
Abstract
In the context of immune-endocrine relationships, we have previously shown that interferon-γ (IFN-γ) inhibits hormone secretion in anterior pituitary (AP) cell cultures. The non-hormone-secreting folliculostellate (FS) cells were found to mediate this inhibitory action. Because in the immune system IFN-γ is a strong stimulator of nitric oxide (NO) release through the induction of NO synthase (NOS), we investigated whether the inducible form of NOS (iNOS) is present in (rat) AP cell cultures, and whether its expression is stimulated by IFN-γ. Immunocytochemistry revealed that under basal in vitro conditions only a very few AP cells contained iNOS. Treatment with IFN-γ caused a sixfold rise in the number of iNOS-positive cells and augmented the intensity of the staining. The increased number of iNOS-expressing cells was paralleled by elevated production of NO. Some of the iNOS-positive cells extended cytoplasmic processes between hormone-secreting cells, which is a characteristic of FS cells. Immunostaining of FS cell-poor and FS cell-enriched populations (obtained by gradient sedimentation) also suggested the presence of iNOS in a subpopulation of FS cells. By double immunofluorescence techniques we found that about 65% of iNOS-expressing cells were positive for S-100, a marker protein for FS cells. However, around 80% of the S-100-positive cells were not labeled for iNOS. On the other hand, the majority of the S-100-negative iNOS-containing cells could not be further identified by antisera against the classical AP hormones, suggesting the presence of iNOS in a still unidentified non-hormone-secreting cell type of the AP gland. This report is the first to demonstrate the expression of the inducible form of NOS in the AP gland. IFN-γ upregulates this expression, showing that cytokines may use the same signaling mechanisms in both the immune and the endocrine system. In addition, a putative new function of a subpopulation of FS cells in the paracrine regulation of the AP gland is suggested.
Keywords
F
In the present study we addressed the question of whether IFN-γ uses the same signaling molecule(s) in the AP gland as it does in the immune system. One of the factors recently discovered and now well documented to play a prominent role in mediating the effects of IFN-γ in the immune system is nitric oxide (NO) (for review see Vilcek and Oliveira 1994). Macrophages activated by IFN-γ produce large amounts of NO. IFN-γ stimulates the expression of the inducible form of NO synthase (iNOS), one of the isozymes catalyzing the formation of NO from L-arginine (Knowles and Moncada 1994; Nathan and Xie 1994; Vilcek and Oliveira 1994). In addition to (macrophage) iNOS, two other forms of NOS have been identified which are constitutively expressed and which, unlike iNOS, are regulated by intracellular Ca2+ (for review see Knowles and Moncada 1994; Nathan and Xie 1994), i.e., the endothelial constitutive NOS (ecNOS) and the brain cNOS (bcNOS). The presence of bcNOS has been demonstrated in the rat AP gland (Ceccatelli et al. 1993; Bredt et al. 1990) and appears to be confined to gonadotrophs and some other cells with FS cell morphology. In the human pituitary (Lloyd et al. 1995), bcNOS was detected in all secretory cell types and in some S-100-positive cells, whereas ecNOS was found in non-identified cells and endothelial cells. cNOS immunoreactivity is upregulated in pituitary adenomas (Lloyd et al. 1995). High levels of bcNOS have been detected in the pituitary GH3 cell line (Wolff and Datto 1992) and of iNOS in the corticotroph cell line AtT20 after induction with interleukin-1β (IL-1β) (Ohta et al. 1993).
In the present study we investigated whether IFN-γ is capable of inducing iNOS in AP cells. Studies were done in vitro because in vivo administration of IFN-γ is known to result in the release of a plethora of other cytokines and factors that may complicate interpretation of the results. To identify iNOS-expressing cells, differentially enriched AP cell populations obtained by unit gravity sedimentation in a serum albumin gradient and double immunofluorescence techniques were used.
Materials and Methods
Antibodies
The polyclonal rabbit anti-iNOS antiserum was a gift from Dr. S. Moncada and Dr. V. Riveros-Moreno of Wellcome Research Laboratories (Beckenham, Kent, UK). The specificity of the antiserum, which was raised against a synthetic peptide fragment of the mouse iNOS molecule, has previously been shown (Hamid et al. 1993). The monoclonal anti-iNOS antibody (IgG2a isotype) was purchased from Transduction Laboratories (Lexington, KY). The antibody had been raised in mice against a 21-kD protein corresponding to amino acids 961–1144 of the mouse macrophage iNOS molecule and also recognizes human, cow, and rat iNOS. The specificity of the antibody has been demonstrated by the supplying company on Western blot and by our pre-adsorption studies as described below. For staining S-100, a polyclonal rabbit antiserum against bovine S-100 was used (Dako; Glostrup, Denmark). For staining AP hormones [prolactin (PRL), growth hormone (GH), corticotropin (ACTH), thyrotropin (TSH), luteinizing hormone (LH)-β, and follicle-stimulating hormone (FSH)-β], the polyclonal rabbit anti-rat antisera used were a gift from Dr. A.F. Parlow through the National Hormone and Pituitary program (NIDDK and NHPP; University of Maryland, Baltimore, MD).
Immunocytochemistry of Paraffin-embedded Sections from Rat Anterior Pituitary Cell Aggregates
Anterior pituitaries (AP) were obtained from 3-month-old random cycling female Wistar rats, and AP cells from pooled pituitaries were prepared as previously described (Denef et al. 1989; Van der Schueren et al. 1982). Animals had been bred and kept in an environment of constant temperature, humidity, and day-night cycle with free access to water and food (Animal Breeding Facility; University of Leuven, Belgium). AP cells were allowed to re-aggregate on a gyratory shaker in a 1.5% CO2/air incubator. These aggregates have been shown to be composed of AP cells displaying ultrastructural and functional integrity (Van der Schueren et al. 1982). In some experiments, AP cells were first fractionated by unit gravity sedimentaton in a bovine serum albumin (BSA) gradient to obtain cell populations differentially enriched in one of the cell types (Denef et al. 1989; Baes et al. 1987). The distribution of the different AP cell types in a large number of independent cell separations has been found to be consistent and has been compiled into an internal laboratory standard pattern (Vankelecom et al. 1989, 1992; Denef et al. 1989; Baes and Denef 1987). After 5 days in culture, aggregates were treated with rat IFN-γ (500 U/ml) (courtesy of Prof. P. Van der Meide; ITRI, Rijswijk, Nederland) for 24 hr. Subsequently, they were fixed in Zamboni fluid (4% paraformaldehyde and 15% saturated picric acid solution in 0.1 M phosphate buffer, pH 7.4) for 4 hr, paraffin-embedded, and sectioned as previously described (Tilemans et al. 1992; Denef et al. 1989).
