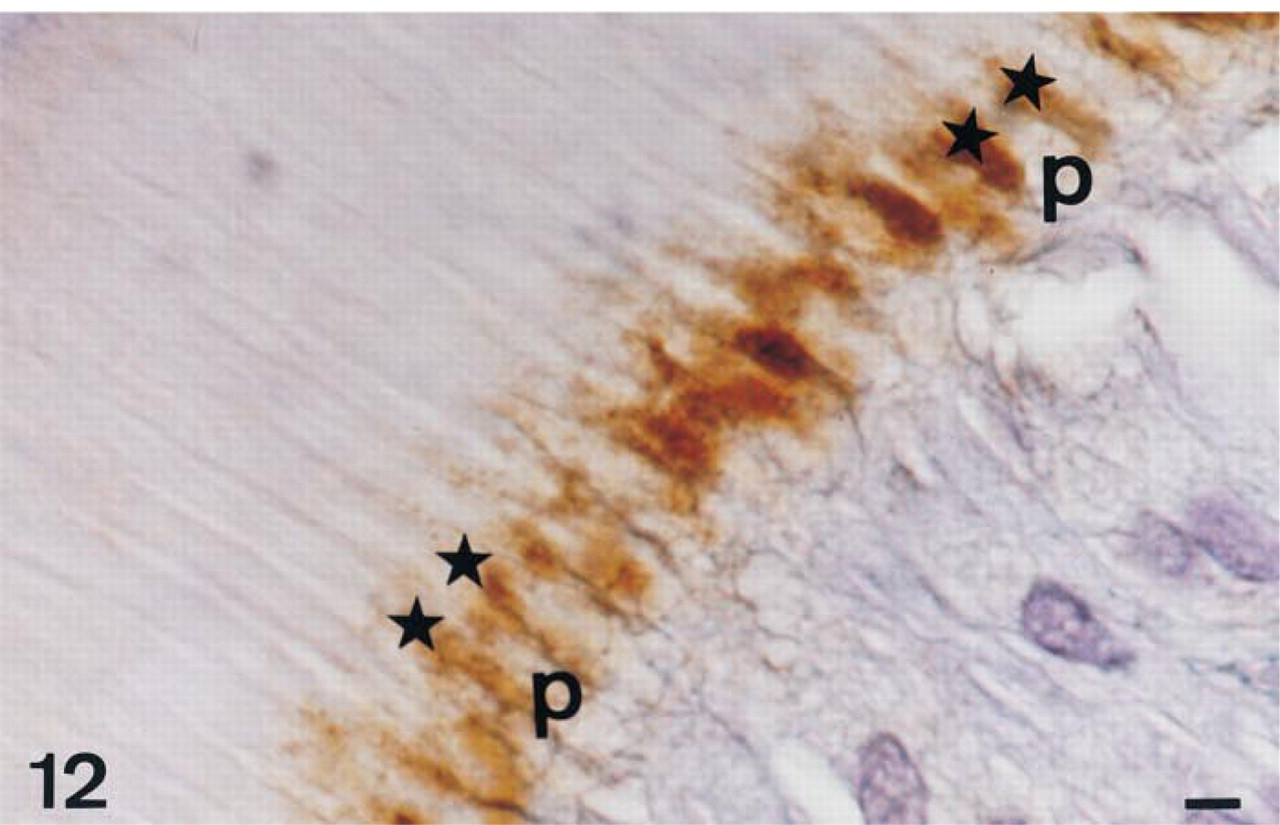
AP immunolabeling in a 56-day-old decalcified rat incisor in the region of mineralization of intercanalicular dentin on a serial section of that shown on Figure 11; 1:1000 dilution of primary antibodies. Staining at the predentin-dentin border (star) was the last to be visualized on serial dilutions. At this dilution, the odontoblast processes are less labeled inside the cytoplasm. This appears clearly when the processes are longitudinally sectioned, surrounded by extracellular TNAP labeling. Odontoblast process, p. Bar = 1.2 μm.
This finding was particularly marked in young rather than old osteocytes. In contrast, during the processes of enamel and dentin formation, initiation of biomineralization was associated not only with upregulation of alkaline phosphatase mRNA/protein/activity in the cells in contact with extracellular matrix (main cells: ameloblasts and odontoblasts, respectively) but also with the subsequent steps of biomineralization. This dental specificity, a biphasic pattern in the main cells, could partly explain the striking phenotype of incisors in TNAP null mutants compared to the bone phenotype (Whyte et al. 1995).
