
Abstract
Steroid acute regulatory protein (StAR) plays an essential role in steroidogenesis because it is responsible for the transfer of cholesterol from cellular stores to the inner mitochondrial membrane. We investigated the distribution and regulation of StAR expression in association with aldosterone production in the rat adrenal gland in vivo. Using nonradioactive in situ hybridization, we demonstrate that the outermost five to seven parenchymal cell layers express the StAR gene only weakly and inhomogeneously. The strongest expression is found in the zona fasciculata and zona reticularis. In addition, some cells in the adrenal medulla also stained positively. To differentiate between functionally active glomerulosa and inactive intermediate cells, we compared the expression pattern of StAR with that of aldosterone synthase. The expression of the latter is localized to two or three cell layers only, located immediately below the capsule. However, the cells of the intermedia are capable of expressing both genes prominently, as shown after stimulation with bilateral nephrectomy for 2 days. All zones of the adrenal cortex by then expressed StAR gene to the same extent. This was accompanied by a 50-fold elevated plasma aldosterone concentration. Our data demonstrate that the width of the aldosterone-producing zone can increase within a short period of time by recruiting hormonally inactive cells to steroidogenesis.
Keywords
A
Recently, an intermediate zone in the rat adrenal cortex was defined by immunohistological criteria, being located between the zona glomerulosa and zona fasciculata (Mitani et al. 1994). Whereas the zonae glomerulosa, fasciculata, and reticularis are functionally characterized by the presence of specific late-step steroidogenic enzymes, the intermediate zone lacks such enzymes. Under unstimulated conditions this zone produces neither aldosterone synthase (CYP11B2) nor 11β-hydroxylase (CYP11B1). The function of the zone is still a matter of debate. The aim of our study was to characterize the tissue-specific regulation of StAR expression in vivo with special respect to the zona glomerulosa and intermedia. To this end we chose two different stimuli of aldosterone production: first, ANGII infusion and second, the model of bilateral nephrectomy, which represents the strongest known experimental stimulus of aldosterone production.
Materials and Methods
Animals, Nephrectomy, Hemodialysis, and Drug Treatment
Adult male Sprague-Dawley rats weighing 400-520 g were housed under alternating 12-hr light and dark cycles at a constant temperature between 20C and 22C. They were fed the standard laboratory diet of Ssniff (R/M-H; Soest, Germany) and had free access to tapwater. All animal experiments were conducted in accordance with federal and local laws as well as institutional regulations.
All animals received chronic indwelling catheters into the femoral artery and vein under Ketamine/Xylazine anesthesia (75 mg/kg and 6 mg/kg, respectively) and were further allocated to one of three groups of six rats each. Each animal received an osmotic minipump (Alzet pumps, model 2ML1; Charles River, Sulzfeld, Germany) which supplied 10 μl per hour at a constant rate. The pumps were implanted
Determination of Plasma Aldosterone and ANGII
Plasma samples were obtained from conscious rats via a catheter at the end of the experiment, 55 hr after operation. Plasma aldosterone and ANGII were measured by specific radioimmunoassays after extraction, as described previously (Sanders et al. 1992; Peters et al. 1996).
Statistical Analysis
Data on plasma aldosterone concentrations were given as means ± SEM. Differences between groups were evaluated by analysis of variance, followed by the post hoc test of Bonferroni. Only p values less than 0.05 were accepted to indicate significant differences.
Tissue Preparation
Rats were anesthetized with Ketamine/Xylazine (75 mg/kg and 6 mg/kg, respectively). Fixation of the adrenal glands was performed by retrograde perfusion from the abdominal aorta with 2% freshly prepared paraformaldehyde in PBS, pH 7.4, for 90 sec at a pressure of 220 mmHg and for 90 sec at a pressure of 170 mmHg, followed by perfusion with 18% sucrose in PBS, adjusted to 800 mOsm/kg for another 3 min at the same pressure level.
For in situ hybridization, the adrenal glands were removed, mounted quickly onto small pieces of styrofoam and then snap-frozen in liquid nitrogen-cooled isopentane. All tissues were stored at −80C.