Immunostaining was performed as reported previously (Tilemans et al. 1992; Denef et al. 1989). Briefly, permeabilisation by trypsin (0.1% bovine pancreatic trypsin, Type III; Sigma Chemical, St Louis, MO) and incubation with pre-immune swine serum (Dako) (1:5 in PBS, pH 7.4) was followed by overnight reaction with the polyclonal rabbit anti-iNOS antiserum (at a dilution of 1:2000 in PBS). Staining was done using the sensitive avidin-biotin complex (ABC) method (ABC-HRP; Dako) with biotinylated swine anti-rabbit Ig (Dako) (1:400) as the secondary antibody and diaminobenzidine (DAB) as the brown color reagent. Appropriate controls were done in which the anti-iNOS antibody or the secondary antibody was omitted.
Quantitative data were obtained by counting the number of iNOS-positive cells per unit aggregate section. Of at least 15–20 sections per condition (control or IFN-γ-treated), the surface area was measured using computer-assisted image analysis (Quantimet 500; Leica, Cambridge, UK) and the immunopositive cells in the section were counted. Data are expressed as number of iNOS-immunoreactive cells/mm2 section area (mean ± SE of the indicated number of independent experiments). Statistical analysis was performed using analysis of variance (ANOVA) methods as described (Vankelecom et al. 1990).
Single and Double Immunofluorescence of Rat Anterior Pituitary Cells in Monolayer Culture
To avoid crossreactivity in double immunofluorescence, antibodies from different species were chosen in combination with distinct fluorophores. The mouse monoclonal anti-iNOS antibody (Transduction Laboratories), however, did not produce a signal in paraffin-embedded aggregate sections, probably due to destruction of the (single) epitope by the paraffin and organic solvent treatment. Different fixation protocols proved unsuccessful. Therefore, we switched to AP cells cultured as monolayers. Staining for hormones in monolayer cells has previously been shown (Vankelecom et al. 1993; Houben and Denef 1991; Carmeliet and Denef 1989). Moreover, clear immunofluorescent signals have been obtained for S-100 protein in AP cell monolayer cultures (Otto et al. 1996).
To obtain AP cell monolayers, cells were prepared as described above, and were seeded on poly-L-lysine (Sigma)-coated coverslips in six-well tissue culture plates (Corning Glass Works; Corning, NY). Cells were kept in defined culture medium (Denef et al. 1989) supplemented with 1% fetal calf serum (FCS) for better cell attachment and were cultured in a 1.5% CO2/air incubator. After 1–2 days in culture, cells were treated with 500 U/ml of rat IFN-γ for 24 hr, washed, and fixed with 1% paraformaldehyde/1.5% glutaraldehyde in PBS for 10 min. To lower background staining, samples were treated with NaBH4 (0.5 mg/ml in PBS). Cells were permeabilized with Triton X-100 (0.1% in PBS) and aspecific binding blocked with BSA (3% in PBS). In the first experiments, staining with the monoclonal anti-iNOS antibody was compared to S-100 staining on separate cell samples, using a fluorescein (FITC)-labeled secondary antibody (rabbit anti-mouse Ig for iNOS and swine anti-rabbit Ig for S-100, both at a dilution of 1:40) (Dako). After the permeabilization and blocking reaction, cells were incubated with the primary antibody for 1 hr (at a dilution of 1:10 for iNOS and 1:500 for S-100 in 0.3% Triton X-100/3% BSA in PBS) and, after rinsing, with the secondary antibody for 30 min. After thorough washing, coverslips with the cell samples were mounted on a glass slide using glycerine (50% in PBS) with 25 mg/ml diazabicyclo[2.2.2]octane (DABCO; Sigma). Cells were examined on a Leica DMRB microscope (Leica; Wetzlar, Germany) using a Leica FITC filterset (I3), and images were recorded on a Leitz Orthomat E semiautomatic camera.
For double immunofluorescence, secondary antibodies labeled with different fluorophores were selected that showed only negligible overlap in absorption and emission spectra. iNOS was visualized with sheep anti-mouse IgG [F(ab')2 fragment] conjugated with 7-amino-4-methylcoumarin-3-acetic-N′-hydroxysuccinimide ester (AMCA) (1:10) (Boehringer; Mannheim, Germany) giving a blue fluorescent signal (excitation optimum 345 nm; emission at 450 nm). For staining S-100 and the AP hormones, a goat anti-rabbit IgG [F(ab')2 fragment] conjugated with indocarbocyanine (Cy3) (1:800) (Jackson ImmunoResearch Laboratories; West Grove, PA) was used, which resulted in a red signal (excitation optimum 550 nm; emission at 570 nm). Images were viewed using Leica filters UVA for AMCA and N2.1 for Cy3. Cells stained in a single staining procedure for iNOS were blue but did not show red fluorescence using the Cy3 filter combination. When staining only for S-100 (or the AP hormones) was performed, a red signal was obtained, whereas no blue color was observed using the AMCA filter set. The procedure used for double immunofluorescence of iNOS and S-100 was similar to the one described for single staining. The procedure for double staining of iNOS and the AP hormones (PRL, GH, ACTH, TSH, LH-β/FSH-β) resulted in high background levels. Therefore, minor modifications were introduced, including extension of fixation time from 10 min to 1 hr and of incubation time with the primary antibodies from 1 hr to overnight, resulting in more satisfying hormone staining and lower backgrounds. Controls were performed in which one of the primary or the secondary antibodies was omitted. In addition, a non-sense mouse IgG2a (Dako) was used as a negative control for iNOS staining.
