
Abstract
Three skeletal tissues of the adult echinoid Paracentrotus lividus (the pedicellaria primordium, the test, and the tooth) were immunolabeled with three sera raised against the total mineralization organic matrix and two specific matrix proteins (SM30 and SM50) from the embryo of the echinoid Strongylocentrotus purpuratus. Two conventional chemical fixation protocols and two high-pressure freezing/freeze-substitution protocols were tested. One conventional protocol is recommended for its good preservation of the ultrastructure, and one high-pressure freezing/freeze-substitution protocol is recommended for its good retention of antigenicity. Immunolabeling was obtained in the three adult tissues. It was confined to the active skeleton-forming cells and to the structured organic matrix. The results indicate that the matrix proteins follow the classical routes of secretory protein assembly and export and suggest that SM30 and SM50 are a part of the tridimensional network formed by the organic matrix before the onset of mineralization. They show that the genetic program of part of skeletogenesis is conserved among different calcification models and developmental stages.
Keywords
S
The larval spicules and adult skeleton share common characteristics. Both are composed of magnesium calcite with a 0.1% (w/w) mineralization organic matrix and both are formed in a vacuole, the calcification site, enclosed in a syncytial pseudopodium generated by the skeleton-forming cells (for review see Dubois and Chen 1989). In each case, the calcification site contains the organic matrix, which will be included in the skeleton during calcification. This organic matrix is composed mainly of proteins and glycoproteins (Weiner 1985; Benson et al. 1986; Swift et al. 1986; Venkatesan and Simpson 1986; Albeck et al. 1996; Killian and Wilt 1996), is structured in concentric sheets bridged by radial threads (Benson et al. 1983; Dubois 1991; Ameye et al. 1998), and controls the properties of the skeleton through structural interactions (Addadi and Weiner 1992; Berman et al. 1993). The latter aspect makes the understanding of its three-dimensional structure and chemistry particularly important (Lowenstam and Weiner 1989).
Two genes controlling the production of intraspicular proteins, i.e., SM30 and SM50, are known from larvae of several echinoid species. SM30 is a 30-kD acidic N-glycoprotein (George et al. 1991), whereas SM50 is a 50-kD alkaline protein (Sucov et al. 1987; Katoh-Fukui et al. 1991). RNA blot analysis reported expression of these genes in adult mineralized tissues (Richardson et al. 1989; George et al. 1991; Kitajima et al. 1996). They showed an accumulation of SM30 and SM50 mRNA in spines and tube feet (the latter contain a terminal calcified ring). Surprisingly, SM30 is apparently not expressed in the heavily calcified test, although SM50 is (Kitajima et al. 1996). However, no report has investigated the cellular localization of the SM30 and SM50 gene products, either in the test or in any other adult calcified tissues. In particular, no data are available on the ultrastructural mapping of these proteins in the calcification site, information that is crucial to assess the role of these proteins in biomineralization.
Accordingly, the goal of the present work was to investigate the ultrastructural localization of SM30 and SM50, using immunogold labeling, in three adult mineralized tissues, i.e., the primordium of an outer appendage (the pedicellaria), the test, and the tooth.
Materials and Methods
Sampling
Pedicellaria primordia and test plates were dissected from individuals of Paracentrotus lividus ranging from 0.8 to 1.2 cm in ambital diameter (i.e., the equatorial diameter of the hemispherical test); teeth were dissected from P. lividus with a minimal ambital diameter of 3 cm. The former echinoids were obtained from the aquaculture in the Centre de Recherches et d′Etudes Côtières (University of Caen, France), whereas the latter were collected intertidally in Morgat (Brittany, France). All the echinoids were kept in a closed-circuit aquarium (35%o, 13C) and fed dried seaweed (Lessonia sp.) once a week. The primordia were processed by conventional chemical fixation (CF) and by high-pressure freezing/freeze-substitution (HPF/FS), the teeth were processed by HPF/FS, and the test plates by CF. All the samples were dissected under a dissecting microscope. The sampled primordia ranged from 100 to 200 μm in size. For CF, the primordia were collected with the underlying test plates using scissors. For HPF/FS, the primordia were cut away from the underlying test plates with a scalpel. (For practical reasons, the test plates could not be processed by HPF; the small size of the holders used in the high-pressure freezer prevents their handling). The teeth were collected with scissors.
Conventional Chemical Fixation (CF)
Two protocols were tested on the pedicellaria primordia and on the test plates. The first protocol (CF1) had been demonstrated to be optimal for the preservation of the spatial organization of the organic matrix within the calcification sites (Ameye et al. 1998), whereas the second one (CF2) aimed to better preserve the antigenicity. In the CF1 protocol, the samples were fixed for 24 hr at 20C in 3% glutaraldehyde (Polysciences; Warrington, PA) in cacodylate buffer (0.1 M, pH 7.8) containing 1% tannic acid (C76H52O46; UCB, Brussels, Belgium) (the solution was adjusted to 1030 mOsM with sodium chloride). The samples were then rinsed in the same buffer and postfixed for 30 minutes at 20C in 1% osmium tetroxide (Acros; Geel, Belgium) in the same buffer (0.1 M, pH 7.8, 1030 mOsM). In the CF2 protocol, the samples were fixed for 3 hr at 20C in 0.75% glutaraldehyde in cacodylate buffer (0.1 M, pH 7.8), adjusted to 1030 mOsM with sodium chloride. They were then rinsed in the same buffer and postfixed for 30 min at 20C in 0.1% osmium tetroxide in the same buffer (0.1 M, pH 7.8, 1030 mOsM). After postfixation and washing in buffer, all the samples processed by the CF1 and the CF2 protocols were dehydrated in a graded ethanol series and embedded in Spurr's resin (Polysciences).
The test plates were decalcified, to allow sectioning by the double-embedding method of Holland and Grimmer (1981). Briefly, embedded samples were abraded until the test plates were exposed at the surface of the block. By immersion of the abraded blocks in a 10% EDTA solution for 12 hr, the stereom of the exposed skeletal plates was selectively dissolved. The blocks were then embedded a second time, allowing the decalcified stereomic space of the test plates to be filled with Spurr's resin.
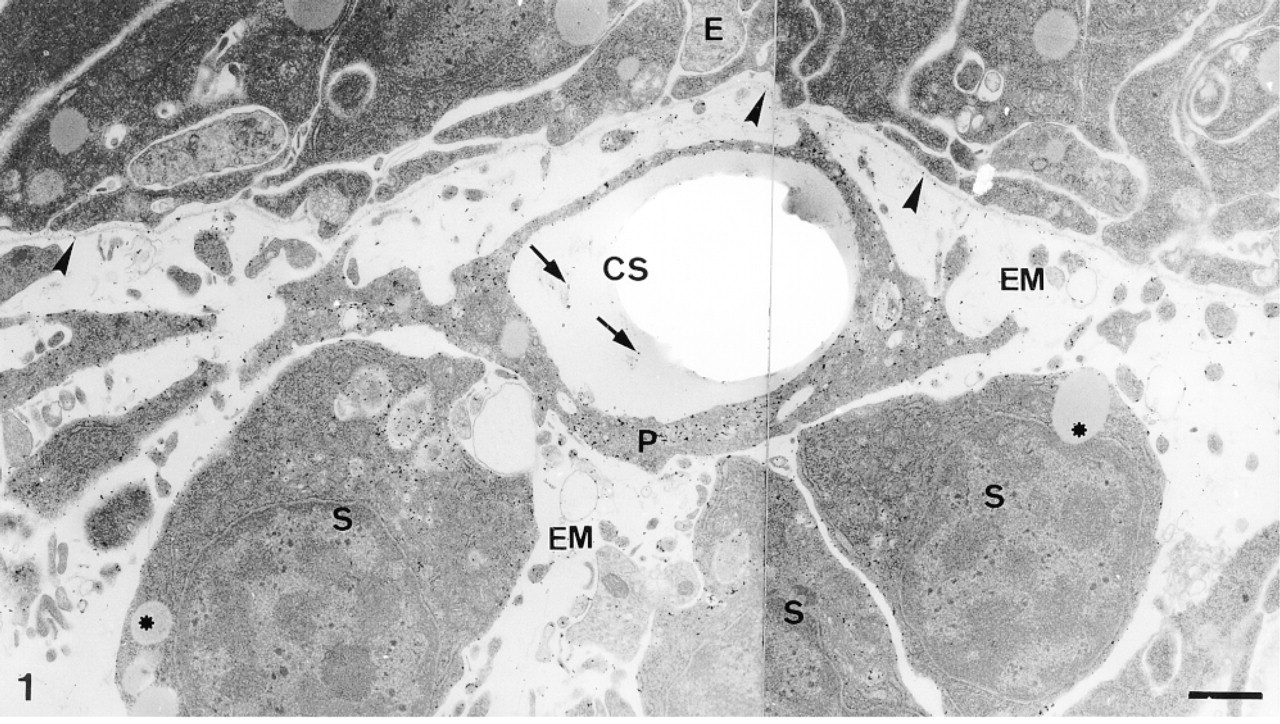
Calcification site (CS) under the epidermis (E). The calcification site (cs) is enclosed in a syncytium pseudopodium (P) generated by sclerocytes (S). Labeling of the organic matrix in the calcification site is indicated by arrows. Note concentration of the labeling in the sclerocytes and their pseudopodia and absence of labeling in the medium gray vesicles (asterisks) of sclerocytes and in the epidermis. Basal lamina (arrowheads) and extracellular matrix (EM) are lightly labeled. Bar = 1 μm.
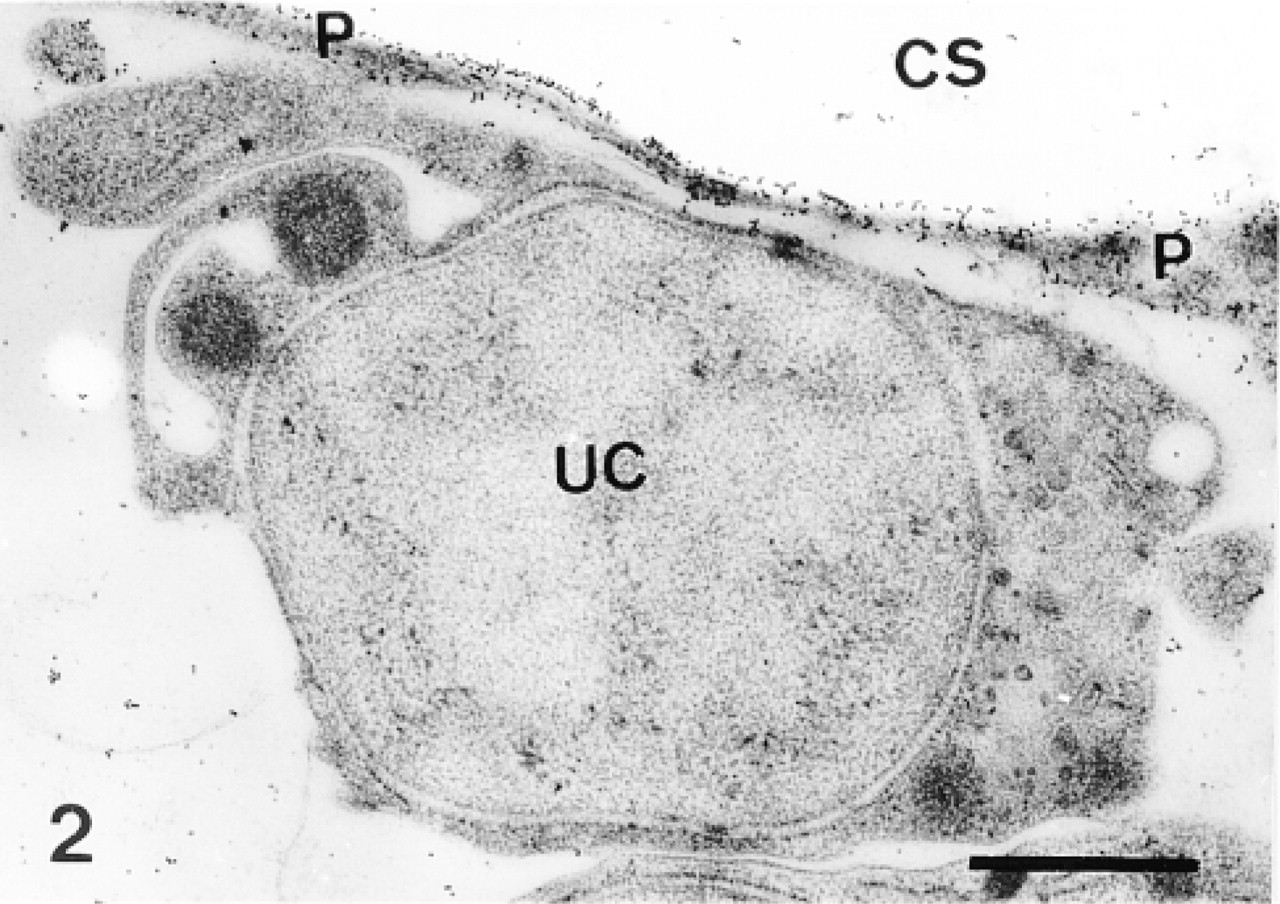
Undifferentiated cell (UC) along a sclerocyte pseudopodium (P) enclosing a calcification site (CS). Labeling is confined to the sclerocyte pseudopodium. Bar = 1 μm.
