
Abstract
We examined the expression of regulated endocrine-specific protein of 18-kD (RESP18) in selected peptidergic and catecholaminergic neurons of adult rat brain. In the hypothalamic paraventricular, supraoptic, and accessory nuclei, RESP18 mRNA was highly expressed in neurons immunostained for oxytocin and vasopressin. RESP18 mRNA was also highly expressed in paraventricular nucleus neurons immunostained for corticotropin-releasing hormone, thyrotropin-releasing hormone, and somatostatin. RESP18 mRNA was expressed in POMC cells of the arcuate nucleus, in neuropeptide Y cells of the dorsal teg-mental nucleus, lateral reticular nucleus, and hippocampus, and in brainstem catechola-minergic neurons. RESP18 mRNA expression was high in all paraventricular and arcuate neurons, but RESP18 protein was detectable in the perikarya of a subset of these neurons, suggesting an important post-transcriptional component to the regulation of RESP18 expression. RESP18 antisera immunostained perikarya but not axon fibers or terminals. Sub-cellular fractionation of homogenates of several hypothalamic nuclei identified RESP18 protein in fractions enriched in endoplasmic reticulum. The presence of 22- and 24-kD RESP18 isoforms in the neural lobe of the pituitary indicated that some RESP18 protein exited the endoplasmic reticulum. The post-transcriptional regulation of RESP18 expression and localization of RESP18 protein primarily to the endoplasmic reticulum suggests that RESP18 plays a regulatory role in peptidergic neurons.
Keywords
R
RESP18 expression is limited to brain, pituitary, peripheral endocrine tissues, and sperm, suggesting that its function is specific to neurons, endocrine cells, and sperm (Bloomquist et al. 1994a,b; Darlington et al. 1996a,b; Schiller and Darlington 1996). In rat pituitary, RESP18 mRNA has been identified in corticotropes, thyrotropes, gonadotropes, somatotropes, and mammotropes of the anterior lobe and in melanotropes of the intermediate lobe (Bloomquist et al. 1994b). RESP18 is expressed in several endocrine cell lines, including AtT-20, GH3, PC12, and RIN cells, but not in fibroblast lines.
Although the function of RESP18 is unknown, expression of RESP18 mRNA is regulated in a cell type-specific manner. In rat melanotropes, the levels of RESP18 mRNA decrease in response to dopaminergic agonists. Dexamethasone increases RESP18 mRNA levels and protein biosynthesis in AtT-20 cells. Insulin, estradiol, and epidermal growth factor treatment lowers RESP18 mRNA levels in GH3 rat somatomammo-trope cells, and nerve growth factor treatment increases RESP18 mRNA levels in rat pheochromocytoma PC12 cells (Bloomquist et al. 1994a,b; Lee et al. 1995).
Recently, RESP18 mRNA has been localized to areas of the brain that control neuroendocrine and autonomic function, including the paraventricular, supraoptic, suprachiasmatic, dorsomedial, ventromedial, and arcuate nuclei of the rat hypothalamus, and the substantia nigra, ventral tegmental nucleus, amygdala, and hippocampus (Darlington et al. 1996a). Most of these neural structures are known to synthesize and secrete peptides such as vasopressin (AVP), oxytocin (OXY), corticotropin-releasing hormone (CRH), thyrotropin-releasing hormone (TRH), neuropeptide-Y (NPY), pro-opiomelanocortin (POMC), somatostatin, and catecholamines (Zimmerman and Nilaver 1984; Sawchenko et al. 1988; Swanson 1991; Armstrong 1995) and we set out to determine if RESP18 is expressed in these neurons. Using a combined in situ hybridization/immunohistochemical approach, we found RESP18 mRNA in AVP, OXY, CRH, TRH, NPY, POMC, somatostatin, and catecholaminergic neurons. We also identified neurons with substantial amounts of RESP18 mRNA and very little RESP18 protein among neurons with high levels of RESP18 mRNA and protein. This discrepancy suggests that post-transcriptional regulation of RESP18 expression occurs in neurons. Finally, Western blot analysis, subcellular fractionation, and immunostaining suggest that much of the RESP18 is located in the endoplasmic reticulum, with small amounts accessing distal parts of the secretory pathway.
Materials and Methods
Thirty-seven adult male Sprague–Dawley rats (300–350 g) were anesthetized with pentobarbital sodium (50 mg/kg) and perfused with 200 ml of phosphate-buffered 4% formaldehyde solution (4C) via the left ventricle. The brain was removed, blocked and cut, and placed in phosphate-buffered 4% formaldehyde (4C) for 1 day, followed by 25% sucrose (4C) for at least 7 days. All animals were handled in accordance with the University of Maryland at Baltimore Institutional Animal Care and Use Committee.
