
Abstract
Dynamin I, a GTPase involved in the endocytic cycle of synaptic vesicle membranes, is believed to support axonal outgrowth and/or synaptogenesis. To explore the temporal and spatial patterns of dynamin I distribution in neuronal morphogenesis, we compared the developmental expression of dynamin with the expression of presynaptic membrane proteins such as SV2, synaptotagmin, and syntaxin in the chick primary visual pathway. Western blots of retina and tectum revealed a steady increase of synaptotagmin and syntaxin from embryonic Day 7 (E7) to E11, whereas for the same time frame no detectable increase of dynamin was found. Later stages showed increasing amounts of all tested proteins until the first postnatal week. Immunofluorescence revealed that SV2, synaptotagmin, and syntaxin are present in retinal ganglion cell axons from E4 on. In later stages, the staining pattern in the retina and along the visual pathway paralleled the formation and maturation of axons. In contrast, dynamin is not detectable by immunofluorescence in the developing retina and optic tectum before synapse formation. Our data indicate that, in contrast to the early expression of synaptotagmin, SV2, and syntaxin during axonal growth, dynamin is upregulated after synapse formation, suggesting its function predominantly during and after synaptogenesis but not in axonogenesis.
S
Developmental studies using growing neurons in primary culture have demonstrated that other presynaptic membrane proteins (referred as SNAREs; Roth-man and Warren 1994), such as syntaxin, SNAP-25, and synaptotagmin, are localized in developing axons before synapse formation (Matteoli et al. 1992; Garcia et al. 1995; Littleton et al. 1995; Sesack and Snyder 1995). Furthermore, cleavage of SNAP-25 or syntaxin by clostridial neurotoxins inhibits axon growth and causes growth cone collapse, indicating its participation in membrane extension (Igarashi et al. 1996; Osen-Sand et al. 1996). Therefore, the presence of these presynaptic membrane proteins in developing axons before synapse formation can be used as a marker for the formation and maturation of axons in a defined neuronal pathway.
The retinotectal system of the chick has long been utilized to analyze the topographical organization of developing retinal ganglion cell (RGC) axons in the optic nerve, the chiasm, and the optic tract, and their formation of synapses in the optic tectum (Rager 1980) (Figure 1). The existence of this specific organization and the timing of RGC axons throughout the primary visual pathway could also be correlated with the expression of cell adhesion molecules found during the formation and maturation of fibers (Rager et al. 1996). Therefore, the developing retinotectal system provides an excellent model for examination of the temporal and spatial distribution of presynaptic proteins during fiber outgrowth and synaptogenesis. We report here the differential regulated expression of dynamin I compared to that of synaptotagmin, SV2, and syntaxin in the primary visual pathway of the chick, which indicates that neuronal dynamin is predominantly expressed after synaptogenesis.
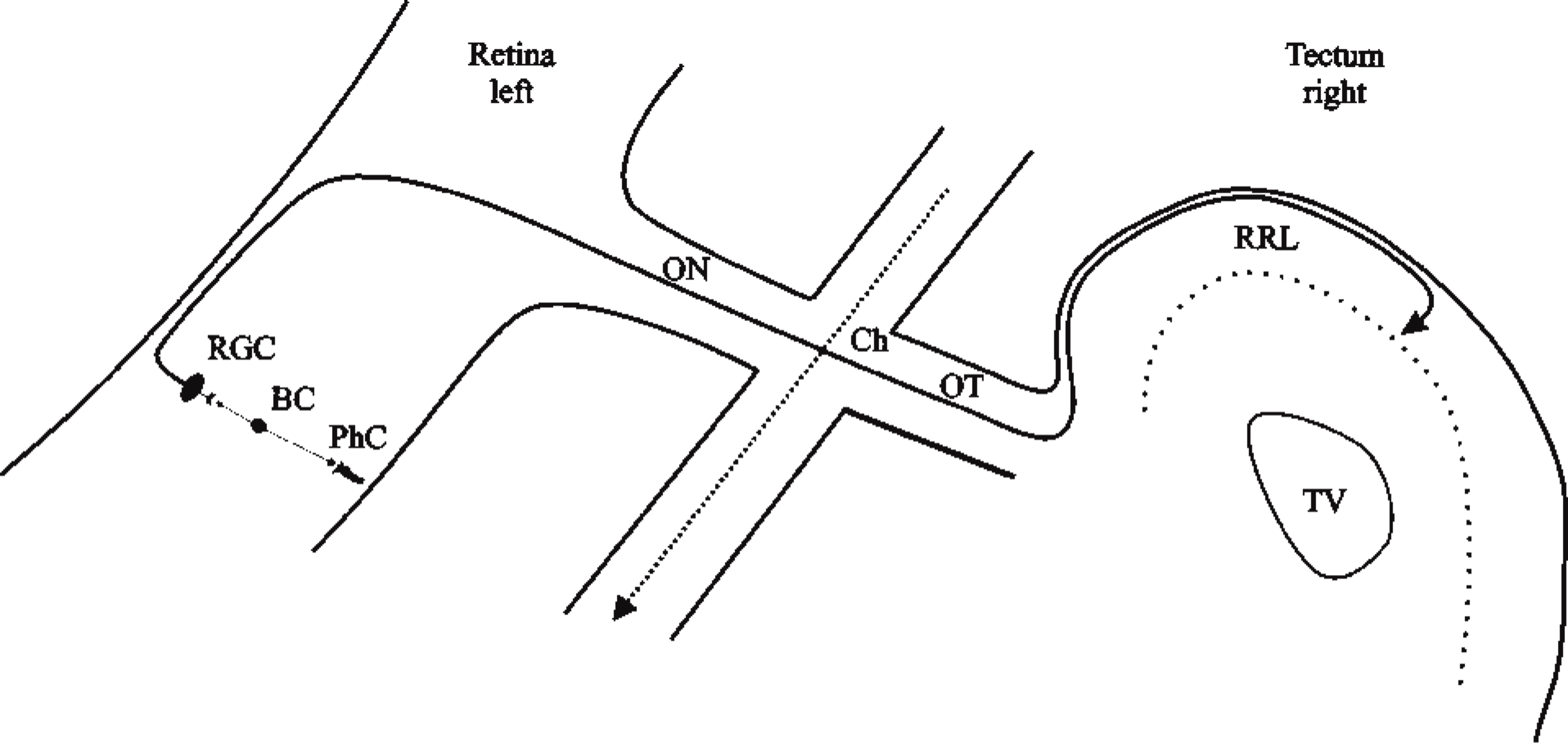
Schematic diagram illustrating the organization of the chick retinotectal system in a simplified manner. The visual pathway starts in the retina with three well-defined cell layers that contain the photoreceptor cells (PhC), which are connected via bipolar cells (BC) to the retinal ganglion cells (RGC). Their axons leave the retina and enter the optic nerve (ON), cross completely in the chiasm (Ch) to the contralateral side, and continue into the optic tract (OT) before the majority of the fibers project to the retinorecipient layers (RRL) of the optic tectum. TV, tectal ventricle.