The developmental pattern of AP mRNA, protein (as shown here), and enzymatic activity (see review by Kurahashi and Yoshiki 1972; Deporter and Ten Cate 1976; Linde and Granström 1980; Orams and Snibson 1982; Gomez and Boyde 1994; Linde and Lundgren 1995; Takano 1995; Wöltgens et al. 1995), although identical in ectomesenchymal and supra-ameloblastic cells, differed significantly in the main cells of the enamel organ, ameloblasts. AP mRNA expression was characterized by a biphasic pattern in which two optimal phases were detected, one at the presecretion stage, as previously shown for enzymatic activity (Takano et al. 1986), and the other at the maturation stage, when AP activity reached its highest levels (Gomez and Boyde 1994). An additional modulation of AP protein (this study)/activity (Gomez and Boyde 1994) levels was demonstrated in maturation stage ameloblasts. These data may correspond to selective functions of ameloblasts throughout the successive steps of amelogenesis (Nanci and Smith 1992). Amelogenesis is a complex process, in which the matrix is elaborated and mineralized during two distinct stages. During the secretion stage, enamel matrix is synthesized and exported, and biomineralization is initiated (Smith et Nanci 1995). When the full thickness of extracellular matrix has been achieved, the maturation stage involves selective proteolysis of matrix proteins and completion of hydroxyapatite crystal growth (Robinson et al. 1995). Several gene/protein expression patterns have been described throughout enamel formation, concerning matrix proteins, such as amelogenins, tuftelin (for review see Zeichner-David et al. 1995), amelin/ameloblastin/sheathlin (Cerny et al. 1996; Krebsbach et al. 1996), enamelin (Hu et al. 1997), and matrix proteases (Bègue-Kirn et al. 1998). More recently, several reports (Bègue-Kirn et al. 1998; Fong et al. 1998; Ritchie et al. 1998) have indicated that enamel proteins are expressed in early odontoblasts, while dentin proteins and their corresponding mRNAs are detected in a mirror configuration in presecretory ameloblasts. Interestingly, this lag time corresponds to the period when AP is jointly and transiently co-expressed in ameloblasts and odontoblasts, as clearly demonstrated here by in situ hy bridization. It can therefore be hypothesized that enamel and dentin proteins are jointly controlled by signaling pathways common to epithelial and ectomesenchymal cells during this critical period, in which dentin and enamel matrix are shared, not yet separated by the formed mineral. Interestingly, this period corresponds to the initiation of enamel/dentin biomineralization which might be interdependent and mediated by mechanisms unrelated to the subsequent stages of separate enamel (Robinson et al. 1995; Nanci and Smith 1992) and dentin formation. For example, it has been established that matrix vesicles are temporarily present in mantle dentin (Linde and Goldberg 1993). Interestingly, concerning the role of TNAP in cartilage matrix vesicles (De Bernard et al. 1986), in our experiment TNAP mRNA was significantly expressed in odontoblasts facing dentin-containing matrix vesicles.
Odontogenic cells synthesize a set of molecules involved in the control of calcium and phosphate bio-availability, presumably useful for nucleation and growth of apatite crystals (see review by Takano 1995), such as calbindin-D9k and calbindin-D28k (Berdal et al. 1996), parvalbumin (Davideau et al. 1993), calcium-ATPase (Borke et al. 1995), and alkaline phosphatase (see review by Takano 1995; and this study). As observed here for AP, the developmental expression patterns of calbindin-D9k, calbindin-D28k (Berdal et al. 1991; Hotton et al. 1995), and parvalbumin (Davideau et al. 1993) have been shown to be based on a biphasic mode concerning the steady-state levels of mRNA and additive cyclic modulations concerning the cytoplasmic levels of proteins during the maturation stage in the incisor. Whether these cellular fluctuations of protein levels are related to variations in the synthesis and/or proteolytic processing in the ruffle- and smooth-ended ameloblasts remains to be investigated. Unfortunately, the rather small quantities of dental epithelial cells that can be used for biochemical investigations in vivo or in vitro further limit such investigations. However, the reliability of the rat incisor sampling methods for biochemical investigations was confirmed in our study, a technical aspect that validates our experimental strategy using this system to investigate gene/protein up- and downregulation by hormones and growth factors in the overall process of tooth formation (see review by Berdal 1997). The EGF receptor pathway may presumably contribute to the presently established fine-tuned intracellular protein levels for AP and previously established for other calcium binding proteins, as suggested by the observed variations of EGF receptors (full-length and truncated forms) in the same experimental system (Davideau et al. 1995).
