
Abstract
Using a high electron resolution staining method, cationic colloidal gold (CCG, pH 1.0) staining, we studied the fine structural localization of sulfated glycosaminoglycans (GAGs) in various maturational stages of guinea pig neutrophils. Azurophil and specific granules of neutrophils reacted positively to CCG, with variety in labeling according to maturation. All immature azurophil and specific granules were labeled selectively. Mature granules lost their affinity with CCG. CCG-positive labeling was also observed in the trans to trans-most Golgi apparatus of promyelocytes and myelocytes. Prior absorption with poly-L-lysine prevented CCG labeling of tissue sections. Mild methylation of ultrathin sections at 37C did not alter CCG labeling, whereas CCG labeling disappeared after active methylation at 60C. Treatment with chondroitinase ABC or heparinase I abolished the majority of CCG labeling. These findings suggest the existence of sulfated GAGs not only in immature azurophil but also in immature specific granules of neutrophils. Sulfation of GAGs occurs in the trans- to trans-most Golgi apparatus of neutrophil granulocytes. A possible correlation between accumulation of sulfated GAGs and maturation of specific granules in neutrophils is also discussed.
Keywords
T
By using cationic colloidal gold (CCG) at pH 1.0, a recently developed method for demonstrating sulfated glycoconjugates with high electron resolution (Skutelsky and Roth 1986; Kashio et al. 1992; Yang et al. 1996), we have been able to clearly show the subcellular distribution of sulfated GAGs at different maturational stages of guinea pig basophils and eosinophils (Yang et al. 1998a,b). In those studies, not only was the ultrastructure of immature and mature granules of basophils and eosinophils well preserved but also the CCG reactive sites were distinctly visualized in relation to cell organelles. We have also described the relationship between sulfated GAGs and granule biogenesis in these two types of granulocytes (Yang et al. 1998a,b).
Several studies have revealed that specific granules of neutrophils also contain many kinds of basic substances such as lysozyme and lactoferrin (Olsson 1969; Payne and Ackerman 1977; Parmley et al. 1979; Sanchez et al. 1992; Lonnerdal and Iyer 1995; Wu et al. 1995). Other novel cationic antibacterial polypeptides in specific granules of neutrophils have been investigated (Nagaoka et al. 1992; Yamashita et al. 1995; Yomogida et al. 1997). Lactoferrin can bind to anionic polysaccharides and its affinity for sulfated GAGs as well as the GAG-binding region of lactoferrin has recently been characterized (Mann et al. 1994; Wu et al. 1995). A recently purified cationic antibacterial polypeptide (CAP 11) from guinea pig neutrophil granules can also bind to negatively charged GAGs because of its basic nature (Yomogida et al. 1997). Such interactions between sulfated GAGs and basic substances of specific granules raise the interesting possibility that sulfated GAGs might also exist in specific granules of neutrophils.
To demonstrate sulfated GAGs in granules of neutrophils, we used Lowicryl K4M-embedded guinea pig bone marrow and the CCG (pH 1.0) staining method in this study. We observed CCG-positive labeling in immature azurophil granules, immature specific granules, and trans- to trans-most Golgi lamellae of granulocytes of premitotic maturational stages. Sulfated GAGs in the cell surface and bone marrow matrix were also revealed. By adding GAGs degrading enzymes and using other control experiments, we tried to analyze the nature of sulfated GAGs of neutrophils. A possible correlation between accumulation of sulfated GAGs and maturation of granules in neutrophils is also discussed.
Materials and Methods
Preparation of Specimens
Three young guinea pigs weighing about 400 g each were used. The animals were anesthetized with ether. Bone marrow was removed from the femur, fixed in half-strength Karnovsky's fixative, and embedded in Lowicryl K4M (Yang et al. 1996). Two-μm-thick semithin sections were cut with an LKB ultratome, stained with 0.1% toluidine blue solution dissolved in 1% borax, and examined by light microscopy to determine the appropriate area of tissue for ultrathin sectioning. Ultrathin sections were cut with an Ultracut N ultratome (Reichert-Jung; Vienna, Austria) and placed on Formvar-coated single-hole nickel grids (Yang et al. 1998a,b).
Ultracytochemical Labeling Using CCG at pH 1.0
Cationic colloidal gold (CCG 10 nm) was purchased from British BioCell International (Cardiff, UK). Ultrathin sections designed for CCG labeling were pretreated with CCG buffer (potassium chloride-HCl buffer, pH 1.0) for 20 min and then labeled with 10 X CCG solution (diluted in the above buffer at pH 1.0) at room temperature (RT) overnight (Yang et al. 1998a,b). After labeling, sections were jet-washed in the same buffer and then washed twice in distilled water for 5 min. Then the sections were counterstained with 2% aqueous uranyl acetate for 3 min and Millonig's lead acetate for 1 min.
Control Experiments
Preabsorption with Poly-L-lysine. Sections were incubated with 10 mg/ml poly-L-lysine (MW 682,600; Sigma, St Louis, MO) solution (dissolved in potassium chloride-HCl buffer, pH 1.0) for 4 hr at RT before labeling with CCG at pH 1.0.
Mild and Active Methylation. CCG labeling was performed after methylation of ultrathin sections with 0.01 N HCl-methanol for 4 hr at 37C (mild methylation) and at 60C (active methylation).
Enzyme Degradation. Sections were incubated in either of the following enzymes: (a) 1 U/ml heparinase I (Flavobacterium heparinum, EC 4.2.2.8; Sigma) dissolved in Tris-buffered saline (TBS), pH 7.0, containing 0.04 M calcium chloride, or (b) 1 U/ml protease-free chondroitinase ABC (Proteus vulgaris, EC 4.2.2.4; Seikagakukogyo, Tokyo, Japan) dissolved in Tris-HCl buffer, pH 8.0. Both incubations were carried out in a moisture chamber at 37C overnight. After incubation, sections were washed three times with the same buffer for 5 min and then with distilled water. Finally, sections were labeled with CCG as previously described. All sections were observed with a Hitachi H7100 electron microscope at 75 kV accelerating voltage.