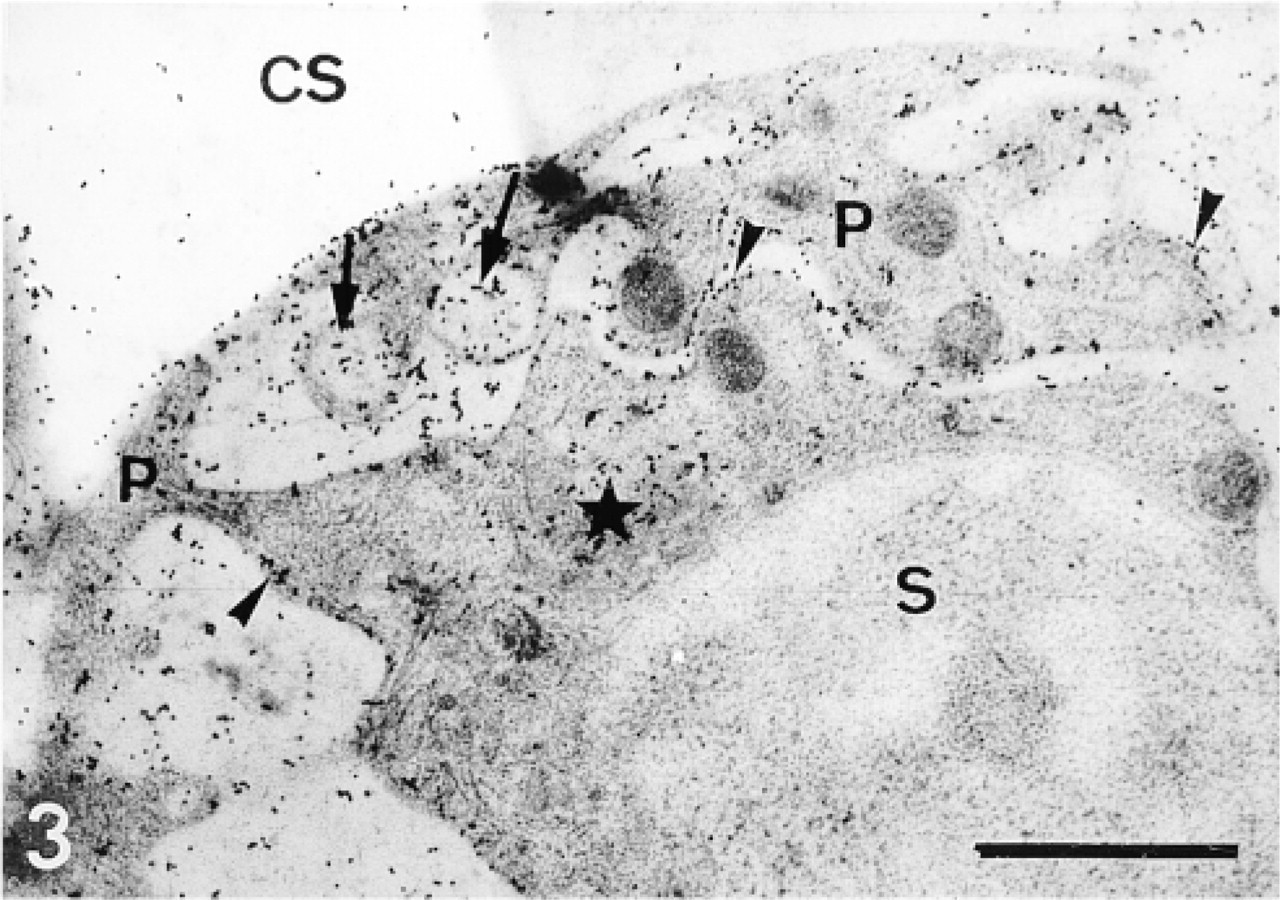
Sclerocyte (S) and its pseudopodium (P). Labeling is confined to the Golgi apparatus (star), the vesicles (arrows), and along the membrane (arrowheads). Note the higher labeling compared with the labeling obtained with the CF2 protocol (Figure 4). CS, calcification site. Bar = 1 μm.
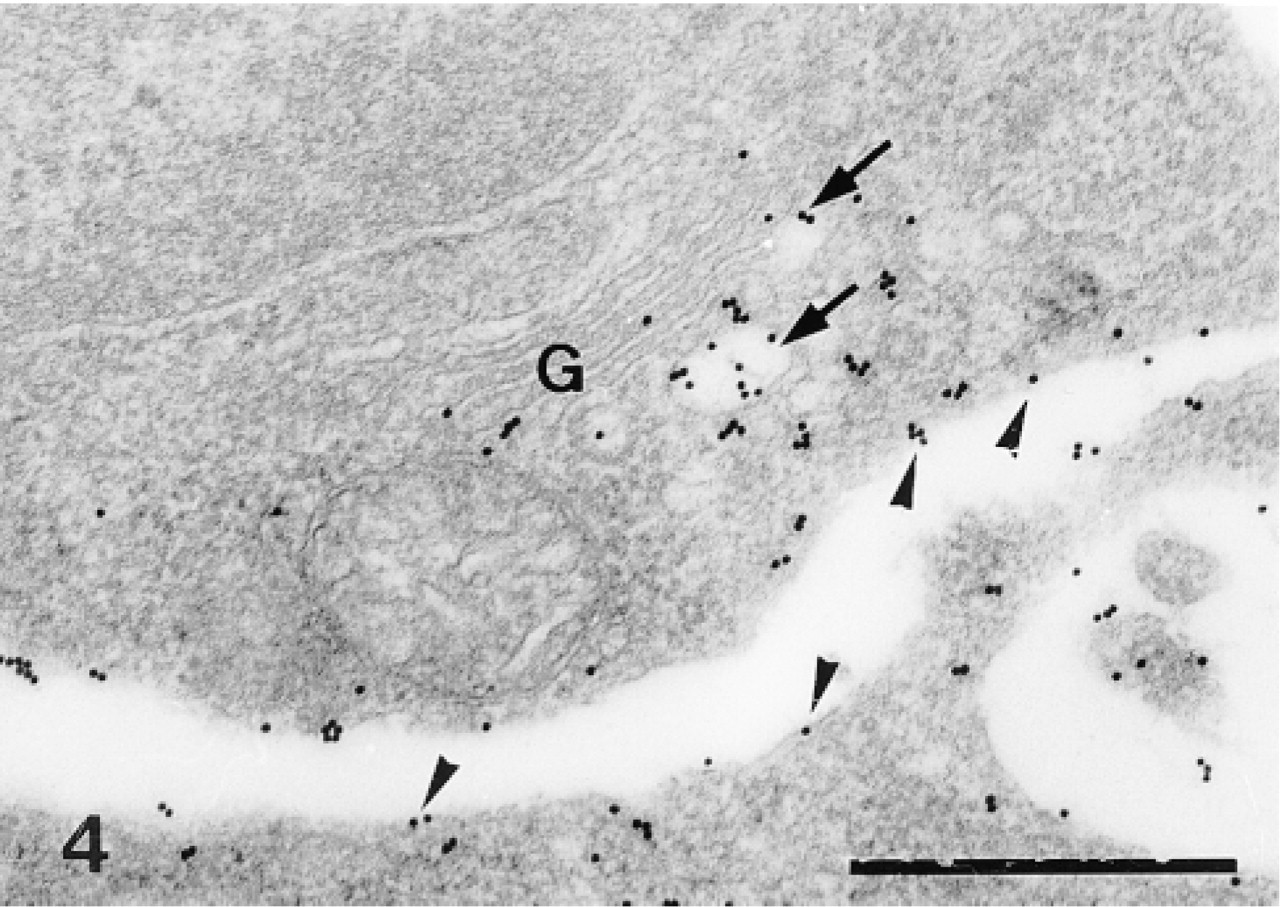
Sclerocyte. Labeling is confined to the trans side of the Golgi apparatus (G), to the Golgi vesicles (arrows), and along the plasma membrane of the sclerocyte (arrowheads). Bar = 500 nm.
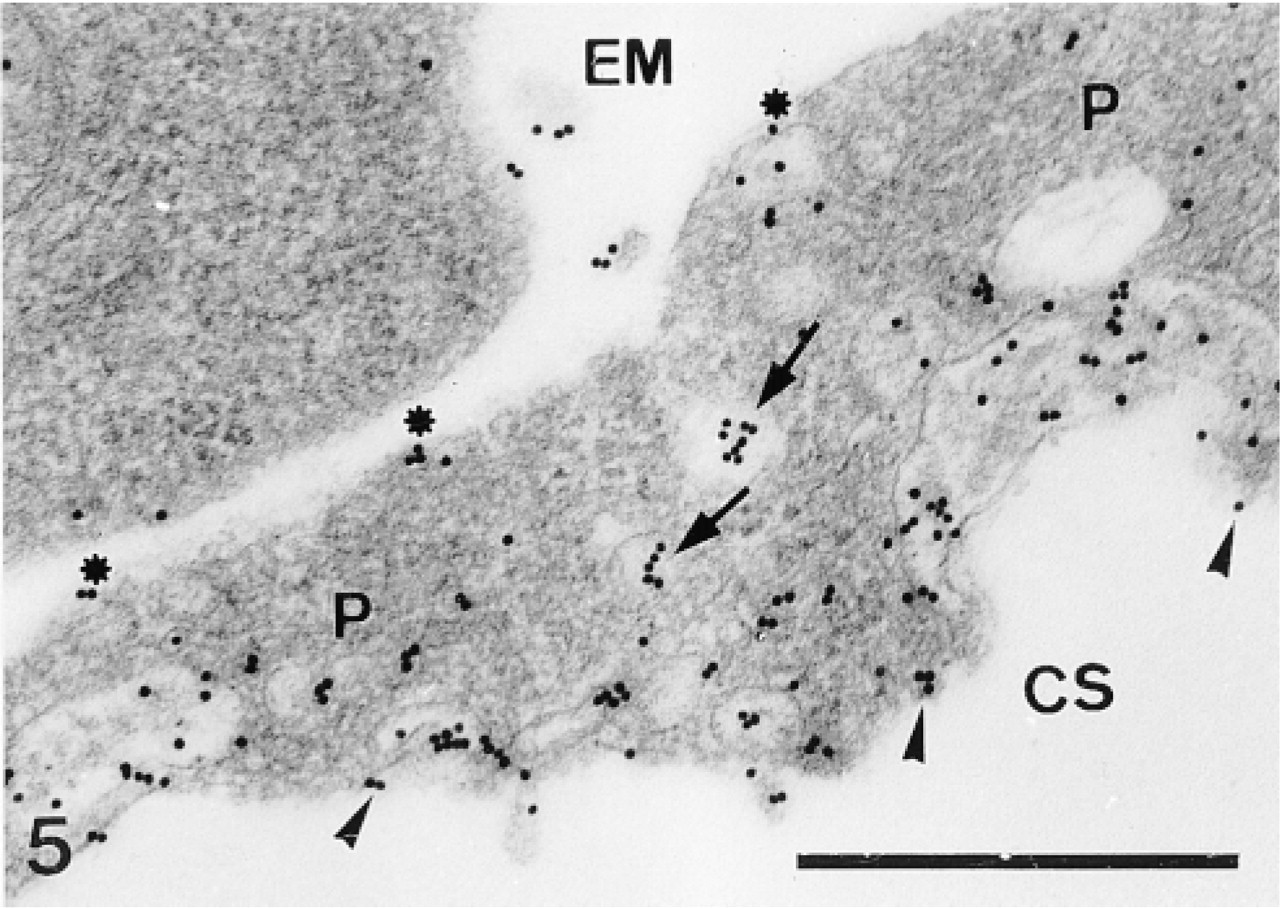
Sclerocyte pseudopodium. Labeling is confined to the vesicles (arrows) and along the inner (arrowheads) and outer pseudopodium (P) membranes (asterisks). CS, calcification site; EM, extracellular matrix. Bar = 500 nm.
Sclerocyte (S) and its pseudopodium (P) enclosing a calcification site (CS). Anti-SM30 labeling is confined to the translucent vesicles (arrowheads). Some gold particles also labeled the organic matrix (arrows). Bar = 500 nm.
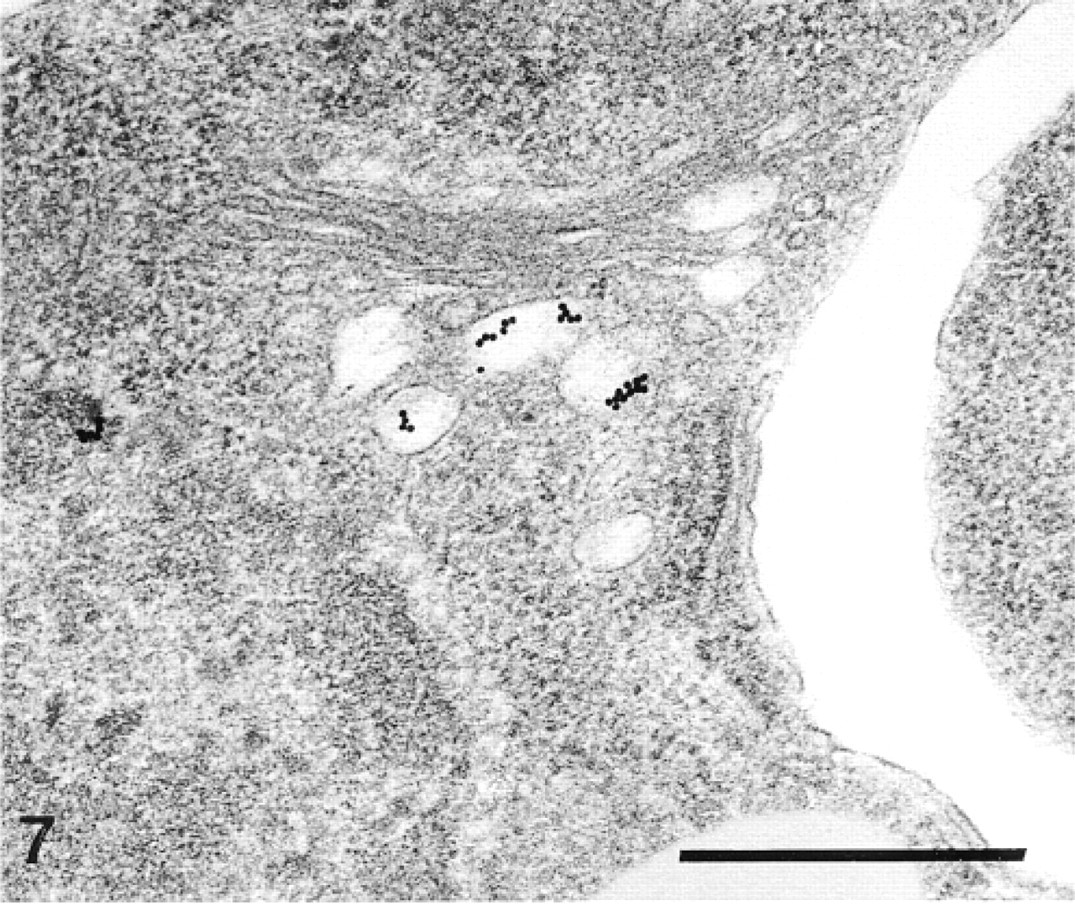
Anti-SM30-labeled Golgi vesicles in a sclerocyte. Bar = 500 nm.
