
Abstract
We describe the synthesis of Bodipy 558/568-modified acetazolamide, a fluorescent inhibitor of carbonic anhydrase and its use to localize the enzyme in living cells. The modified acetazolamide, with its specific sulfonamide group intact, labeled cells at concentrations as low as 10−9 M, with a minimal loading time of 5 min. The staining was decreased by 57.4% by preincubating cells with unaltered acetazolamide (1:100) or with trifluoromethane sulfonamide, 6-ethoxyzolamide, and 5-(3-hydroxybenzoyl)-thiophene-2-sulfonamide. The efficacy of the inhibitor was unchanged by the fluorescent label, as determined by an acridine orange assay that detects acidification of osteoclasts, the cell model used in this study. This compound should prove to be useful for studying carbonic anhydrase in many organisms because of the high degree of conservation of the active site of this enzyme.
T
Osteoclasts are the large multinucleated cells that resorb bone by secreting hydrogen ions derived from the action of carbonic anhydrase (Hunter et al. 1988, 1991; Gay 1996). Carbonic anhydrase, the enzyme that catalyzes the reversible hydration of CO2 to carbonic acid, has been identified in osteoclasts of chickens (Gay and Mueller 1974; Anderson et al. 1982), rats (Väänänen and Parvinen 1983; Marie and Hott 1987; Sundquist et al. 1987), mice (Jilka et al. 1985), and humans (Väänänen 1984). Inhibition of bone resorption by acetazolamide and other inhibitors of carbonic anhydrase has been demonstrated in vivo (Waite et al. 1970; Waite 1972; Conaway et al. 1973; Kenny 1985; Brown et al. 1990) and in vitro, in whole-bone explants (Minkin and Jennings 1972; Mahgoub and Stern 1974; Hall and Kenny 1985a,b,1986,1987; Raisz et al. 1988) and isolated osteoclasts (Hunter et al. 1991). Genetic evidence for carbonic anhydrase II involvement in bone resorption has been identified as an autosomal recessive syndrome of osteopetrosis, a disease characterized by failure of bone resorption (Sly et al. 1983,1985). Given the importance of carbonic anhydrase for osteoclast function, although this enzyme has been localized in osteoclasts by autoradiography, histochemistry, and immunocytochemistry, its demonstration in living cells opens new avenues of research to investigate the intracellular function of this enzyme.
This report presents the labeling of carbonic anhydrase in living osteoclasts with a fluorescent-modified acetazolamide, of which the inhibitory sulfonamide group remains unaltered. This is the first study of carbonic anhydrase localization in living cells. Another fluorescent sulfonamide, 5-dimethylamino-naphthalene-1-sulfonamide (DNSA), has been used to label carbonic anhydrase in fixed tissue sections of rat kidney, skeletal muscle, and thymocytes (Dermietzel et al. 1985). The fluorescent-modified acetazolamide should prove to be a useful tool for studying carbonic anhydrase, because sulfonamides, particularly acetazolamide, have been shown to bind carbonic anhydrases in a wide variety of prokaryotic and eukaryotic organisms, including higher mammals and plants (for reviews see Maren 1967; Bundy 1977). Because the carbonic anhydrase II isoform predominates in many cell types, including osteoclasts, sulfonamide-specific fluorescence will reflect levels of carbonic anhydrase II in specific cell types.
Materials and Methods
Synthesis of Bodipy 558/568-modified Acetazolamide
The fluorescent Bodipy 558/568-modified acetazolamide (AZ-Bd) was synthesized in three steps, as shown in Figure 1. First, acetazolamide (Compound 1) was deacetylated in ethanol-HCl to give Compound 2. The amino group of Compound 2 was then used as the site of dye attachment. To reduce any interference of the dye with the activity of the drug, β-alanine was used as a linker to separate the dye and the drug moiety. Thus, N-t-butoxycarbonyl β-alanine (Compound 3) was conjugated to Compound 2 via the mixed anhydride method to give Compound 4. Deprotection of the amino group of Compound 4 with trifluoroacetic acid and subsequent reaction with the N-hydroxysuccinimidyl (NHS) ester of Bodipy 558/568 propionic acid yielded AZ-Bd in an overall 61% yield. Acetazolamide was purchased from Sigma (St Louis, MO) and Bodipy 558/568 NHS ester was provided by Molecular Probes (Eugene, OR). The synthesis of AZ-Bd was performed at Molecular Probes under the direction of Dr. Fei Mao. All other chemicals were purchased from Aldrich Chemicals (Milwaukee, WI) and were used as received without further purification. Column chromatography was performed using medium grade silica gel (35–75 μm) from EM Science (Gibbstown, NJ) and an appropriate solvent system as indicated for each compound below. The identities of all the products were confirmed by NMR spectroscopy as recorded on a 400-MHz Bruker FT NMR spectrophotometer. Excitation and emission spectra were recorded on a F-4500 Hitachi fluorescence spectrophotometer. The absorption and emission spectra of Bodipy 558/568 did not change after conjugation (data not shown).