Quantitation of CCG Labeling
The CCG labeling density of neutrophil granules was calculated by using a Macintosh computer. Electron micrographic negatives (X 10,000) were scanned into the computer with a Nikon Film Scanner. For each experiment and at each stage, at least five neutrophil granulocytes with 30-50 granules were used. After the scale was set, the granule areas (Ai) were measured by NIH Image software. The number of gold particles (Ni) was counted manually. The mean density of gold particles was calculated as =$Ni/$Ai (number/μm2). One-way analysis of variance (ANOVA) was used to analyze the labeling difference between granulocytes with or without treatment with enzymes. Student's t-test was used to analyze the labeling difference between promyelocytes and myelocytes. A p value of less than 0.05 was considered significant.
Results
We divided the development of neutrophil granulocytes into promyelocyte, myelocyte, band form (including metamyelocyte), and segmented neutrophils according to the methods of earlier investigators (Bainton and Farquhar 1966; Wetzel et al. 1967; Bainton et al. 1971; Brederoo and Daems 1978). For convenience of description, we describe only azurophil and specific granules of neutrophils in this study.
Promyelocytes
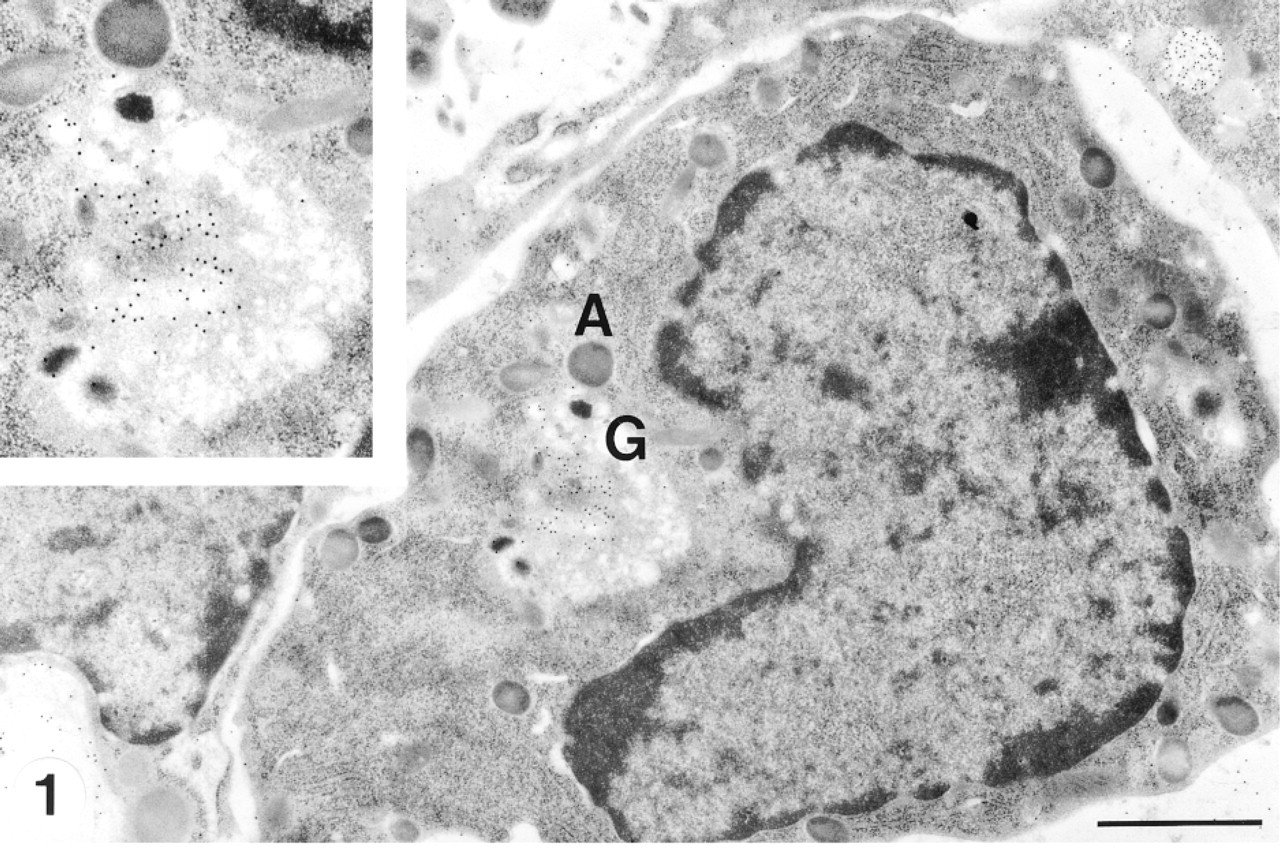
A very early promyelocyte has an irregular nucleus and sparse cytoplasm with few granules. The Golgi complex (G) is situated near the concave face of the nucleus. (
Promyelocytes are the largest cells of the neutrophil series and represent the first recognizable stage in development. The nucleus shows a dispersed euchromatin pattern and a small peripheral zone of moderately condensed heterochromatin (Figures 1 and 2). Sometimes one or two nucleoli can be observed in an ultrathin section (Figure 2). The Golgi complex is situated at the concave side of the nucleus (Figures 1 and 2). Azurophil granules at different developmental stages varied in size and electron density.