To confirm the specificity of the monoclonal anti-iNOS antibody, we performed pre-adsorption studies, using the mouse macrophage lysate that is provided by Transduction Laboratories as a positive control for iNOS immunoreactivity. Antigen (lysate at a dilution of 1:10) and antibody were incubated overnight at 4C. The mixture was further used for single immunofluorescence as described above. As a negative control antigen, human endothelial cell lysate was used (Transduction Laboratories). This preparation was shown by the supplying company to contain ecNOS but no iNOS. Staining for S-100 and pituitary hormones by polyclonal rabbit anti-rat antisera has in our previous work been shown to be specific (Denef et al. 1989).
Proportions of double (iNOS + S-100) stained cells and of single (iNOS or S-100) stained cells were obtained by counting as many cells per field as possible before fading occurred. In general, about 300–400 immunofluorescent cells were counted per independent experiment. Data are expressed as mean ± SE of four independent experiments. In the Results section, only a limited number of (representative) color microphotographs are shown; additional pictures are available on request.
Uptake of the Dipeptide β-Ala-Lys-N∊-AMCA as a Marker for FS Cells
Recently, Otto et al. (1996) reported that FS cells, but no other AP cells, contain a dipeptide carrier of which the presence can be made visible by uptake of the fluorescent dipeptide β-Ala-Lys-N∊-AMCA. We used this dipeptide as an additional marker for FS cells in monolayer cultures. AP cells seeded on coverslips (see above) were incubated with 15 μM β-Ala-Lys-N∊-AMCA (kindly provided by Dr. C. Otto and Dr. K. Bauer; Max-Planck Institute, Hannover, Germany) for 3 hr. After thorough washing, cells were fixed and processed as described above.
Nitrite Concentration in AP Cell Monolayer Culture Supernatant as a Measurement of NO Production
AP cells were cultured for 2 days at a density of 200,000 cells/ml in serum-free defined medium. Medium was removed and cells were stimulated with IFN-γ (100 U/ml) for 24 hr. Nitrite (NO2 −) levels in the supernatant were determined spectrophotometrically (absorbance at 540 nm) after mixing 100-μl samples with 100 μl Griess reagent [sulfanilamide 2% (w/v), N-(1-naphthyl)ethylenediamine 0.2% (w/v), phosphoric acid 4% (v/v); all products from Sigma]. NaNO2 (Sigma) was used as a standard. In preliminary experiments, nitrate (NO3 −) levels were also measured by comparing NO2 − levels in cell supernatants before and after treatment with nitrate reductase (Sigma) (Matthys et al. 1995). NO2 − levels did not change (data not shown), implying that NO3 − is undetectable in our samples (NaNO3 from Sigma was used as a positive sample and standard). Therefore, in subsequent assays, only NO2 − levels were determined.
Results
IFN-γ Induces iNOS Expression in AP Cells, Some of Which Resemble FS Cells
Cell re-aggregates from primary rat AP cells were treated with IFN-γ (500 U/ml) for 24 hr, and paraffin-embedded sections were stained for iNOS using a rabbit polyclonal anti-iNOS antiserum. As shown in Figure 1A, iNOS was detectable in AP cells under basal conditions. However, only a very small number of cells showing only a weak signal were detectable, i.e., 103 ± 11 cells/mm2 aggregate section area (mean ± SE; n = 4) (which corresponds to 5–10 cells per average section of one aggregate). Treatment with IFN-γ (Figure 1B) clearly upregulated the expression of iNOS: the number of iNOS-positive cells increased sixfold (605 ± 13 cells/mm2 after IFN-γ-treatment, n = 4; IFN-γ-treated vs. control p<0.001 by ANOVA), and the staining became more intense.
A closer look revealed that some of the iNOS-positive cells exhibited the morphology typical of FS cells (S-100-positive cells). They displayed thin cell processes protruding between the other AP cells (see arrows in Figure 2A). Other iNOS-containing cells displayed a round or polygonal morphology. In addition, some S-100-positive cells were observed as round cells without the typical stellate shape (for an example see Figures 2D, 3A, and 4A).
To obtain further evidence that iNOS is expressed in FS cells, immunostaining was performed on AP cell populations that were differentially enriched in FS cells as obtained by unit gravity sedimentation in a BSA gradient. iNOS-positive cells were observed in populations that contain a high number of FS cells [Figures 1C-1F: BSA gradient fractions (BSA fr) 2 and 3]. In BSA fr 3, 26 ± 3 cells/mm2 were positive for iNOS in control cultures, and 417 ± 24 cells/mm2 expressed iNOS after IFN-γ treatment (n = 3; IFN-γ-treated vs control p<0.001 by ANOVA). For BSA fr 2, no quantitative data were collected because of relatively large necrotic areas in the center of the aggregates. Only very few iNOS-positive cells were found in populations that are poor in FS cells (Figures 1G-1J: BSA fr 4 and 7–9) (for proportions of FS cells in different BSA fr see Table 1 and Vankelecom et al. 1989, 1992). Some iNOS-positive cells of BSA fr 2 and 3 (as well as 7–9: see below) displayed the typical cytoplasmic extensions of FS cells (see arrows in Figures 2B, 2C, and 2E for iNOS compared to Figures 2D and 2F for S-100).
To test the functional activity of iNOS in IFN-γ-treated AP cells, NO2 − levels were determined in AP cell monolayer cultures as a measure of NO production (Table 1). NO production was stimulated by IFN-γ, and the level of stimulation was highest in populations with the highest proportion of FS cells (BSA fr 2 and 3), confirming our immunocytochemical data.
Because there were clearly more S-100-positive cells (FS cells) than iNOS-containing cells in the aggregates (for examples see Figures 2C and 2D), we hypothesized that (part of) iNOS is localized in a subpopulation of FS cells. The finding that BSA fr 4 still contains a considerable amount of S-100-positive cells (3–4%) (Vankelecom et al. 1989, 1992; see also inset of Figure 1H), but almost no iNOS-positive cells (Figures 1G and 1H) was concordant with this idea. On the other hand, BSA fr 7–9, known to contain FS cells that sediment through the gradient as clumps (Vankelecom et al. 1989, 1992), also contained some iNOS-immunopositive cells after IFN-γ treatment (Figure 1J), some of them displaying cytoplasmic extensions (Figures 2E and 2F).