Synthesis of Compound 2: 2-amino-1,3,4-thiadiazole-5-sulfonamide Hydrochloride
A suspension of acetazolamide, Compound 1 (3.36 g, 15.1 mmol), was gently refluxed in a mixture of ethanol (40 ml) and concentrated hydrochloric acid (6 ml) at 85C. After 90 min of reaction, the suspension became a clear solution. After an additional 4 hr of reaction, the solution was cooled to room temperature (RT) and allowed to stand overnight. The colorless crystals were collected by filtration and washed with cold water and ethanol. The crystals were dried under house vacuum at 50C for 24 hr to give Compound 2 in 90% yield.
Synthesis of Compound 4: 2-(3-(N-t-butoxycarbonylamino) propionylamino)-1,3,4-thiadiazole-5-sulfonamide
To N-t-butoxycarbonyl-β-alanine, Compound 3 (1.05 g, 5.54 mmol), in 10 ml dry tetrahydrofuran (THF) was added triethylamine (0.93 ml, 6.65 mmol) in one portion. While the solution was stirred at 0C in an ice-water bath, isobutyl chloroformate (0.86 ml, 6.65 mmol) was added dropwise under nitrogen. The solution was stirred at 0C for 1 hr, then placed at RT for 10 min, followed by cooling to 0C again. Compound 2 (1.0 g, 4.6 mmol) in 4 ml dry THF was added dropwise, followed by the addition of triethylamine (0.93 ml, 6.65 mmol) dropwise. The resulting solution was stirred at RT for 24 hr. The solvent was removed by rotary evaporation and the residue was purified by silica gel column chromatography eluting with 1:10 methanol-chloroform to give 1.3 g (80% yield) of Compound 4 (Rf = 0.3).
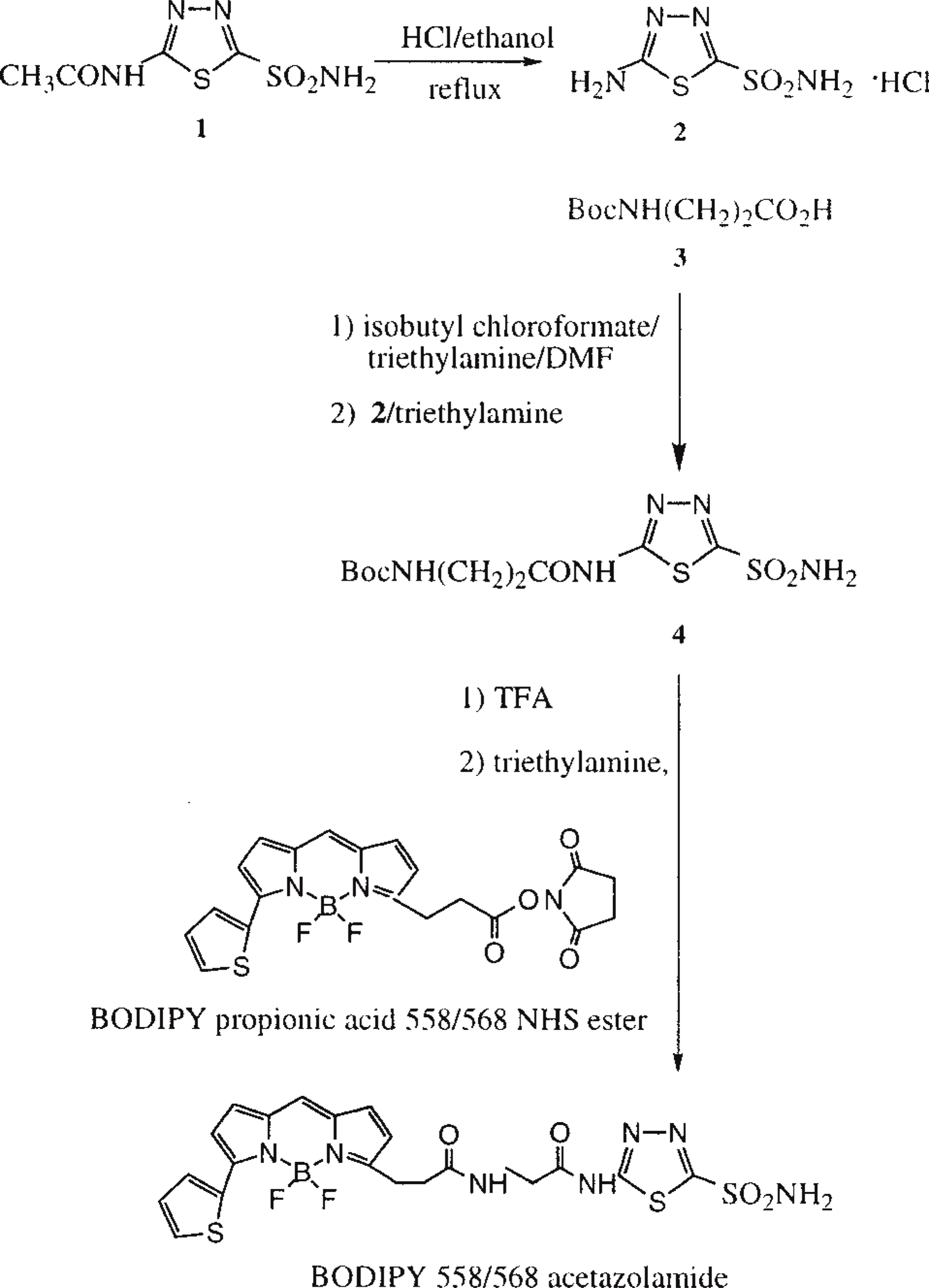