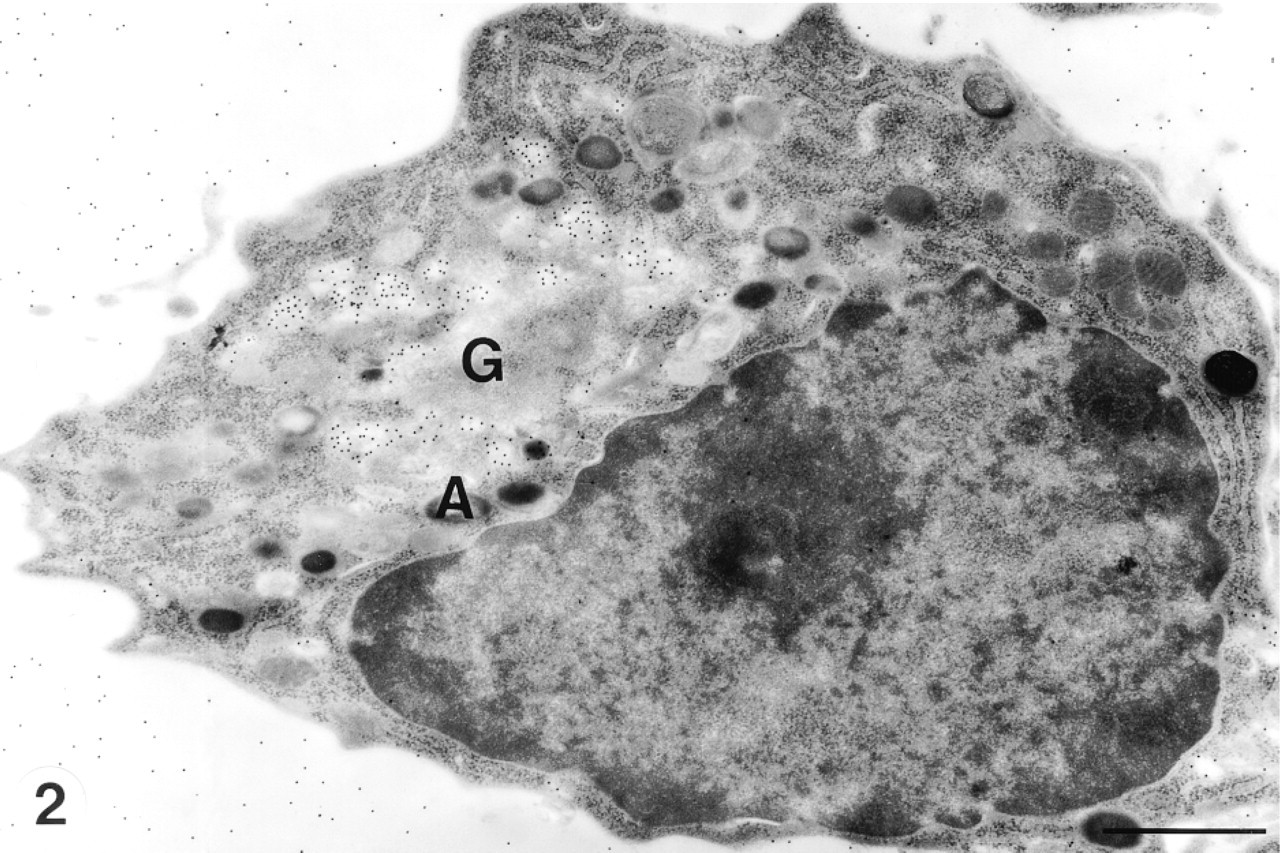
A neutrophil promyelocyte contains an irregular nucleus with a nucleolus. An active Golgi complex (G) is situated near the nucleus. The trans- to trans-most Golgi lamellae and electron-lucent immature azurophil granules are strongly labeled by CCG (pH 1.0). Mature electron-dense azurophil granules are seen in the peripheral cytoplasm and are not labeled with CCG. CCG labeling in bone marrow matrix indicates the presence of GAGs in it. A, azurophil granule. Bar = 1.0 μm.
CCG labeling was observed in the trans- to transmost Golgi apparatus, small vesicles in the Golgi area, and immature azurophil granules (Figures 1 and 2). The immature azurophil granules were electron-lucent. They were spherical or ellipsoid and were present around the Golgi area and elsewhere in the cytoplasm (Figure 2). Mature azurophil granules were highly electron-dense. They were present throughout the cytoplasm and showed decreased or absent labeling of CCG (Figures 1 and 2). Sulfated GAGs in the cell surface and bone marrow matrix were also labeled with CCG (Figures 1 and 2).
Myelocytes
Myelocytes were smaller than promyelocytes, with a round or elongated shape. The nucleus contained clumped and peripherally condensed heterochromatin. Both azurophil and specific granules were present at this stage. Immature azurophil granules had an electron lucent periphery with a somewhat dense core, whereas mature ones were electron-dense (Figure 3). Immature specific granules showed irregular flocculated contents, whereas mature ones were electron-lucent with a moderately electron-dense periphery (Figure 4). The Golgi apparatus had some stacks of cisternae. Small vesicles arising from the convex face of the Golgi apparatus indicated active specific granulogenesis. At this maturational stage, CCG positively labeled the trans- to trans-most Golgi apparatus, small vesicles in the Golgi area, immature azurophils, and immature specific granules (Figures 3 and 4).
Band Form (Including Metamyelocytes)
Because it is difficult to differentiate metamyelocytes and band form neutrophils in ultrathin sections, they are described together. These cells possessed variably shaped nuclei with peripherally condensed heterochromatin. The Golgi apparatus was relatively small and no immature, aggregating granules were associated with it. Negligible CCG labeling was observed at this stage (Figure 5). Both azurophil and specific granules were present, the latter outnumbering the former.
Segmented Neutrophils
The segmented neutrophil is the smallest cell of the neutrophil series. It is characterized by a segmented nucleus. The Golgi apparatus is small, with no granulogenesis. No CCG labeling was observed at this stage.
Control Experiments
Preabsorption with poly-L-lysine abolished CCG labeling at all stages of neutrophils. CCG labeling remained unchanged after mild methylation at 37C but disappeared after active methylation at 60C (Figure 6). Treatment with chondroitinase ABC reduced CCG binding by about 92%, and treatment with heparinase I reduced CCG binding by about 97% in cells at all maturational stages (Figures 7 and 8; Table 1). The number of CCG labeled gold particles was greater in promyelocytes than that in myelocytes in sections not treated with GAG-degrading enzymes (Table 1).
