
Abstract
We investigated the intracellular localization of CRH in transiently transfected COS-7 cells expressing the full-length rat corticotropin-releasing hormone (CRH) precursor cDNA. CRH synthesized by transfected COS-7 cells is mainly stored intracellularly. In contrast, CHO-K1 cells expressing the same CRH precursor stored and released equal amounts of immunoreactive (IR)-CRH. Ultrastructural analysis revealed that CRH is stored in electron-dense aggregates in the RER of transiently transfected COS-7 cells and does not migrate into the Golgi apparatus. On the basis of the different intracellular localization, storage, and release of CRH in COS-7 and CHO-K1 cells, we hypothesize that the intracellular trafficking of CRH within the constitutive secretory pathway for protein secretion not only depends on its primary amino acid sequence but might also be influenced by intracellular conditions or factors.
Keywords
I
The establishment of the stably transfected CHO-K1 cell line expressing the rat CRH precursor has been previously described (Castro et al. 1995b). COS-7 cells were transiently transfected using the plasmid pGWH1/rCRH. Both expression vectors encode exactly the same full-length rat CRH precursor cDNA under the control of exactly the same sequence of the major immediate early human cytomegalovirus (HCMV) promoter. The data shown in Tables 1 and 2 compare the storage and secretion of CRH within stably