In Situ Hybridization
A full-length cDNA of StAR protein was obtained from rat adrenal cDNA. Because the published cDNA sequences of rat StAR differ in their 5′-region (Lee et al. 1997; Mizutani et al. 1997), we used an adaptor-ligated cDNA, prepared from rat adrenal polyA+ RNA using the Marathon cDNA Amplification Kit (Clontech; Heidelberg, Germany) according to the manufacturer's instructions. First- and secondstrand cDNA synthesis was followed by the ligation of an adaptor to both cDNA ends. Amplification of the StAR cDNA was performed using the Expand Long Template PCR system (Boehringer; Mannheim, Germany) with a specific antisense primer (5′-GTGCAGGTAGATGTGGTGGGC-3′), hybridizing immediately downstream of the 3′-end of the StAR coding sequence and a sense primer hybridizing to the adaptor. The obtained fragment was cloned into pBluescript and verified by sequencing. A specific probe for detection of the CYP11B2 transcript was also generated by PCR from the full-length cDNA, which had been prepared from the rat adrenal cDNA. The used sense (5′-GGGATGTCATCTCTGAGTATG-5′) and anti-sense (5′-GATTGCTGTCGTGTCAAC-3′) primers give rise to a fragment of 185
DIG1-11-UTP-labeled sense and anti-sense riboprobes were synthesized from the linearized plasmids using T3 and T7 RNA polymerases, respectively. In situ hybridization was carried out as described previously (Obermüller et al. 1997). Cryostat sections (6 μm thick) were transferred onto silanecoated glass slides. Sections were postfixed in 4% paraformaldehyde (in PBS, pH 7.4) for 20 min, rinsed three times in PBS, and washed in DEPC-treated bidistilled water for 10 min. A mild deproteinization step was performed by immersing slides in 0.1 M HCl for 10 min, followed by two short rinses (5 min each) in PBS. Slides were then acetylated for 20 min in 0.1 M triethanolamine, pH 8.0, containing 0.25% acetic anhydride. After rinsing in PBS, slides were stepwise dehydrated (8 min each in 70%, 80%, 95% ethanol) and air-dried. Sections were incubated with prehybridization solution (50% deionized formamide, 50 mM Tris-HCl, pH 7.6, 25 mM EDTA, pH 8.0, 20 mM NaCl, 0.25 mg/ml tRNA from yeast, 2.5 × Denhardt's solution) at 46C for 2 hr, followed by incubation for 16 hr at 42C in a moist chamber with 25 μl hybridization mixture (final concentrations: 50% deionized formamide, 20 mM Tris-HCl, pH 7.6, 1 mM EDTA, pH 8.0, 0.33 M NaCl, 0.2 M DTT, 0.5 mg/ml tRNA, 0.1 mg/ml sonicated denatured DNA from fish sperm, 1 × Denhardt's solution, 10% dextran sulfate, and denatured riboprobe 10 ng/μl), covered with a siliconized coverslip. Slices were washed once in 2 × SSC (1 × SSC = 150 mM NaCl, 15 mM sodium citrate, pH 7.0) at room temperature for 30 min, followed by 1 hr at 49C in 1 × SSC and 0.5 × SSC with 50% formamide each, and then rinsed at room temperature in 0.5 × SSC for 15 min, in 0.2 × SSC for 10 min, and equilibrated twice for 5 min in Tris buffer (100 mM Tris-HCl, 150 mM NaCl, pH 7.4). Incubation of slides with alkaline phosphatase-coupled anti-digoxigenin antibody (dilution 1:500) as well as color reactions were performed according to the manufacturer's protocol (Boehringer).
The specificity of the obtained in situ hybridization signal was verified by parallel incubation with anti-sense and sense riboprobes on alternate sections. Throughout all experiments, sense probes did not produce any detectable signal.
Results
Plasma ANGII and Aldosterone Concentrations
After a period of 55 hr, plasma ANGII levels were elevated about tenfold by ANGII infusion and decreased to less than 30% of controls by bilateral nephrectomy (sham 59 ± 13 pg/ml; ANGII infusion: 607 ± 110 pg/ ml; NX 18 ± 2 pg/ml). At 55 hr after chronic infusion of ANGII, aldosterone levels were increased (27 ± 12 pg/ml) compared with levels in sham-operated rats (11 ± 3 pg/ml). Owing to the slight differences and small number of rats, the effect did not reach significance level. In contrast, bilateral nephrectomy increased plasma aldosterone concentrations prominently and significantly (465 ± 90 ng%; p < 0.001). Corticosterone levels were not different between the groups (sham 29 ± 8 μg%; ANGII 41 ± 13 μg%; NX 48 ± 10 μg%).