Discussion
In the past few decades, histochemical and other cytochemical studies pointed out that sulfated GAGs exist in the azurophil granules and Golgi apparatus but not in specific granules of neutrophils (Horn and Spicer 1964; Dunn and Spicer 1969; Hardin and Spicer 1971; Murata and Spicer 1973; Payne and Ackerman 1977; Spicer et al. 1978; Murata et al. 1979, 1992; Parmley et al. 1980, 1983, 1986). However, these methods could not differentiate the reactive sites in relation to granule ultrastructure owing to the relatively low electron resolution. By using CCG at pH 1.0, a recently developed method for specific staining of sulfated GAGs with high electron resolution, it has become possible to demonstrate clearly the CCG-reactive sites with cell organelles (Yang et al. 1996, 1998a,b). In this study, we observed positive CCG labeling in immature azurophil granules but not in mature ones of neutrophils by virtue of the high electron resolution of CCG staining. This result corresponds with the observations of Parmley et al. (1980, 1983). The existence of GAGs in the Golgi apparatus and the involvement of the Golgi apparatus in the formation of secretory granules have been documented (Spicer et al. 1978; Murata et al. 1979, 1992; Parmley et al. 1980, 1983). We observed more precisely the positive CCG labeling in the trans- to trans-most Golgi apparatus of promyelocytes and myelocytes with high resolution. This indicates that sulfation of GAGs of neutrophil granules also takes place in the trans- to trans-most Golgi apparatus as in many other types of cells (Young 1973; Yang et al. 1996, 1998a, b).
It is fascinating to note the positive CCG labeling in the immature specific granules of neutrophils. This indicates that the content of specific granules is negatively charged, probably due to the presence of sulfated GAGs. This result agrees with reports that suggest that GAGs are present in smaller quantities in secondary lysosomal granules (Olsson 1969; Payne and Ackerman 1977; Parmley et al. 1979; Kolset and Gallagher 1990). The coexistence of sulfated GAGs and other basic substances, such as lysozyme and elastase, in azurophil granules has been well documented (Olsson and Gardell 1967; Murata 1974). It is well known that specific granules of neutrophils also contain many kinds of basic substances such as lysozyme, lactoferrin (Olsson 1969; Payne and Ackerman 1977; Parmley et al. 1979; Sanchez et al. 1992; Lonnerdal and Iyer 1995), and other novel cationic antibacterial polypeptides (Nagaoka et al. 1992; Yamashita et al. 1995; Yomogida et al. 1997). Like other basic substances, lactoferrin binds to polyanionic macromolecules, including sulfated GAGs (Sanchez et al. 1992; Wu et al. 1995). The putative GAG binding region of lactoferrin has recently been characterized to be localized in the N-terminus (Zou et al. 1992; Mann et al. 1994; Wu et al. 1995). An intriguing feature of the crystallographic structure of lactoferrin is that a large number of basic amino acids are surface-exposed near the N-terminus (Anderson et al. 1990; Wu et al. 1995). A recently purified cationic antibacterial polypeptide (CAP 11) from guinea pig neutrophil granules can also bind to negatively charged GAGs because of its basic nature (Yomogida et al. 1997). Because specific granules contain so many kinds of basic substances that can interact with GAGs, it is reasonable to assume that GAGs coexist with lactoferrin and other basic substances in the specific granules of neutrophils for neutralization. This idea was first suggested two decades ago (Olsson 1969; Payne and Ackerman 1977; Parmley et al. 1979; Kolset and Gallagher 1990). Our study is the first report to give direct histochemical evidence to support it.
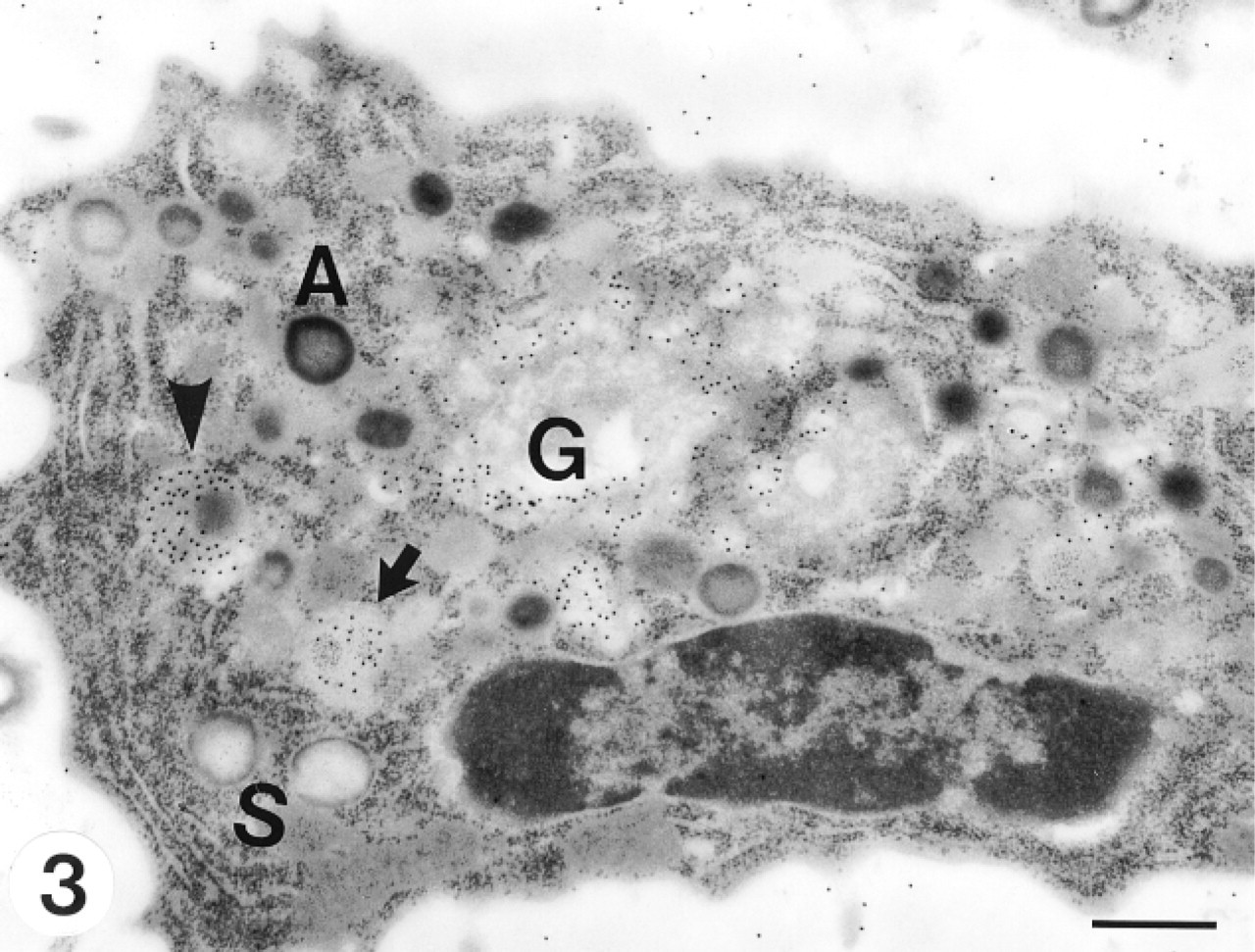
A neutrophil myelocyte is elongated, with dense peripheral heterochromatin in its nucleus. It contains both azurophil and specific granules at various stages of development. Small vesicles arising from the convex face of the Golgi apparatus indicate active specific granulogenesis. Immature specific granules show irregular flocculated contents (arrow). CCG positively labeled the trans- to trans-most Golgi apparatus, small vesicles in the Golgi area, immature azurophil (arrowhead) and immature specific granules (arrow). G, Golgi complex; A, azurophil granule; S, specific granules. Bar = 0.5 μm.
