
Abstract
Alexa 350, Alexa 430, Alexa 488, Alexa 532, Alexa 546, Alexa 568, and Alexa 594 dyes are a new series of fluorescent dyes with emission/excitation spectra similar to those of AMCA, Lucifer Yellow, fluorescein, rhodamine 6G, tetramethylrhodamine or Cy3, lissamine rhodamine B, and Texas Red, respectively (the numbers in the Alexa names indicate the approximate excitation wavelength maximum in nm). All Alexa dyes and their conjugates are more fluorescent and more photostable than their commonly used spectral analogues listed above. In addition, Alexa dyes are insensitive to pH in the 4-10 range. We evaluated Alexa dyes compared with conventional dyes in applications using various conjugates, including those of goat anti-mouse IgG (GAM), streptavidin, wheat germ agglutinin (WGA), and concanavalin A (ConA). Conjugates of Alexa 546 are at least twofold more fluorescent than Cy3 conjugates. Proteins labeled with the Alexa 568 or Alexa 594 dyes are several-fold brighter than the same proteins labeled with lissamine rhodamine B or Texas Red dyes, respectively. Alexa dye derivatives of phalloidin stain F-actin with high specificity. Hydrazide forms of the Alexa dyes are very bright, formaldehyde-fixable polar tracers. Conjugates of the Alexa 430 (ex 430 nm/em 520 nm) and Alexa 532 (ex 530 nm/em 548 nm) fluorochromes are spectrally unique fluorescent probes, with relatively high quantum yields in their excitation and emission wavelength ranges.
T
Some of these aims have been obtained by sulfonation of certain cyanine dyes. Sulfonation decreases the inherent tendency of molecules to form aggregates, presumably due to the increased polarity imparted by the sulfonic acid moiety (Mujumdar et al. 1993). Ring sulfonation also increases the brightness of many dyes in aqueous media, including the carbocyanines Cy3 and Cy5 (Wessendorf and Brelje 1992) and some pyrenes, such as Cascade Blue (Whitaker et al. 1991).
All Alexa dyes were used in succinimidyl ester form for conjugation with the primary amines of biomolecules. This reactive form of dyes yields the most stable conjugates and allows reactions to take place efficiently at pH 7.5-8.5 or at an even lower pH when required (Haugland 1995).
We report here the superior, largely unparalleled fluorescence emission and photostability of conjugates of a new series of sulfonated compounds named Alexa 350, Alexa 430, Alexa 488, Alexa 532, Alexa 546, Alexa 568, and Alexa 594 dyes. The number in each of their names refers to the approximate excitation maxima of the dyes, which nearly match the principal wavelength of several commonly used excitation sources. All of the Alexa dyes exhibit pH-insensitivity over a very broad range, and those that absorb at wavelengths greater than 480 nm have the high molar extinction coefficient typical of fluoresceins or rhodamines.
Sulfonation of the aminocoumarin or rhodamine molecule to obtain the Alexa dyes was performed by substituting one or more hydrogen atoms in the dyes with sulfonic acid or one of its salts. The introduction of sulfonic acid groups bestows negative charges on the dye that make the water-insoluble zwitterionic rhodamine or neutral coumarin molecules much more hydrophilic, allowing conjugations to be performed also in the absence of organic solvents. Furthermore, the net negative charges of the fluorophores decrease the interaction between molecules that strongly hinders the fluorescence yields of rhodamine conjugates. Therefore, the coumarin and rhodamine molecules have been modified by sulfonation to yield probes for the commonly used excitation wavelengths from UV to the 594-nm line of the orange He-Ne laser. We have prepared very bright conjugates of Alexa 532, a highly fluorescent dye that can be excited at the optimal emission line of the frequency-doubled Nd-YAG laser, a wavelength range in which there are few other readily available fluorescent dyes, the conjugates of which are only weakly fluorescent. Both the Alexa 350 and Alexa 430 dye, which are based on 7-aminocoumarin, have lower extinction coefficients (∼19,000 cm−1 M−1) but exhibit high Stokes shifts, greater than 70 nm.
Our results show that the Alexa dye conjugates of antibodies, streptavidin, and other proteins (e.g., DNase, fibrinogen) are the brightest and among the most photostable low molecular weight fluorescent probes available.
Materials and Methods
Succinimidyl esters of the Alexa 350, Alexa 430, Alexa 488, Alexa 532, Alexa 546, Alexa 568, Alexa 594 dyes, carboxyrhodamine 6G, carboxytetramethylrhodamine and Texas Red-X, and fluorescein isothiocyanate, Texas Red sulfonyl chloride and propidium iodide, as well as streptavidin, its conjugates and anti-α-tubulin mouse monoclonal antibody (MAb) were all from Molecular Probes (Eugene, OR). Goat anti-mouse IgG (GAM) and F(ab′)2 fragment of GAM were from Fortron Bio Science (Morrisville, NC). ConA and WGA were from EY Laboratories (San Mateo, CA). Deoxyribonuclease, fibrinogen, and transferrin were from Sigma Chemical (St Louis, MO). Cy3 conjugates of streptavidin and GAM were from Amersham Life Science (Arlington Heights, IL). The biotinylated mouse MAb CD16/CD32 was purchased from PharMingen (San Diego, CA). All cell culture supplies were obtained from GIBCO-BRL (Gaithersburg, MD). RAW264.7 cells (mouse macrophages, Abelson leukemia) and BPAE cells were obtained from American Type Culture Collection (Rockville, MD) and were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal calf serum. The antiphotobleaching reagents tested were SlowFade and SlowFade Light, ProLong (Molecular Probes), Vectashield (Vector Laboratories; Burlingame, CA), Fluorsave (Calbiochem; La Jolla, CA), Immuno-mount (Shandon; Pittsburgh, PA), Gel Mount, Crystal Mount, Clarion (Biomeda; Foster City, CA), Aqua-Poly Mount (Polysciences; Warrington, PA), and Cytoseal (Stephens Scientific; Riverdale, NJ). All commercial antifade reagents and mounting media were applied according to the manufacturer's instructions. All salts and other chemicals were of reagent grade or of high purity.
Absorbance measurements were performed with a Hitachi U-2000 spectrophotometer (Hitachi Instruments; Boulder, CO). Fluorescence measurements were performed using an SLM-Aminco fluorometer SLM Instrument (Urbana, IL). Flow cytometry data acquisition was performed using a Becton-Dickinson FACS Vantage flow cytometer (San Jose, CA). The 488-nm line of an air-cooled argon-ion Enterprise laser was used at 100 mW. Sample acquisition and analysis were performed using CellQuest v.1.2 software. The photomultiplier tubes referred to as FL1, FL2, and FL3 were equipped with 530 ± 15-nm, 575 ± 13-nm and 633 ± 15-nm emission filters, respectively. A 560SP dichroic filter was used between the FL1 and FL2 channels. Single cell populations were identified using a plot of the forward angle light scatter vs side scatter. Electronic gating was used for selecting live cells from the propidium iodide-positive dead cells. For every sample, at least 10,000 events were collected and used for data analysis.