Scheme for the synthesis of Bodipy 558/568-modified acetazolamide (AZ-Bd). Numbers in bold correspond to the compounds whose synthesis is detailed in Materials and Methods.
Synthesis of Bodipy 558/568-modified Acetazolamide
Compound 4 (30 mg, 85.2 μmol) was dissolved in 2 ml of trifluoroacetic acid (TFA) at 0C and the solution was stirred at RT for 10 min. The excess TFA was removed by rotary evaporation and the residue was dried under high vacuum at RT for 3 hr. The solid was redissolved in 10 ml dry THF, then stirred at RT. Triethylamine (160 μl, 1.15 mmol) was added in one portion, followed by dropwise addition of Bodipy propionic 558/568 NHS ester (38 mg, 85.8 μmol) in 5 ml THF. The resulting solution was stirred at RT for 24 hr. The solvent was removed by rotary evaporation and the residue was purified by preparative TLC using ethyl acetate for elution, yielding 42 mg (85%) of Bodipy 558/568-modified acetazolamide. ∗∗∗λex(MeOH or H2O) = 558 nm; λem(MeOH or H2O) = 568 nm.
Bone Isolation
Tibias of 2.5- to 3-week-old male chicks (Peterson-Arbor Acre) were removed and cleaned of adherent tissue. The bones were split longitudinally and marrow removed with forceps. Residual marrow was rinsed away in minimal essential media (MEM) (Sigma) supplemented with 10% calf serum. The bone halves were then split into smaller pieces (∼0.1–1 cm) with a razor blade. These smaller pieces of bone were stored in MEM plus 5.0% fetal bovine serum (FBS) (Sigma) in the incubator until use. The bone fragments were used to label osteoclasts attached to the endosteal surface and were not in the incubator longer than 12 hr.