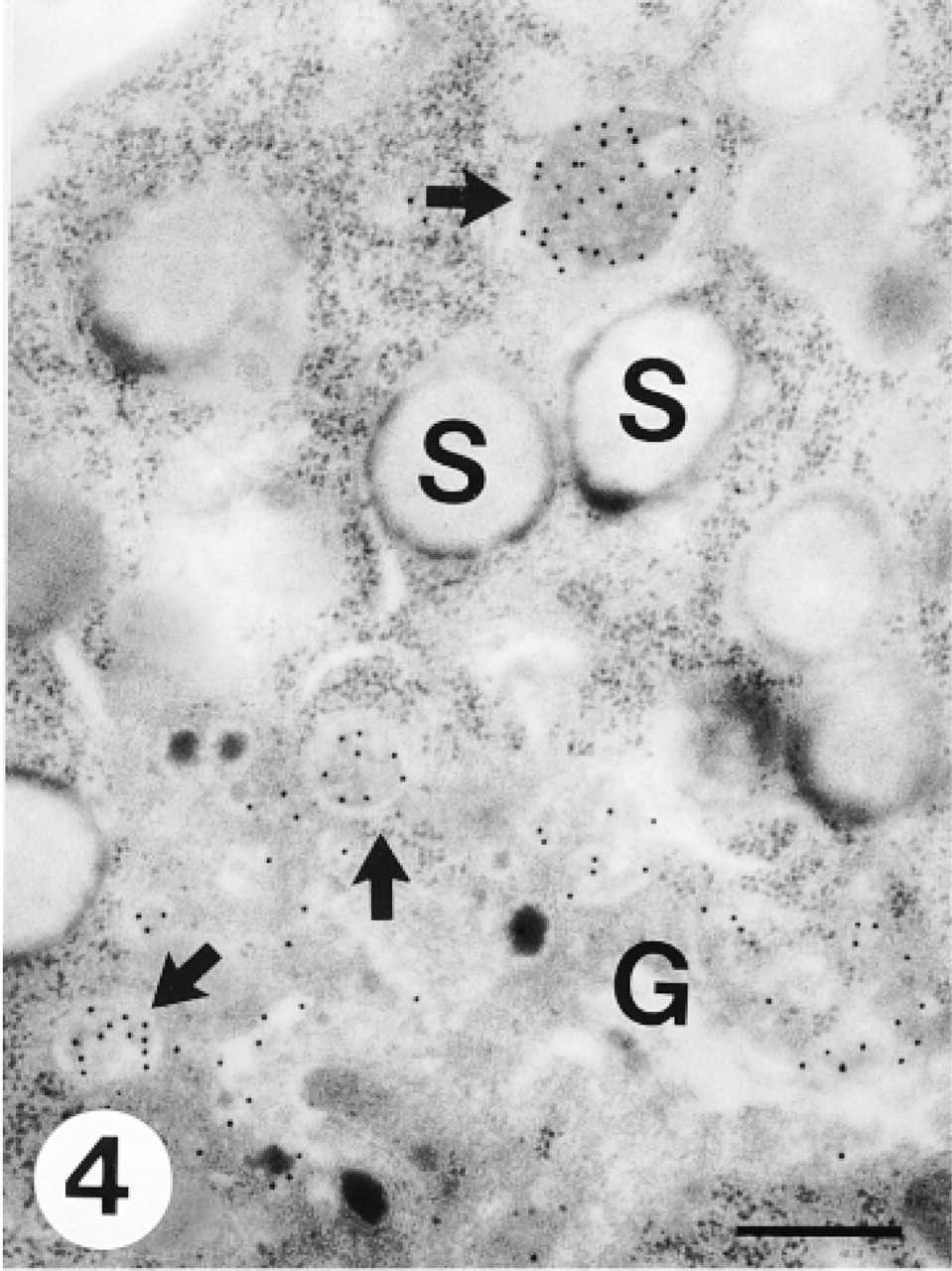
A portion of a myelocyte containing many specific granules. The Golgi complex (G) is small but is still involved in granulogenesis. CCG labeling is observed in the trans- to trans-most Golgi apparatus and immature specific granules with flocculated contents (arrows), but not in mature specific granules (S). Bar = 0.36 μm.