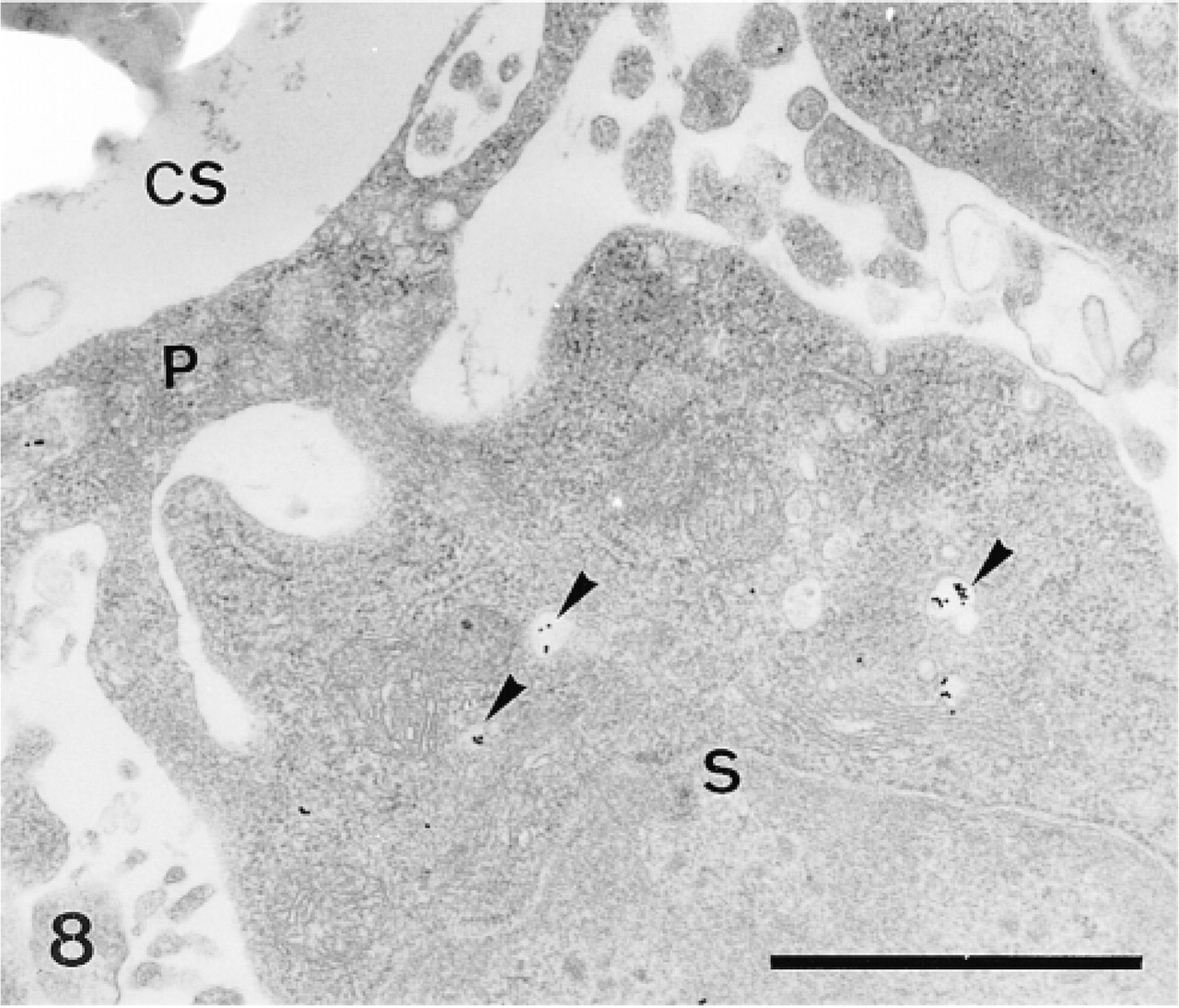
Sclerocyte (S) and its pseudopodium (P) enclosing a calcification site (CS). Anti-SM50 labeling is confined to the translucent vesicles (arrowheads). Bar = 1 μm.
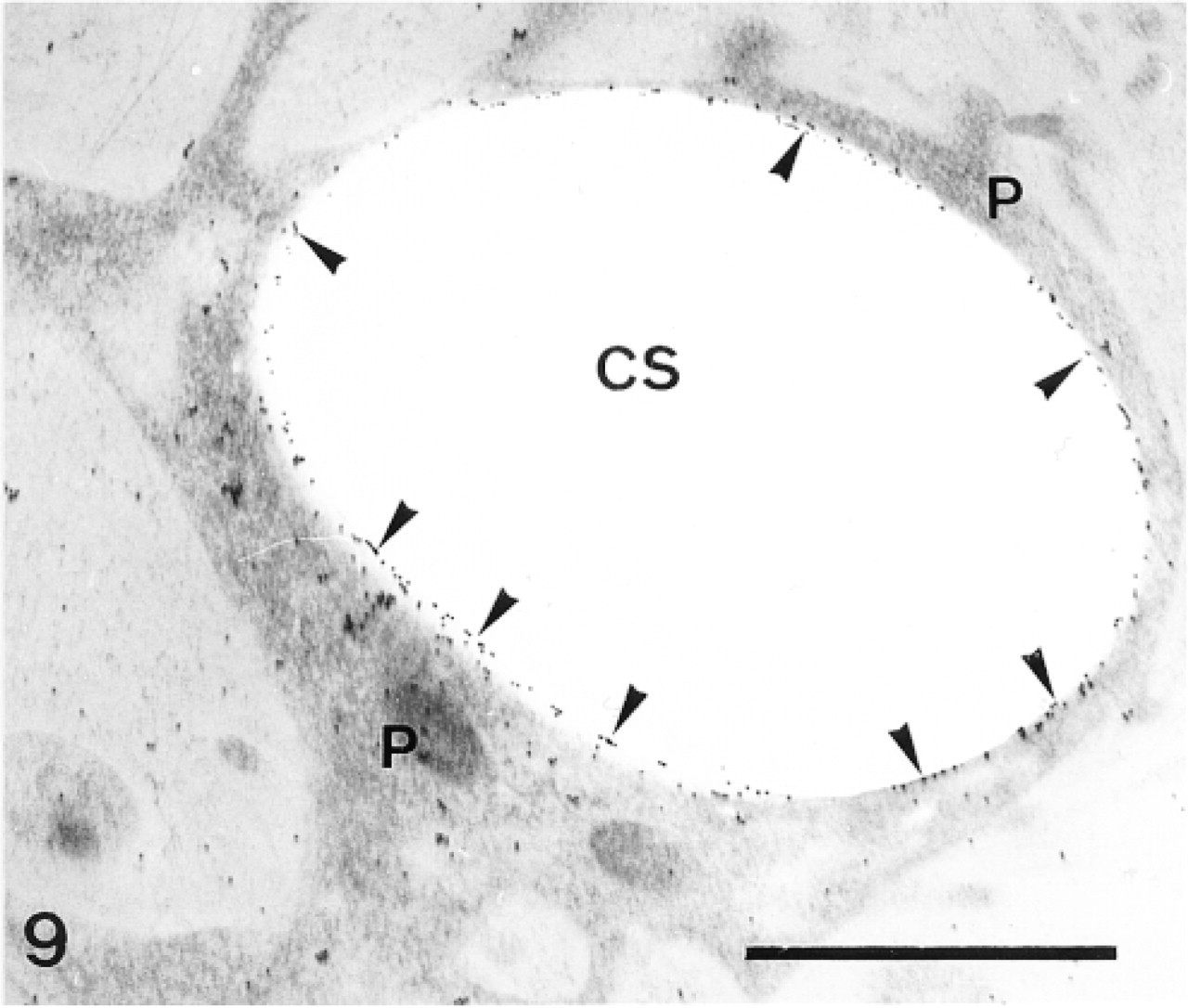
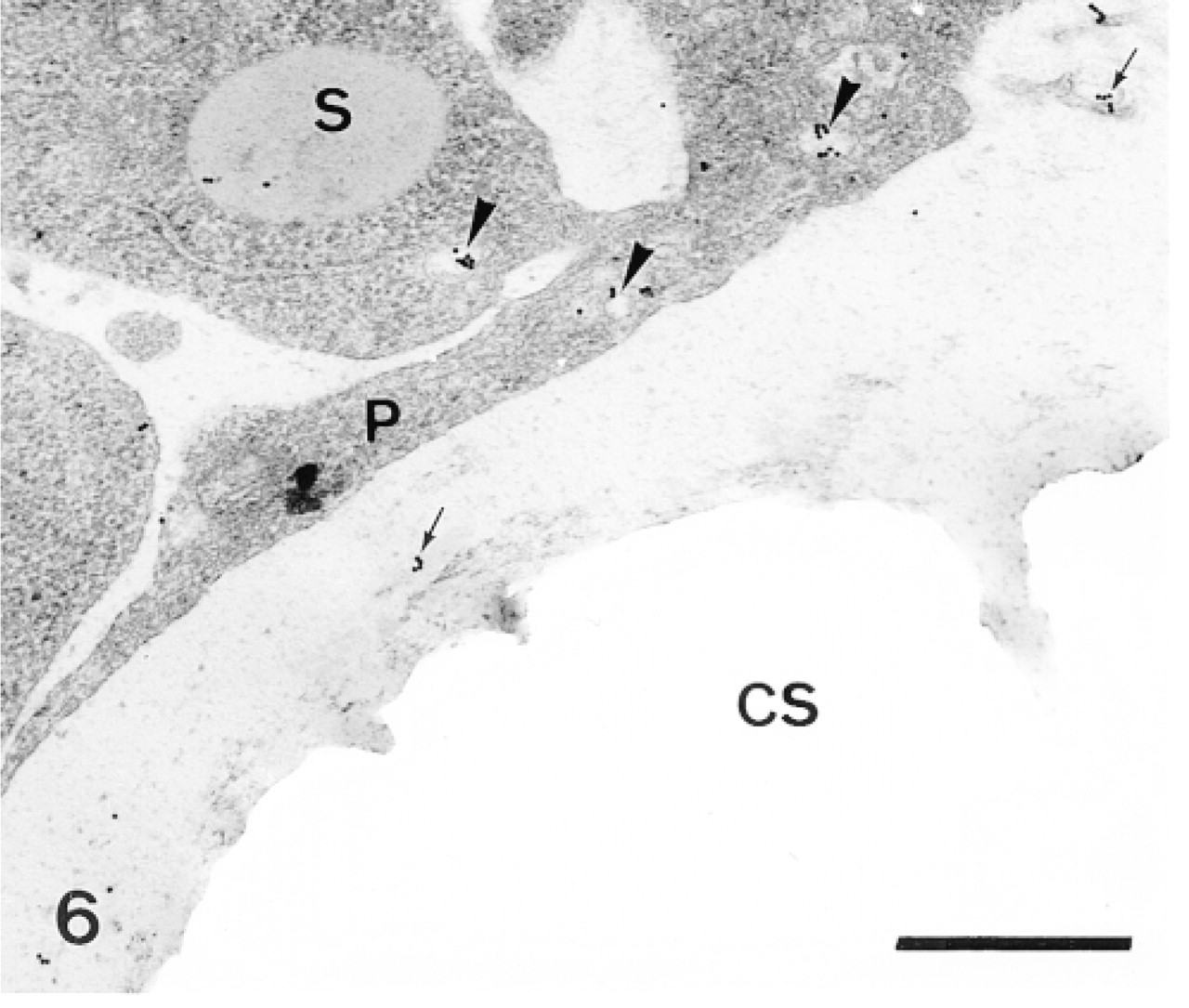
Sclerocyte pseudopodium (P) enclosing a calcification site (CS). Anti-SM50 labeling is confined to the cytoplasm of the pseudopodium (P) and along the perforation within the calcification site (arrowheads). Bar = 1 μm.
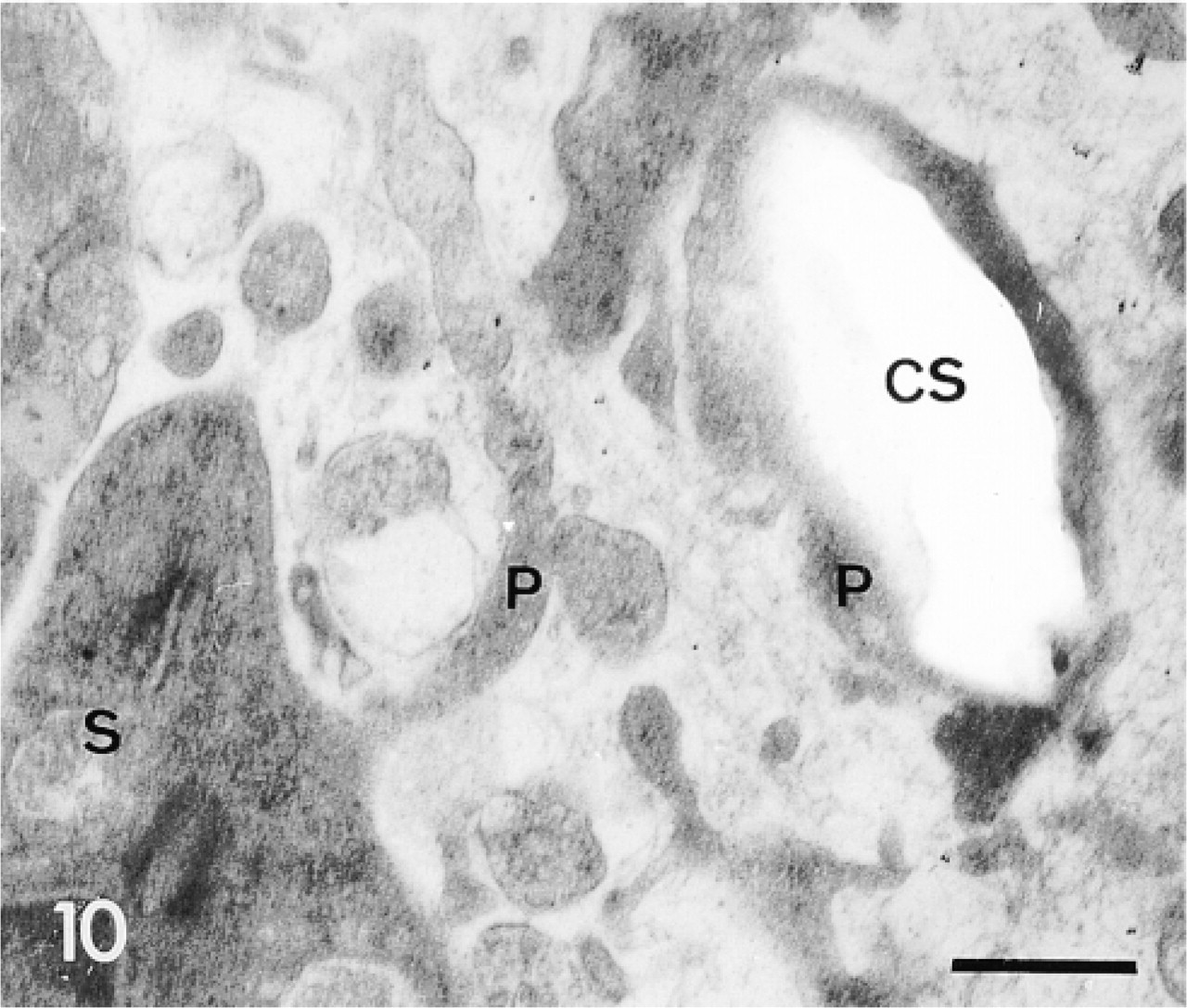
Control experiment: sclerocyte and its pseudopodium surrounding a calcification site (CS). Note absence of secondary antibody labeling in the sclerocyte (S) and in its pseudopodium (P). Bar = 1 μm.