Carbonic Anhydrase Labeling and Imaging
Pieces of tibia were prerinsed in MEM for 5 min at 37C, then labeled with AZ-Bd for 5–30 min at concentrations ranging from 10−9 M to 5 × 10−7 M. The pieces were then rinsed three times for 5 min, to remove excess label. For competitive studies, cells were preincubated with the unlabeled inhibitors for 5 min, then co-incubated for 5–30 min with AZ-Bd plus inhibitors. The bone pieces were placed in MEM in a chamber covered by a glass coverslip. Fluorescent images were generated with the BioRad MRC-600 laser scanning confocal imaging system integrated with an upright optical microscope using the YHS filter block with an excitation beam at 568 nm and a 10% neutral density filter. Images were constructed by projecting the 2–4-μm sections generated using either a x 10 (Leitz, 0.25 NA) or x25 (Leitz, 0.5 NA) objective. All projections were made in the Comos software package provided with the BioRad system. The same gain and contrast levels were maintained throughout each experiment to allow valid comparisons to be made. Trifluoromethane sulfonamide (TFMS) was provided by Molecular Probes. Acetazolamide and 6-ethoxyzolamide were purchased from Sigma. Merck, Sharpe and Dohme provided 5-(3-hydroxybenzoyl)-thiophene-2-sulfonamide (HTS).
Osteoclast Isolation for Acidification Assay
Osteoclasts were isolated from the tibias of 2.5- to 3-week-old chicks according to Hunter et al. (1991), with the following modifications. The tibias were subjected to a digestion in trypsin (0.03%, 20 min) before the endosteal surface was scraped. Cells were filtered through polypropylene meshes (Small Parts; Miami Lakes, FL), plated on fibronectin-coated plates (0.02 mg/ml), and incubated at 37C in 5% CO2, 95% air for 10 min. Nonadherent cells were then transferred to another set of plates and incubated for 12.5 min. The adherent cells from both sets of plates were resuspended by gentle swirling (six times) in MEM + 5% FBS, after which approximately 105 cells/ml were plated on 12-mm round glass coverslips. At 24 hr in culture, the coverslips were rinsed in MEM and placed in fresh medium (MEM + 0.5% FBS). Acridine orange experiments were performed at Day 4 in culture.
Osteoclast Acidification Assay