The present cellular and tissue distribution of immunoreactive AP is consistent with the proposed pathways for AP synthesis and processing (Van Hoof and De Broe 1994). AP presents several isoforms. A form anchored to the membrane via a glycosyl-phosphatidylinositol domain has been shown to be enzymatically active in osteoblasts in vitro (Anagnoustou et al. 1996). On the other hand, a cleaved AP, either active or inactive as assessed by histoenzymology (De Bernard et al. 1986), is present in the extracellular milieu, including the serum compartment (Van Hoof and De Broe 1994). Previous immunolocalizations and the present study have been performed in bone (Bonucci et al. 1992) and cartilage (De Bernard et al. 1986). The AP protein is distributed in the cellular compartment, where it is presumably synthesized, in the membranes of cells and matrix vesicles and, finally, in the extracellular matrix. The present investigation extends the concept of the existence of two states of AP, i.e. anchored and cleaved/extracellular, in mineralized dental tissues, as suggested by previous distinct studies on dentin, cementum, bone, and cartilage (Kurahashi and Yoshiki 1972; Deporter and Ten Cate 1976; Linde and Granström 1980; Orams and Snibson 1982; Gomez and Boyde 1994; Groeneveld et al. 1995; Linde and Lundgren 1995; Takano 1995; Woltgens et al. 1995). More specifically, (a) during dentin biomineralization, the junction between predentin and dentin was particularly rich in TNAP protein, which would be enzymatically active as shown by histoenzymology (Larsson 1973), and (b) during the maturation stage of enamel formation, the outermost layer of enamel contained TNAP, which would be enzymatically active as shown by histoenzymology (Gomez and Boyde 1994). The tissues studied in this experiment (except mantle dentin) do not mineralize via matrix vesicles, presumably like bone and in contrast to cartilage, in which the role of AP in biomineralization has been extensively investigated (see review by Anderson and Morris 1993). The spatial organization and growth of crystals in these dental systems have been proposed to be under the control of matrix macromolecules, mainly collagen Type I and noncollagenous proteins such as bone sialoprotein, phosphoproteins in dentin (for review see Butler and Ritchie 1995), and very unique enamel proteins, including amelogenins (for review see Zeichner-David et al. 1995), amelin/ameloblastin/sheathlin (Cerny et al. 1996; Krebsbach et al. 1996; Nanci et al. 1998), and enamelin (Hu et al. 1997), which are variably phosphorylated. The present investigation showed that AP is distributed in the extracellular compartment of both enamel and dentin. Functional investigations of AP have been recently highlighted by the phenotype of TNAP null mutants, which is particularly striking in teeth. This enzyme appears to be a protein phosphatase during osteoblast differentiation, locally increasing the available levels of Pi and binding ionic calcium for crystal formation during biomineralization (for review see Whyte 1994). The extracellular sites of AP protein/activity were located at the predentin/dentin border, specifically at the site of secretion of dentin phosphoproteins (Rabie and Veis 1995). These molecules are called phosphophoryns, because they provide Pi and covalently bind phosphates to crystals that nucleate heterogeneously and grow at the mineralization front. In this specific area, our data support the hypothesis that AP could be bound to collagen Type I, as shown previously (Wu et al. 1991), and could dephosphorylate phosphophoryns, making Pi available for hydroxyapatite crystals, as previously suggested.
A striking feature of the comparison among bone, enamel, and dentin is that proteins important for the formation of mineralized tissues, e.g., TNAP, are expressed not only in the “main” cells, which secrete matrix proteins, osteoblasts, ameloblasts, and odontoblasts (for review, see Kurahashi and Yoshiki 1972; Deporter and Ten Cate 1976; Linde and Granström 1980; Orams and Snibson 1982; De Bernard et al. 1986; Takano et al. 1986; Bonucci et al. 1992; Gomez and Boyde 1994; Linde and Lundgren 1995; Woltgens et al. 1995; and this study), but also and very intensely in “satellite” cells, adjacent to the main cells: osteoprogenitor, supra-ameloblastic, and subodontoblastic cells. The dental specificities of TNAP vs bone were (a) marked overexpression during completion of biomineralization and (b) the organized extracellular gradient of TNAP distribution compared with the diffuse appearance observed in bone matrix. These properties may be key elements in the differential sensitivity of bone and teeth to phosphatase disturbances, as illustrated by experimental hypophosphatasia (Whyte et al. 1995).
Footnotes
Acknowledgements
Supported by EA 2380 funds, PHRC AOM 96067, and Laboratories Novartis, Santé Familiale.