transfected CHO-K1 cells and transiently transfected COS-7 cells expressing the CRH precursor. Transiently transfected COS-7 cells store more than 90% of total immunoreactive (IR)-CRH (Table 1). The transfection efficiency for COS-7 cells is 40-60% (results not shown). In contract, stably transfected CHO-K1 cells store and release approximately equal levels of IR-CRH (Table 2). It is possible that some IR-CRH could also be recovered in the medium due to cell lysis, which in both cell types at the time of assaying was 5%, as assessed by trypan blue exclusion. To investigate the trafficking of the CRH precursor within the secretory pathway, i.e., constitutive vs regulated pathway in transfected COS-7 and CHO-K1 cells, we stimulated the cells with 10−5 M forskolin, 10−3 M 8-Br-cAMP, and 51mM KCl. None of these secretagogues stimulated the release of IR-CRH (results not shown), thus indicating targeting of proCRH to the constitutive secretory pathway.
IR-CRH stored and released in transiently transfected COS-7 cells (mean ± SEM) a
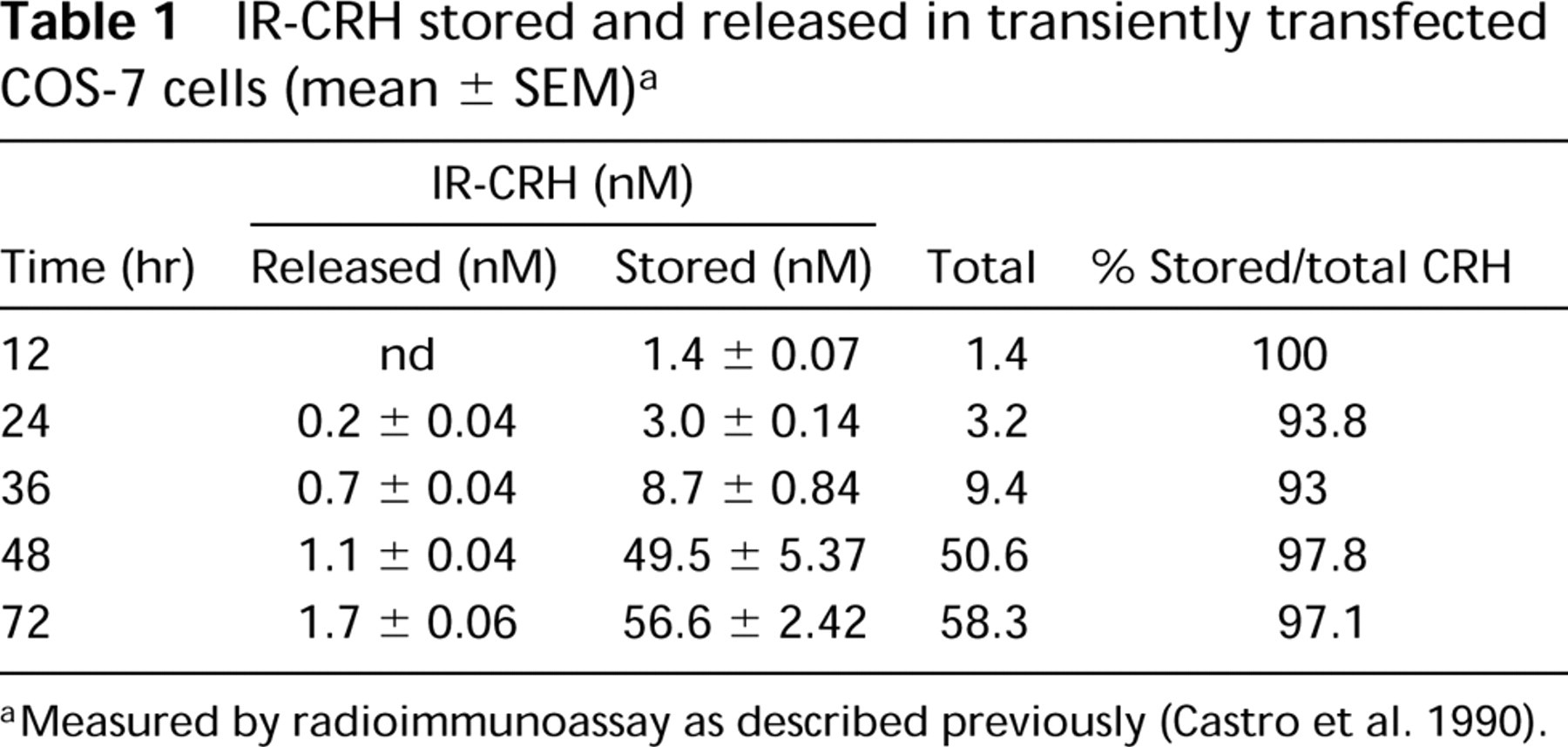
Measured by radioimmunoassay as described previously (Castro et al. 1990).
IR-CRH stored and released in stably transfected CHO-K1 cells (mean ± SEM) a
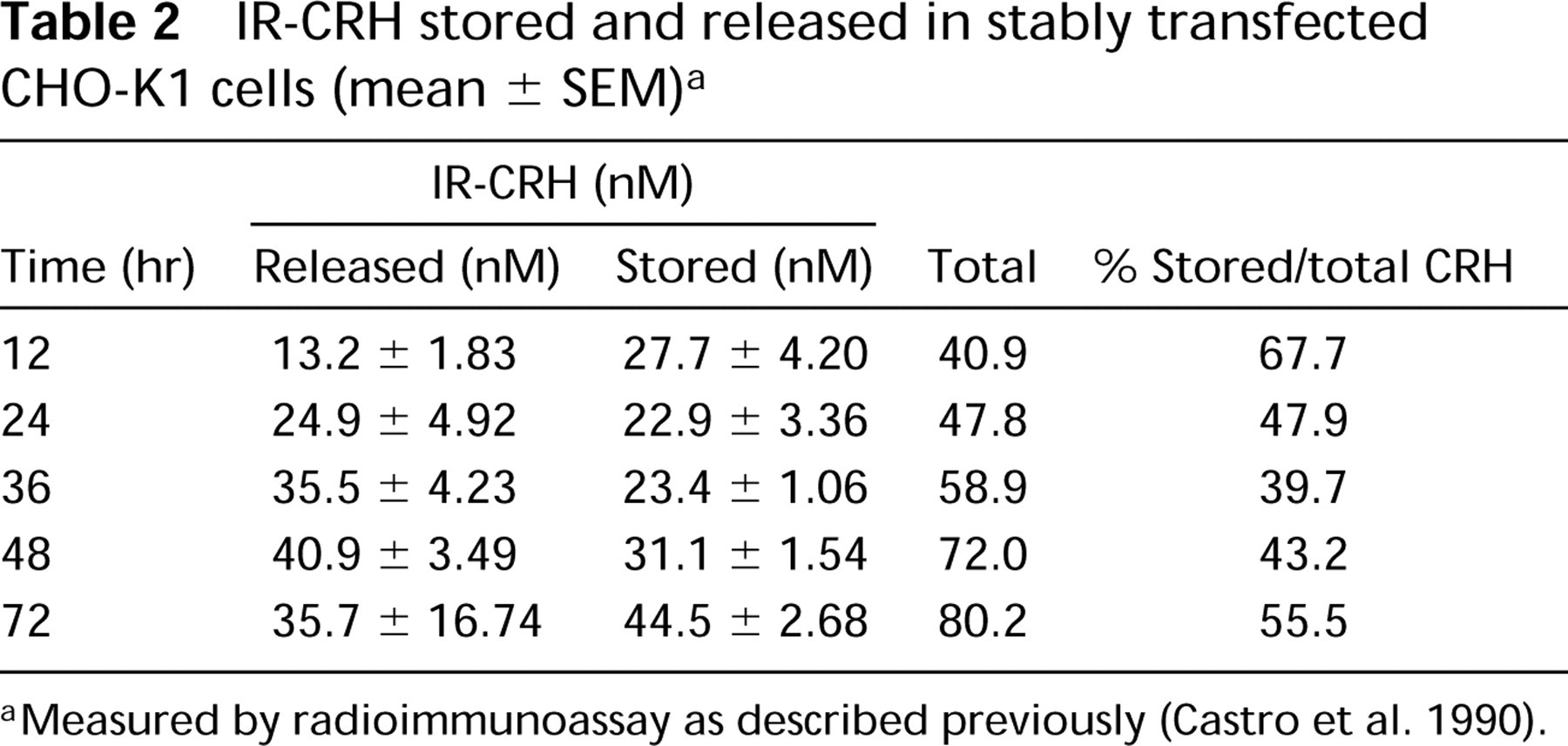
Measured by radioimmunoassay as described previously (Castro et al. 1990).