Osteoclasts were neutralized with 20 mM NH4Cl in MEM for 15 min, rinsed in MEM plus treatment (three times for 30 sec), and allowed to recover in the presence or absence of carbonic anhydrase inhibitors for 10–20 min. The cells were then incubated for 10 min in treatment plus 10 μM acridine orange (Polysciences; Warrington, PA) as previously described (Hunter et al. 1988). The fluorescent intensities of 20 cells per coverslip per time per treatment were measured using a Leitz MPV photometer (excitation 455–490 nm, emission 510 nm, suppression filter 515 nm). For each experiment, one coverslip was incubated in 20 mM NH4Cl and 10 μM acridine orange to establish the level of background fluorescence.
Statistics
The data in Table 1 were compared using a one-factor ANOVA to show that the competition values were different from the uncompeted values.
Results
Synthesis of Bodipy 558/568-modified Acetazolamide
The synthesis scheme for Bodipy 558/568-modified acetazolamide is shown in Figure 1 and is described in Materials and Methods. The resultant compound is excited at 558 nm and emits at 568 nm.
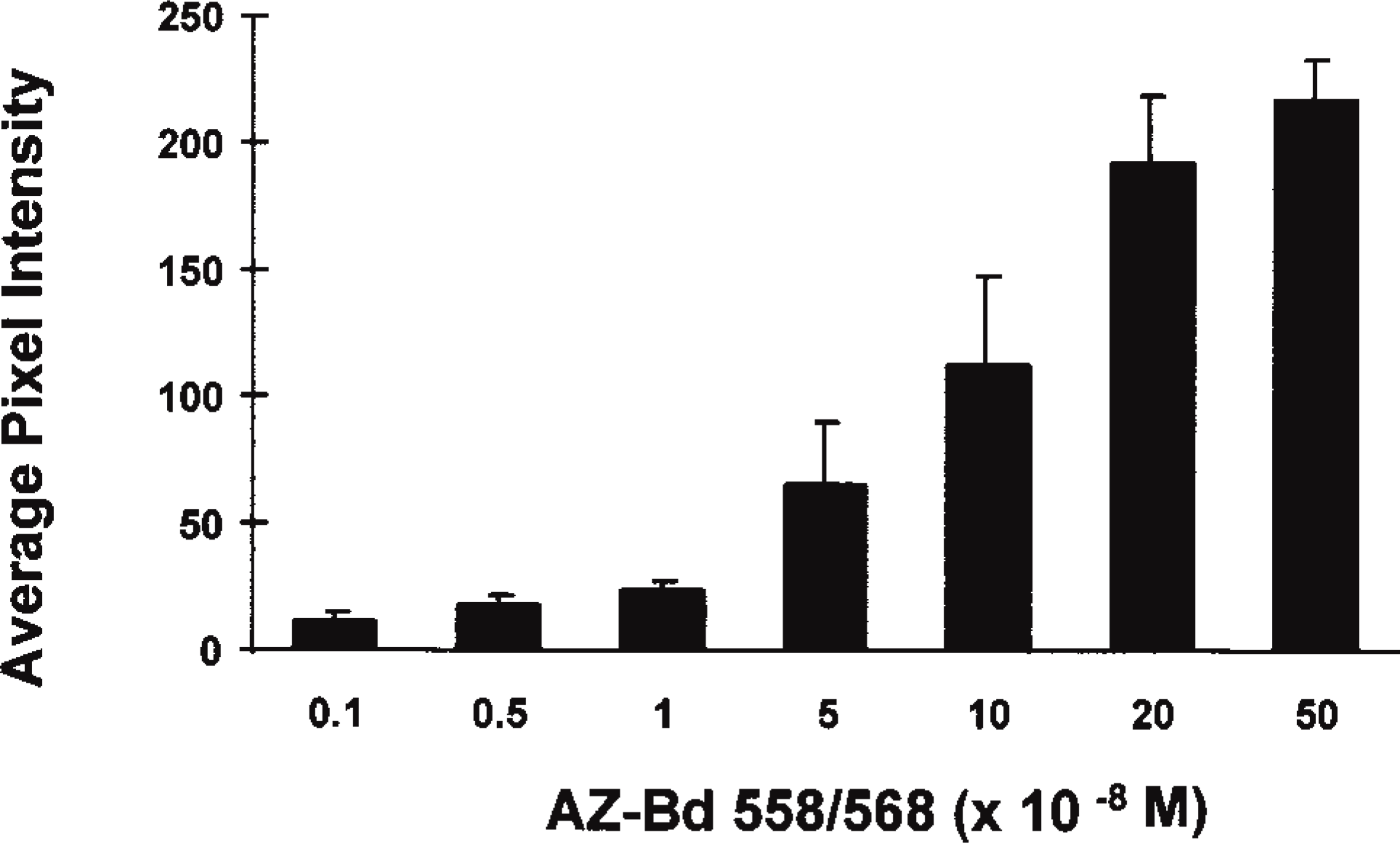
Intracellular concentration profile for Bodipy 558/568-modified acetazolamide. Osteoclasts on bone surfaces were labeled with 10−9 M to 5 × 10−7 M AZ-Bd for 15 min and pixel intensities were determined using a confocal microscope system. Values presented are mean ± SD for four experiments combined for an average of 80 cell intensities per bar.
Cell Labeling and Competition Assays
Freshly isolated osteoclasts on bone were labeled with AZ-Bd in a dose-dependent manner, as shown in Figure 2. The labeling was specifically localized throughout the cytoplasm of osteoclasts, with a 100-fold excess of unlabeled acetazolamide eliciting a 57.4 ± 2.0% decrease in fluorescent staining of cells on bone (Figure 3). Ethoxyzolamide, trifluoromethane sulfonamide (TFMS), and 5-(3-hydroxybenzoyl)-thiophene-2-sulfonamide (HTS) all competed with the labeled acetazolamide for cell staining (Table 1). Intensity of staining was decreased by approximately 50% with a 100-fold excess of unlabeled inhibitors.
Inhibitory Effectiveness of Labeled Acetazolamide
This assay is based on an earlier study in which cultured osteoclasts were shown to produce acid after neutralization with NH4Cl as a result of carbonic anhydrase activity (Hunter et al. 1991). To assess if fluorescent labeling affected the activity of the inhibitor, osteoclasts were neutralized, then allowed to recover in the presence of the AZ-Bd. AZ-Bd decreased acid production as efficiently as unlabeled inhibitors, as shown in Table 2.