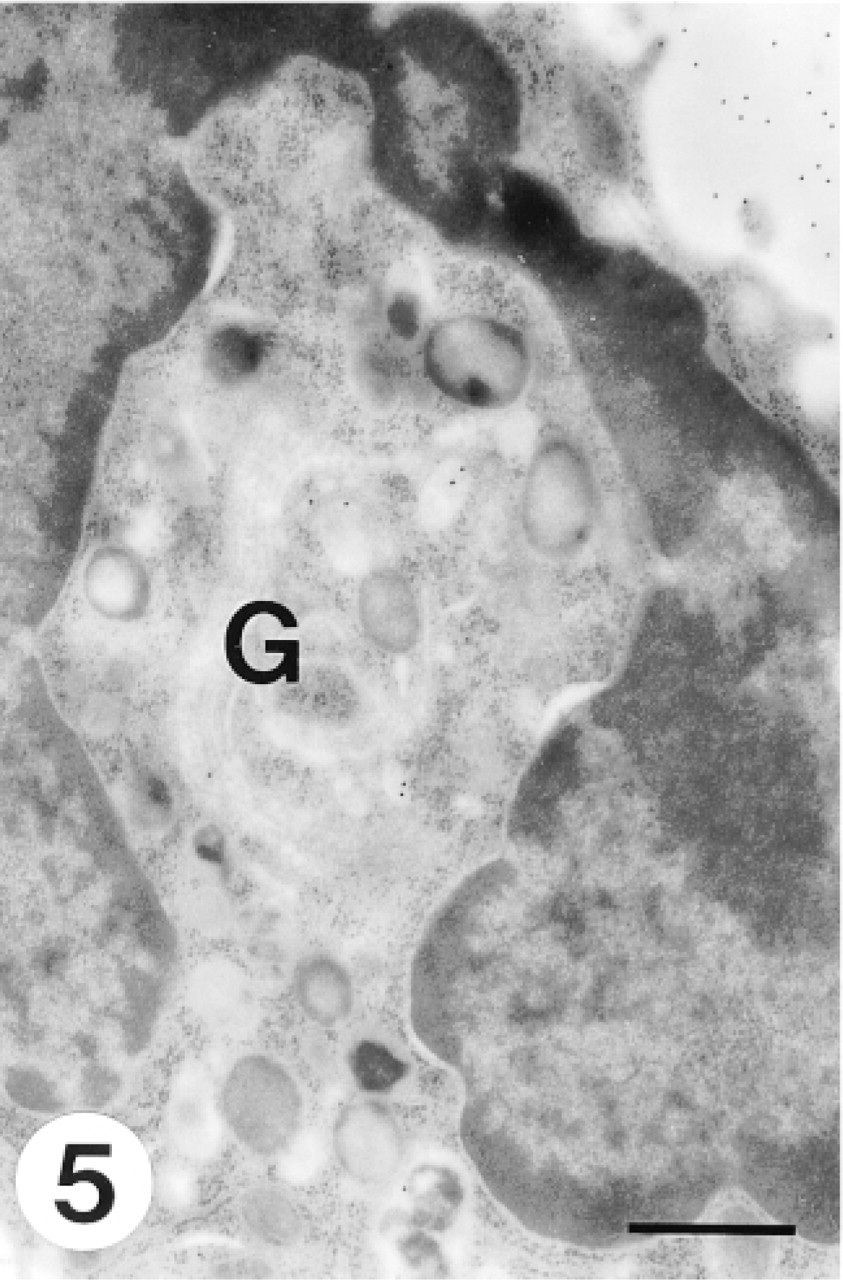
A band form neutrophil shows a characteristic horseshoe-shaped nucleus. The Golgi apparatus is small and no immature aggregating granules are associated with it. Negligible CCG labeling was observed in the Golgi region. Bar = 0.5 μm.
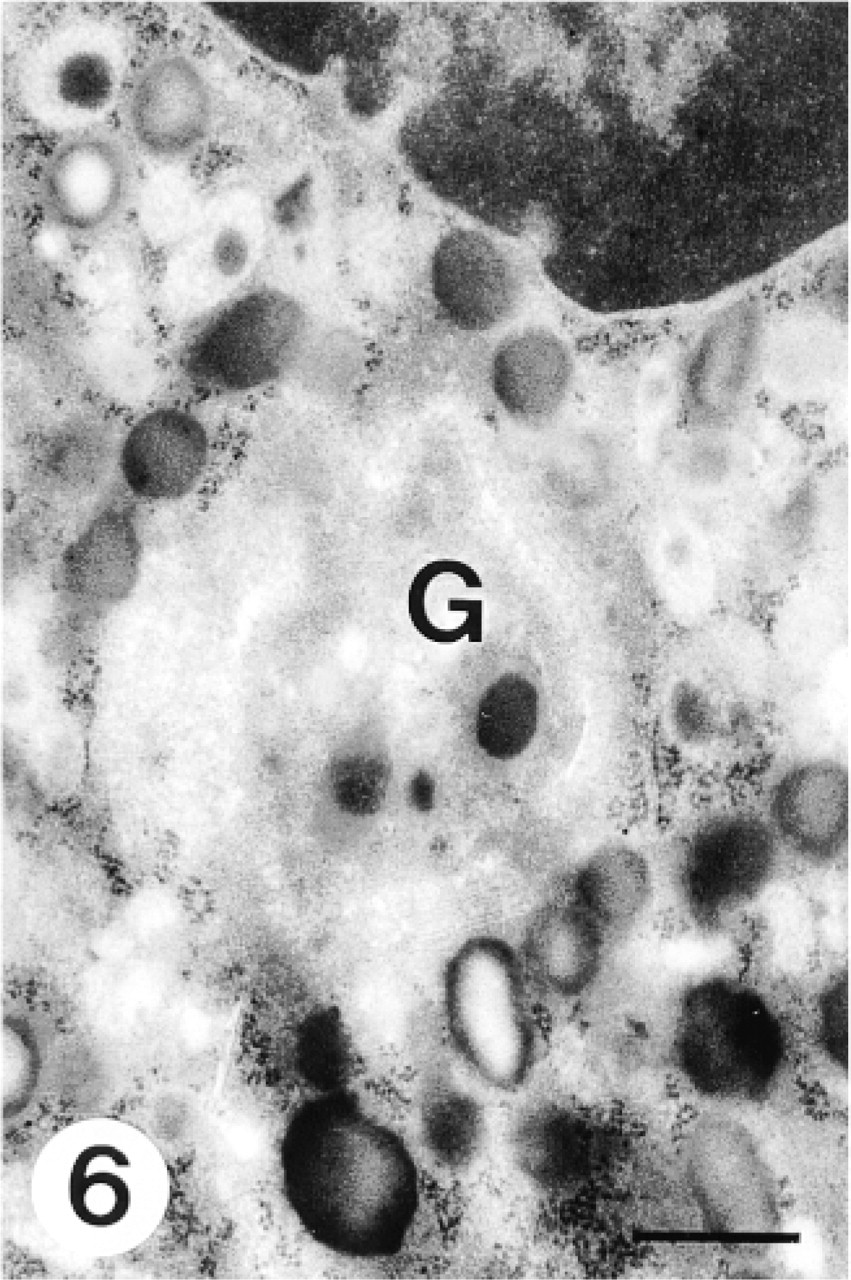
This micrograph shows that CCG labeling in the Golgi complex (G) of a myelocyte disappears after active methylation at 60C. Bar = 0.5 μm.