As can be seen in Figure 1D, staining in BSA fr 2 was prominent along the peripheral borders of the aggregates, which may be partly due to the fact that there were some necrotic areas in the center of the aggregate. Necrotic zones have consistently been observed in aggregates highly enriched in FS cells (unpublished observations). However, in aggregates from BSA fr 3 cells (Figure 1F), as well as from unfractionated AP cell populations (Figure 1B), expression of iNOS was also most pronounced in the peripheral cell layers, and necrotic foci were not observed in these aggregates.
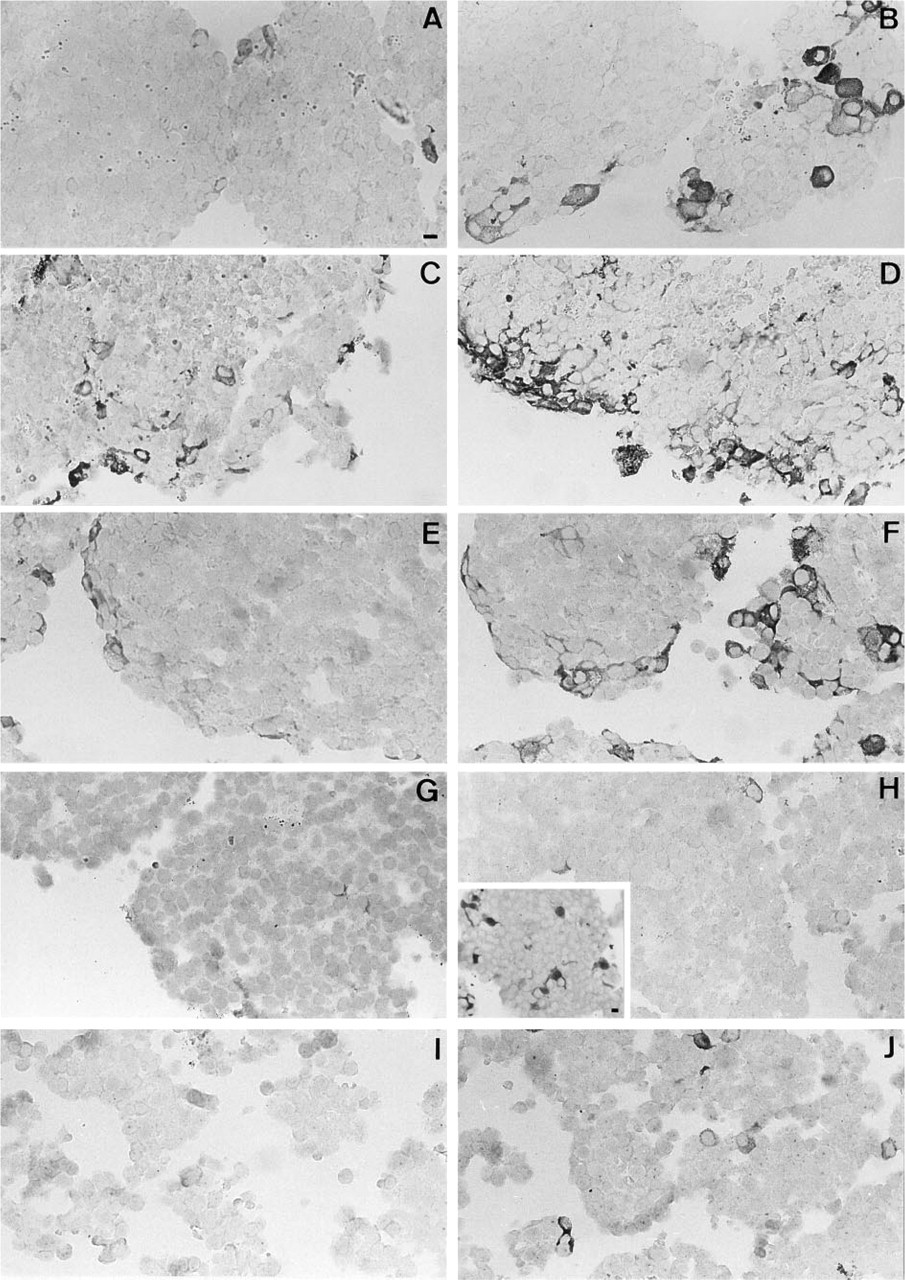
iNOS immunoreactivity in AP cell re-aggregates: upregulation by IFN-γ and (partial) co-sedimentation with FS cells.