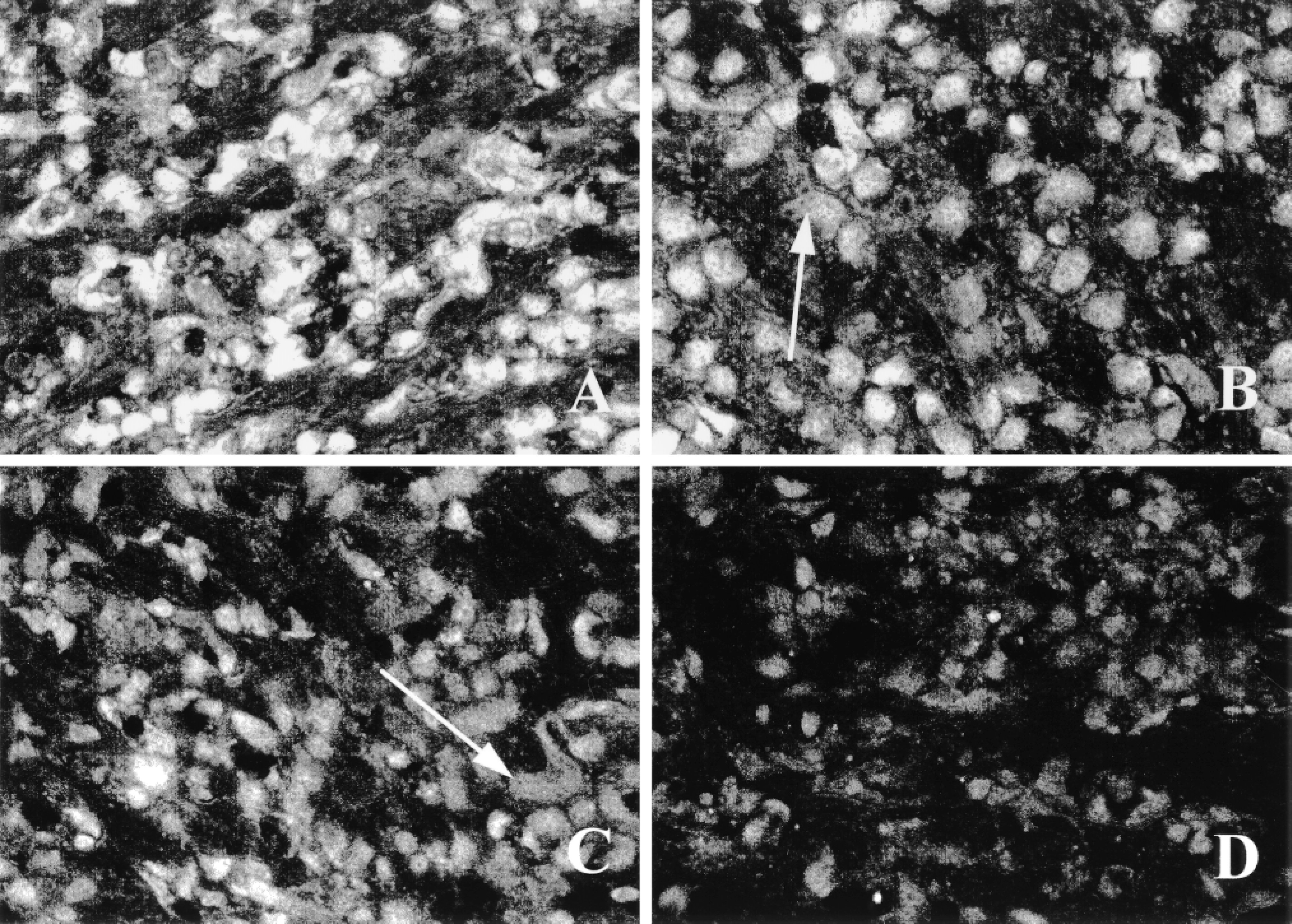
Composite of a competition assay with unmodified acetazolamide. Osteoclasts on bone were preincubated for 5 min with no (A), 5 × 10−8 M (B), 5 × 10−7 M (C), or 5 × 10−6 M (D) acetazolamide before adding 5 × 10−8 M AZ-Bd for 15 min. The 1:100 competition (D) decreased the fluorescent intensity of staining by approximately 57% (see Table 1). Unstained nuclei of the multinucleated osteoclasts are evident (arrows).