To investigate the molecular forms of CRH synthesized by transiently transfected COS-7 cells, cell extracts were subjected to Western blotting as previously described (Castro et al. 1996) (data not shown). Three different anti-CRH antibodies (Matthew, 3B3, and Hannah) were employed. Two bands of approximately 21 and 19 kD were detected. The former band could account for the full-length CRH precursor and the latter for the CRH precursor devoid of its signal peptide. No other bands corresponding to smaller CRH peptides were observed, indicating that the CRH precursor did not undergo endoproteolytic processing within COS-7 cells.
To determine the time course of biosynthesis of CRH, COS-7 cells were metabolically labeled using [35S]-methionine/[35S]-cysteine at 48 hr post transfection. CRH was detected using immunoprecipitation followed by SDS-PAGE and autoradiography as described previously (Castro et al. 1995a) (Figure 1). IR-CRH was detectable in cell extracts at 20 min after labeling and accumulated with time, but did not appear in culture supernatant until 2 hr after labeling. The major immunoreactive product observed in both cell extracts and culture medium was a band of 19 kD. The higher molecular weight band of 21 kD appeared in cell extracts after 4 hr labeling (Figure 1B, Lane 7). No other smaller molecular weight CRH-derived peptides, such as CRH(1-41), were detected. No bands corresponding to IR-CRH were seen in extracts or culture medium of nontransfected COS-7 cells metabolically labeled for 4 hr.