Epifluorescence microscopy was performed with either a Nikon Labophot II microscope equipped with a QuadFluor fluorescence adapter or with a Nikon Diaphot TMD inverted fluorescence microscope. Either a X40/0.75 NA Plan Fluor, X40/1.3 NA Fluor, or a X60/1.4 NA Plan Apo objective lens and optical filter sets (Omega Optical; Brattleboro, VT) appropriate for each dye were used. The signal from fluorescently labeled tubulin in the cells was monitored in a Nikon Diaphot TMD microscope equipped with a X40 Fluor 1.3 NA oil objective lens, 100-W Hg lamp, and Quantix CCD camera. Quantitative fluorescence intensity data were acquired using MetaMorph software. Optical filters used were as follows: Alexa 350, ex 365 ± 8 nm/em 450 ± 33 nm (XF06); Alexa 430, ex 455 ± 35 nm/em ≥515 nm (XF18); Alexa 488, ex 485 ± 11 nm/em 530 ± 15 nm (XF22); Alexa 532, ex535 ± 17.5 nm/em 590 ± 17.5 nm (XF32); Alexa 546, ex 546 ± 5 nm/em 580 ± 15 nm (XF37); Alexa 568, ex 560 ± 20 nm/em 610 ± 20 nm (XF41); Alexa 594, ex 590 ± 22.5 nm/em 660 ± 16 nm (XF44) (XF numbers represent Omega Optical filter sets). Color photomicrographs were acquired with Ektachrome Elite II ASA 400 slide film. For the photobleaching experiments, digital images were acquired with Star-1 cooled CCD camera (Photometrics; Tucson, AZ), saved in 12-bit format, and processed with Image-1 software (Universal Imaging; Malvern, PA). Some postacquisition image processing was performed with Photoshop software (Adobe Systems; San Jose, CA).
The stability to pH of Alexa dyes was monitored by measuring their absorption spectra in buffers at pH 4-9 after 10-60 min of exposure in the presence and absence of 1 mM CaCl2.
Conjugations of the proteins with the various dyes were performed at pH 8.3, with the exception of those with FITC (Banks and Paquette 1995) and Texas Red (Titus et al. 1982), which were done at pH 9.0. When conjugations are performed by dissolving the succinimidyl ester of the dyes in aqueous media, the stock solution of the dyes must be used immediately after preparing it because the reactive portion of the dye hydrolyzes rapidly. Because we used the same dye stock solution to perform multiple conjugations simultaneously, we chose to dissolve the dyes in an organic solvent, which preserves the reactivity of the dyes much longer. Therefore, the dyes were dissolved at 10 mg/ml in anhydrous dimethylformamide (DMF) and aliquots were immediately added to the protein solutions to obtain the desired dye-to-protein molar ratios while stirring. In the case of Lucifer Yellow, which is not stable in its activated form, the dye was made into its succinimidyl ester derivative immediately before starting the conjugation procedure. The reaction mixtures were incubated for 90 min at room temperature, stopped by the addition of hydroxylamine at pH 8.0 to obtain the final concentration of 0.15 M, and incubated for an additional 30 min. The conjugates were purified from unreacted dye by size-exclusion chromatography using Bio-Gel P-30 (Bio-Rad Laboratories; Hercules, CA) or Matrex Cellufine GH-25 (Amicon; Beverly, MA), with or without a small addition of Toyopearl HW-40 F (TosoHaas; Montgomeryville, PA) on the top of the column. Absence of unreacted dye was assayed by thin-layer chromatography on silica gel plates using water:acetonitrile (20:80) as solvent for the conjugates of Alexa and Cy3 dyes and using chloroform:methanol:acetic acid (70:25:5) for all the other dye conjugates. The degree of labeling was determined as the ratio of moles of fluorophore to moles of protein (F/P) as previously described (Haugland 1995). The relative quantum yield (RQY) of the conjugates was calculated using the ratio of the integrated emissions of the conjugate and the appropriate dye standard. The conjugates being compared were matched for identical absorbance to that of the dye standard at the excitation wavelength.
Immunofluorescence staining was performed using commercially prepared Hep-2 human cell slides and human anti-nuclear antibody, both supplied by INOVA Diagnostics (San Diego, CA). The cells were stained with mouse anti-human IgG, followed by GAM-dye conjugate or biotinylated protein G and streptavidin-dye conjugates. Images were viewed with the fluorescence microscope as described above and the integrated fluorescence intensity of each field was quantitated with Image-1 software.
Cell staining for flow cytometry experiments was performed as follows. Approximately 1.0 million RAW 264.7 cells, suspended in PBS, pH 7.2, containing 1% bovine serum albumin and 2 mM sodium azide, were incubated with 5 μg of biotinylated anti-mouse CD16/CD32 antibody or the equivalent volume of PBS for 30 min on ice. After washing with the same buffer, both antibody-labeled and unlabeled cells were incubated in triplicate with 2 μg of the Alexa 488, Oregon Green 488, fluorescein, Rhodol Green, or BODIPY FL conjugates of streptavidin. After a 30-min incubation on ice, the cells were washed twice. To discriminate dead cells, 1 μl of a 1 mg/ml aqueous solution of propidium iodide was added before flow cytometry data acquisition.
The multicolor image (see Figure 8) was obtained by incubating bovine pulmonary artery epithelial (BPAE) cells in culture medium for 2 hr at 37C with 2 μM Alexa 594 hydrazide, which, as expected, localized in the lysosomes (Storrie and Ferris 1985). After removal of the excess hydrazide, the cells were fixed, permeabilized, and incubated with a cocktail of anti-bovine cytochrome oxidase Subunit V mouse MAb 3G5-F7-G3 and anti-human cytochrome oxidase Subunit I MAb 5D4-F5 to label mitochondria. Using a mixture of two or more MAbs often generates a stronger signal because two or multiple haptens of the same targeted protein are probed simultaneously. The incubation was carried out for 60 min, the cells were washed, and the mitochondria were labeled with 10 μg/ml Alexa 350-conjugated GAM for 30 min. Finally, to visualize actin filaments, the cells were incubated with Alexa 488-phalloidin (Barak et al. 1980). Quantitative imaging of immunofluorescent staining of fixed cells was used to measure the brightness and photostability of IgG conjugates of the Alexa 350, Alexa 430, Alexa 488, Alexa 532, Alexa 546, Alexa 568, and Alexa 594 dyes with IgG in PBS, Mowiol, and SlowFade (Longin et al. 1993) or other commercially available mounting media listed above. Photobleaching measurements were carried out using BPAE cells fixed in 3.75% formaldehyde and permeabilized with 0.1% Triton X-100 in PBS. Nonspecific staining was blocked by incubating cells with PBS containing 6% BSA. Cells on coverslips were incubated with anti-α-tubulin mouse MAb for 60 min and subsequently labeled for 30 min with a GAM conjugate of an Alexa fluorophore. After washing in PBS, the coverslip was inverted over 10 μl of mounting medium on a slide and sealed with paraffin. Using a variety of mounting media, measurements were performed on cells having similar morphology and fluorescence distribution.