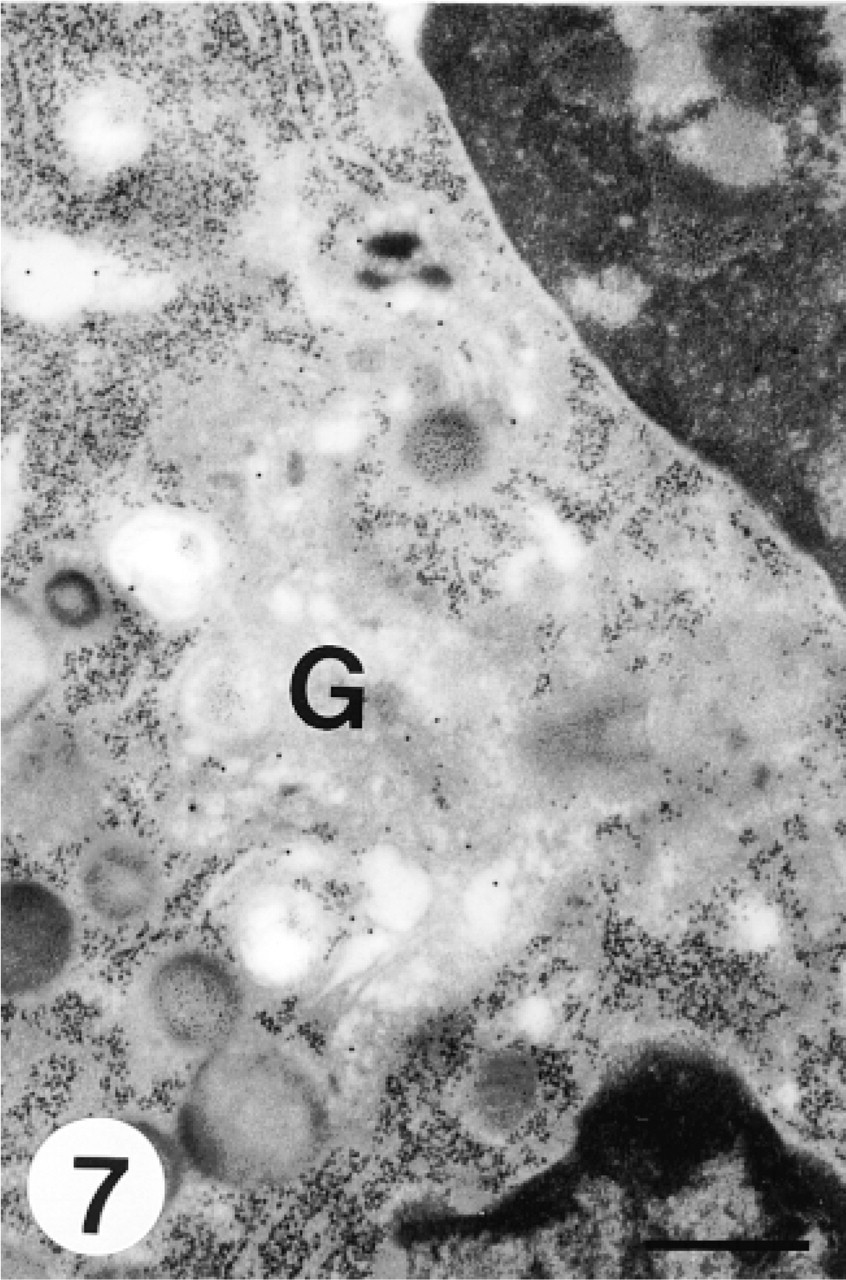
This micrograph shows negligible labeling in the Golgi region (G) of a myelocyte after heparinase I treatment. Bar = 0.5 μm.