Crossreactivity of dynamin DG-1 antiserum in chick brain. Western blot analysis revealed that DG-1 recognizes chicken brain dynamin. Similar to the human (Lane A), rat (Lane B), and mouse (Lane C) neocortex an immunopositive band at 100 kD was found in the homogenate of the chick hyperstriatum (Lane D).
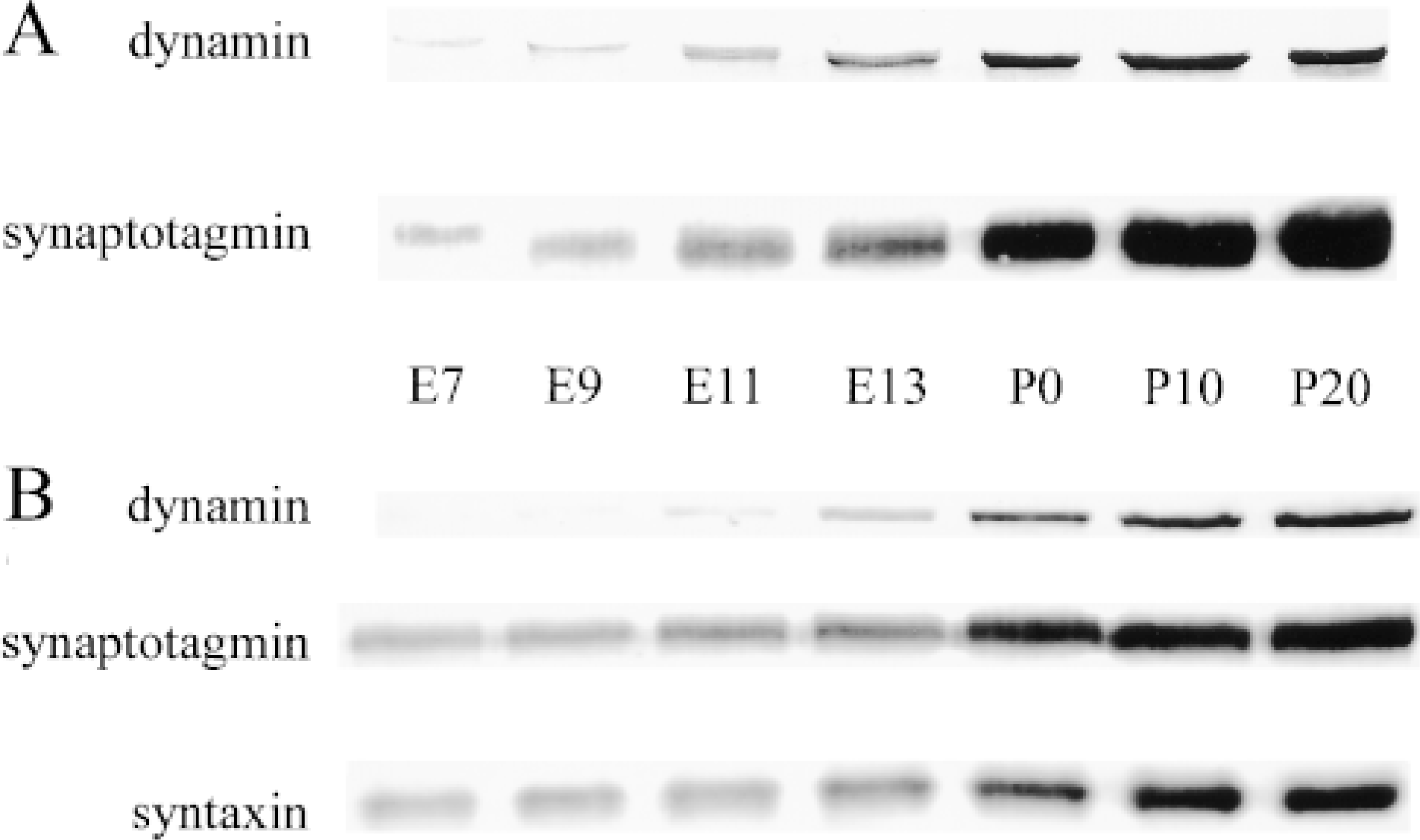
Developmental expression of presynaptic proteins in the chick retina and tectum. (A) In Western blots from retinae, a steady increase of protein amounts was detected for synaptotagmin between E7 and P0. In contrast, dynamin exhibited first signals from E7, but increasing immunoreactivity was not found before E13. (B) In the chick tectum, protein expression for synaptotagmin and syntaxin was found from E7 in increasing amounts. First clear dynamin signals were detected at E11 and increased to steady state at P20.