Distribution of CyP11B2 and StAR mRNA in the Adrenal Glands of Control Animals
By in situ hybridization with CyP11B2 anti-sense probe, staining was observed in parenchymal cells of the zona glomerulosa only, clustered within the first two to three subcapsular cell layers (Figure 1a). No staining was observed in the capsule, the inner parts of the cortex, or in the medulla. This demonstrates the specificity of the probe used, because no crossreaction with CYP11B1 mRNA occurred.
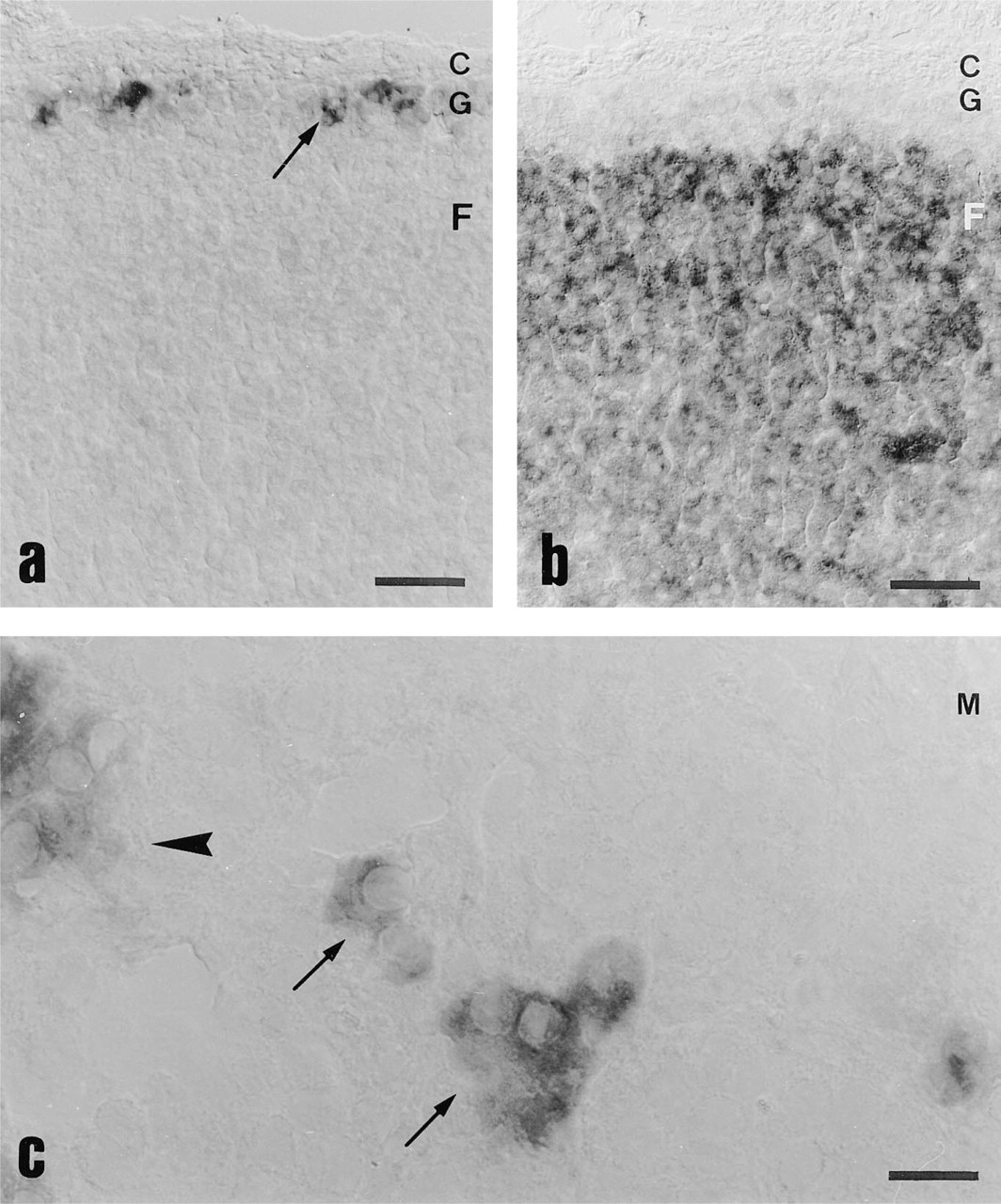
Distribution of CYP11B2 and StAR mRNA in the adrenal glands of control animals. (
In contrast, StAR anti-sense probe revealed only a faint and inhomogeneous staining in the parenchymal cells of the zona glomerulosa and intermedia (the outermost five to seven cell layers) (Figure 1b), with a slight preference for the subcapsular cell layer. An intense homogeneous signal was observed in parenchymal cells of the zona fasciculata and zona reticularis. In the adrenal medulla, some single cells or groups of two to three cells stained positive for StAR mRNA (Figure 1c), whereas CYP11B2 always was negative in this area.