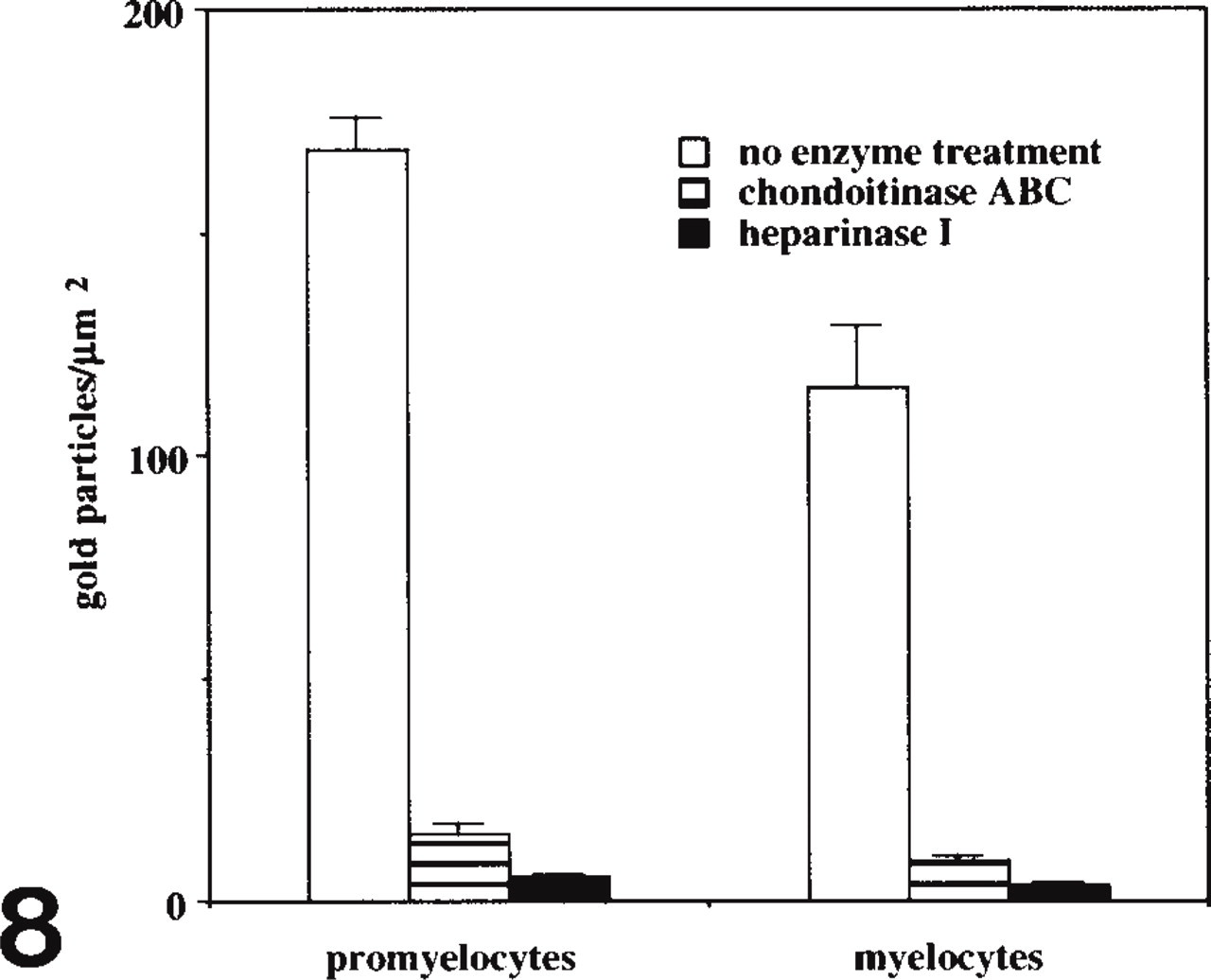
Histogram showing the effect of treatment with GAG-degrading enzymes on the labeling of CCG in promyelocytes and myelocytes.
GAGs in azurophil granules function by complexing with certain basic components and holding them inactive to facilitate granule storage (Hardin and Spicer 1971; Olsson 1971; Parmley et al. 1980, 1983). The loss of cationic staining in mature azurophil granules might be due to the masking of reactive sites or the decrease of concentration in the granules after intracytoplasmic processing (Parmley et al. 1980, 1983). The physiological significance of the binding of GAGs to lactoferrin and other cationic antibacterial peptides in specific granules is unclear. It is reported that proteoglycan sulfation and storage parallel storage of basic secretory proteins (Blair et al. 1991). We assumed that, like the situation with azurophil granules, GAGs in specific granules might also provide a negatively charged matrix to bind cationic proteins present in the granules, thus facilitating the biogenesis and maturation of granules. A recent study points out that the mRNA of antibacterial cationic peptide was expressed only in neutrophil precursor cells, such as promyelocytes and myelocytes, but was not detected in the mature neutrophils (Nagaoka et al. 1992). Further studies will be necessary to determine whether the loss of cationic staining in mature specific granules and mature neutrophils is due to the masking of reactive sites or to the loss of GAGs in mature granules and cells.
Previous biochemical studies have determined that the major GAGs of neutrophils are mainly chondroitin-4-sulfate (Olsson and Gardell 1967; Murata 1974; Kolset and Gallagher 1990; Kjellén and Lindahl 1991). That preabsorption with poly-L-lysine prevented CCG labeling suggests that the polyanionic nature of the GAGs is important for CCG binding. CCG labeling did not change after mild methylation at 37C but disappeared after active methylation at 60C. This indicates that the positive CCG reaction products are sulfated GAGs (Yang et al. 1998b). The observation that treatment with chondroitinase ABC and heparinase I abolished the same percentage of CCG binding in promyelocytes and myelocytes suggests that the content of GAGs in specific granules is almost the same as that in azurophil granules. Because chondroitinase ABC degraded chondroitin sulfate and dermatan sulfate, and heparinase I degraded chondroitin sulfate, dermatan sulfate, heparan sulfate, and heparin (Orenstein et al. 1978), the content of GAGs might include chondroitin sulfate, dermatan sulfate, and heparan sulfate. Studies using antibodies specific for special types of GAGs will be necessary to differentiate the types of GAGs in the granules.
Labeling density of gold particles per square micrometer (mean ± SEM) in granules of promyelocytes and myelocytes with or without treatment with GAG-degrading enzymes a

aParentheses indicate the percentage decrease after enzyme treatment.
∗ p<0.001.
∗∗ p<0.05.
The existence of GAGs in specific granules of neutrophils is a new finding. It has not been previously reported, probably owing to the limitations of methodology in previous studies. Further studies, e.g., double staining by using CCG and immunogold with monoclonal antibodies against GAGs or lactoferrin, will provide more interesting evidence for this idea.