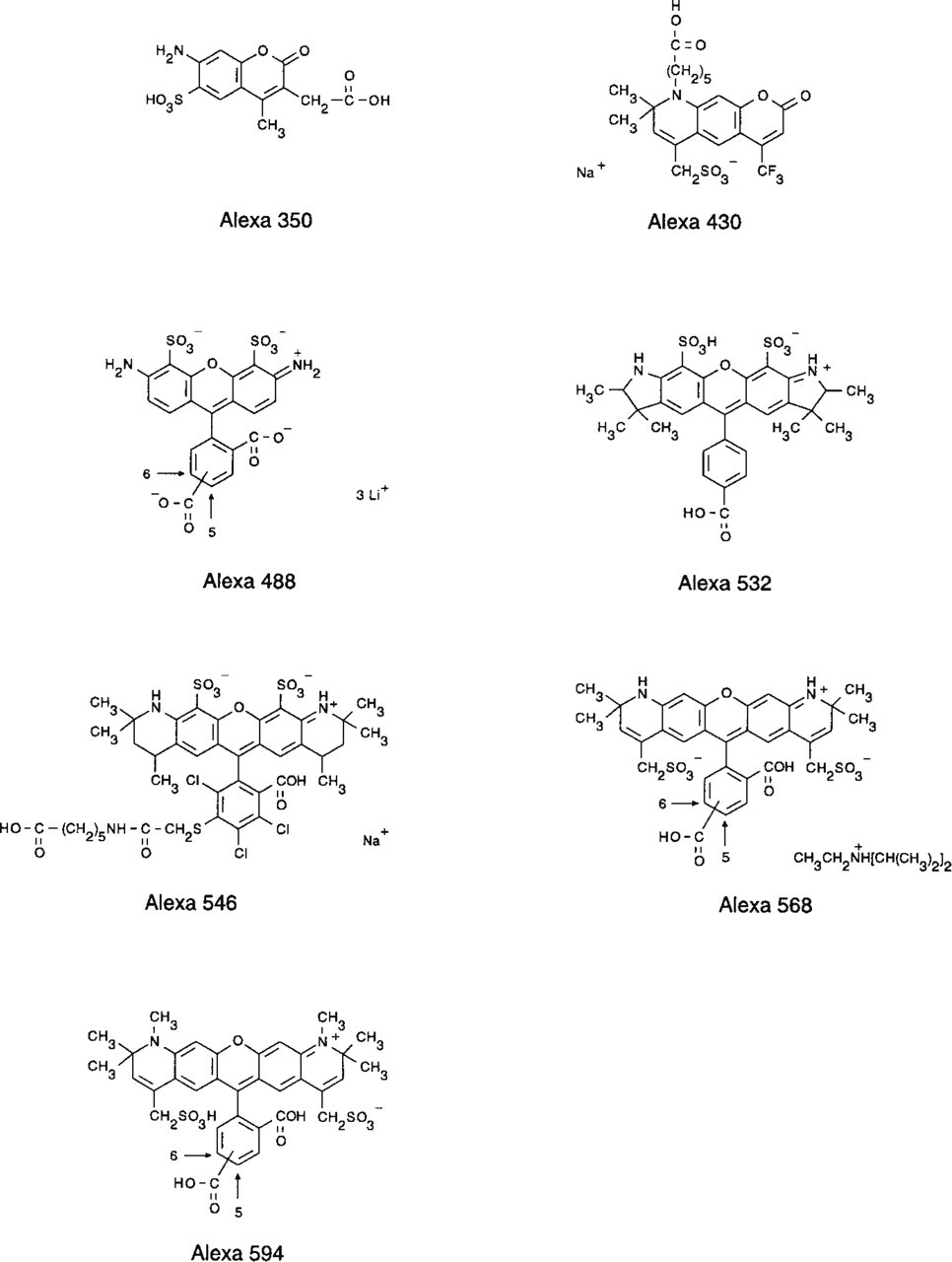
Chemical structures of the Alexa dyes.
Results and Discussion
The chemical structures of the Alexa dyes are shown in Figure 1. The normalized excitation and emission spectra of the Alexa dye series are illustrated in Figure 2. The excitation and emission ranges of the Alexa dyes cover the entire spectrum from ultraviolet to red, comprising wavelengths that match conventional light sources and including Alexa 430 and Alexa 532 dyes that, to our knowledge, represent the best dyes with good relative quantum yield in their excitation wavelength range. The Alexa 430 dye, which emits at 525 nm, is an ideal candidate as an acceptor dye from a donor that is excitable in the UV range. Alexa 532 conjugates are suitable for excitation by the frequency-doubled Nd-YAG laser.
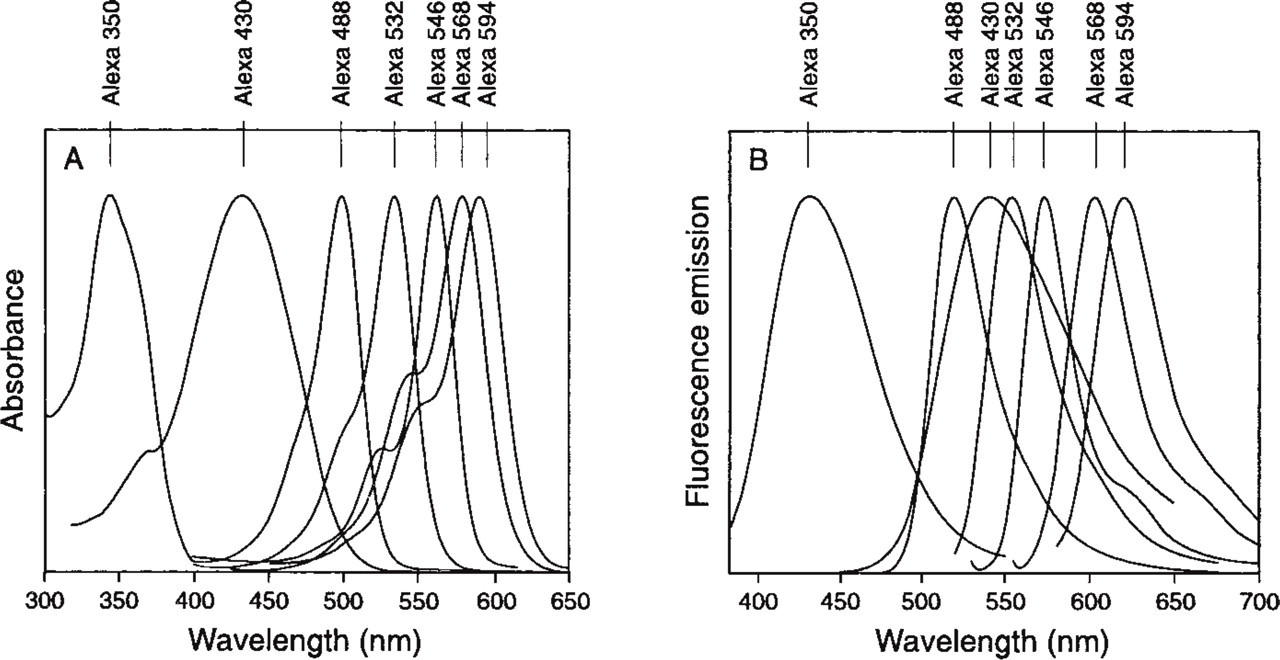
Normalized absorption, (
Addition of 1 mM CaCl2 or dissolving each Alexa dye at identical concentration in buffers of various pH, from pH 4.0 to pH 9.0, did not change the intensity of the fluorescence emission or the profile of the excitation/emission spectra (data not shown).