Discussion
We describe here the synthesis and activity of a fluorescent inhibitor of carbonic anhydrase and demonstrate that it can be used to label living cells. AZ-Bd enters osteoclasts, the cells that produce acid to resorb bone, and retains its capacity to inhibit carbonic anhydrase, as assessed by an assay that measures cytosolic acidification after neutralization with ammonium chloride. As expected, fluorescence was found throughout the cytoplasm of osteoclasts, with nuclei devoid of staining, as was reported earlier by [3H]-acetazolamide autoradiography (Gay and Mueller 1974) and immunohistochemistry (Gay et al. 1974). Neighboring cells on bone surfaces were not labeled. The fluorescent dye used to assess acid production, acridine orange, is sequestered in cells on exposure to H+ ions and fluoresces in the orange at low pH and in the green at a neutral pH (de Duve et al. 1974; Moriyama et al. 1982). Inhibition of acid production by AZ-Bd and the nonfluorescent sulfonamides was approximately 60%. The reason for incomplete inhibition could be explained by a recent study that revealed the presence of the Na+/H+ exchange mechanism in osteoclasts, a process that could contribute to recovery from neutralization (Gupta et al. 1996). Also contributing to incomplete inhibition is the spontaneous hydration of CO2 that is known to occur in tissues.
Competition assay results for carbonic anhydrase inhibitors a
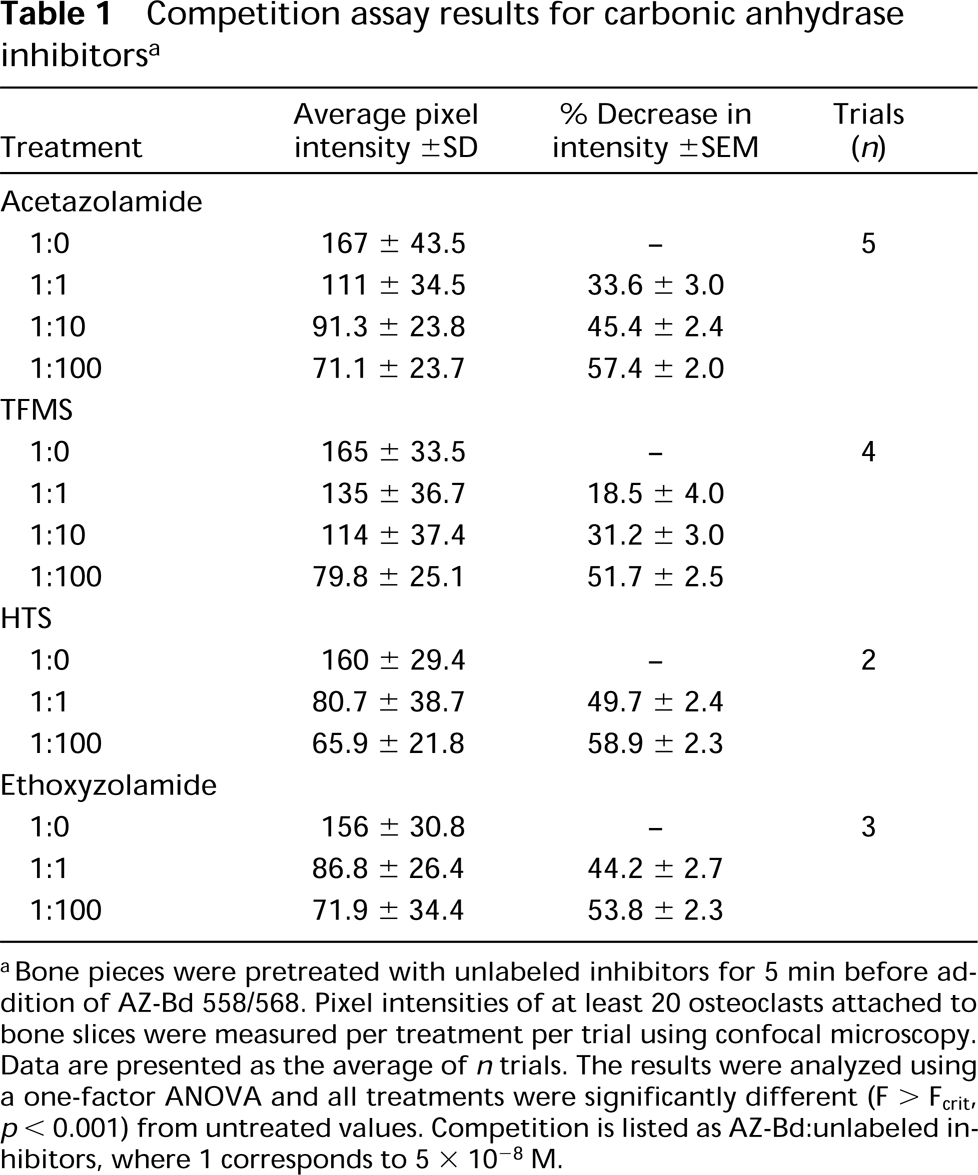

aBone pieces were pretreated with unlabeled inhibitors for 5 min before addition of AZ-Bd 558/568. Pixel intensities of at least 20 osteoclasts attached to bone slices were measured per treatment per trial using confocal microscopy. Data are presented as the average of n trials. The results were analyzed using a one-factor ANOVA and all treatments were significantly different (F < Fcrit, p > 0.001) from untreated values. Competition is listed as AZ-Bd:unlabeled inhibitors, where 1 corresponds to 5 × 10−8 M.