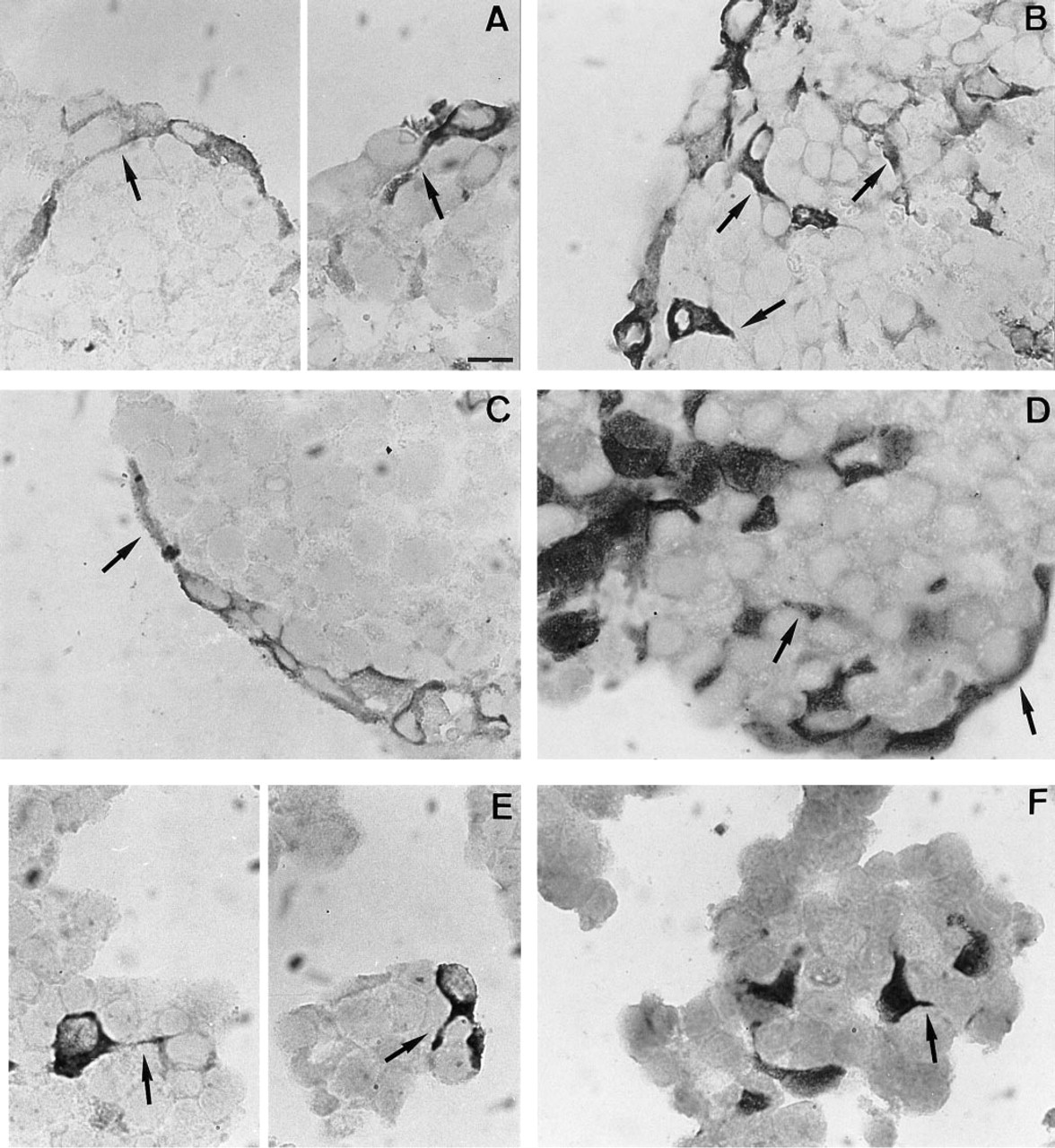
Morphological resemblance between some of the iNOS-positive cells and FS cells in AP cell re-aggregates. Higher magnification images of paraffin-embedded sections reveal that some iNOS-containing cells in unfractionated cell populations
Morphological resemblance between iNOS-positive cells and S-100-containing (FS) cells could also be observed in AP monolayer cell cultures treated for 24 hr with IFN-γ and processed for immunofluorescence using the monoclonal antibody against iNOS (see Figure 3). Cultured in the presence of a small amount of FCS (1%), FS cells nicely displayed their starlike morphology (Figure 3A). Such morphology was also seen in some of the iNOS-positive cells (Figure 3B). Again, some non-stellate cells contained iNOS immunoreactivity. On the other hand, some non-stellate cells were also S-100-positive. Further evidence for the morphological resemblance was based on the recent observation that FS cells, but no other AP cells, bear a dipeptide carrier of which the presence can be made visible by uptake of the fluorescent dipeptide β-Ala-Lys-N∊-AMCA (Otto et al. 1996). Cells accumulating β-Ala-Lys-N∊-AMCA (see Figure 3C) displayed the stellate morphology as observed for a portion of the iNOS-containing cells. However, some non-stellate cells also accumulated β-Ala-Lys-N∊-AMCA.
Stimulation of NO production by IFN-γ in AP cell populations differentially enriched in FS cells
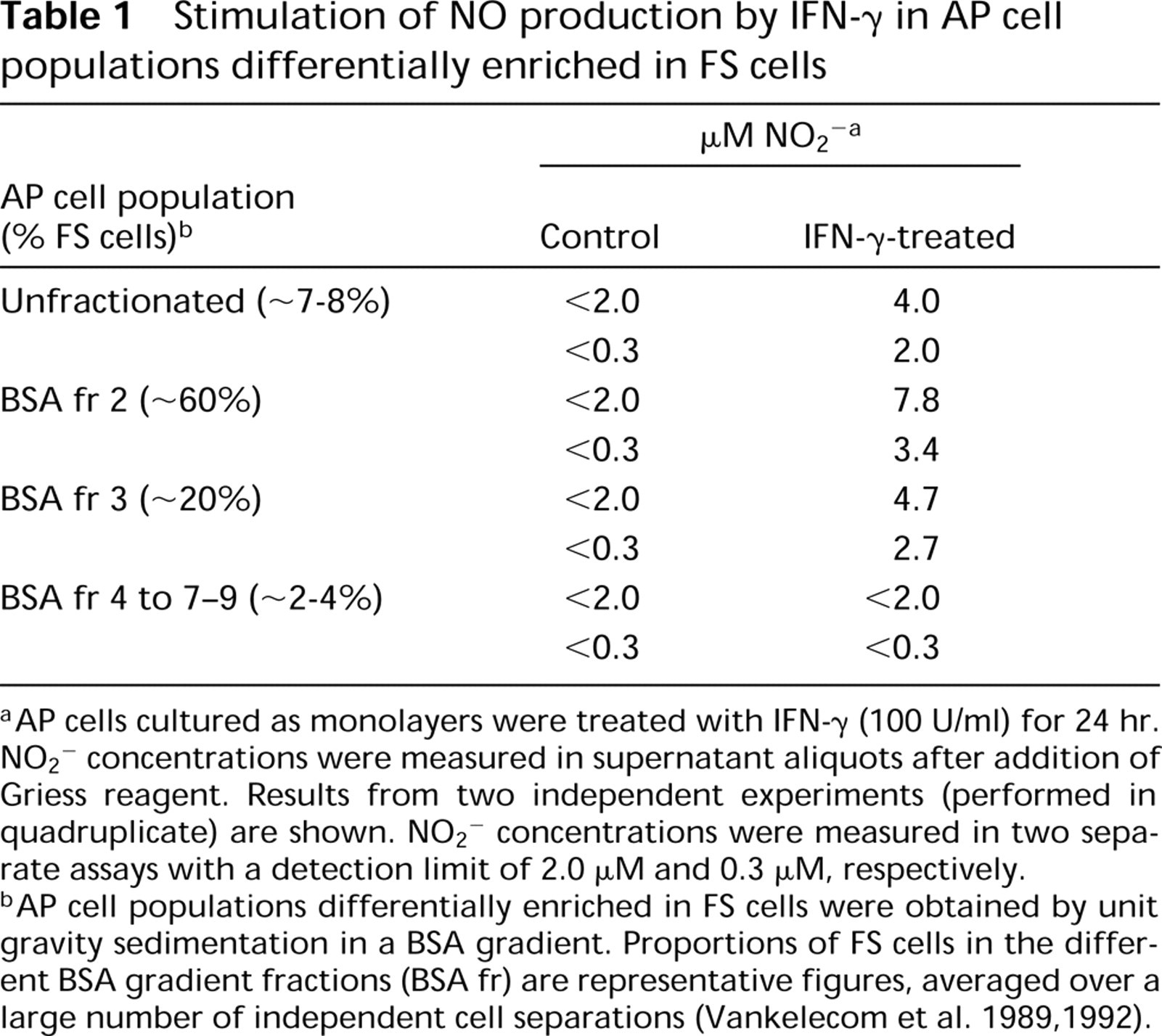
aAP cells cultured as monolayers were treated with IFN-γ (100 U/ml) for 24 hr. NO2 − concentrations were measured in supernatant aliquots after addition of Griess reagent. Results from two independent experiments (performed in quadruplicate) are shown. NO2 − concentrations were measured in two separate assays with a detection limit of 2.0 μM and 0.3 μM, respectively.