Effect of ANGII on Expression of StAR and CYP11B2
Under stimulation with ANGII, no significant change in StAR mRNA was observed in the outer adrenal cortex (Figures 2a and 2b), while the signal per cell for CYP11B2 mRNA became more intense and the number of positive cells increased (Figures 2c and 2d). There was no increase in width of the aldosteroneproducing zone.
Effect of Nephrectomy on Expression of StAR and CYP11B2
The prominent increase of plasma aldosterone concentrations by nephrectomy was accompanied by an intense staining for StAR mRNA in the zona glomerulosa and intermedia (control, Figure 3a∗; nephrectomy, Figure 3b∗). The entire outer cortex became positive to a degree similar to that of the zona fasciculata and zona reticularis (Figure 3b). Furthermore, after nephrectomy we observed that an area of five to seven cell layers now stained intensely positive for CYP11B2 (Figure 3c∗), whereas the zona fasciculata/ reticularis and medulla still remained without staining (Figure 3c).
Discussion
StAR protein is an important factor in steroidogenesis, because there is evidence that the delivery of cholesterol to the inner mitochondrial membrane is the ratelimiting step of steroidogenesis (Simpson et al. 1972; Jefcoate et al. 1987). Although regulation of this protein is well documented in vivo for the zona fasciculata, its regulation by factors stimulating aldosterone production has thus far been investigated only in vitro. In this study we characterize the distribution of StAR mRNA in the rat adrenal gland, its induction in the zona glomerulosa and intermedia in vivo, and its contribution to changes in plasma aldosterone levels.
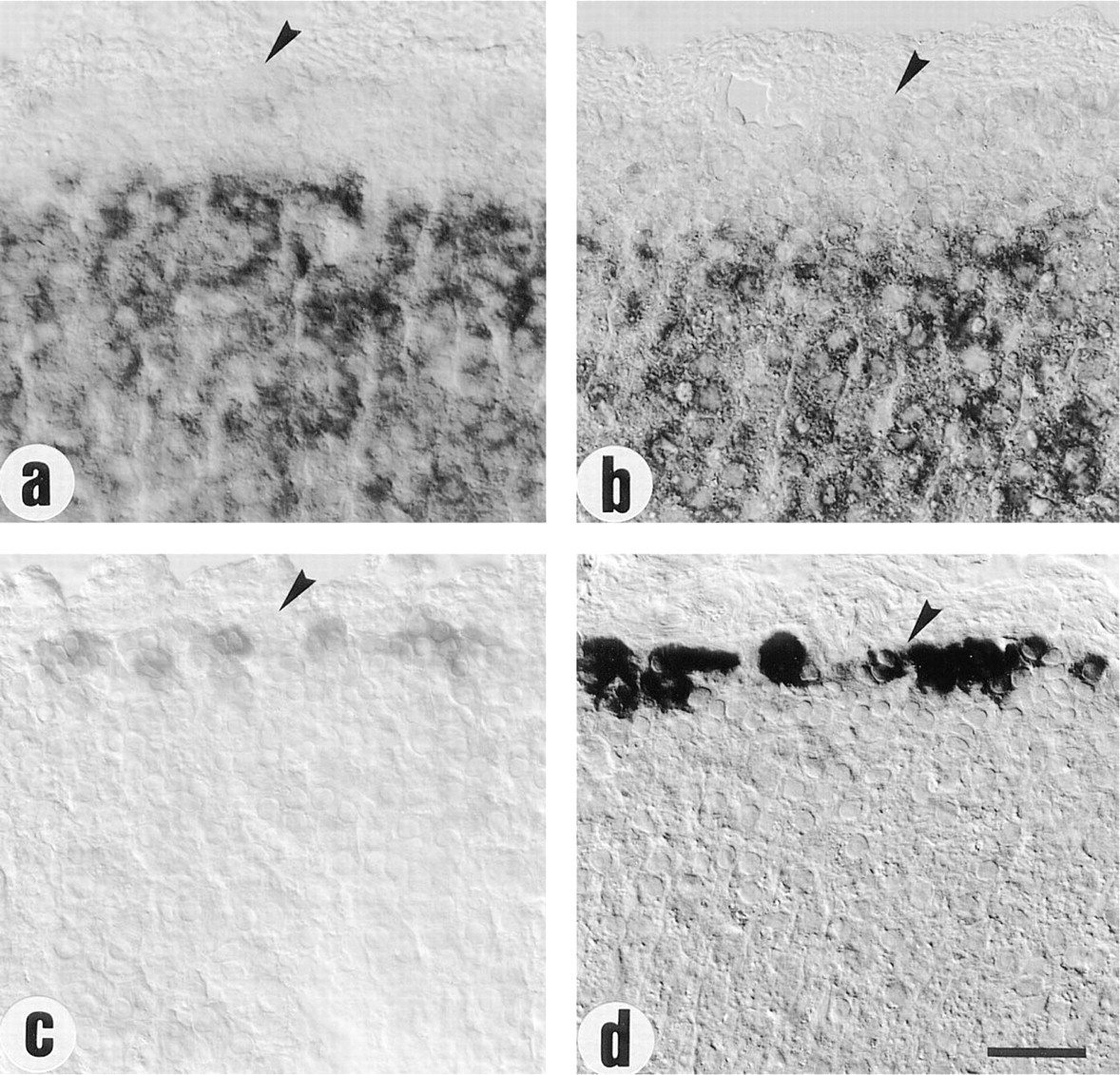
Effect of ANGII on StAR and CYP11B2 mRNA levels in the outer adrenal cortex. Representative tissue sections from glands of (
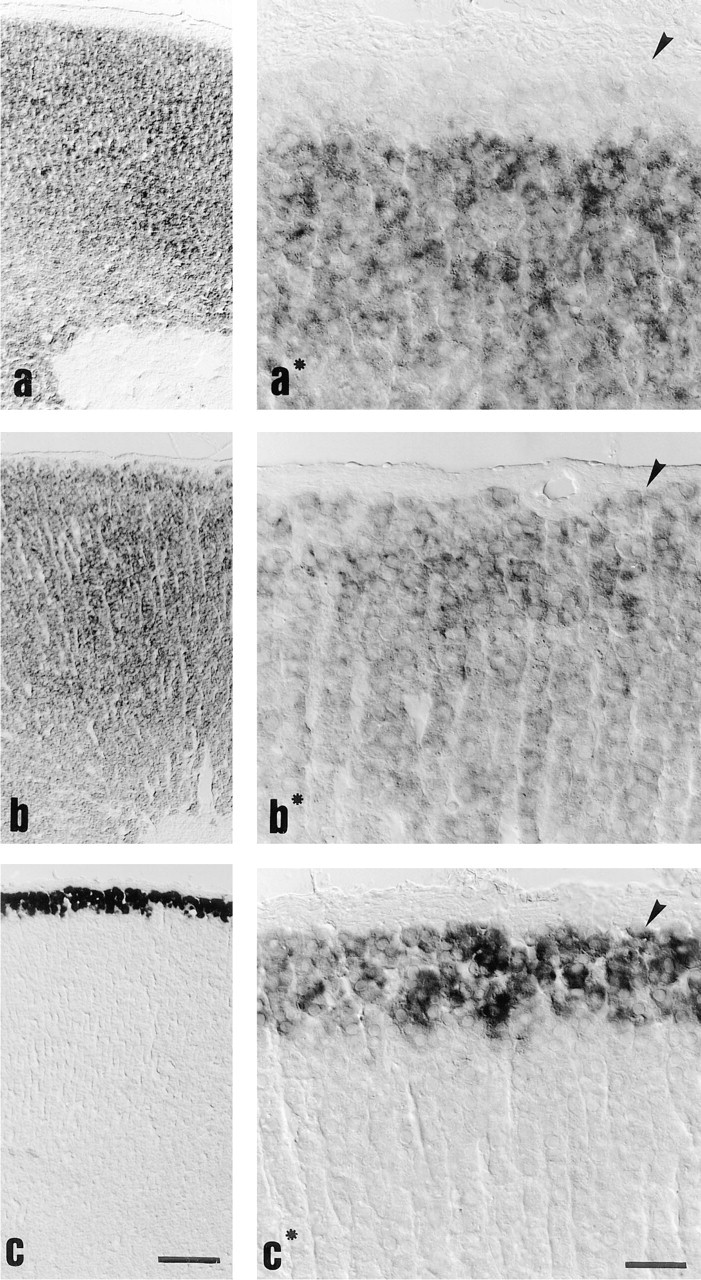
Cellular distribution of StAR mRNA compared to CYP11B2 expression after nephrectomy. (