Control experiment: epidermal basal lamina of the pedicellaria primordium. Note secondary antibody labeling of the basal lamina (arrowheads) and of the dermal extracellular matrix (arrows). E, epidermis; M, mesoderm. Bar = 1 μm.
High-pressure Freezing (HPF)
HPF was used on the pedicellaria primordia and on the plumulae of the teeth. The samples were sandwiched between two aluminum plates and cryoimmobilized with a high-pressure freezer (HPM 010; Balzers Union, Balzers, Liechtenstein) (Moor 1987). After immobilization they were immediately transferred to liquid nitrogen, in which they were stored.
Freeze-Substitution (FS)
The frozen sandwiches were opened under liquid nitrogen and the samples (still attached to one of the plates) were freeze-substituted using a freeze-substitution unit (Balzers FSU 010). Two protocols were tested. The first one (HPF/FS1) attempted to preserve the spatial organization of the organic matrix, whereas the second (HPF/FS2) attempted to better preserve the antigenicity.
In the HPF/FS1 protocol, the samples were freeze-substituted in anhydrous acetone with 0.2% gallic acid (low molecular weight tannic acid; C7H6O5; Fluka, Buchs, Switzerland) and 6% glutaraldehyde (Polysciences) for 48 hr at 183K and 12 hr at 213K, washed three times at 213K with anhydrous acetone, transferred at 213K to anhydrous acetone with 2% osmium tetroxide (Johnson Matthey; Herts, UK), and kept for 30 hr at 213K, 8 hr at 243K, and 1 hr at 273K. The samples were then washed three times in anhydrous acetone and embedded stepwise in Epon/Araldite (30%, 70%, 100%) (Fluka). The infiltration times were 3 hr at each resin concentration. The final polymerization was completed at 333K.
In the HPF/FS2 protocol, the samples were freeze-substituted in anhydrous ethanol with 0.5% uranyl acetate (Sigma; St Louis, MO) at 183K, 213K, and 243K for 8 hr at each step (Müller et al. 1980) and for 1 hr at 273K. The samples were then washed three times in anhydrous acetone and embedded stepwise in LR Gold (33%, 66%, 100% resin) (Fluka). The infiltration times were respectively 3 hr, 12 hr and 2 hr. The polymerization was carried out at 253K.
Microtomy, Immunolabeling, and Electron Microscopy
Ultrathin sections were cut on an Ultracut UCT Leica with diamond knives (Diatome; Biel, Switzerland), picked up on gold grids, and immunolabeled. Three primary sera (anti-total matrix, anti-SM30, and anti-SM50) raised in rabbit against larval organic matrix components of the echinoid Strongylocentrotus purpuratus were used. The characterization of the antitotal matrix serum was reported in Benson et al. (1986); the anti-SM30 and the anti-SM50 sera characterizations were reported in Killian and Wilt (1996).
Anti-total matrix, anti-SM30, and anti-SM50 single immunolabeling was applied to sections placed on Formvar-coated or uncoated grids. To block unspecific staining, the sections were immersed in drops of 1 M NH4Cl in PBS buffer (0.01 M, 0.15 M NaCl) for 1 hr. They were then rinsed three times with PBS and incubated in primary antibodies diluted 1:500 for 1 hr at 20C. After six rinses with PBS, goat anti-rabbit IgG conjugated to 10- or 15-nm gold particles (Biocell; Cardiff, UK) diluted 1:100 with PBS (0.01 M, NaCl 0.15 M) plus 0.1% Tween-20 (Sigma) and 1% BSA (Sigma) was applied to the grids for 1 hr at 20C. The sections were then rinsed six times with PBS and three times with Milli-Q (Millipore; Bedford, MA) water.
Immunolabeling omitting the primary antibody was used as control. All the sections were stained with uranyl acetate and lead citrate and examined in a Philips EM 300 or a JEOL 2000 FX II transmission electron microscope.
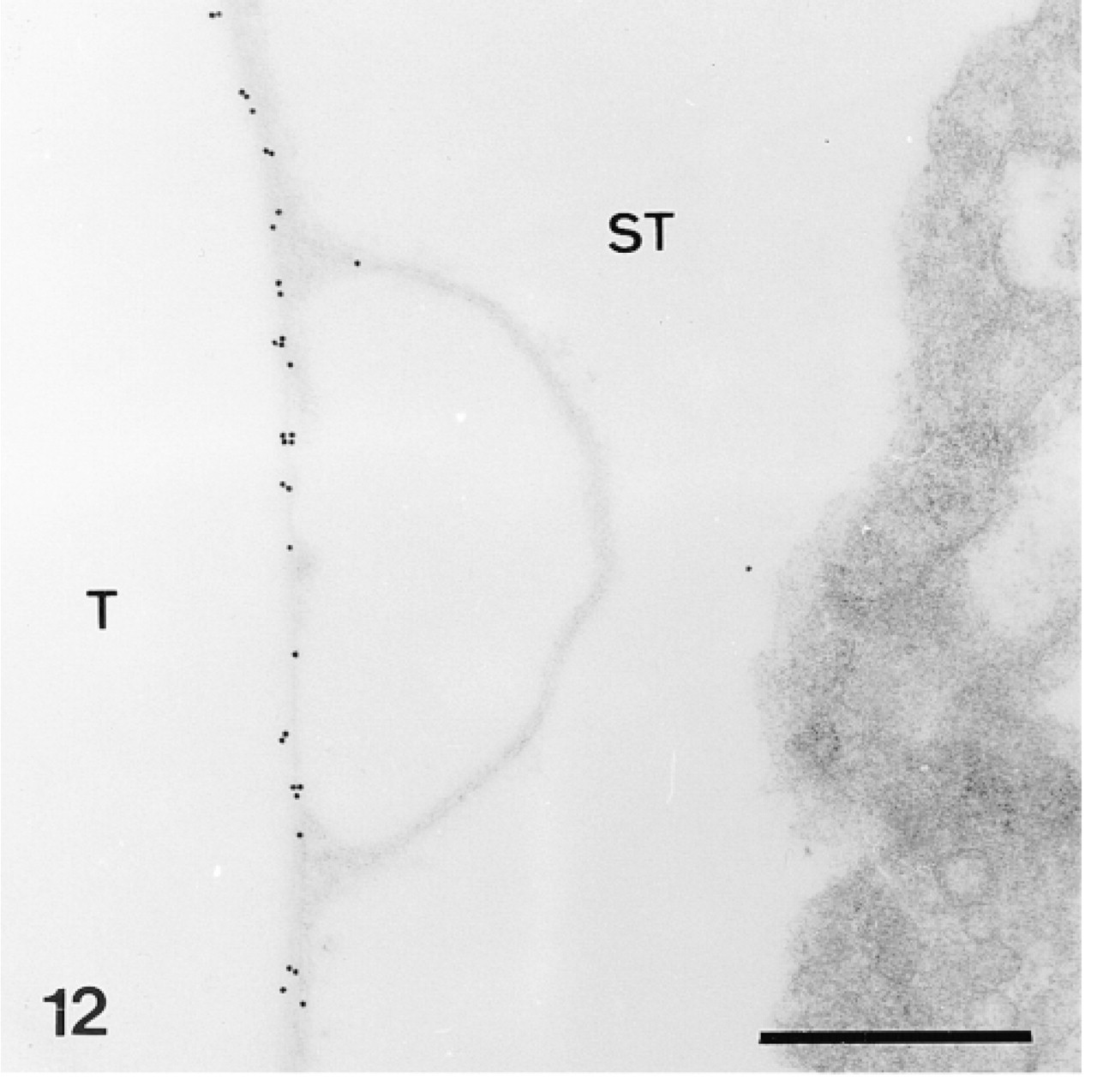
Trabecular coat labeled by anti-total matrix serum in a mature skeletal plate. ST, stroma; T, trabecula. Bar = 500 nm.
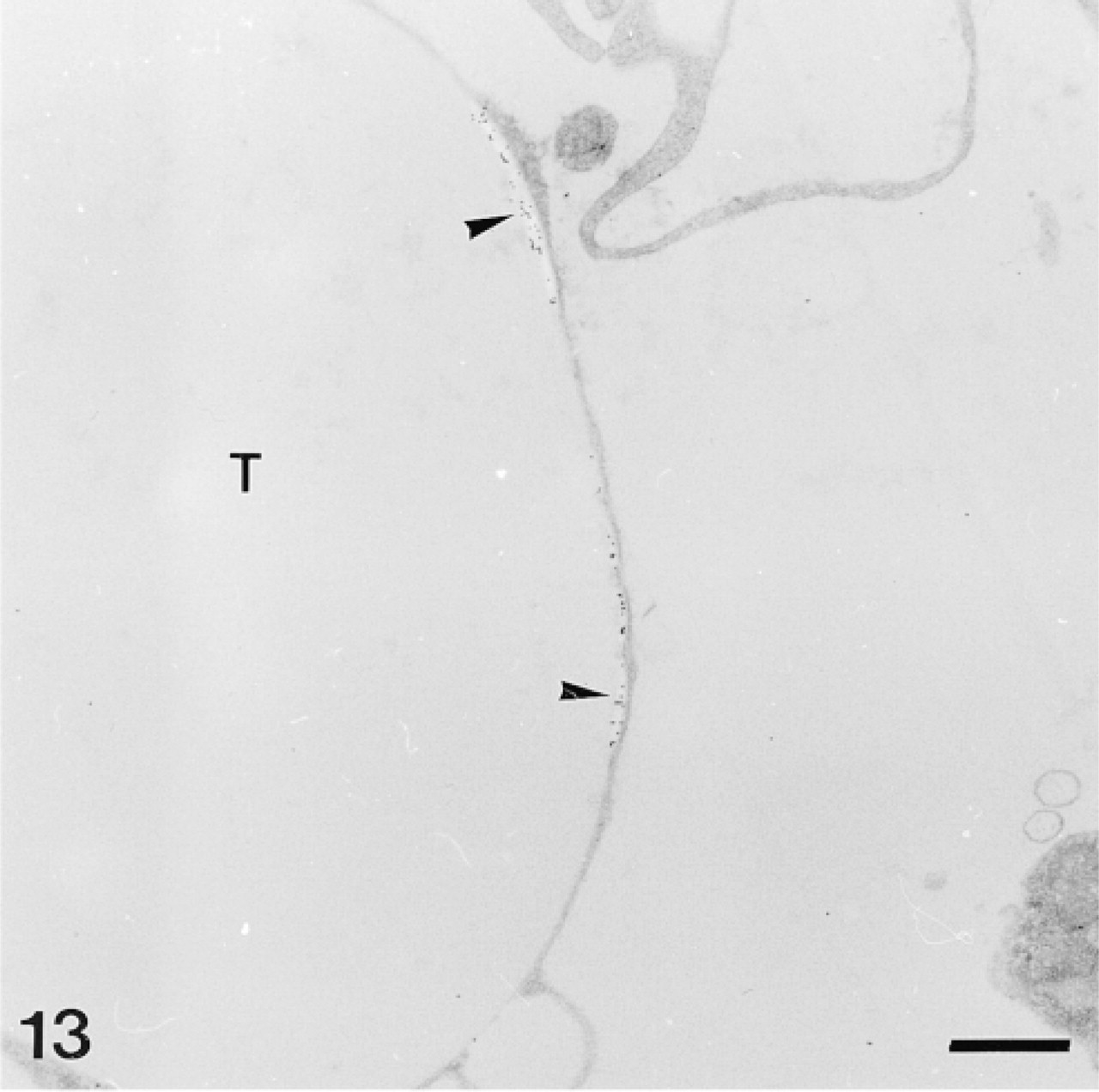
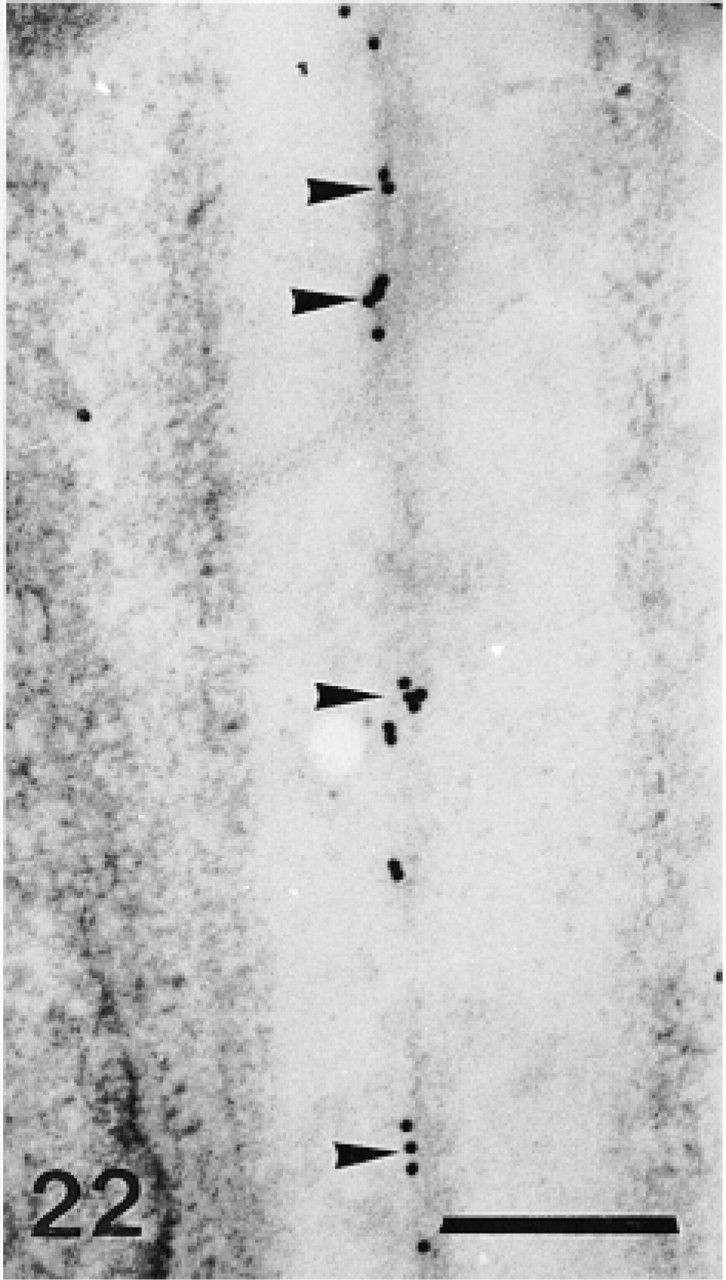
Anti-SM30-labeled trabecular coat in a mature skeletal plate. Note aggregative distribution of the anti-SM30 labeling and the stereomic electron-lucent areas lining this labeling (arrowheads). T, trabecula. Bar = 1 μm.