bAP cell populations differentially enriched in FS cells were obtained by unit gravity sedimentation in a BSA gradient. Proportions of FS cells in the different BSA gradient fractions (BSA fr) are representative figures, averaged over a large number of independent cell separations (Vankelecom et al. 1989, 1992).
The specificity of the monoclonal anti-iNOS antibody was confirmed by pre-adsorption experiments: incubating the antibody with iNOS-containing macrophage lysate abolished the staining (Figure 3D), whereas human endothelial cell lysate known to contain ecNOS but no iNOS did not affect the staining (data not shown).
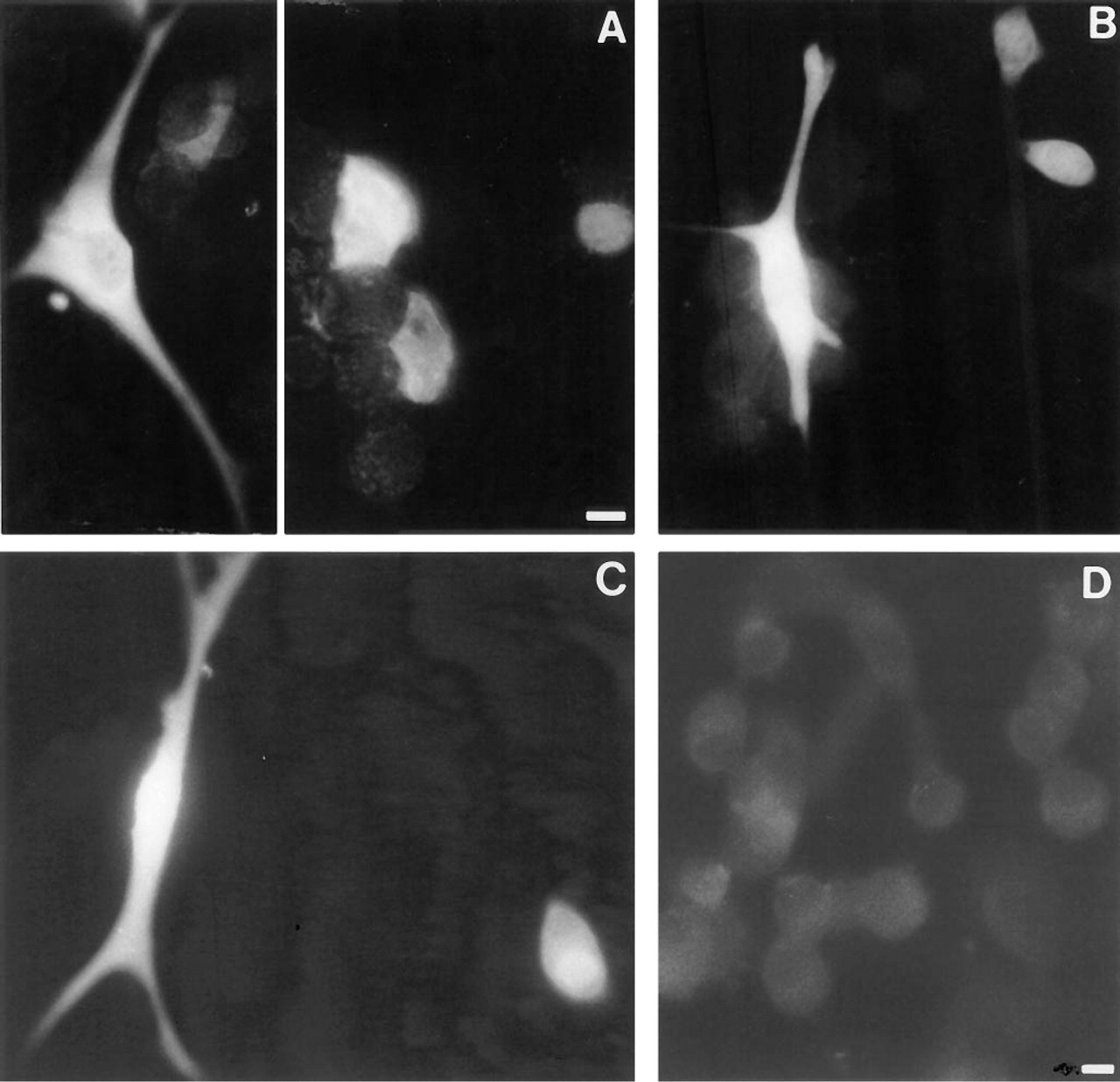
Morphological resemblance between some of the iNOS-positive cells and FS cells in AP cell monolayer cultures, and specificity test of the monoclonal anti-iNOS antibody. Immunofluorescence for S-100
Localization of iNOS in a Subpopulation of FS Cells and in Still Unidentified Cells as Revealed by Double Immunofluorescence
To conclusively identify the iNOS-expressing cell type(s), double immunofluorescence was performed. The mouse monoclonal antibody against iNOS and the rabbit polyclonal antisera against AP hormones and S-100 were used. Immunostaining with the anti-iNOS monoclonal antibody was unsuccessful in paraffin-embedded aggregate sections. In contrast, AP cells cultured as monolayers and fixed by brief glutaraldehyde and paraformaldehyde treatment provided a suitable alternative (see Materials and Methods).