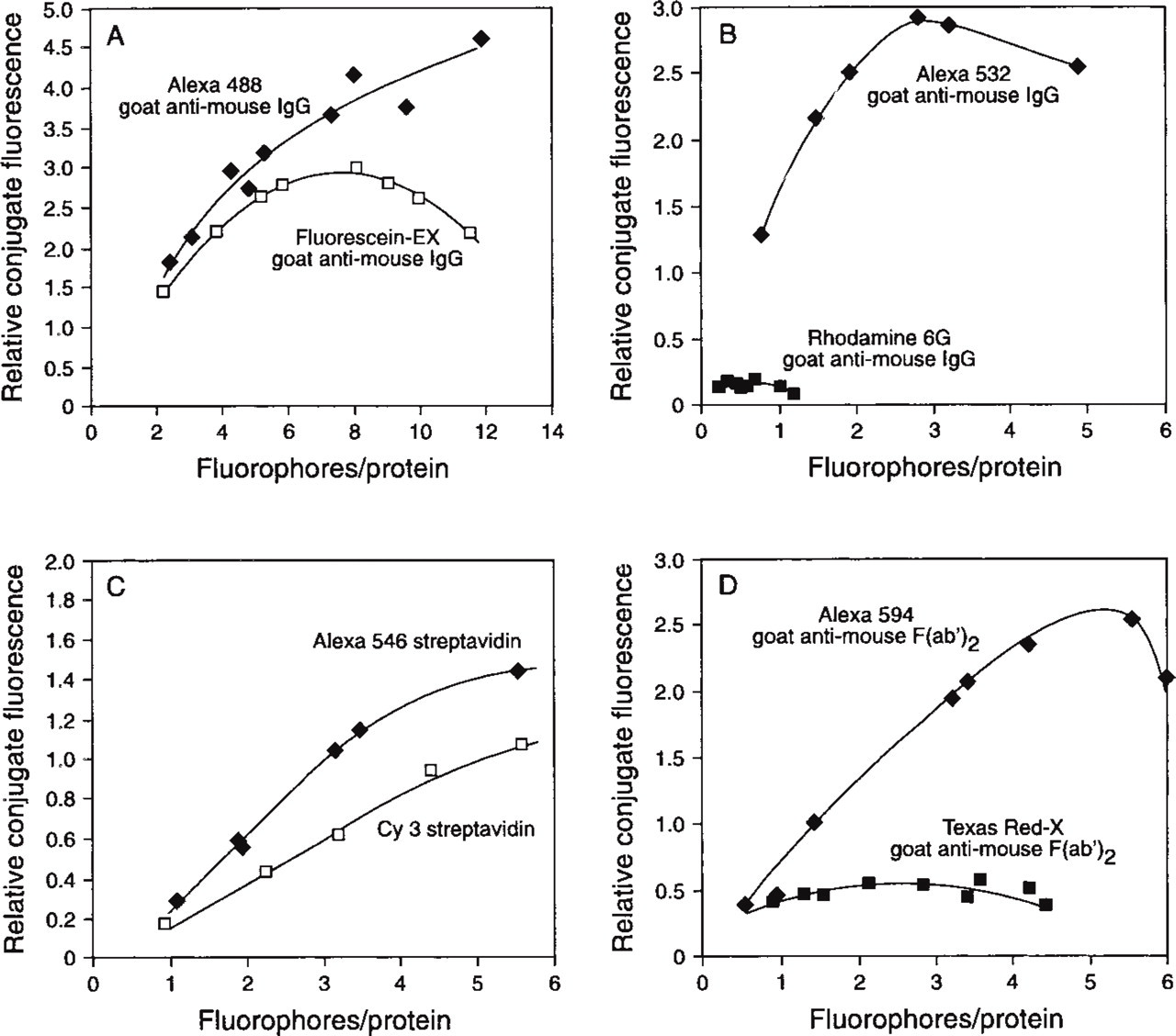
Comparison of the conjugate fluorescence of Alexa 488-GAM with fluorescein-EX-GAM (
To compare the fluorescence intensity of the protein conjugates of Alexa dyes and the conjugates of dyes with similar spectral properties, we derivatized various proteins with dyes at different dye-to-protein molar ratios. Simultaneously, we were able to determine the optimal degree of labeling for each protein, i.e., the number of dyes per protein that yields the most fluorescent conjugate with good preservation of biological activity. In general, less precipitation of proteins was observed during conjugation with Alexa dyes than with spectrally analogous dyes, such as Texas Red in comparison to Alexa 594 dye or tetramethylrhodamine in comparison to Alexa 546 dye. On conjugation, Alexa dye conjugates do not undergo the alteration in their absorption spectra typical of rhodamine dye conjugates. The absorbance spectra of conjugates of tetramethylrhodamine labeled with more than one mole of dye per mole of protein show a pronounced increase in the shoulder at 520 nm, apparently due to interaction between the dyes or between the dye and the protein. A concomitant loss in fluorescence is observed because the new conjugate labeled with aggregated tetramethylrhodamine dyes, which has an absorption maximum at 520 nm, is not fluorescent (Ravdin and Axelrod 1977). The differences in fluorescence intensity among some of the Alexa dye conjugates and the conjugates of the corresponding dye of similar spectral properties are illustrated in Figures 3–5 and Table 1.
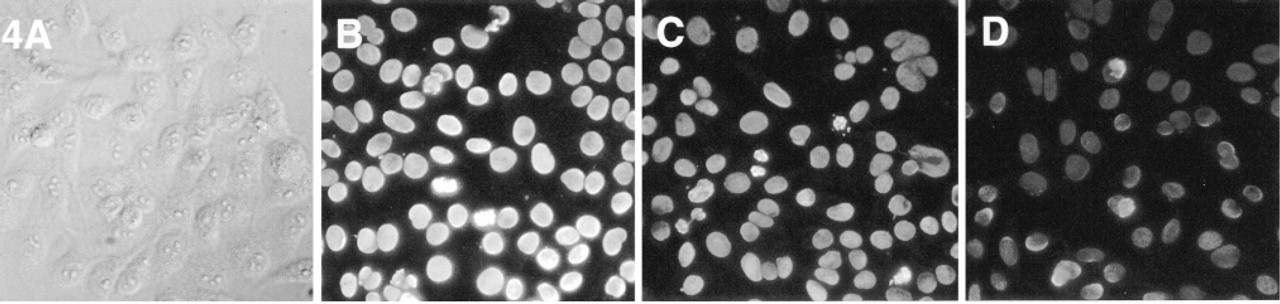
Immunofluorescent staining of Hep2 cells obtained with Alexa 488-GAM F(ab′)2 conjugates compared to conjugates with other green fluorescent dyes labeled with a similar number of fluorophores. DIC image (
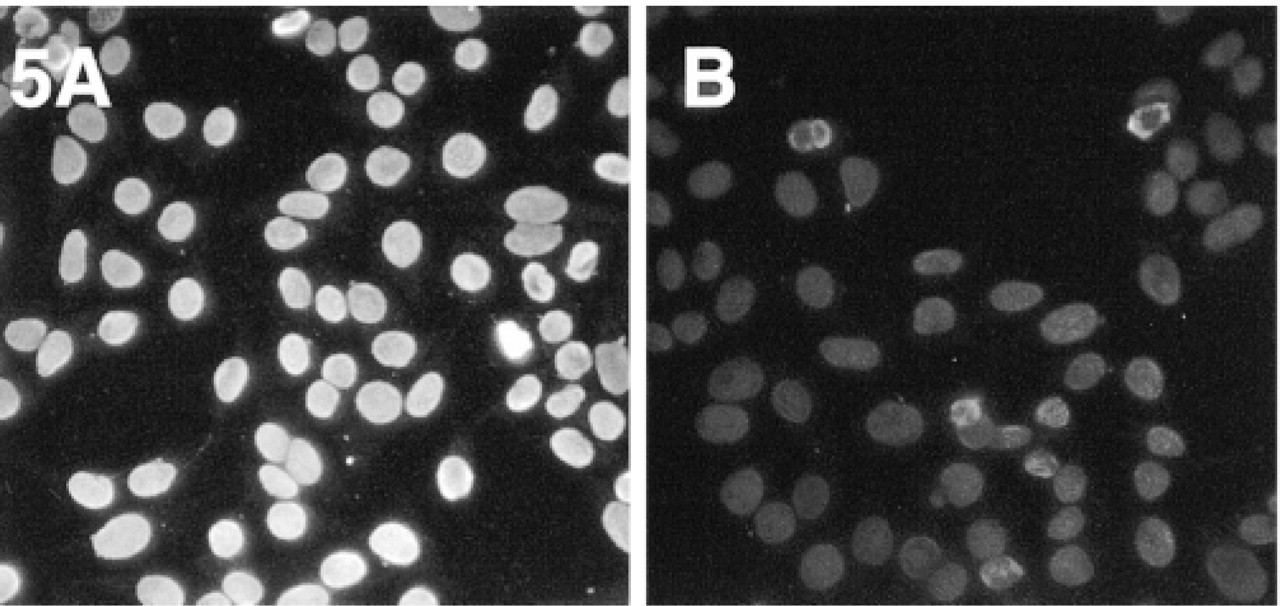
Experiments and results analogous to those illustrated in Figure 4, showing the superior brightness of the images obtained with Alexa 546-GAM, F/P = 4.0 (
Figure 3A shows that the conjugate of GAM with Alexa 488 dye is more fluorescent than the GAM conjugate of fluorescein (FITC) at the optimal degree of labeling. The curves of fluorescence intensity vs degree of labeling typically show a broad peak because the conjugate fluorescence is being quenched after reaching the optimal degree of labeling. However, Alexa 488-GAM is still very bright even at high numbers of dye molecules per molecules of antibody, whereas the fluorescence of fluorescein-GAM is generally dimmer and quenched at above 8 dyes/GAM. Alexa 532 conjugates of GAM are more than 10-fold brighter than GAM labeled with rhodamine 6G (Figure 3B). As shown in Figure 3C, Alexa 546-streptavidin conjugates are brighter than the conjugates of Cy3. In addition, the Alexa 594-GAM F(ab′)2 conjugate (Figure 3D) is about fivefold brighter than the corresponding conjugate of Texas Red-X. We have previously shown that Texas Red-X conjugates are, in general, already more fluorescent than the conjugates made with Texas Red sulfonyl chloride (Lefevre et al. 1996).