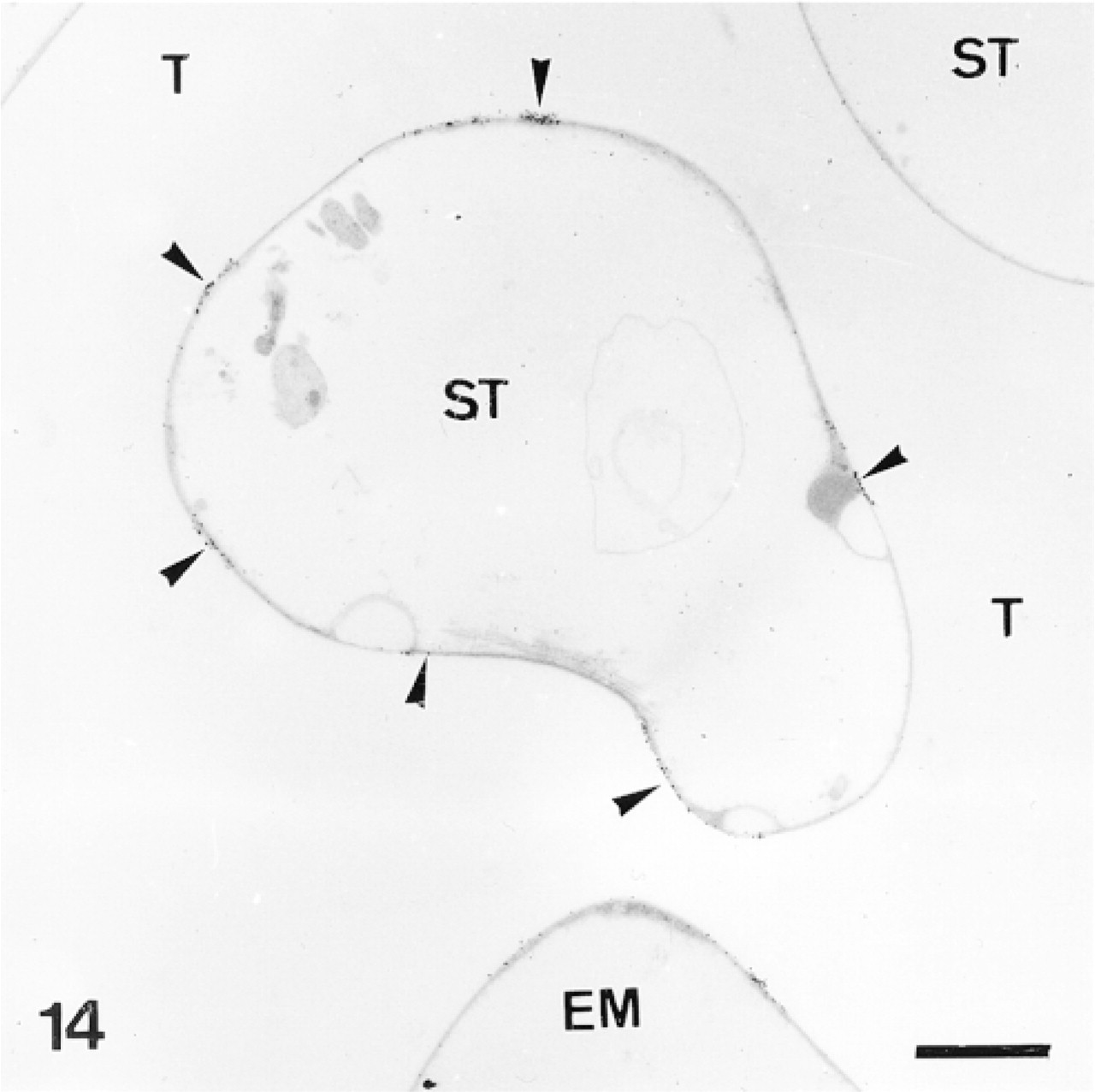
Anti-SM50-labeled trabecular coat in a mature skeletal plate. Network of trabeculae (T) delimits a complementary network filled by connective tissue: the stroma (ST). Note aggregative distribution of anti-SM50 labeling (arrowheads). EM, extracellular matrix. Bar = 2 μm.
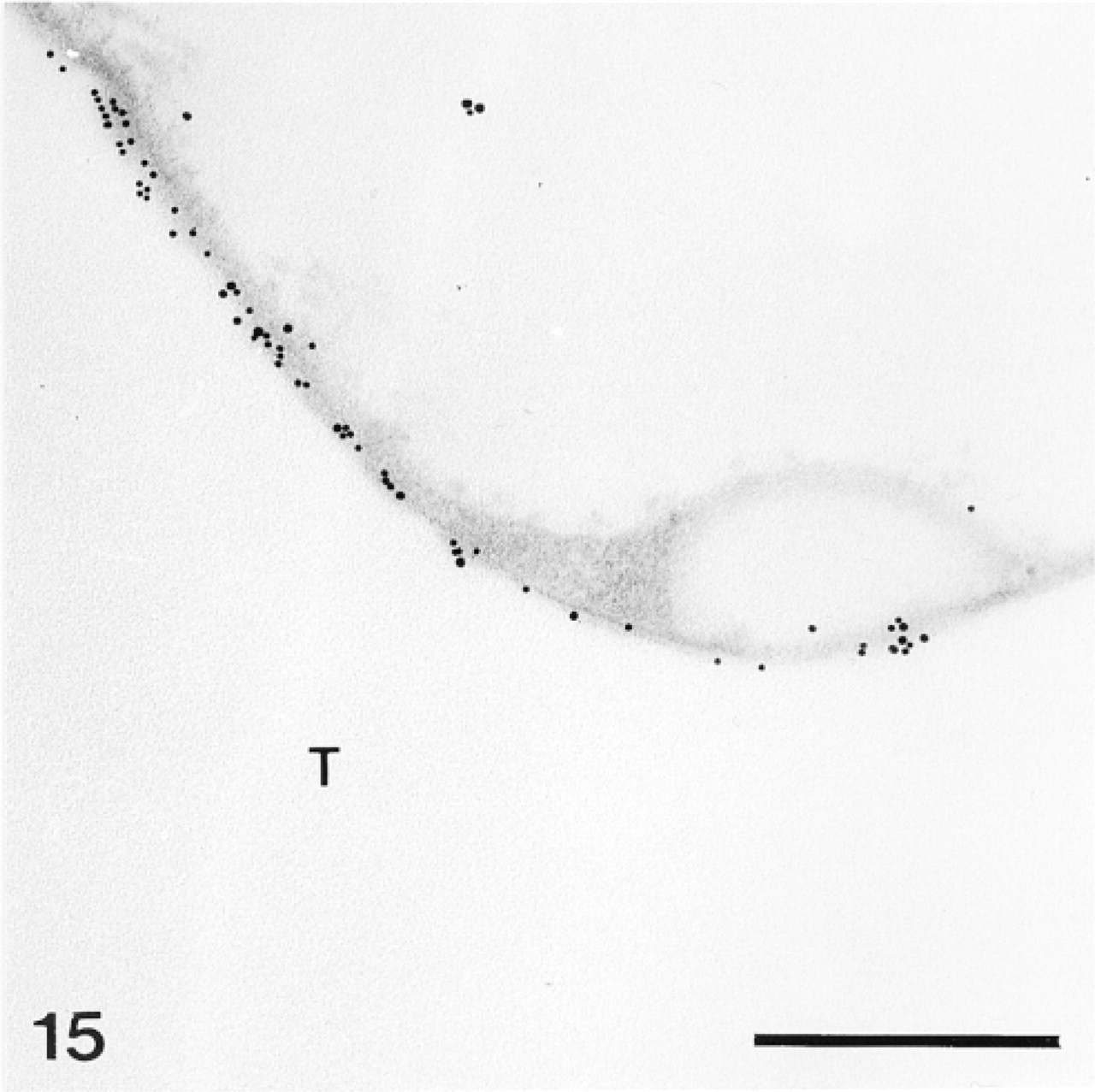
Magnified area of Figure 14. Anti-SM50-labeled area of the trabecular coat. T, trabecula. Bar = 500 nm.
Results
Preservation of Antigenicity According to the Fixation Protocol
Three primary sera (anti-total matrix, anti-SM30, and anti-SM50) raised against larval organic matrix components of the echinoid S. purpuratus were used on pedicellaria primordia, test plates, and teeth of the echinoid P. lividus. The anti-total matrix serum is known to react with at least eight components of the larval organic matrix, whereas the anti-SM30 and anti-SM50 sera are specific to, respectively, the SM30 and SM50 proteins (Benson et al. 1986; Killian and Wilt 1996). Two conventional fixation protocols (CF1 with tannic acid and CF2 without tannic acid) and two high-pressure freezing/freeze-substitution protocols (HPF/FS1 with gallic acid and HPF/FS2 without gallic acid) were tested to determine the best method for preserving antigenicity. No labeling was obtained with the CF1 protocol (not shown). With the HPF/FS1 protocol, the anti-total matrix labeling was rather weak (Figure 18) and the anti-SM30 and anti-SM50 labeling was not applied on sections of tissues fixed with this protocol. With the CF2 and HPF/FS2 protocols, the same structures were labeled. However, the HPF/FS2 labeling was much stronger although the poorer contrast of membranes (particularly of the membranes of the Golgi stacks and vesicles) made the HPF/FS2 labeling less precise than the CF2 labeling (compare, e.g., Figures 6 and 9).
Immunolabeling of the Pedicellaria Primordia
The pedicellariae are small appendages consisting of three movable jaws on a stalk. They form on the test plates within primordia, which are made up of a cluster of tightly packed dermal cells covered by the epidermis (Burke 1980; Ameye et al. 1998). The pedicellaria primordia contain two main types of dermal cells: undifferentiated cells and sclerocytes. The sclerocytes extend long syncytial pseudopodia which delimit the calcification sites (Figure 1) (see Ameye et al. 1998). These contain structured organic material (the so-called mineralization organic matrix) and sometimes show perforations corresponding to newly deposited mineral that was displaced during sectioning.
In the primordia, the anti-total matrix serum labeled the sclerocytes, the sclerocyte pseudopodia, and the organic matrix of the calcification sites (Figure 1). The epidermis (Figure 1) and the undifferentiated cells of the primordia (Figure 2) were not labeled. In the sclerocytes, the labeling was observed on the trans side of the Golgi stacks, in the electron-translucent Golgi vesicles, and along the plasma membranes (Figures 3 and 4) but not in the medium gray vesicles (Figure 1). In the sclerocyte pseudopodia, the labeling was localized along the inner and outer pseudopodium membrane and in the vesicles (Figure 5). The anti-SM30 and anti-SM50 labeling was weaker than the antitotal matrix labeling (compare Figures 6 and 8 with Figure 1). They presented the same localization and were observed in the translucent Golgi vesicles of the sclerocyte cell bodies (Figures 6–8), in the translucent vesicles of the sclerocyte pseudopodia (Figure 6), on the organic matrix of the calcification sites, and along the edges of the perforations within the calcification sites (Figure 9). The latter labeling did not result from a nonspecific effect due to the perforation. Indeed, no labeling was observed along the accidental perforations localized outside calcification sites. With each of the three sera separately, the basal lamina and the extracellular organic matrix of the primordia were lightly labeled (Figure 1).
Immunolabeling of the Test Plates
The test plates comprise a tridimensional mineral network of trabeculae (the stereom) which delimits an internal and complementary network filled by a connective tissue (the stroma, Figure 14) containing some sparse sclerocytes. The trabeculae are surrounded by an outer organic matrix layer; the trabecular coat. In our sections, no sclerocyte cell body was observed and the trabecular coat was the only structure to be labeled (Figures 12–15). It was labeled uniformly by the anti-total matrix serum and by patches with the anti-SM30 and anti-SM50 sera (Figures 13 and 14). For the anti-SM30 labeling, the labeled patches were often bordered by electron-lucent areas of the stereom (Figure 13).