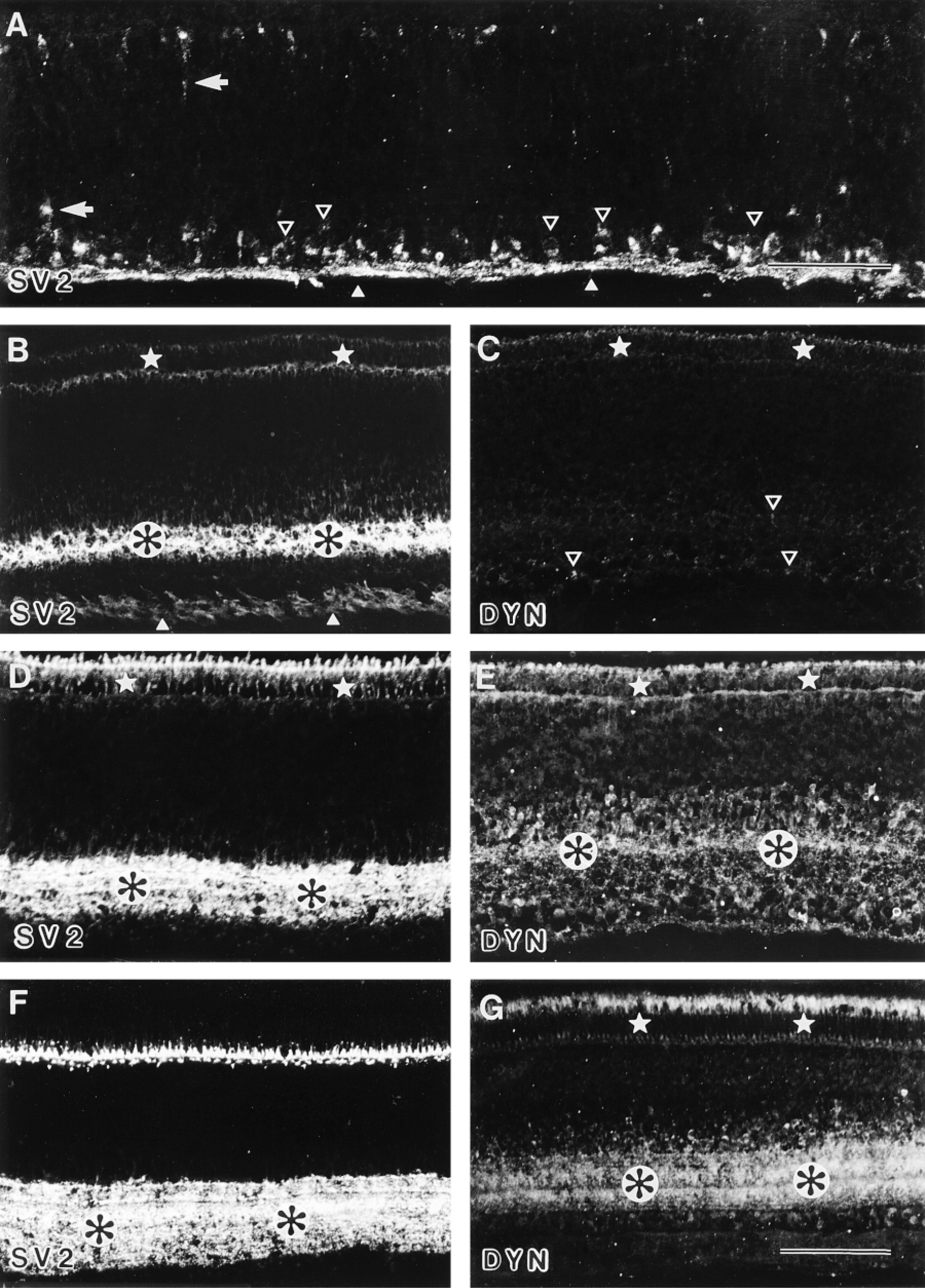
Distribution of SV2 and dynamin in the developing retina. (A) E4: SV2 was present in ganglion cell perikarya (open arrowheads) and the optic fiber layer (arrowheads) of the chick central retina. Immunofluorescence for SV2 was also expressed in young neurons migrating toward the ganglion cell layer (arrows). Bar = 50 μm. (B,C) E10: SV2 was present in the inner plexiform layer (black asterisks in white circles) and was less prominent in the developing photoreceptor cells (stars). Note the weak immunofluorescence in the optic fiber layer (arrowheads). In contrast to SV2, the immunofluorescence for dynamin (DYN) was very faint in photoreceptors (stars) in ganglion and amacrine cells (open arrowheads). (D,E) E13: SV2 was seen in the inner plexiform layer (black asterisks in white circles) and was more prominent in the developing photoreceptor cells (stars). Compared to SV2, dynamin (DYN) immunoreactivity was detected also in developing photoreceptor cells (stars), less prominent in the inner plexiform layer (black asterisks in white circles), and in cells of the outer part of the inner nuclear layer. (F,G) P0: SV2 was present in the inner plexiform layer with two horizontal stripes of less immunofluorescence (black as terisks in white circles) and in the outer plexiform layer. Dynamin (DYN) was present predominantly in the inner plexiform layer, endowed also with two bands of lesser immunofluorescence (black asterisks in white circles) and within the photoreceptor cell bodies (stars). In addition, some immunofluorescent cells for DYN were found in the inner nuclear layer of the retina. Note the differential staining pattern within the photoreceptor cells for SV2 compared to that of dynamin. Bar = 100 μm.
Materials and Methods
Animals
White Leghorn chicks from hatching (P0) until post-hatching Day 20 (P20) and chick embryos [(E3-E20, Stages 19-45, according to Hamburger and Hamilton (1951)] were used in this study.
Antibodies
For immunohistochemistry and Western blotting the following mouse monoclonal antibodies (MAbs) and/or rabbit polyclonal antisera were used. A rabbit antiserum directed against dynamin [DG-1 antibodies were generated against a human dynamin I 6x His-tag fusion protein lacking the proline-rich domain (Butler et al. 1997); kindly provided by P. De Camilli, New Haven, CT] and a mouse MAb generated from rat dynamin I specific in chick tissue (cl. 41; Transduction Laboratories, Lexington, KY) were used. For the integral synaptic vesicle membrane protein SV2 (Buckley and Kelly 1985), a mouse MAb (DSHB; Iowa City, IA) recognizing both of the two similar brain-specific proteins of the SV2 class (Bajjalieh et al. 1993) was used. The mouse MAb against synaptotagmin (cl. 41.1; kindly provided by R. Jahn, Göttingen) was generated with recombinant synaptotagmin used as antigen and directed against synaptotagmin I as affinity ligand (Brose et al. 1992). For syntaxin, a mouse MAb HPC-1 (Sigma; Munich, Germany) was used. The specificity of this antibody and its use for immunohistochemistry and immunoblotting of neural tissue have been described previously (Barnstable et al. 1985).