As shown in Figures 4A and 4B, iNOS-positive (blue) cells were also S-100-positive (red). A large number of the S-100-positive cells (77.9 ± 4.3%, mean ± SE; n = 4) did not stain for iNOS. On the other hand, one third of the iNOS-positive cells (33.2 ± 1.6%; n = 4) failed to stain for S-100 (see arrowheads in Figures 4C and 4D). Only very sporadically, iNOS immunoreactive material was seen in cells that stained for LH-β/FSH-β (Figures 4E and 4F, arrow), or PRL (Figures 4G and 4H; no double-stained cell shown). In contrast, iNOS was never found in cells staining for ACTH (Figures 4I and 4J), GH, or TSH. Appropriate controls in which one of the primary antibodies or the secondary antibodies was omitted did not show any sign of aspecific staining or staining overlap. Substituting the mouse monoclonal anti-iNOS antibody with a mouse non-sense antibody of the same isotype (IgG2a) abolished blue immunofluorescent staining (data not shown).
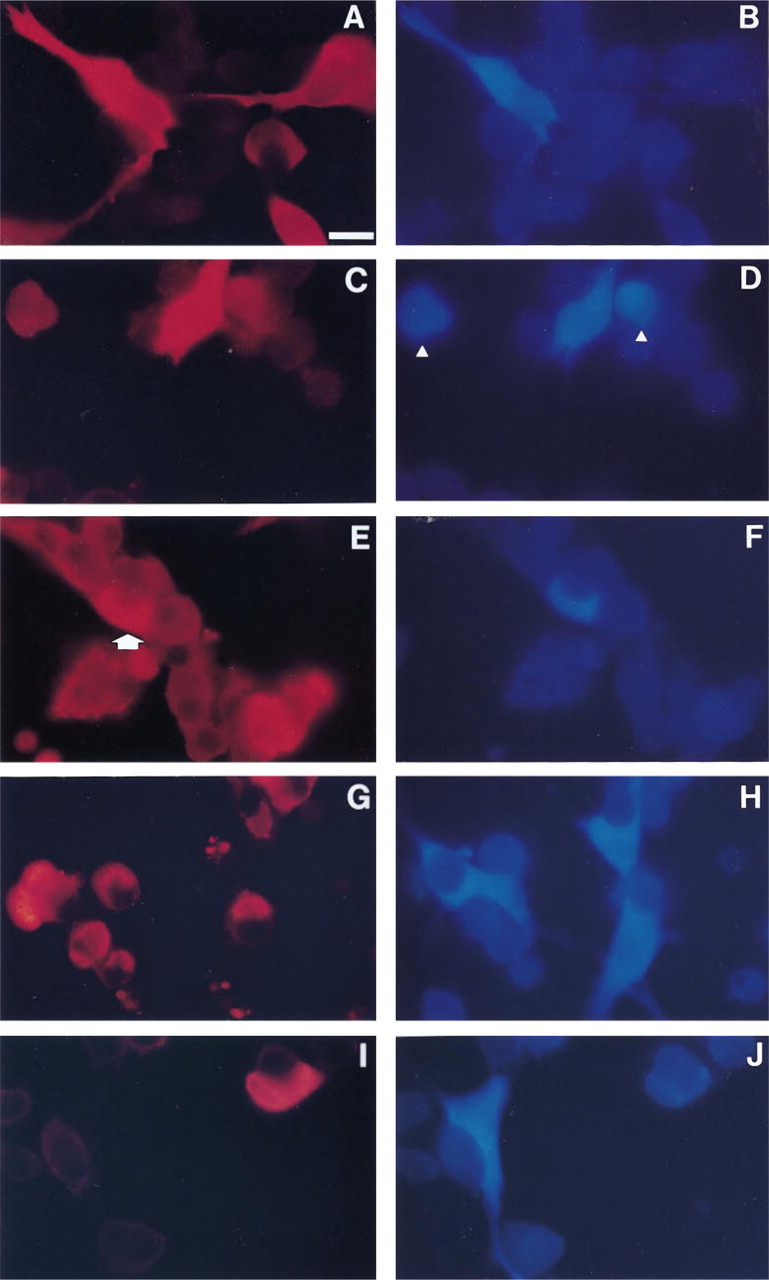
Localization of iNOS in a subpopulation of FS cells and in as yet unidentifiable AP cells, as revealed by double immunofluorescence. About 20% of the S-100-containing (FS) cells
Discussion
In this study we demonstrated that in AP cell aggregate cultures a few cells express iNOS and that IFN-γ causes a drastic increase in the expression of this enzyme. The number of cells immunoreactive for iNOS increased by a factor of 6 and the intensity of the immunostaining significantly increased as well. Enhanced expression of the enzyme was paralleled by elevated NO production, indicating that immunoreactive material represents functional iNOS.