In control rats, the StAR gene was expressed with high intensity in the zona fasciculata and zona reticularis. The zona glomerulosa and the intermediate zone exhibited only faint staining, if at all. This is in contrast to the human adrenal, in which no considerable difference between zona glomerulosa and zona fasciculata was observed by immunohistological means, whereas the zona reticularis stained to a lower extent than the outer regions (Pollack et al. 1997). In the adrenal medulla, we detected single cells with StAR gene expression, whereas aldosterone synthase was always negative in this area. The existence of steroidogenic cells in the adrenal medulla has been reported previously (Bornstein et al. 1991,1994). Expression of 17 α-hydroxylase has been observed in single cells or groups of cells in the medulla of pig and human, some being in connection to the adrenal cortex. The detection of StAR mRNA in the rat adrenal medulla supports the view that steroids are produced at that site and suggests interactions with chromaffin cells.
The pattern of StAR expression throughout the cortex and its upregulation by bilateral nephrectomy provides us with important information regarding the role of the intermediate zone. Adaptation processes in the adrenal cortex were believed to occur slowly and to involve proliferation of cells. Recently, Mitani et al. (1996) described the effects of long-term stimulation by combined administration of ACTH and a low-salt diet. After 2-3 weeks of stimulation, the intermediate zone decreased in width but did not completely disappear. Furthermore, replicating cells were found around the boundary of the zona glomerulosa and zona fasciculata. This suggested the presence of a stem-cell zone in this area. However, we observed that after stimulation with bilateral nephrectomy the staining for StAR was homogeneously distributed in the entire adrenal cortex, including the outermost zones. The existence of undifferentiated stem cells in the intermediate zone is therefore rather unlikely.