An example of the difference in the immunofluorescent staining obtained with Alexa 488-GAM F(ab′)2 and Oregon Green 488 or fluorescein-EX conjugates, prepared with the same lot of GAM and similar degree of labeling, is shown in Figure 4, in which the images of the nuclei of Hep-2 cells were obtained using the same exposure time. A differential interference contrast (DIC) image is included to show that, as expected, only the nuclei are stained. An analogous comparison between GAM conjugates of Alexa 546 and a commercially available Cy3-GAM conjugate is shown in Figure 5, demonstrating the superior fluorescence emission of the Alexa 546 conjugate.
Comparison of the fluorescence of Alexa conjugates with that of conjugates made with conventional dyes
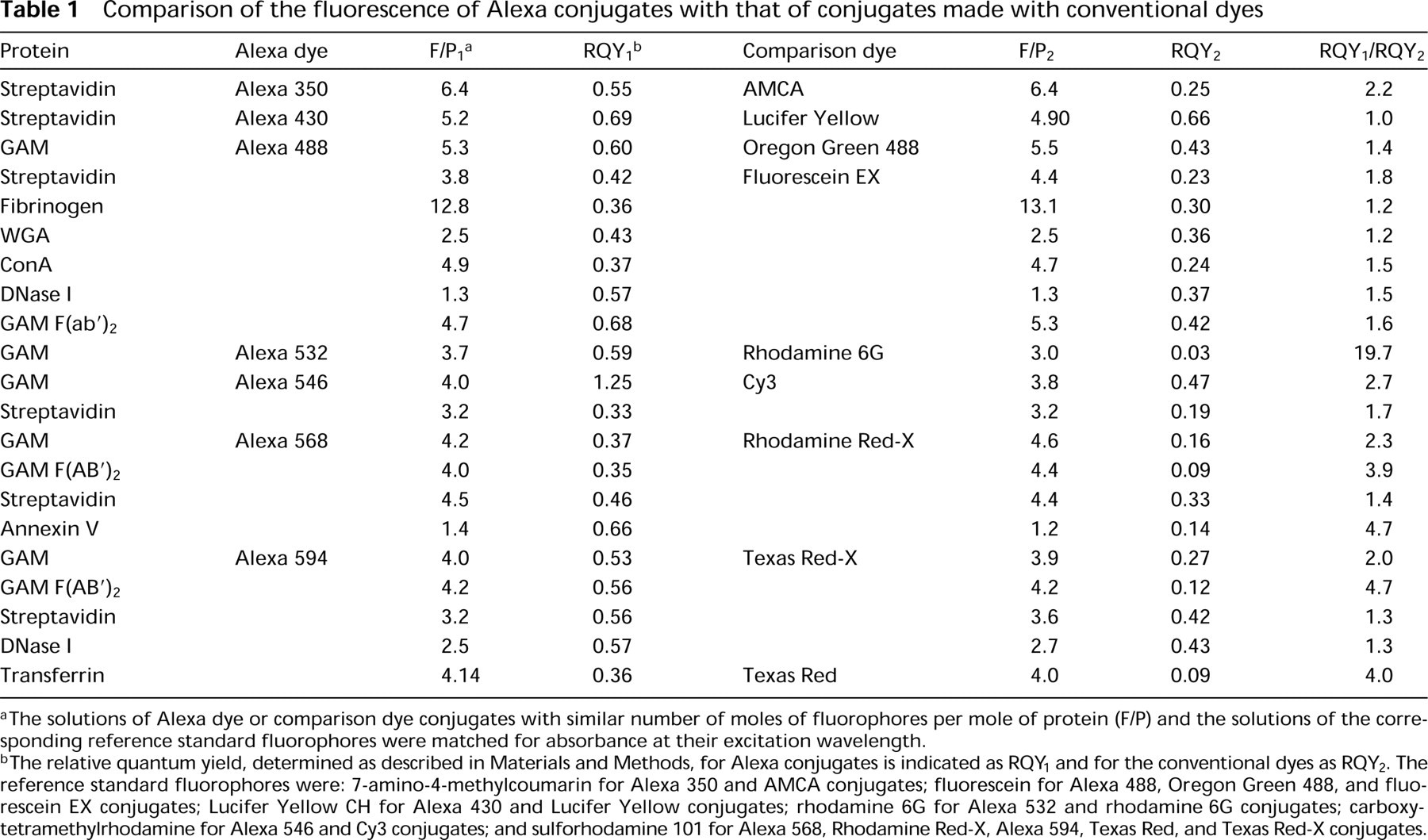
The solutions of Alexa dye or comparison dye conjugates with similar number of moles of fluorophores per mole of protein (F/P) and the solutions of the corresponding reference standard fluorophores were matched for absorbance at their excitation wavelength.
The relative quantum yield, determined as described in Materials and Methods, for Alexa conjugates is indicated as RQY1 and for the conventional dyes as RQY2. The reference standard fluorophores were: 7-amino-4-methylcoumarin for Alexa 350 and AMCA conjugates; fluorescein for Alexa 488, Oregon Green 488, and fluorescein EX conjugates; Lucifer Yellow CH for Alexa 430 and Lucifer Yellow conjugates; rhodamine 6G for Alexa 532 and rhodamine 6G conjugates; carboxytetramethylrhodamine for Alexa 546 and Cy3 conjugates; and sulforhodamine 101 for Alexa 568, Rhodamine Red-X, Alexa 594, Texas Red, and Texas Red-X conjugates.