Immunolabeling of the Teeth
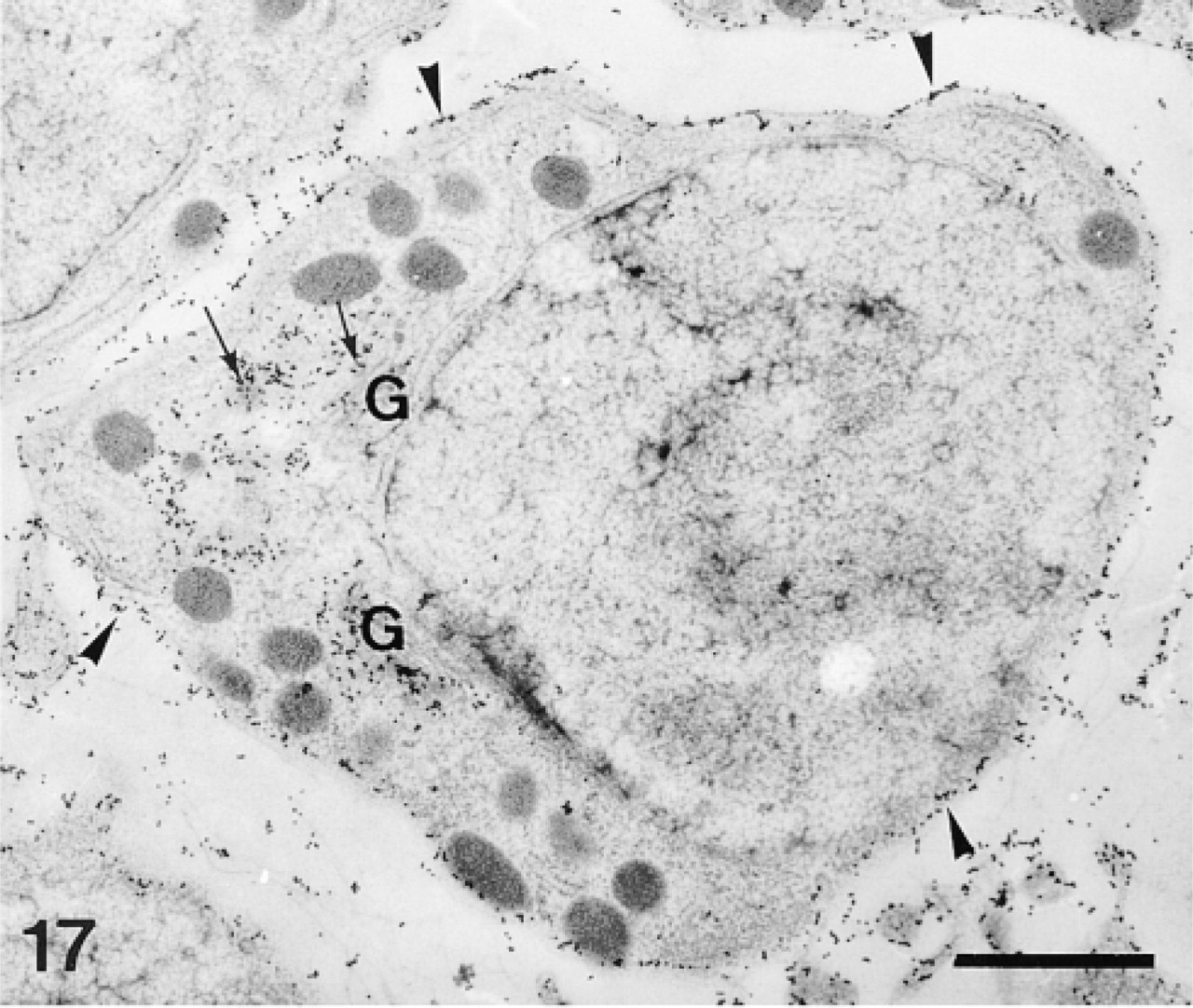
The plumula of the tooth contains two main cell types, the odontoblasts and the preodontoblasts (Märkel et al. 1986). The odontoblasts are organized in parallel rows, which are separated from each other by calcified primary plates. The odontoblasts of each row extend one long syncytial pseudopodium which delimits one calcification site, in which one primary plate develops. The odontoblasts are characterized by a thin layer of cytoplasm devoid of Golgi apparatus (Figure 16). The preodontoblasts are located at the extremities of the rows of odontoblasts, ready to be incorporated in the odontoblast syncytia. Unlike the odontoblasts, they have a more developed cytoplasm and are actively involved in the secretion of proteins, as attested to by their many Golgi stacks (Figure 17).
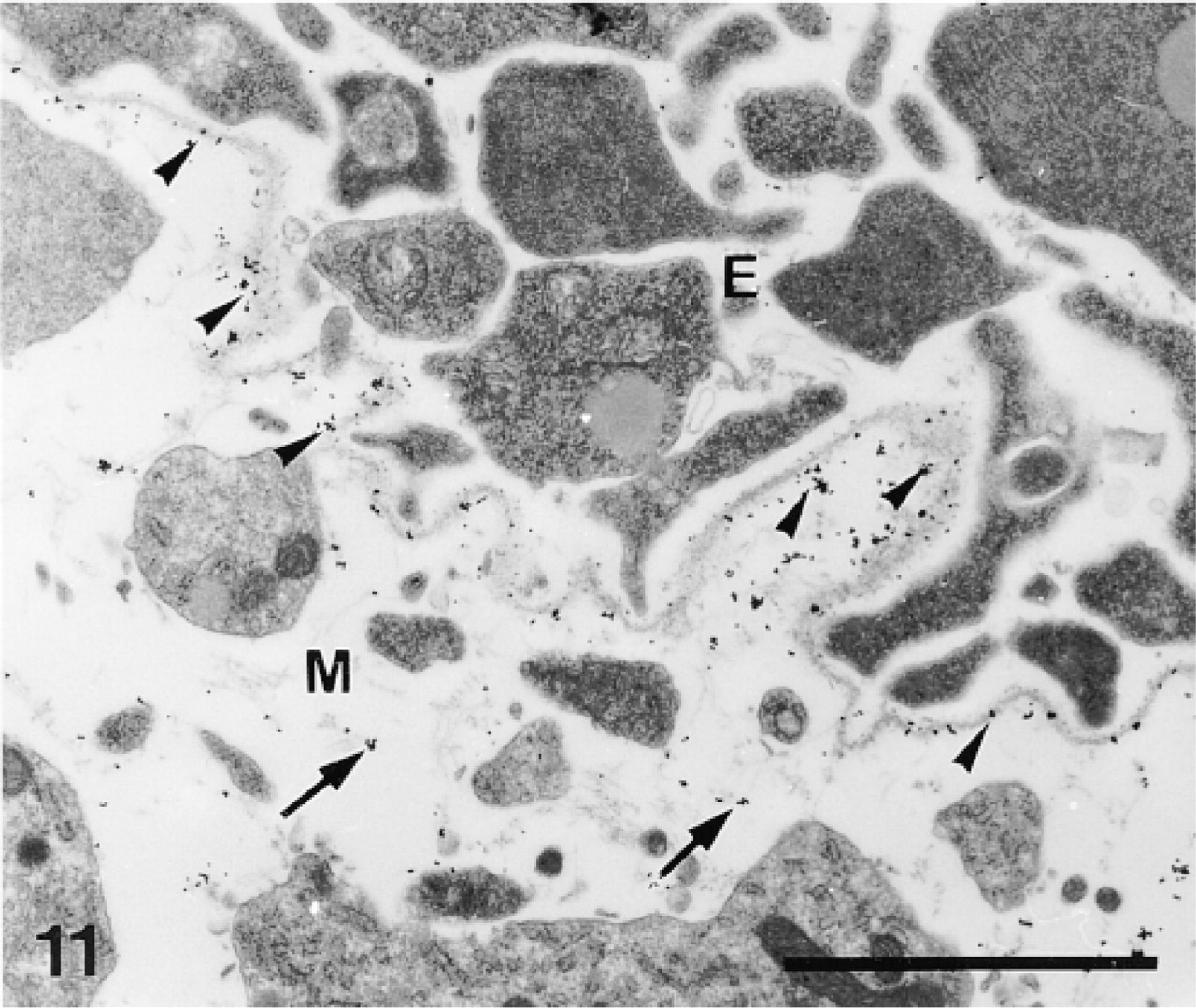
The anti-total matrix serum labeled the odontoblasts and their pseudopodia (Figure 16), the preodontoblasts (Figures 17 and 18), the organic matrix of the primary plates (Figure 19), and the margins of the perforations in the calcification sites (Figure 16). No labeling was observed on the extracellular matrix or along accidental perforations outside of the calcification sites. This indicates that the labeling along the perforations of the calcification sites did not result from a nonspecific effect caused by the perforation. In the odontoblasts and their pseudopodia, labeling was observed on the plasma membranes of the odontoblasts and on the inner and outer membranes and the hyaloplasm of the pseudopodia (Figure 16). In the preodontoblasts, labeling was observed in the Golgi stacks, in the medium electron-dense Golgi vesicles, and along the plasma membranes (Figures 17 and 18).
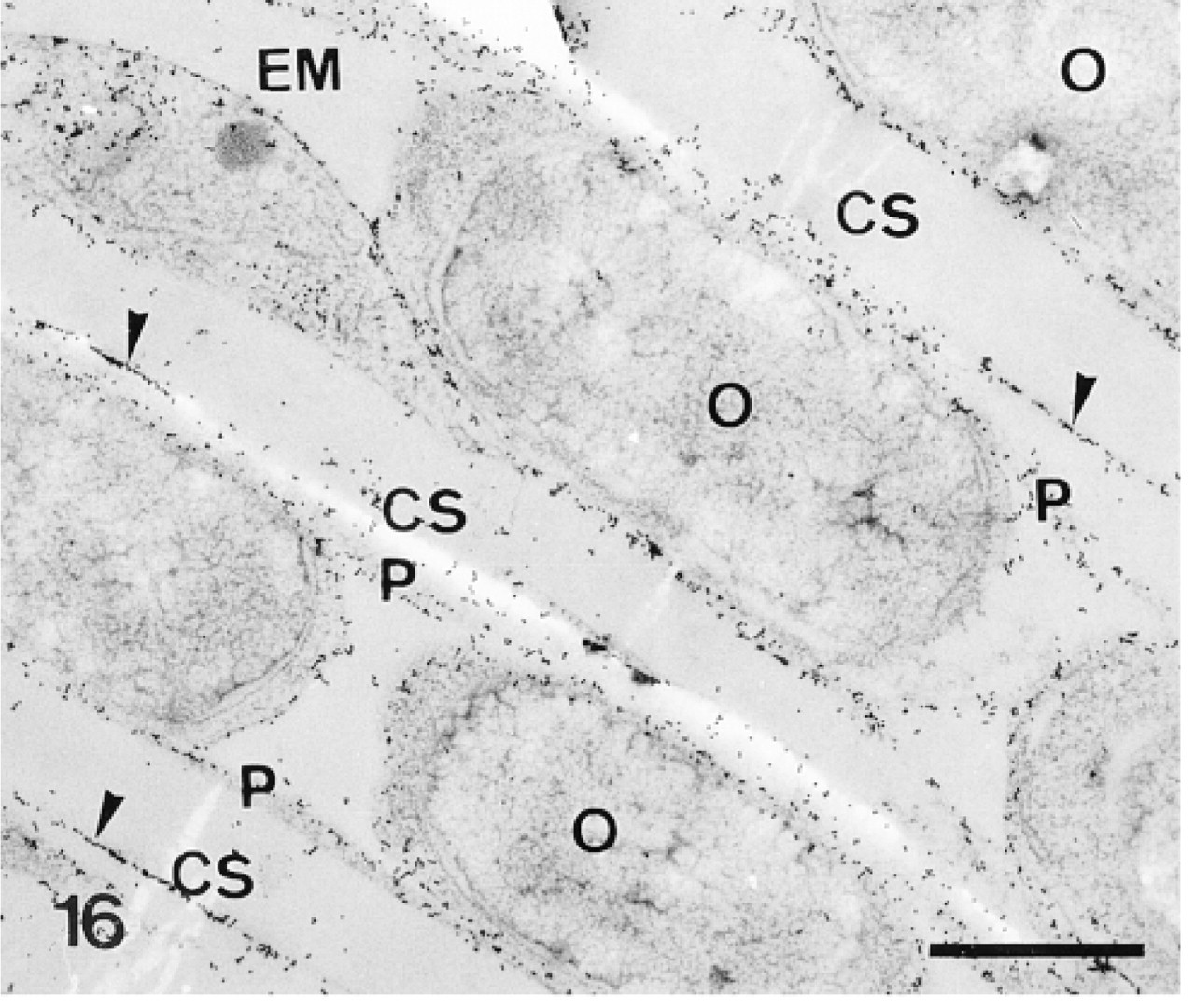
Calcification sites (CS) enclosed in odontoblast pseudopodia (P) and separated from each other by rows of odontoblasts (O). Anti-total matrix labeling is observed on the odontoblasts, the odontoblast pseudopodia, and the organic matrix (arrowheads). Note absence of labeling in the odontoblast nuclei and in the extracellular matrix (EM). Bar = 1 μm.
Preodontoblast. Anti-total matrix labeling is observed on the Golgi stacks (G), the Golgi vesicles (arrows), and along the plasma membrane (arrowheads). Bar = 1 μm.
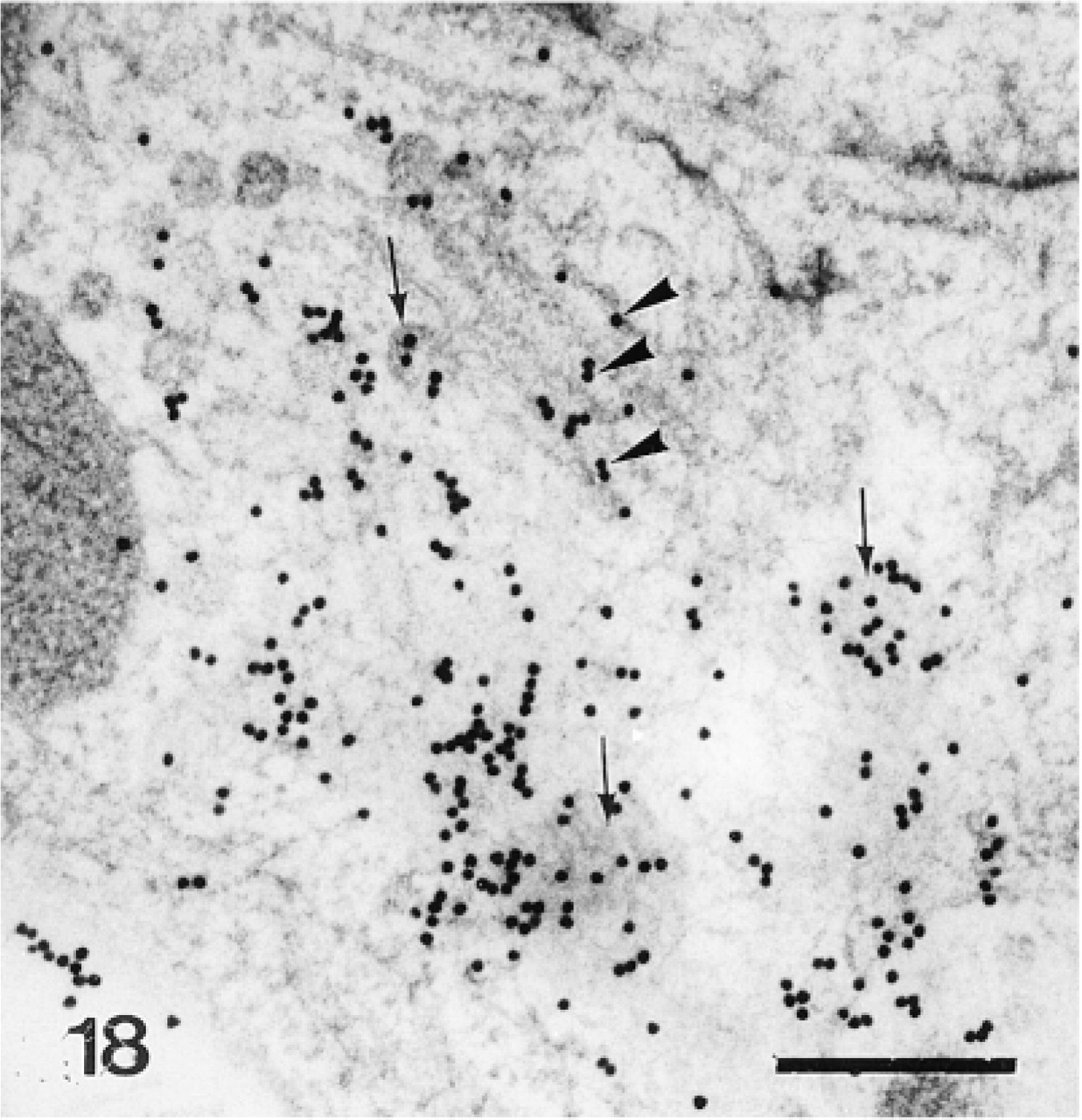
Magnified area of Figure 17. Note anti-total matrix labeling of the Golgi apparatus (arrowhead) and the Golgi vesicles (arrows). Bar = 500 nm.