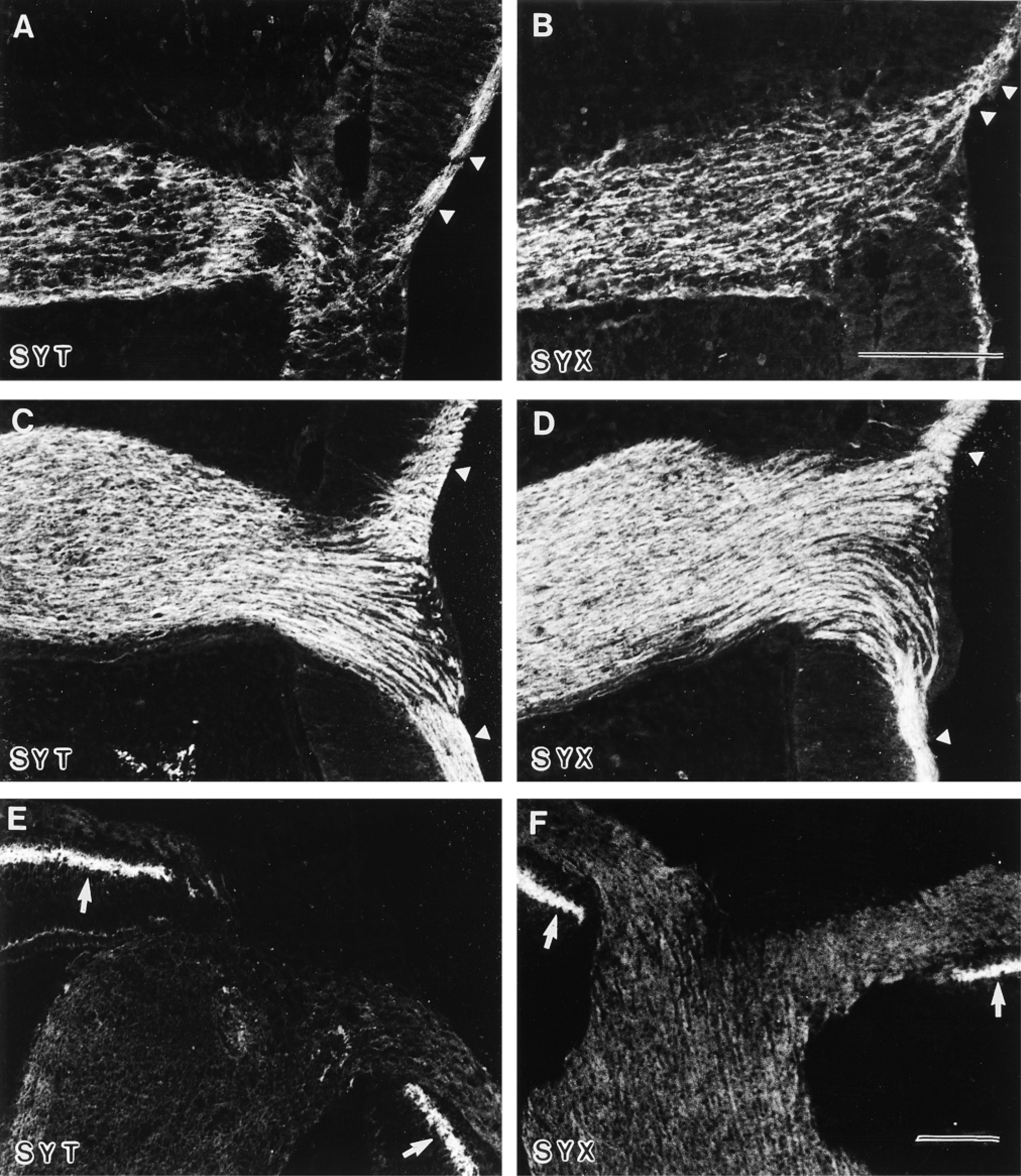
Distribution of synaptotagmin and syntaxin in the developing optic nerve. (A,B) E5: Synaptotagmin (SYT) and syntaxin (SYX) immunofluorescent axons from the optic fiber layer (arrowheads) reach the optic nerve through the optic nerve head. (C,D) E6: Immunofluorescence for synaptotagmin (SYT) and syntaxin (SYX) was found in all axons of the optic fiber layer (arrowheads) and in fiber bundles of the optic nerve at this developmental stage. (E,F) E9: In contrast to E6, the immunostaining for synaptotagmin (SYT) and syntaxin (SYX) disappears from the axons and fiber bundles. Note the appearance of immunofluorescence in the inner plexiform layer of the retina (arrows). Bars = 100 μm.