A comparison of the brightness of green fluorescence-emitting dyes, as analyzed by flow cytometry, is plotted in Figure 6, where the fluorescence output of the Alexa 488 and of Oregon Green 488 conjugates of streptavidin outperform that of the fluorescein, BO-DIPY FL, and Rhodol Green conjugates. All the streptavidin conjugates compared in this experiment were labeled with a similar number of dye molecules per streptavidin molecule. Although photostability is not a prerequisite in most flow cytometry experiments, the higher fluorescence intensity of Alexa 488 conjugates is advantageous for applications in which a direct strong signal is needed to detect sparse receptors or structures.
A more detailed demonstration of the superior fluorescence of Alexa dye-protein conjugates is shown in Table 1. This table compares the relative quantum yields of diverse protein conjugates of Alexa dyes to those of the corresponding conjugates of dyes of similar excitation and emission, at similar numbers of moles of dye per mole of protein. With the exception of the Alexa 430 dye (see below), Alexa dyes consistently yielded conjugates of higher fluorescence intensities. The amount of increase in fluorescence is different for each protein and is of particular significance for the streptavidin conjugates. In fact, in our experience, the emission of fluorophores is considerably more quenched on conjugation with biotin binding proteins, such as avidin, streptavidin, or NeutrAvidin, than on conjugation with antibodies. Conjugates of the Alexa 430 dye (ex 430 nm/em 540 nm) have a fluorescence intensity and the unusually broad Stokes shift of approximately 110 nm similar to those of Lucifer Yellow. Moreover, the Alexa 430 dye is available as an amine-reactive succinimidyl ester, whereas amine-reactive forms of Lucifer Yellow are unstable, limiting the use of this dye as a label for biomolecules (Brelje et al. 1993).
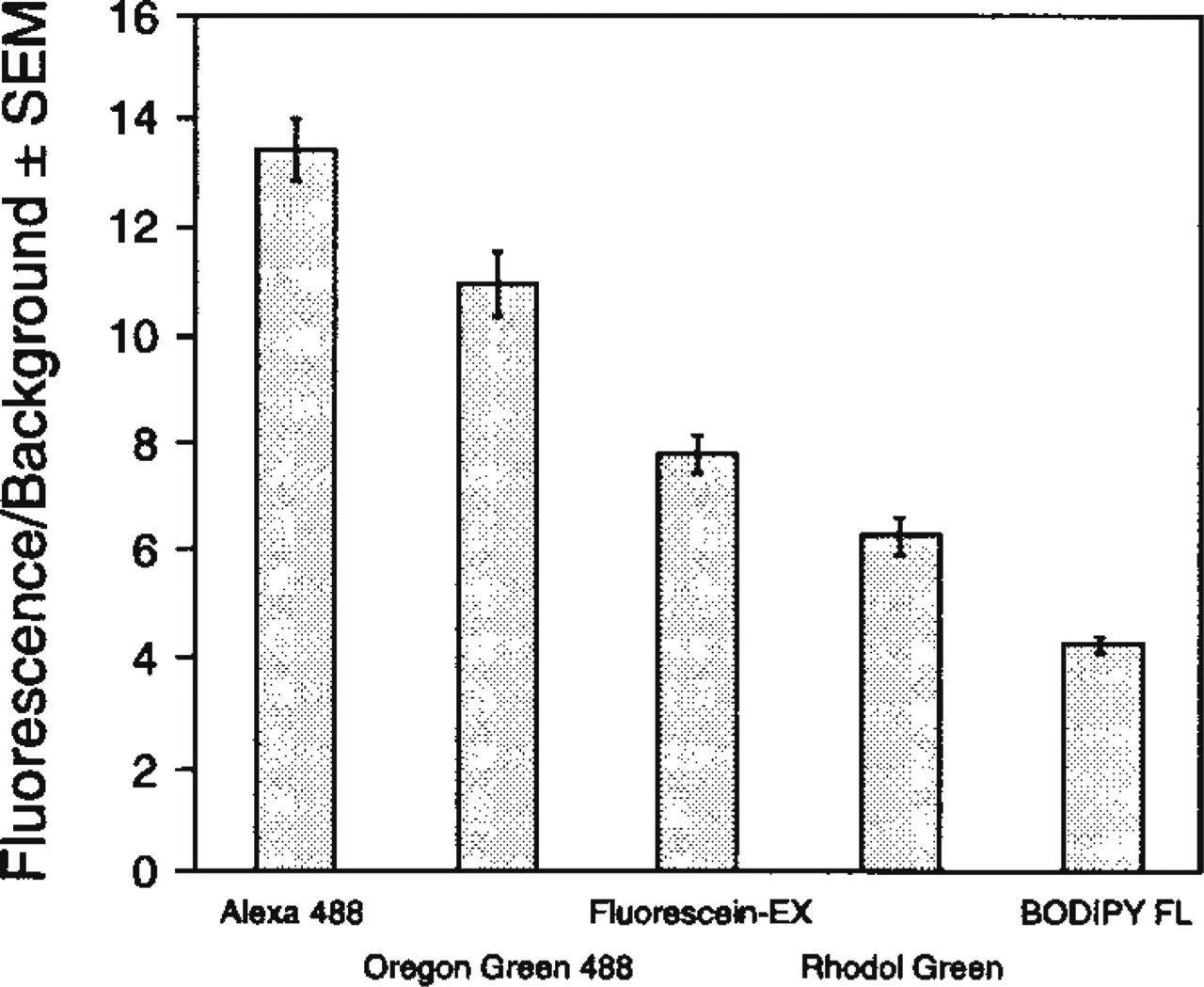
Streptavidin conjugates of the Alexa 488, Oregon Green 488, fluorescein-EX, and BODIPY FL dyes, labeled with a similar number of moles of fluorophores per mole of protein, were used to detect by flow cytometry live RAW264.7 cells treated with biotinylated anti-mouse CD16/CD32 antibody.