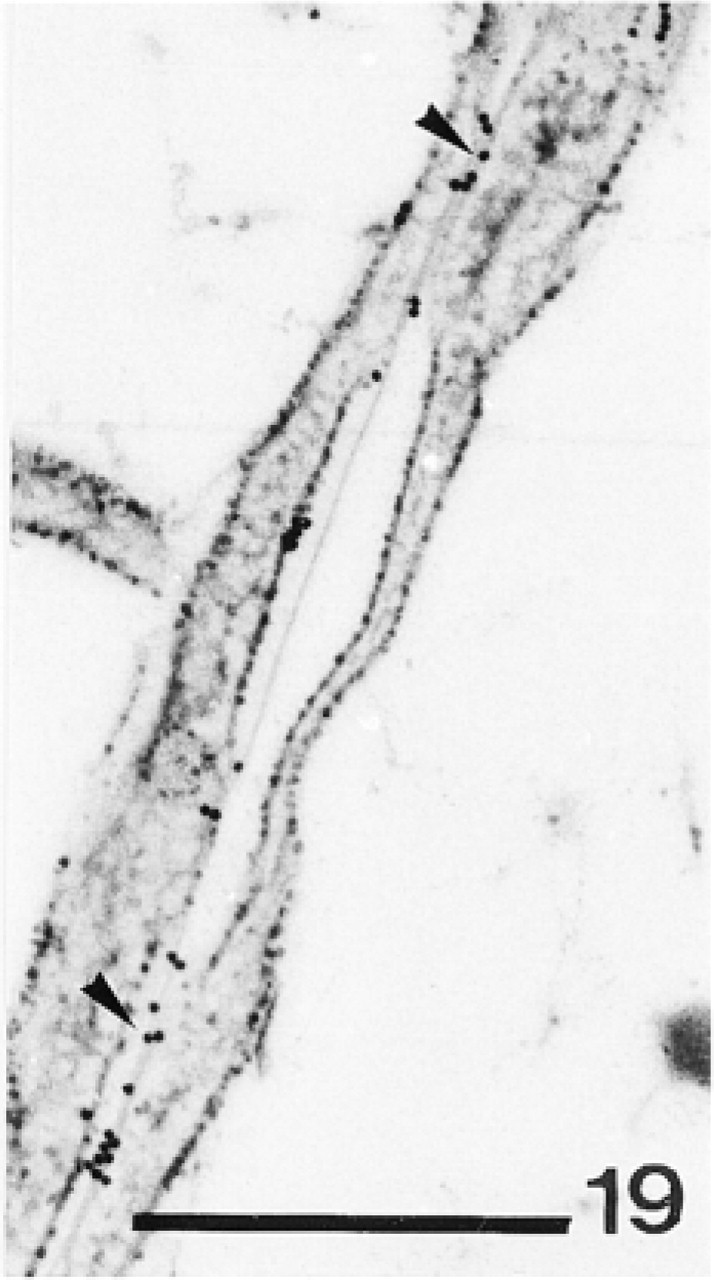
Calcification site. Note anti-total matrix labeling of the organic matrix (arrowheads). Bar = 250 nm.
Control experiment: secondary antibody labeling in the tooth. Note very low background labeling. CS, calcification site; O, odontoblast. Bar = 1 μm.
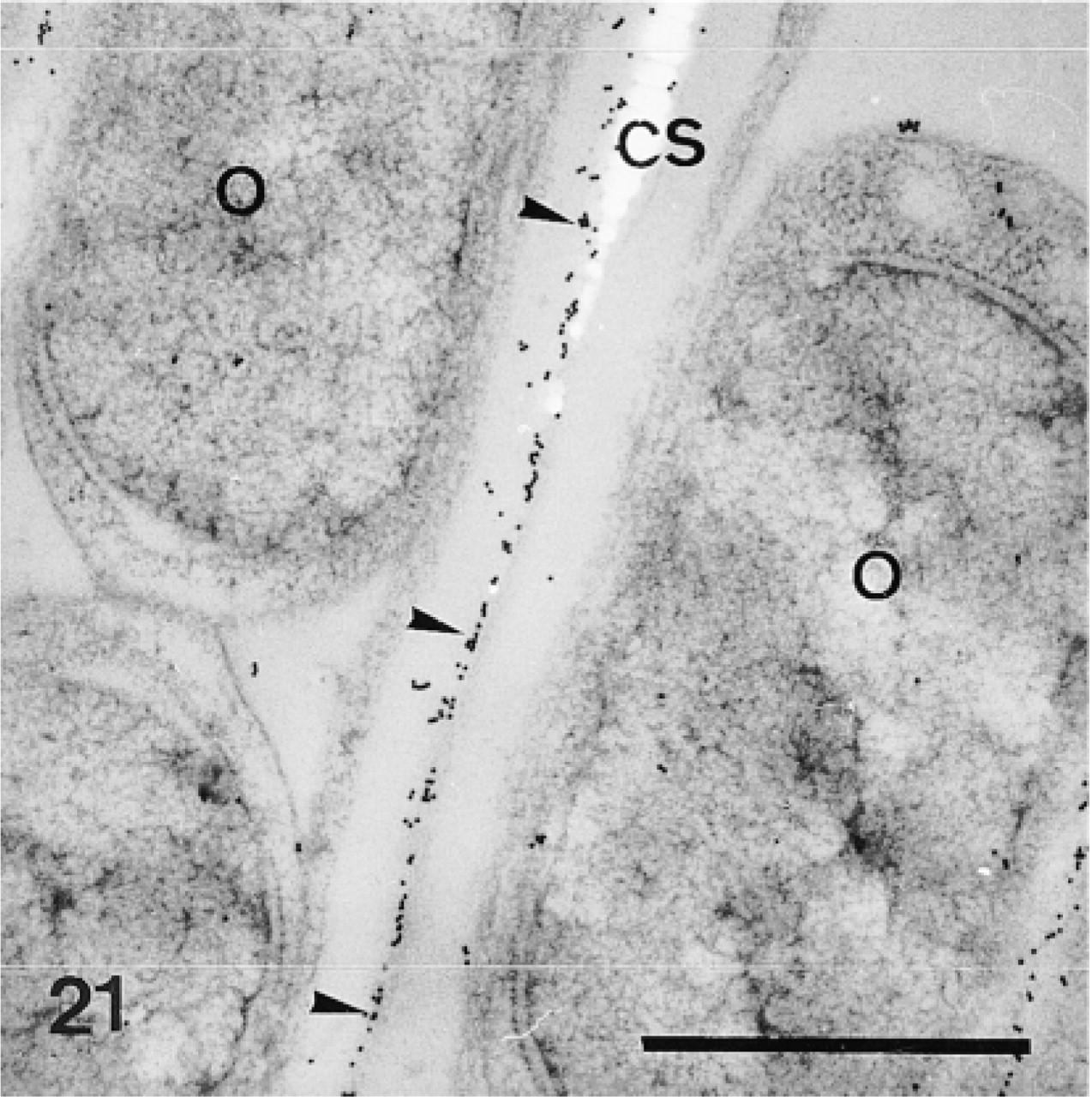
Odontoblasts (O) and calcification site (CS). Anti-SM50 labeling is confined to the organic matrix (arrowheads). Bar = 1 μm.
Calcification site. Anti-SM30 labeling is confined to the organic matrix (arrowheads). Bar = 250 nm.
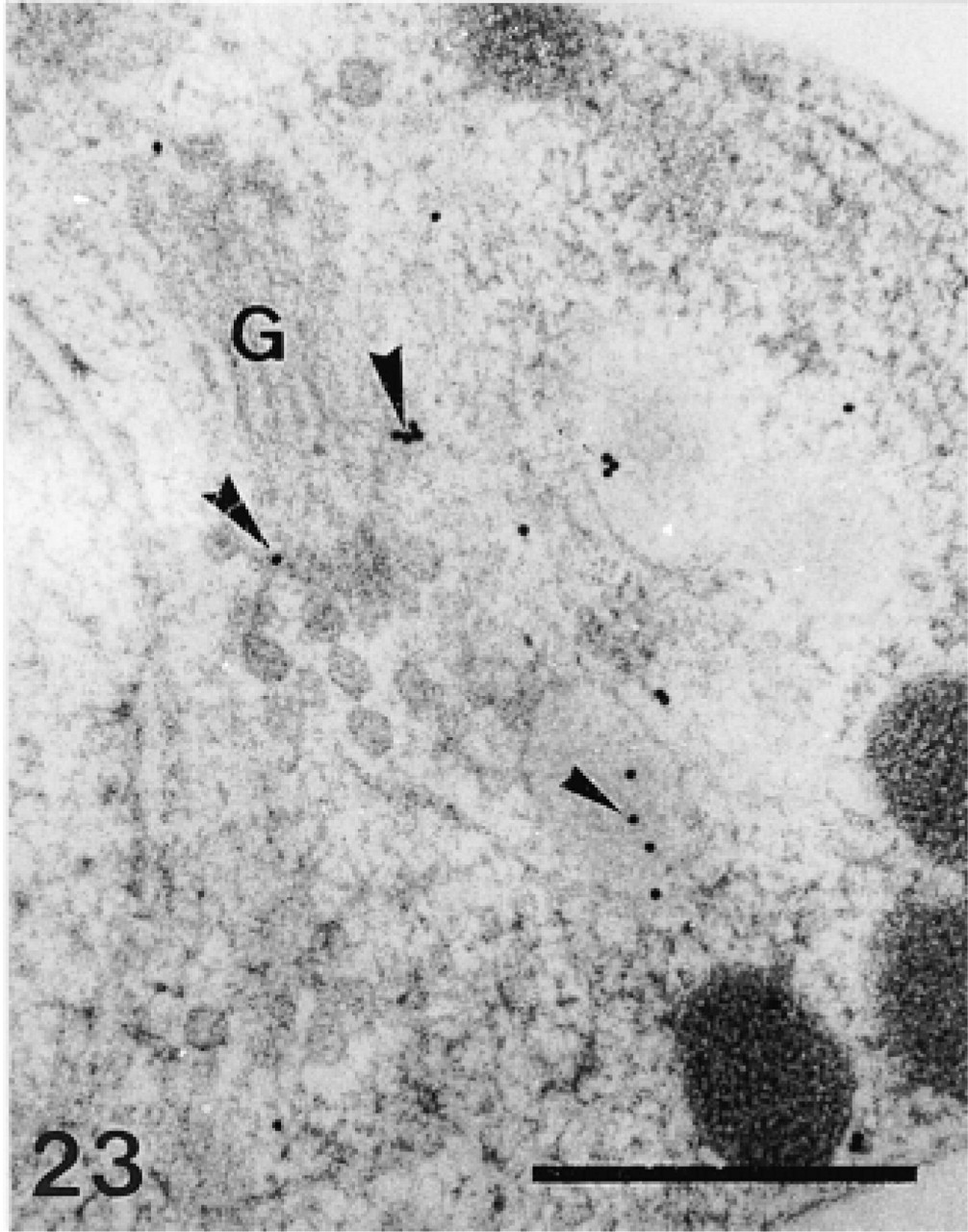
Anti-SM50 labeling of the Golgi apparatus (G) in a preodontoblast. Bar = 500 nm.
The anti-SM30 and anti-SM50 labeling was practically identical. Their intensity was lower than the intensity of the anti-total matrix labeling (compare Figures 21 and 22 with Figure 16 and Figure 23 with Figure 17). The anti-SM30 and anti-SM50 labeling was confined to the Golgi apparatus of the preodontoblasts (Figure 23) and to the organic matrix of the calcification sites (Figures 21 and 22). No labeling was observed in the extracellular matrix or along the plasma membrane of the odontoblasts. The only difference between the anti-SM30 and the anti-SM50 labeling was that the organic matrix was more weakly labeled with the anti-SM30 serum than with the anti-SM50 serum (compare Figures 22 and 21).