Acetazolamide was less effective than AZ-Bd and the other sulfonamides in blocking acid production. There are several possible explanations for this, including increased lipid solubility of the labeled acetazolamides due to the Bodipy side arm. The pKa of the −SO2NH2 side arm and lipid solubility play roles in the effectiveness of the inhibitor at pH 7.4. The pKa of −SO2NH2 at pH 7.4 is 9.1, 8.0, and 5.9, and the lipid partition coefficients are 0.001, 30, and 0.006 for acetazolamide, ethoxyzolamide, and TFMS, respectively (Maren 1992; Maren and Conroy 1993). The ideal inhibitor would be both water soluble and lipid soluble, thus having a low pKa and a high lipid partition coefficient. Therefore, a possible explanation for why it was necessary to pretreat cells with unlabeled inhibitors, before addition of AZ-Bd for competition studies could be the consequence of an increase in lipid solubility of the fluorescent compound due to the Bodipy side arm.
Specificity of binding was demonstrated by competition with acetazolamide as well as ethoxyzolamide, HTS, and TFMS. Staining decreased with increasing unlabeled inhibitor. Osteoclasts still adherent to bone were used to assess the effectiveness of the dye because these cells have been shown to contain carbonic anhydrase (Gay and Mueller 1974; Gay 1996) and are actively producing acid, a carbonic anhydrase-dependent process (Hunter et al. 1988,1991). Staining of cultured cells was observed but with much less intensity (not shown). Asotra et al. (1994) have shown that carbonic anhydrase levels decrease in nonresorbing vs resorbing osteoclasts.
Seven distinct isozymes of carbonic anhydrase, designated CA I-CA VII, have been characterized from mammals (Tashian et al. 1991). Few enzymes approach the distributional and functional diversity of carbonic anhydrase, which may be cytosolic (I, II, III, and VII), mitochondrial (V), membrane-bound (IV, V), or secreted (VI), and these range in distribution from their presence in many cell types throughout the animal kingdom (II) to more limited expressions in plants and bacteria (Fernley 1988). Inhibition of the isoforms of carbonic anhydrase by sulfonamides is variable, with the most research focused on isoform II (Maren and Conroy 1993; Sly and Hu 1995). The cells examined in this study, avian osteoclasts, have been shown to contain solely CA II, as reviewed earlier (Gay 1996). Sulfonamide inhibition of the isoforms varies, with CA II and V being the most sensitive, forms I, IV, and VI being somewhat sensitive, and form III being insensitive to inhibition (Sly and Hu 1995). Very little is known about the inhibition of form VII. Carbonic anhydrase from the green algae Chlamydomonas reinhardtii and from spinach and pea chloroplasts had some affinity for the sulfonamides (Sly and Hu 1995). Therefore, a fluorescent carbonic anhydrase inhibitor should be useful for studying CA in a variety of ways, including localization studies, determining kinetic data, and quantitation of activity or levels.
Effect of carbonic anhydrase inhibitors on osteoclast acidification a
aIsolated osteoclasts were neutralized in NH4Cl, treated with carbonic anhydrase inhibitors for 20 min, then stained with acridine orange in the presence of the inhibitor (10−8 M). The fluorescent intensity of 20 cells per treatment per experiment was recorded using a photometer attached to a fluorescent microscope. Data presented are averages of three experiments. Untreated cells were allowed to recover from NH4Cl neutralization in media without inhibitor and non-recovery cells were those stained with acridine orange in the presence of 20 mM NH4Cl to establish a baseline fluorescence (see Materials and Methods).
Footnotes
Acknowledgements
Supported by NIH grant DE 04345.
We would like to thank Virginia Gilman for assistance with editing, proofreading, and figure preparation.