Effect of mounting medium on brightness and photostabilty of specific cell labeling with Alexa IgG conjugates; brightness is depicted as initial intensity value (0 sec), then as a percentage of the initial intensity at 10 and 90 sec
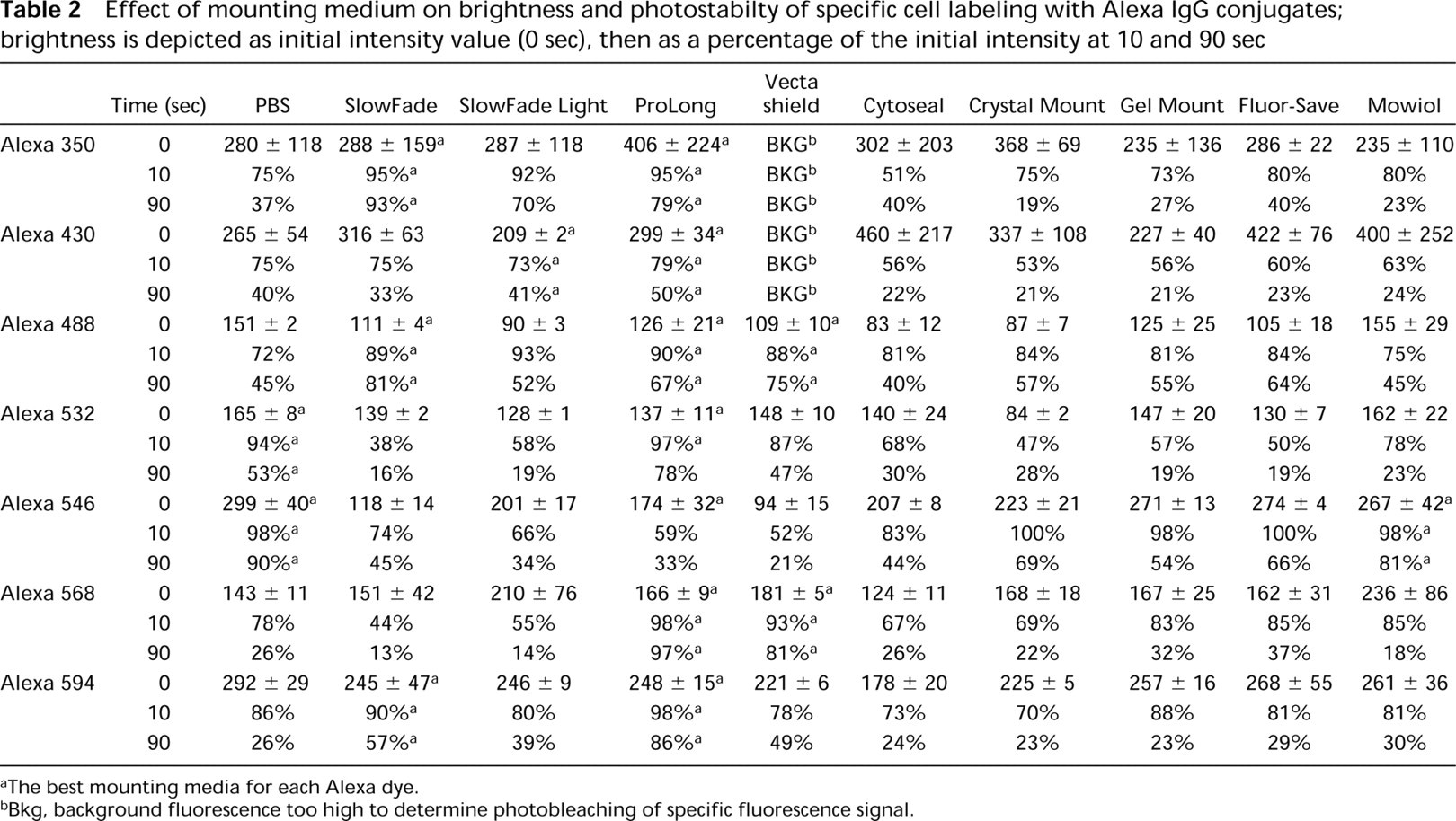
The best mounting media for each Alexa dye.
Bkg, background fluorescence too high to determine photobleaching of specific fluorescence signal.
The remarkable difference in photostability between the Alexa 488 dye and that of fluorescein, as well as the potential utility of this property, is illustrated in Figure 7. Two discrete intracellular targets, actin bundles and microtubules, were labeled with fluoresceinphalloidin and mouse anti-α-tubulin MAb followed by Alexa 488-GAM, respectively. When observed with a typical filter set optimized for fluorescein conjugates, both the actin bundles and the tubulin filaments have bright green fluorescence and the structures cannot be visually distinguished (Figure 7A). However, the fluorescence of the fluorescein-phalloidin rapidly photobleaches, leaving only the Alexa 488-GAM-stained tubulin filaments visible (Figure 7D). Using pixel-by-pixel measurements and pseudocoloring of the actin bundles in red, one can use a single optical filter set and simple image processing techniques to reconstruct the staining pattern of the actin bundles (Figure 7E). This technique permits resolution of both the actin and the tubulin staining (Figure 7F), thus doubling the number of targets that can normally be discriminated by exciting with a single filter set when conventional approaches are used. Similar results were obtained by reversing the staining pattern, i.e., by visualizing actin with Alexa 488-phalloidin and tubulin with fluorescein-GAM conjugate (not shown). Time-resolved fluorescence photobleaching measurements of Alexa dyes and similarly colored second (or even third) dyes that have faster photobleaching rates but similar spectra should be very useful for simultaneous multicolor techniques in which the targets are spatially resolved, such as fluorescence in situ hybridization or detection on DNA chips or Western blots. In particular, combination of the Alexa 488 dye with fluorescein should be helpful when the argon-ion laser is used as the excitation source.
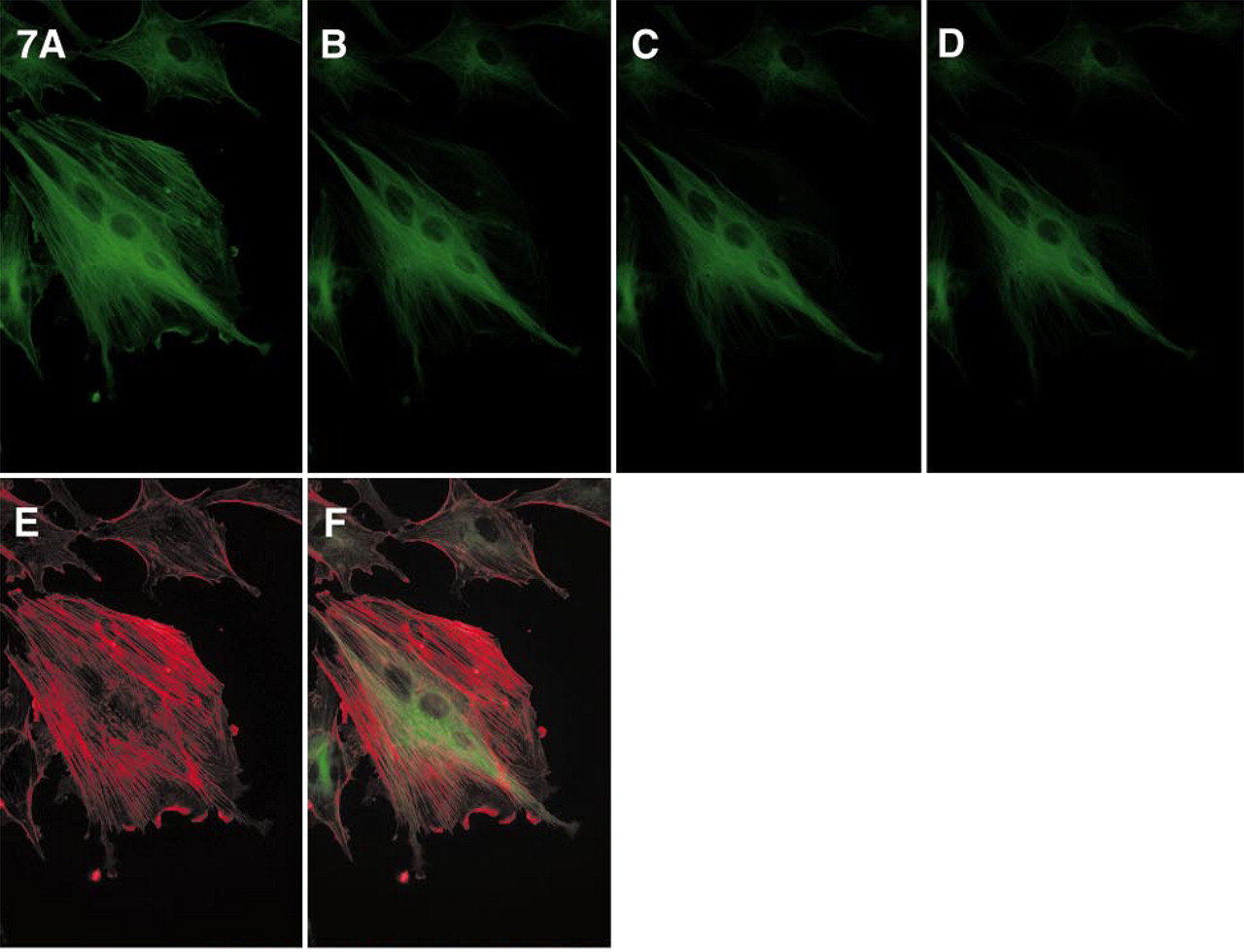
Simultaneous imaging of F-actin (stained with fluorescein-phalloidin) and tubulin (stained with Alexa 488-GAM) using single excitation/emission, based on the difference in photobleaching rates of the Alexa 488 and fluorescein dyes. The samples were imaged with a Quantix CCD camera as described previously, using a Nikon Diaphot TMD microscope equipped with an ex 485 ± 11-nm/em 530 ± 15-nm (XF22) optical filter set to continuously illuminate the specimen.