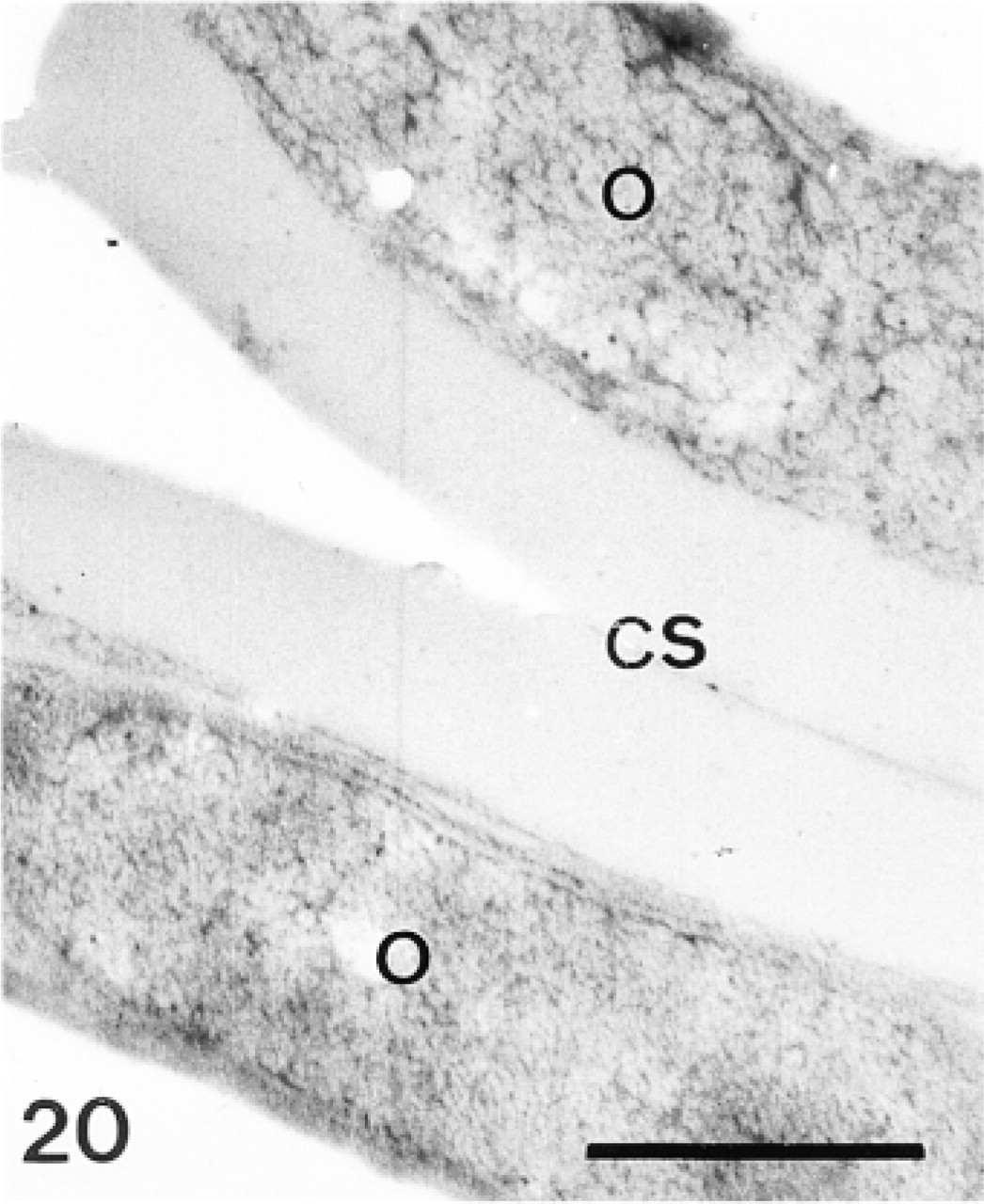
A summary of the structures labeled by the three sera studied is presented in Table 1. Control experiments omitting the primary sera showed that secondary antibodies induced a background, restricted to some components of the extracellular matrix and to the epidermic basal lamina, in the primordia and the test plates (Figures 10, 11, and 20).
Discussion
Four protocols, two conventional chemical protocols (CF1 with tannic acid and CF2 without tannic acid) and two high-pressure freezing/freeze-substitution protocols (HPF/FS1 with gallic acid and HPF/FS2 without gallic acid), were tested. The CF1 and HPF/FS1 protocols better preserved the spatial organization of the organic matrix but prevented any labeling. These protocols contain tannic or gallic acid, which are known to improve the preservation of the spatial organization of the echinoid organic matrix (Ameye et al. 1998) but also to decrease tissue antigenicity (Berryman et al. 1992). The two protocols without tannic and gallic acid (CF2 and HPF/FS2) allowed successful immunolabeling with the three sera; the same structures were labeled with both protocols. However, it is noteworthy that a better preservation of the ultrastucture was achieved by the CF2 protocol, whereas a higher preservation of the antigenicity was ensured by the HPF/FS2 protocol. The better preservation of the ultrastructure (particularly of the membranes) with the CF2 protocol is probably due to the presence of osmium tetroxide, which preserves the ultrastructure in two ways: by preserving, at least in part, the unsaturated lipids of the membranes (Hayat 1989) and by staining cell membranes (Hayat 1993). The higher intensity of labeling with the HPF/FS2 protocol can be explained by three factors: the use of uranyl acetate in the fixative solution, the high-pressure freezing/freeze-substitution procedure, and the embedding in LR Gold resin. Uranyl acetate has little or no adverse effect on immunolabeling (Humbel and Schwarz 1989), and the high-pressure freezing/freeze-substitution followed by embedding in LR Gold resin maintains the hydration shell of molecules, causing superior preservation of the antigenicity (Humbel and Schwarz 1989; Kellenberger 1991; Hippe-Sanwald 1993). Moreover, LR Gold does not bind covalently to cell components (and therefore does not chemically alter the epitopes of antigens) and polymerizes at low temperature (and thus avoids the harmful heat effect; Humbel and Schwarz 1989). If one compares the results obtained with the two protocols, the labeling with the CF2 protocol showed better microscopic resolution, whereas higher intensity of labeling with the HPF/FS2 protocol gave a more accurate picture of the in vivo localization of the antigens.
Except for the background due to labeling by the secondary antibodies of the extracellular matrix and epidermal basal lamina (in the primordia and the test plates), the anti-total matrix, anti-SM30, and anti-SM50 labeling was confined to the skeleton-forming cells and to the mineralization organic matrix (i.e., the organic material within the calcification sites) (see Table 1). The sclerocytes (in the pedicellaria primordium) and the preodontoblasts (in the tooth) had their Golgi stacks, Golgi vesicles and vesicles of the pseudopodia labeled by the three sera (Table 1). This indicates that the adult matrix glycoproteins follow the classical routes of secretory glycoprotein assembly and export, as already demonstrated in echinoid larvae (Benson et al. 1989). The glycoproteins are modified in the Golgi apparatus, exported in Golgi vesicles to the pseudopodium of the skeleton-forming cells, and eventually secreted into the calcification sites. Interestingly, the medium gray vesicles of the skeleton-forming cells were not labeled by any of the three antisera studied. The same is true in the echinoid larva (see Figure 3 in Benson et al. 1989). This indicates that these vesicles probably do not store or secrete spicule matrix proteins, contrary to the suggestion by Ingersoll and Wilt (1998).
The plasma membranes of all three skeleton-forming cells studied (sclerocytes, preodontoblasts, and odontoblasts) were labeled by the anti-total matrix serum but not by the anti-SM30- and anti-SM50-specific sera. This labeling is not related to the secretion route of the glycoproteins. It might be due to a crossreaction of the polyclonal serum with the msp 130 protein. The msp 130 is a membrane glycoprotein present in both echinoid larvae and adults (Anstrom et al. 1987; Parks et al. 1988; Drager et al. 1989). In larvae, it is restricted to the skeleton-forming cells (i.e., primary mesenchyme cells) (Farach et al. 1987) and shares an asparagine-linked anionic carbohydrate epitope with the SM30 (Brown et al. 1995; Farach-Carson et al. 1989). Because the specific anti-total matrix labeling in adults is also confined to the skeleton-forming cells and recognizes a membrane protein, we suggest that some antibody of the anti-total matrix serum is raised against the asparagine-linked anionic carbohydrate epitope of the SM30 and recognizes this epitope on the msp 130. It is noteworthy that the anti-SM30 antibody does not crossreact with that epitope (Brown et al. 1995) and, accordingly, did not label the plasma membranes.
Structures specifically labeled by the anti-total matrix, anti-SM30, and anti-SM50 sera in the pedicellaria primordium, test plates, and tooth of the echinoid Paracentrotus lividus a
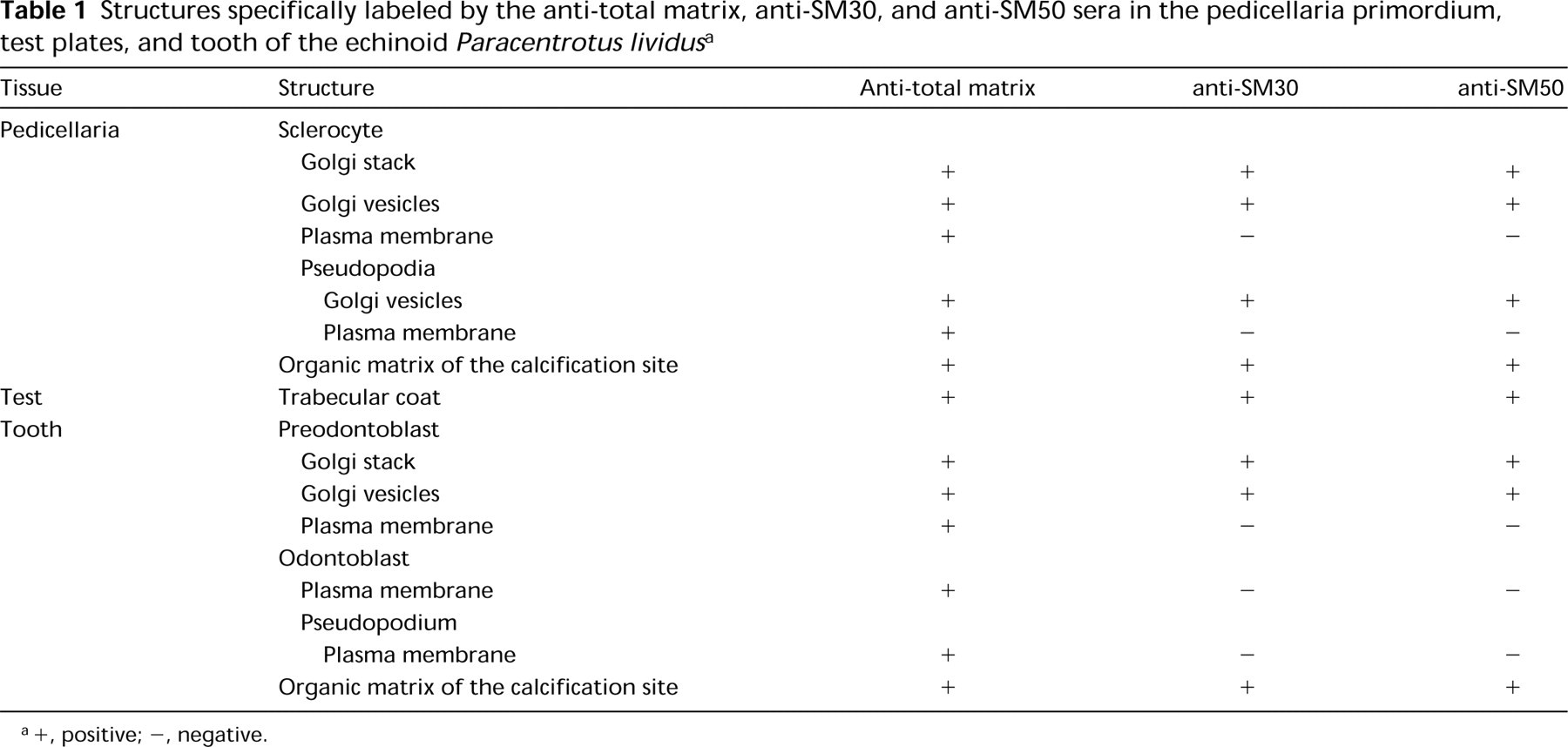
+, positive; -, negative.
In this study, the odontoblasts did not present well-developed Golgi apparatus and were not labeled by the anti-SM30 and anti-SM50, indicating that they were not involved at this time in the synthesis of these important matrix proteins. This is surprising because these cells closely surround still growing tooth plates (Märkel 1969; Wang et al. 1997). In contrast, odontoblasts located between older but still growing primary plates have well-developed Golgi apparatus, indicating active synthesis of proteins (unpublished observations). This implies that the synthesis of matrix proteins by the skeleton-forming cells is sometimes discontinuous during the formation of a skeletal element. Temporal variation in the synthesis of some organic matrix components was also evident in the larval skeleton-forming cells during spicule formation (Harkey et al. 1995; Guss and Ettensohn 1997).
In the tooth, anti-SM30 and anti-SM50 labeling is restricted (within the calcification site) to the structured organic matrix that is pre-formed before the onset of biomineralization (see Ameye et al. 1998), indicating that both proteins are probably not in soluble form within the calcification site. (Precipitation due to preparation of the samples could have occurred but, if the proteins were soluble, they would have been formed on both the matrix and the membrane of the calcification site.) Labeling on the edges of the perforation within these sites probably results from shrinking of the sections under the electron beam around these holes. This shrinking probably produced an artificial “concentration” of the labeled matrix.
Unlike Kitajima et al. (1996), we did detect SM30 in the test plates. This could be due either to the lower sensitivity of the RNA immunoblot analysis used by these authors in comparison with immunoelectron microscopy or to an actual absence of SM30 RNA transcripts. The absence of anti-SM30 labeling in the sclerocyte pseudopodia of the mature test supports the latter hypothesis. The detection of SM30 in the test of Paracentrotus lividus (present study) probably rules out the suggestion that SM30 expression could be linked to orientation of the crystallographic axes of the considered ossicles (each echinoderm ossicle is a single crystal of high-magnesium calcite) (see Kitajima et al. 1996). Indeed, test plates of both Paracentrotus lividus and Hemicentrotus pulcherrimus (the species studied by Kitajima and co-authors) show similar c-axis orientations: c-axis is tangential to the plate in P. lividus and tangential to oblique (maximum of 45°) in H. pulcherrimus (Raup 1962).
The present results demonstrate the first immunolabeling of organic matrix components in the adult echinoid skeleton and tooth. The presence of SM30 and the SM50 in postmetamorphic tegumentary skeleton has previously been documented from RNA blot experiments (Richardson et al. 1989; George et al. 1991; Killian and Wilt 1996; Kitajima et al. 1996), but this is the first report of the presence of SM30 and SM50 in the tooth. This immunolabeling confirms that at least part of the skeletogenesis genetic program is conserved among different developmental stages (from larvae to adults) and also between different calcification models (i.e., the tegumentary skeleton and the tooth). This conclusion is in agreement with the morphological and crystallographic similarities shown by the larval and adult skeletons (see Dubois 1990; Beniash et al. 1997; Wang et al. 1997).
Footnotes
Acknowledgements
Supported by a FRIA grant (to LA) by FRFC grant 2.4512.95, and by NFSR grant 1.5.201.98. PhD is a Research Associate of the NFSR (Belgium). Contribution from the Centre Interuniversitaire de Biologie Marine (CIBIM).
We thank Ph. Grosjean of the Centre de Recherches et d′Etudes Côtières (University of Caen) for providing the echinoids. LA is grateful for the hospitality shown to him by Professor Mueller and colleagues while a visitor at Laboratory for Electron Microscopy 1 (ETH, Zürich, Switzerland) to carry out the cryotechnical part of this work.