iNOS expression after IFN-γ treatment was found in (S-100-positive) FS cells, as indicated by the following observations. First, the morphology of some of the iNOS-positive cells resembled that of FS cells. Second, iNOS-containing cells co-sedimented with FS cells in a BSA gradient. Moreover, NO levels after IFN-γ stimulation in differentially enriched AP cell populations were higher when more FS cells were present. Third, double immunofluorescence data clearly showed iNOS immunoreactivity in some of the cells positive for S-100, indicating expression of iNOS in a subpopulation of FS cells. Importantly, however, a portion of the iNOS-positive cells did not contain S-100. These particular iNOS-containing cells failed to stain for the AP hormones (ACTH, PRL, GH, LH/FSH, TSH) apart from some very rare cells exhibiting PRL or LH/FSH immunoreactivity. These findings suggest the presence of iNOS in an AP cell type that remains to be identified. An attractive hypothesis is that iNOS may be expressed in progenitor cells. It has recently been shown that nerve growth factor (NGF) induces NO production in PC12 cells and that NO is responsible for cell growth arrest (cytostasis), a prerequisite for entry into the terminal differentiation pathway (Peunova and Enikolopov 1995). IFN-γ has been shown to facilitate NGF-induced differentiation of PC12 cells (Improta et al. 1988), possibly through enhanced production of NO by IFN-γ. Furthermore, after NGF treatment PC12 cells produce an IFN-like product which, in turn, may induce NO production and trigger differentiation events in these cells (Tirone and Shooter 1989). In analogy with these findings, it is tempting to speculate that progenitor cells in the AP might be driven into differentiation by NO. In view of this hypothesis, rare cells co-expressing iNOS with PRL or with LH/FSH may represent progenitor cells in the transition state on their way to becoming fully differentiated lactotrophs or gonadotrophs.
In view of the reported heterogeneity of FS cells (Al-laerts et al. 1990a, 1991a), which is also illustrated by the presence of S-100 and the uptake of β-Ala-Lys-N∊-AMCA in non-stellate cells, we cannot rule out the possibility that iNOS is expressed in FS cells that are not identifiable by S-100. Along the same line, iNOS may be localized in hormone-secreting cells that have lost their capacity to synthesize hormones in culture. This problem, however, has been minimized by the use of short-term cultures (1–2 days), thereby reducing the probability of de-differentiation of specific AP cells.
Thus far, other investigators have not found iNOS immunoreactivity in the AP which is probably because no inducing stimulus such as IFN-γ was added to their test system (Lloyd et al. 1995). However, the presence of iNOS in the corticotroph cell line AtT20 after IL-1β treatment (Ohta et al. 1993) is in contrast to our finding that corticotrophs do not express iNOS immunoreactivity, as demonstrated by the absence of iNOS in ACTH-containing cells with double immunofluorescence techniques and as further documented by the different morphology in aggregate sections and the distinct BSA gradient sedimentation pattern of both cell types (highest percentage of ACTH cells in BSA fr 4) (Vankelecom 1992; Denef et al. 1989; Baes and Denef 1987). This discrepancy may be accounted for by the use of different stimulatory cytokines or may reflect the neoplastic state of the AtT20 cells. The latter would be in line with the presence of high levels of bcNOS in pituitary adenomas (Lloyd et al. 1995) and in the pituitary cell line GH3 (Wolff and Datto 1992).
The expression of iNOS by FS cells after IFN-γ stimulation is interesting in view of the putative macrophage/dendritic cell-like nature of at least part of the FS cells (Denef 1994; Vankelecom et al. 1989, 1993; Allaerts et al. 1990a, 1991a; Stokreef et al. 1986). Moreover, IFN-γ, which is the principal macrophage-activating factor in the immune system, also activates FS cells to exert inhibitory actions on hormone secretion, possibly mediated by (a) paracrine factor(s) (Vankelecom et al. 1992).
The present study also showed that iNOS-positive cells clearly tend to be located at the periphery of the AP aggregates. This peculiar distribution is not the consequence of insufficient penetration of IFN-γ into the aggregate, because AP cell aggregates are fully permeable to macromolecules (Allaerts et al. 1990b). The peripheral localization may be due to the non-random distribution of iNOS-expressing cells in the aggregate, for which differences in cell adhesion properties of iNOS-expressing cells with other AP cell types may be responsible. Non-random distribution of AP cell types within the aggregate has been demonstrated in our previous work (Tilemans et al. 1995; Allaerts et al. 1991b).
Several authors have reported that NO may be implicated in the regulation of AP hormone release (Duvilanski et al. 1995; Rivier 1995; Brunetti et al. 1993; Ceccatelli et al. 1993; Kato 1992). In recent work, we were able to demonstrate that NO produced by iNOS is involved in the inhibitory effect of IFN-γ on AP hormone secretion, as revealed by the use of specific iNOS inhibitors (Vankelecom et al., submitted for publication).
In summary, our findings demonstrate that the inducible form of NOS is expressed in the AP gland and is strongly upregulated by IFN-γ. It is detected in a subpopulation of FS cells and in other as yet unidentified cells. The presence of iNOS, in addition to the expression of cNOS as demonstrated by others, suggests an important regulatory role for NO in the AP gland. Moreover, the presence of both forms in one cell type, i.e., the FS cell, further emphasizes the significance of FS cells in the paracrine regulation of AP cell development, growth, and function, and in integrating hormonal responses to immune and other stress reactions.
Footnotes
Acknowledgements
This work was supported by grants from the Fund for Scientific Research-Flanders, Belgium (FWO), and the Flemish Ministry of Science Policy (Concerted Research Actions). HV is a Postdoctoral Fellow of the FWO, and PM is a Postdoctoral Fellow of the University of Leuven.
We thank Dr C. Otto and Dr K. Bauer (Max-Planck Institute, Hannover, Germany) for providing the fluorescent dipeptide β-Ala-Lys-N∊-AMCA, and for transferring fixation and fluorescent staining technology on AP cell monolayers. We also thank Dr J. Steel (ICRF; Histopathology, London, UK), Dr S. Moncada, and Dr V. Riveros-Moreno (Wellcome Research Laboratories, Beckenham, UK) for supplying the polyclonal anti-iNOS antibody. We further thank K. Rillaerts, L. Seghers, and T. Mitera for skilfull technical assistance.