Associated with a 50-fold elevation of plasma aldosterone concentration, together with the rise of StAR mRNA, the CYP11B2-positive region increased from one or two to five to seven cell layers. This demonstrates that an increase in width of the aldosteroneproducing zone can happen within a short period of time and that all cells in the intermediate zone can become steroidogenically active. It also suggests that the increase in glomerulosa width in our experiment is solely the result of activation of previously quiescent cells and not of proliferation.
Surprisingly, ANGII administration did not induce a notable increase of StAR mRNA, whereas it stimulated CYP11B2 expression considerably. A stimulation of StAR protein by ANGII in vitro has been reported (Clark et al. 1995a). Probably, the induced changes in our experiment are too small to be detected by in situ hybridization. However, consistent with the absent rise of StAR mRNA, plasma aldosterone levels also did not increase significantly. Therefore, in our experiment aldosterone levels correlated even better with StAR than with CYP11B2 mRNA.
In summary, we have demonstrated changes in the distribution pattern of StAR mRNA in the rat adrenal gland after acute stimulation in vivo and concomitant changes in aldosterone production. The abundance of StAR mRNA in all layers of the outer adrenal cortex after stimulation confirms the function of the intermediate zone as a reservoir of steroidogenic cells.
Footnotes
Acknowledgments
Supported by grant Pe 366/3-2 from the Deutsche Forschungsgemeinschaft and by the Forschungsfond der Fakultät für Klinische Medizin Mannheim of the University of Heidelberg.
We thank Hiltraud Hosser for technical support and I. Ertel for photographic work.