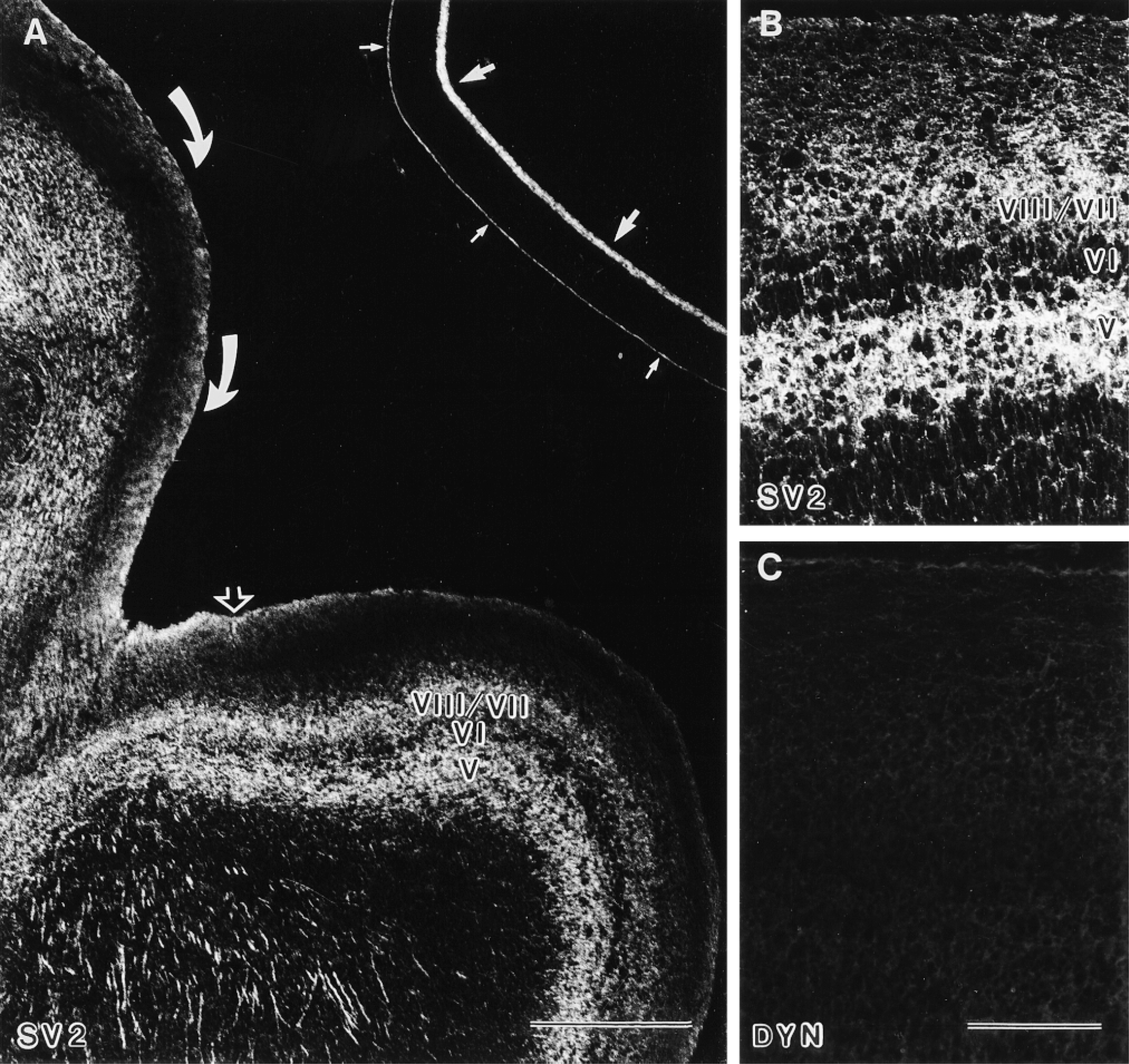
Distribution of SV2 and dynamin in the developing optic tract and optic tectum. (A) E9: The optic tract (curved arrows) and the tectal surface (open arrow) showed immunostaining for SV2. In the optic tectum, immunofluorescence was detected in plexiform Layer VII and in the relatively cell-sparse zone V, whereas Layer VI, a dense population of piriform cells, showed less immunofluorescence for SV2 [roman numbers indicate the tectal layers according to La Vail and Cowan (1971)]. The retina exhibited immunofluorescence for SV2 in the inner plexiform layer (arrows) and in the photoreceptor cells (small arrows). Bar = 500 μm. (B,C) E9: Higher magnification of the optic tectum revealed immunofluorescence for SV2 predominantly in Layers VIII/VII and V (B) but not for dynamin (DYN; C). Bar = 100 μm.
Western Blotting
For protein detection using the Western blot technique, samples of human, rat, and mouse neocortex and from chick hyperstriatum, retina, and tectum were homogenized and processed according to standard protocols (for further details see Grabs et al. 1994,1996; Bergmann et al. 1996). Lanes of 10 μg equal total protein were run on SDS-PAGE gels under reducing conditions and transferred to nitrocellulose. For immunodetection, all primary antibodies (see above) were tested in different dilutions and used under saturating conditions to compare different antigens. Antigen-antibody complexes were coupled to HRP-conjugated secondary goat anti-rabbit or goat anti-mouse IgG antibodies, respectively, and visualized using DAB/H2O2 substrate protocols.