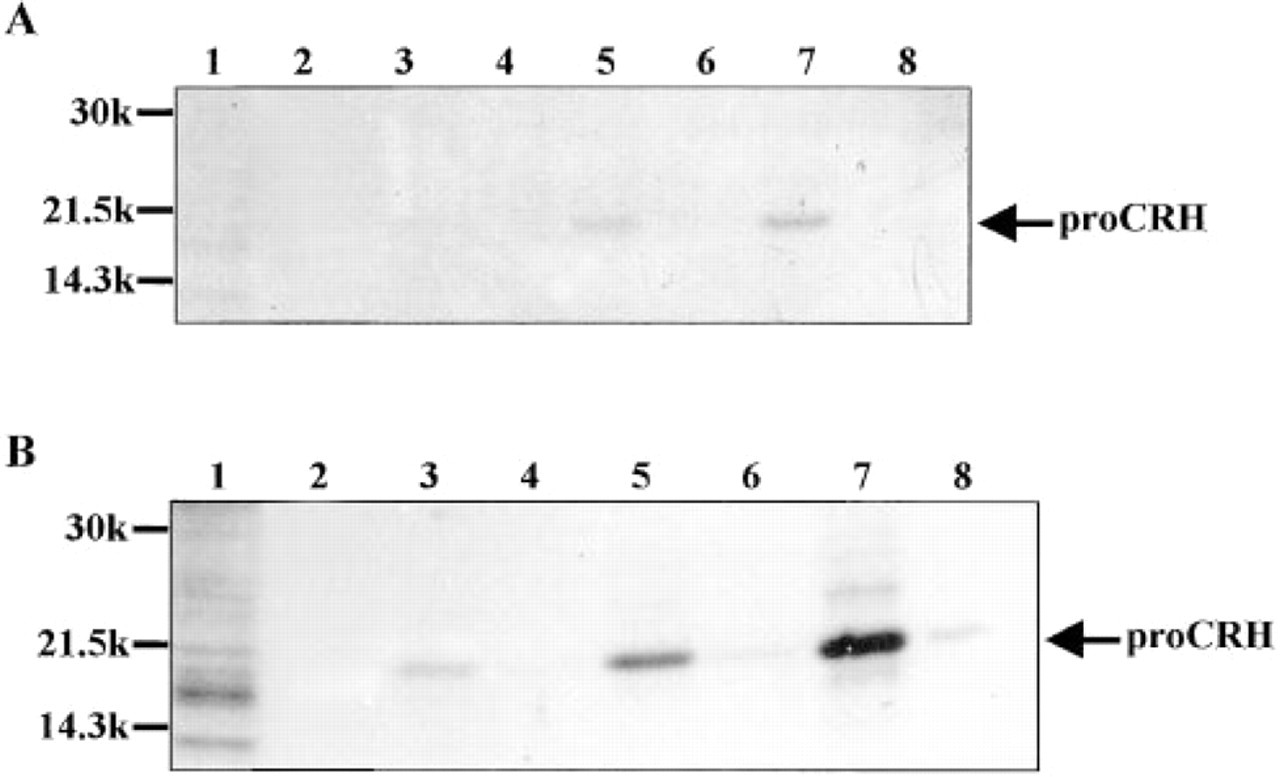
Metabolic labeling of transiently transfected COS-7 cells followed by SDS-PAGE and autoradiography.
To elucidate the intracellular localization of IR-CRH within transiently transfected COS-7 cells, we performed immunoelectron microscopy. Figure 2A shows a transfected COS-7 cell next to an untransfected cell. The transfected cell contains the electron-dense structures within tubular membrane-bound compartments (Figures 2A, 2B, and 2D). The membranes are studded with ribosomes, identifying the organelle as the RER. Immunogold particles surrounding electron-dense aggregates confirm that they are immunoreactive for CRH (Figure 2C). Figure 2D shows the Golgi apparatus (G), which does not contain any electron-dense structures (arrows). Therefore, in transfected COS-7
Electron microscopy of COS-7 cells transfected with the rat CRH precursor cDNA.