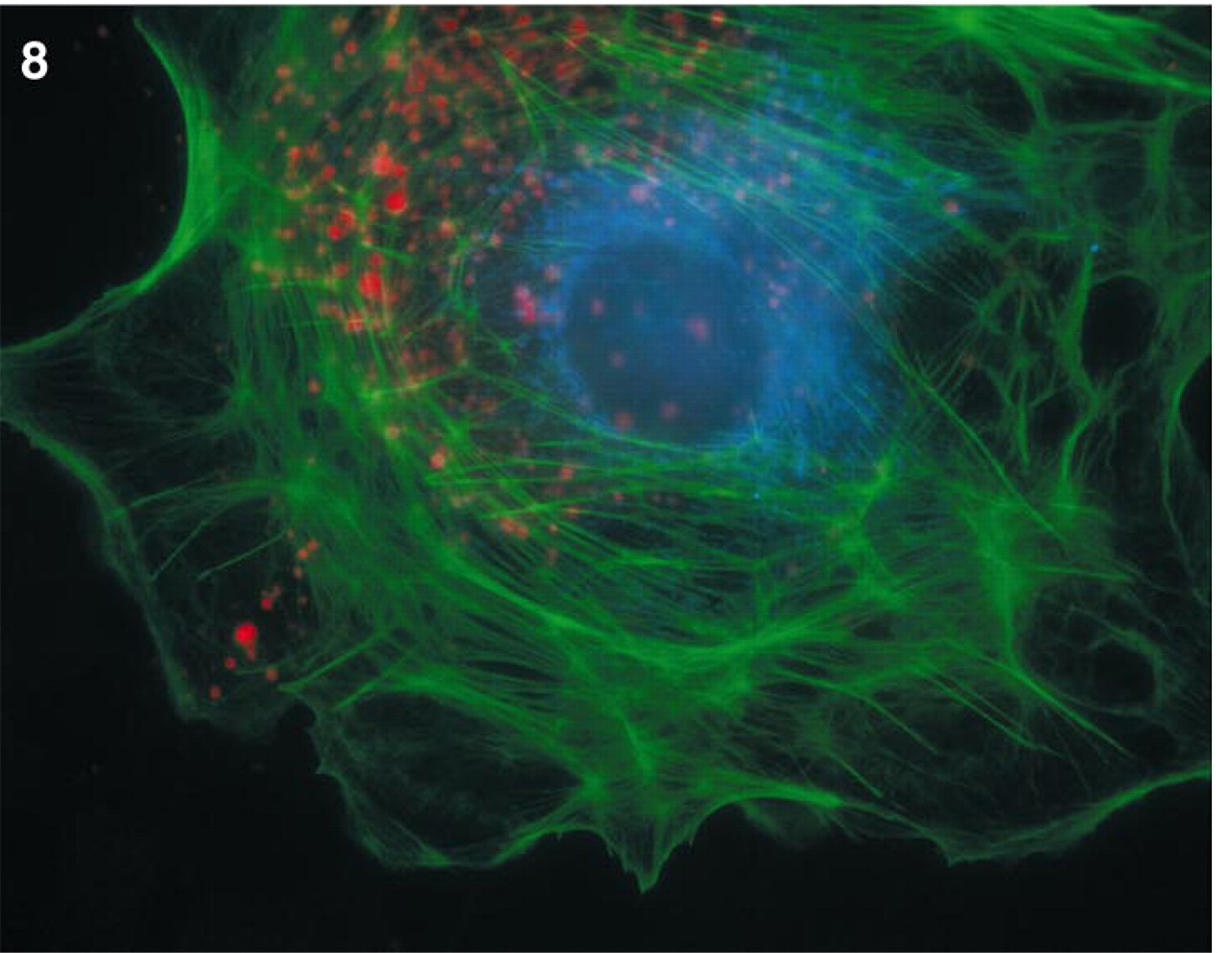
Living BPAE cells stained for lysosomes with Alexa 594 hydrazide and after fixation for mitochondria with the Alexa 350 conjugate of GAM as described in Materials and Methods and for actin with Alexa 488-phalloidin. Images were acquired using a Quantix CCD camera, as described previously, using a Nikon Diaphot TMD microscope and MetaMorph software. With the exception of filter XF 43 (ex 580 ± 14 nm/em 630 ± 15 nm), which is used for Alexa 594 hydrazide, all other filters are the same as described in Table 2.
From the results shown in Table 2, it is clear that antifade reagents can be used to decrease loss of fluorescence of the Alexa dyes through photobleaching but that no single antifade reagent is ideal for all Alexa dyes. In particular, it is noteworthy that the background signal generated by Alexa 350 conjugates was quite high in many of these antifade reagents, to the extent that the rate of photobleaching of specific cell staining could not be determined, because of high background fluorescence (indicated as “Bkg” in the table). ProLong antifade medium was relatively effective as a photoprotectant for all but one Alexa 546 conjugates, which were markedly less bright in this medium than in any other medium tested. The performance of Clarion and Aqua-Poly Mount media was inferior to that of the other antifade agents that we tested. Therefore, those two media were not included in Table 2. Immuno-mount yielded results similar to Gel Mount and was also omitted from the table.
Various combinations of Alexa dyes can be used for multicolor imaging. One example is shown in Figure 8, in which lysosomes of BPAE cells were first loaded by pinocytosis of the red fluorescent Alexa 594 hydrazide. After fixation of the hydrazide with formaldehyde, actin filaments of the cells were subsequently stained with the green fluorescent Alexa 488-phalloidin and mitochondria with a blue fluorescent Alexa 350-GAM conjugate.
The performance of Alexa dye conjugates is superior to that of the commonly used dyes in most applications. However, like fluorescein, Cy3, and Cy5, Alexa dyes carry a net negative charge. In some cases the negative charge of fluorophores may cause a nonspecific electrostatic interaction with positively charged cell structures (Mahmudi-Azer et al. 1998). In these few cases, conventional neutral dyes, such as BODIPY FL, rhodamine, or Texas Red, may still be the proper choice, notwithstanding their lower fluorescence output.
In summary, we have developed a series of sulfonated coumarin and sulfonated rhodamine dyes that we term Alexa dyes, with spectra that cover the entire visible range. Conjugates of these dyes tend to be brighter and more resistant to photobleaching than those of any alternative dyes that have similar spectral properties. The water solubility of their succinimidyl esters permits conjugations without addition of organic solvents, and their protein conjugates tend to be more resistant to precipitation than alternative dyes such as rhodamines and Cy3.