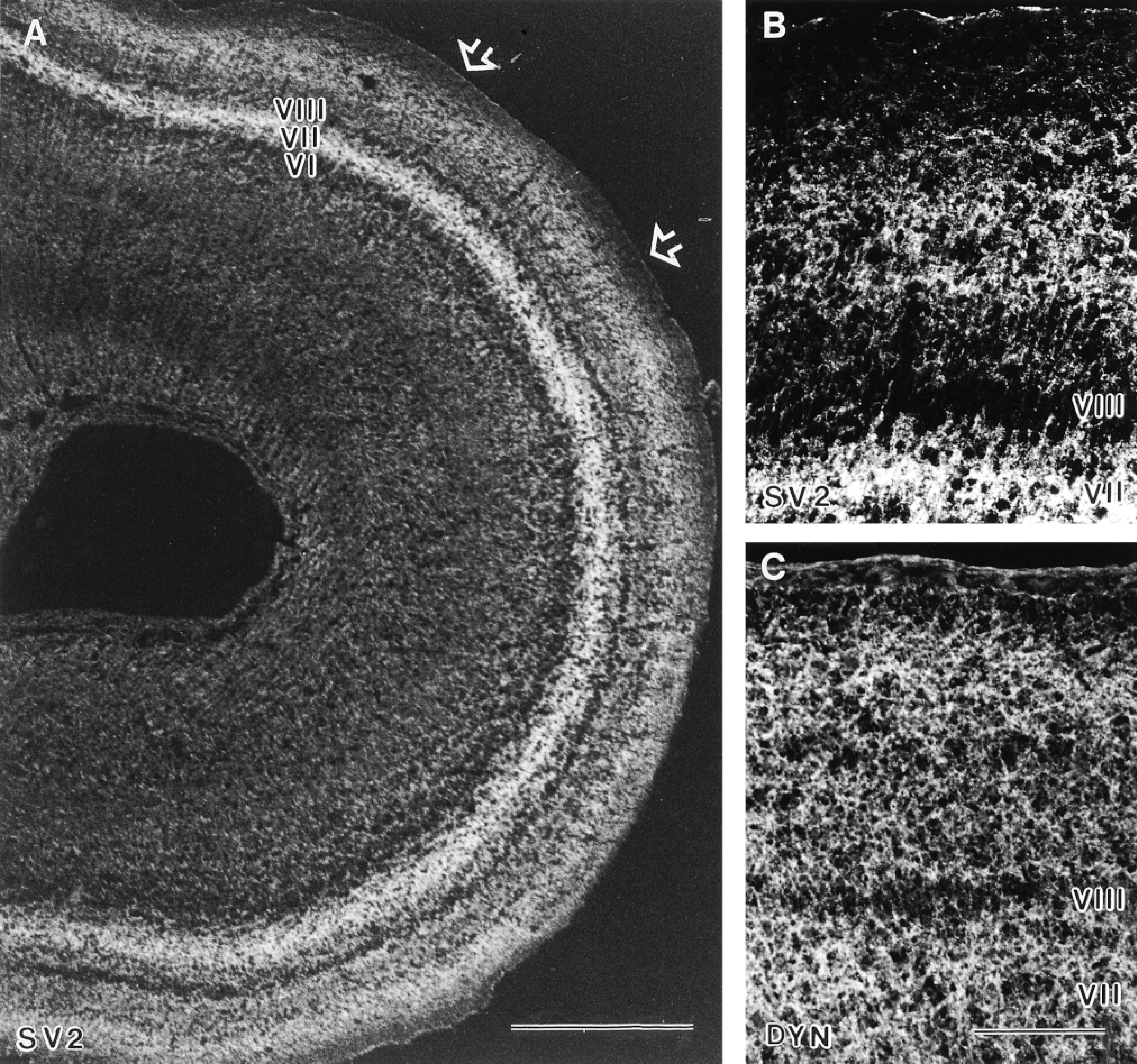
Distribution of SV2 and dynamin in the developing optic tract and optic tectum. (A) E13: In the stratum opticum (open arrows), immunofluorescence for SV2 has virtually disappeared. Immunofluorescence for SV2 was found in tectal Layer VII (future layer h) and in the retinorecipient layers superficially to Layer VIII. Layer VIII (future layer g) and Layer VI (future layer i) showed less immunostaining for SV2 [roman numbers indicate the tectal layers according to La Vail and Cowan 1971)]. Bar = 500 μm. (B,C) E13: Higher magnification of the ventrolateral part of the optic tectum revealed immunofluorescence not only for SV2 (B) but now also for dynamin (DYN; C). Immunostaining was found predominantly in the plexiform tectal Layer VII and in the retinorecipient layers superficial to Layer VIII. Bars = 100 μm.
Immunohistochemistry
For preparation of tissue sections, the eye cups, optic nerves, optic tracts, optic tecta, and whole brains were dissected out and immersion-fixed for 4-6 hr with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) or perfusion-fixed with the same fixative. The tissue was mounted with Tissue Tek embedding medium (Miles; Elkhart, IN) and rapidly frozen on a mounting block cooled over liquid nitrogen. Frontal cryosections were cut at10 μm. For immunofluorescence, primary antibodies were diluted 1:100 (dynamin, SV2, and syntaxin) or 1:500 (synaptotagmin) in goat serum dilution buffer (20 mM NaPO4, 15% normal goat serum, 450 mM NaCl, 0.3 % Triton X-100). Antigen-antibody complexes were visualized using fluoresceinor CY3-conjugated goat anti-rabbit or goat anti-mouse IgGs and were examined with a Leitz DM-RBE microscope equipped for epifluorescence (for further details see Grabs et al. 1994; Grosse et al. 1998). Controls were incubated with 2% normal goat serum in PBS instead of the specific antibodies.
Results
Western blot analysis revealed that the rabbit antiserum DG-1 also recognizes neuronal chicken dynamin. Dynamin was detected in immunoreactive bands at 100 kD in neocortex homogenates of human, rat, and mouse brain and in the hyperstriatum homogenate of the chick brain (Figure 2).
Western blots of equal total protein of the chick retina revealed only borderline signals for dynamin between embryonic Day 7 (E7) and E11, with no detectable increase of the protein during this period (Figure 3A). In the chick tectum, only very weak immunoreactivity for dynamin was found before E11 (Figure 3B). In contrast, both areas exhibited a steady increase of the proteins involved in exocytosis from E7 (for synaptotagmin see Figures 3A and 3B; for syntaxin see Figure 3B). From E13 until the third postnatal week (postnatal Day 20, P20), increasing amounts of all the proteins investigated, including synaptotagmin, syntaxin, and dynamin were detected. Until adulthood, all tested proteins kept the high levels of expression as seen on P20 (Figures 3A and 3B).
Immunofluorescence in the chick retina revealed that synaptotagmin, SV2 (Figure 4A), and syntaxin are present in retinal ganglion cells (RGCs) and their axons from E4 on. First, immunoproducts are seen in the central part of the retina, where first RGCs are generated. With further development, immunoreactivity was detected in RGCs in all parts of the retina. This process of maturation, which is reflected in both the size and the structure of the inner plexiform layer, becomes visible by intense immunofluorescence for synaptotagmin, SV2, and syntaxin in the central retina from E8 on (for E10 see Figure 4B; for E13 see Figure 4D). In contrast, dynamin was not detectable by immunofluorescence in the retina before E10. Initial, very faint staining in the photoreceptor and ganglion cell perikarya was seen from E10 on (Figure 4C). At E13, dynamin was found in developing photoreceptors and their pedicles, in cells of the outer part of the inner nuclear layer, and in the inner plexiform layer (Figure 4E). The appearance of dynamin immunofluorescence in the inner plexiform layer of the central retina coincides temporally and spatially with the formation of the first synapses in the inner plexiform layer of the chick retina around E13 (Hughes and Lavelle 1974; Daniels and Vogel 1980). In later retinal development, immunofluorescence for dynamin was seen predominantly in the inner plexiform layer and in the photoreceptor cells in the developing retina (Figure 4G). Similarly, immunofluorescence for synaptotagmin, SV2, and syntaxin was found not only in the inner plexiform layer but also in the developing photoreceptor cells (for SV2 see Figure 4D). After hatching, strong immunofluorescence was concentrated in the photoreceptor terminals in the outer plexiform layer (for P0 see Figure 4F). Interestingly, two horizontal stripes of lesser immunoreactivity were seen for all the proteins investigated in the inner plexiform layer (Figures 4F and 4G), most probably corresponding to the stratification of amacrine cell processes and RGC dendrites, which divide the inner plexiform layer of the avian retina into typical sublaminae (Ramón y Cajal 1893; Rager 1980; Rager et al. 1996).
Starting with E4, we found first immunopositive fibers for synaptotagmin (Figure 5A), SV2, and syntaxin (Figure 5B) leaving the retina through the optic fiber layer and the optic nerve head and entering the optic nerve. In this way the fibers fill the optic stalk rapidly, entering the gaps between undifferentiated neuroepithelial cells and the holes left by dying cells until E6 (compare Figs. 5A and 5B to Figure 5C and 5D). After E10, immunoreactivities for synaptotagmin (Figure 5E), syntaxin (Figure 5F), and SV2 in the optic nerve and chiasm disappeared. Immunosignals in the ganglion cell axons along the optic tract (for E9 see Figure 6A) were found between E7 and E12 for synaptotagmin, syntaxin, and SV2. Finally, first immunostaining for these proteins in the superficial layers of the optic tectum was found at E8 (for E9 see Figures 6A and 6B). In contrast, dynamin was not detectable by immunofluorescence in the developing optic tectum at these developmental stages (Figure 6C). From E11, detectable immunostaining for dynamin was found in the superficial layers of the optic tectum (for E13 see Figure 7C). From E13 until adulthood, all the proteins investigated, including synaptotagmin, syntaxin, SV2 (Figures 7A and 7B), and dynamin (Figure 7C), were detected in the optic tectum.
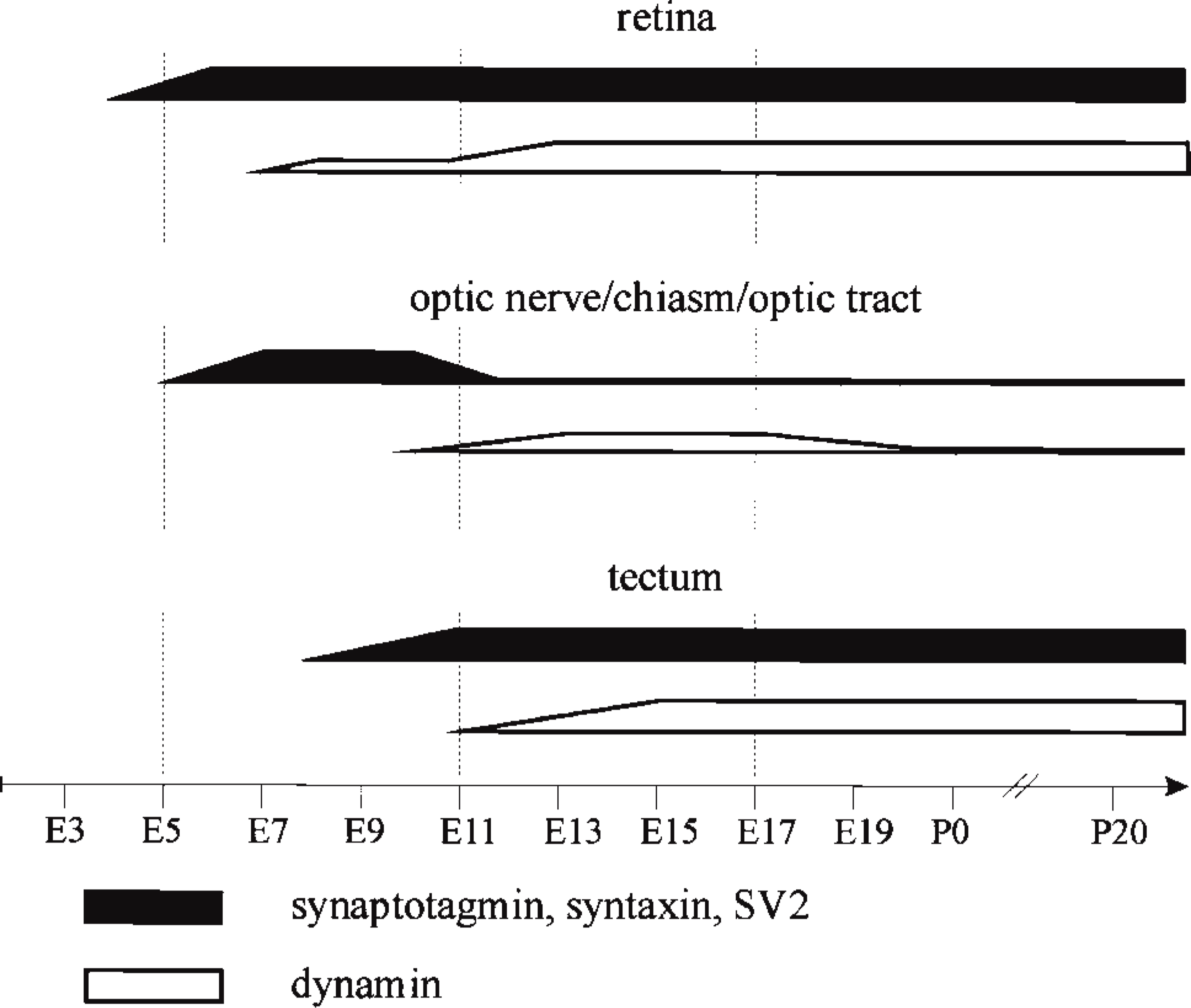
Appearance of presynaptic proteins in the chick retinotectal system. In the retinotectal pathway, immunoreactivity for presynaptic proteins such as synaptotagmin, syntaxin, and SV2 was similar and could be found from E4 in the retina, 1 day later in the optic nerve, and in first immunopositive retinal fibers reaching the optic layers of the tectum around E8 (black). However, dynamin appears around E7 in the retina but is detectable in immunofluorescence only after its upregulation in onset of synapse formation in the inner plexiform layer around E13 (Hughes and LaVelle 1974; Daniels and Vogel 1980) (white). Similarly, detectable immunofluorescence of dynamin was found in the retinorecipient layers of the optic tectum only after the appearance of first functional synapses from E11 on (Rager 1980) (white). In summary, the appearance of immunoreactivity for presynaptic proteins such as synaptotagmin, syntaxin, and SV2 in all investigated areas of the retinotectal projection follows the development of the system and precedes the expression of dynamin.
Discussion
In this study we applied the Western blotting and immunofluorescence techniques to analyze the developmental expression of presynaptic proteins, comparing dynamin I with the presynaptic membrane proteins synaptotagmin, syntaxin, and SV2, to learn more about the biogenesis and recycling of the plasma membrane during constitutive axon and synapse formation. Our data demonstrate that dynamin is upregulated only after synapse formation, whereas the other proteins are expressed during axonal growth (Figure 8). In addition, the temporospatial distribution of the presynaptic membrane proteins during development of the chick primary visual system reveals that the appearance of these proteins is closely correlated with the chronotropic pattern of axons described in previous morphological studies (Rager 1980).
Expression and Distribution of Dynamin Correlate with Synaptogenesis
Although the mechanisms underlying endocytic retrieval of presynaptic membrane material after exocytosis have not been fully established, it is widely accepted that the clathrin-dependent endocytosis is largely mediated and controlled by a number of proteins, including dynamin (De Camilli 1995; Shupliakov et al. 1997). Thus far, only a few contradicting interpretations about the developmental expression of these proteins are available. The increased expression of dynamin I during neuronal development was reported to support either neurite formation (Torre et al. 1994) or synaptogenesis (Nakata et al. 1991; Faire et al. 1992). To determine the involvement of neuronal dynamin in axon outgrowth and /or synaptogenesis, different anti bodies against dynamin were used in this study. Surprisingly, no detectable immunofluorescence for dynamin was found before synapse formation in the developing retina and optic tectum. With establishment of functional synapses (Hughes and LaVelle 1974; Daniels and Vogel 1980), immunoreactivity in the synaptic layers increased from the central to the peripheral part of the retina. In conformity with the temporospatial gradient of maturation, ganglion cell synapses in the retinorecipient layers (Rager 1980) displayed dynamin from the rostral to the caudal pole of the tectum. However, because these superficial layers also receive minor inputs from local circuits, intratectal synapses in this region cannot be excluded.
The absence of a dynamin immunofluorescent signal during axon development may be due to the lack of enrichment of the soluble protein in the axons and growth cones before functional synapses have appeared. In addition, our Western blot analysis of the retina and the tectum revealed a steady increase of the SNARE proteins syntaxin and synaptotagmin before synapse formation, whereas no detectable increase was found for dynamin during this period. It is possible that growing axons deliver only a minor fraction of dynamin compared to the SNARE membrane components, suggesting a higher rate of membrane insertion than membrane retrieval, which may cause axon elongation. Alternatively, our observations may imply that dynamin is not involved in constitutive membrane recycling during axon elongation and that its upregulation is triggered by synaptogenesis, possibly due to mechanisms involving electrical activity.
Expression and Distribution of SV2, Synaptotagmin, and Syntaxin Correlate with Axonogenesis
Given the complexity of the presynaptic localization of SNAREs (Otto et al. 1997) transported by fast axonal transport (Li et al. 1996), vesicles in developing axons containing these proteins may represent precursors for synaptic vesicles and/or for the axon plasma membrane. Therefore, these constitutive vesicles may provide the material for axon membrane elongation, independent of the synaptic pathway (Kraszewski et al. 1995). A recent study in cultured hippocampal neurons using a defective Herpesvirus vector to express an exogenous protein, the lymphocyte transmembrane protein CD8α, showed that the newly synthesized membrane protein first appeared at the growth cone and that it was then redistributed over the plasma membrane surface (Craig et al. 1995). When analyzed at the light microscopic level, the axon staining pattern of the RGCs for synaptotagmin, syntaxin, and SV2, first in growth cones and then in the entire axon plasma membrane (our unpublished data), supports the hypothesis that an additional mechanism of endocytic recycling contributes to the further spread of the protein in the membrane (Craig et al. 1995).
In summary, our data on the developing chick retinotectal system demonstrate that the presynaptic membrane proteins synaptotagmin, syntaxin, and SV2 are expressed immediately after axon outgrowth as residents of heterogeneous organelles that probably represent a mixture of precursors for synaptic vesicles and the axon plasma membrane. Furthermore, the redistribution of these proteins during synapse formation, from the axon to the presynaptic terminal, is closely correlated with the chronotropic pattern of axons in the chick retinotectal projection and with the functional development of retinotectal synapses studied by electrophysiological techniques using both field potential analysis and single unit recording, which indicate that synaptic transmission is not effective before E11 (Rager 1976,1980). In contrast, proteins of regulated synaptic membrane retrieval, such as dynamin, become upregulated after synapse formation, indicating their participation in regulated endocytosis only. It remains to be established if other proteins are necessary to internalize components of the constitutive axonal plasma membrane. Developmental expression studies of such proteins that are implicated in endocytosis, such as amphiphysin, AP 2, and clathrin, will probably be useful in identifying proteins of constitutive membrane retrieval.
Footnotes
Acknowledgements
Supported by Swiss National Fond, grant 3100-049523. 96/1.
We thank R. Jahn (Göttingen, Germany) and P. DeCamilli (New Haven, CT) for generously supplying antibodies, and L. Clement, M. Kaczorowski, Ch. Marti, B. Scolari, and C. Weber for excellent technical assistance.