In Situ Hybridization
Frozen sections (12 μm) were placed on gelatin-coated glass slides for RESP18 in situ hybridization using RESP18 riboprobes as previously described (Bhat et al. 1992; Bloomquist et al. 1994a; Darlington et al. 1996a). Full length sense and anti-sense riboprobes were synthesized from linearized pBS.RESP plasmid using T3 and T7 RNA polymerase, respectively, and 125 μCi of [35 S]-UTPγS (1000 Ci/mmol; Du-Pont–NEN, Boston, MA). Probe (2 × 106 cpm) diluted in 50 μl of hybridization buffer (50% formamide, 0.6 M NaCl, 10 mM Tris-HCl, 1 mM EDTA, 1 × Denhardt's, 0.2 mg/ml of yeast tRNA, 10% dextran sulfate, 10 mM dithiothreitol, pH 7.4) was placed on each slide, covered with parafilm, and incubated at 56C overnight. After incubation the sections were rinsed in 2 × SSC and washed sequentially in 2 × SSC with 1 μg/ml of RNAse A at 37C (Worthington Biochemicals; Freehold, NJ) (1 hr), 2 × SSC (30 min), 1 × SSC (30 min), 0.5 × SSC (30 min), 0.2 × SSC (10 min), 0.2 × SSC at 60C (60 min), 0.2 × SSC at room temperature (RT; 10 min), and PBS (10 min).
Immunocytochemistry
Tissue sections processed for in situ hybridization were then analyzed by immunocytochemistry. The tissue sections were blocked in 10% normal goat serum/PBS (30 min) and 3% H2O2/PBS (10 min) and were then rinsed in PBS for 10 min. Slides were then incubated overnight at 4C in 100 μl of rabbit polyclonal primary antiserum diluted in 10% normal goat serum/PBS. Primary antibodies were visualized using an avidin–biotin kit with 3,3′-diaminobenzidine as the substrate (Vector Laboratories; Burlingame, CA). No distinct immunostaining was present when RESP18 antiserum was preincubated with purified RESP18 antigen (50 ng/ml) or when control rabbit serum was used. No distinct immunostaining was present in any neurons when primary antiseruma was omitted from the immunocytochemical protocol.
Sections were rinsed and sequentially dehydrated (70%, 95% ethanol), air-dried, placed on film for 1–3 days, dipped in Kodak NTB-3 photographic emulsion (Eastman Kodak; Arlington, VA), exposed for 10–14 days, and developed. The sections were rinsed in ddH2O, counterstained with 0.4% methyl green for 1 min, rinsed in ddH2O, destained with 70% ethanol, dehydrated (95%,100% ethanol) into xylene, and coverslipped.
Antisera
We used rabbit polyclonal antisera to RESP18 (JH1162 and JH1163, 1:4000 dilution), POMC (Georgie, 1:2000), AVP and OXY (V4 and V10, 1:1000 dilution; gift of Dr. Hal Gainer, National Institutes of Health), somatostatin (SS175c, 1:5000 dilution; gift of Dr. Bryan Noe, Emory University), CRH (1:1000 dilution), TRH (1:1000 dilution; gift of Dr. Eduardo Nillni, Brown University), dopamine β mono-oxygenase (DβM, JH2049, 1:5000 dilution), neuropeptide Y (JH3, 1:2000 dilution), and BiP/GRP78 (1:2000; Affinity BioReagents, Neshamic Station, NJ) (Eipper and Mains 1978; Bloomquist et al. 1994a,b; Darlington et al. 1996a; Paquet et al. 1996). We also used monoclonal antisera to synaptophysin (#902 314, 1:1500; Boehringer Mannheim, Indianapolis, IN) and synaptotagmin (1:3000 dilution; gift of Dr. Richard Scheller). Synaptotagmin was used as a marker for large dense-core vesicles, small dense-core vesicles, synaptic vesicles, and plasma membrane sheets (Calakos and Scheller 1996). Synaptophysin was used as a marker for synaptic vesicles, some of which may be associated with plasma membrane sheets in nerve terminals (Calakos and Scheller 1996). Goat anti-rabbit and goat anti-mouse immunoglobulins linked to horseradish peroxidase were from Amersham (Arlington Heights, IL).
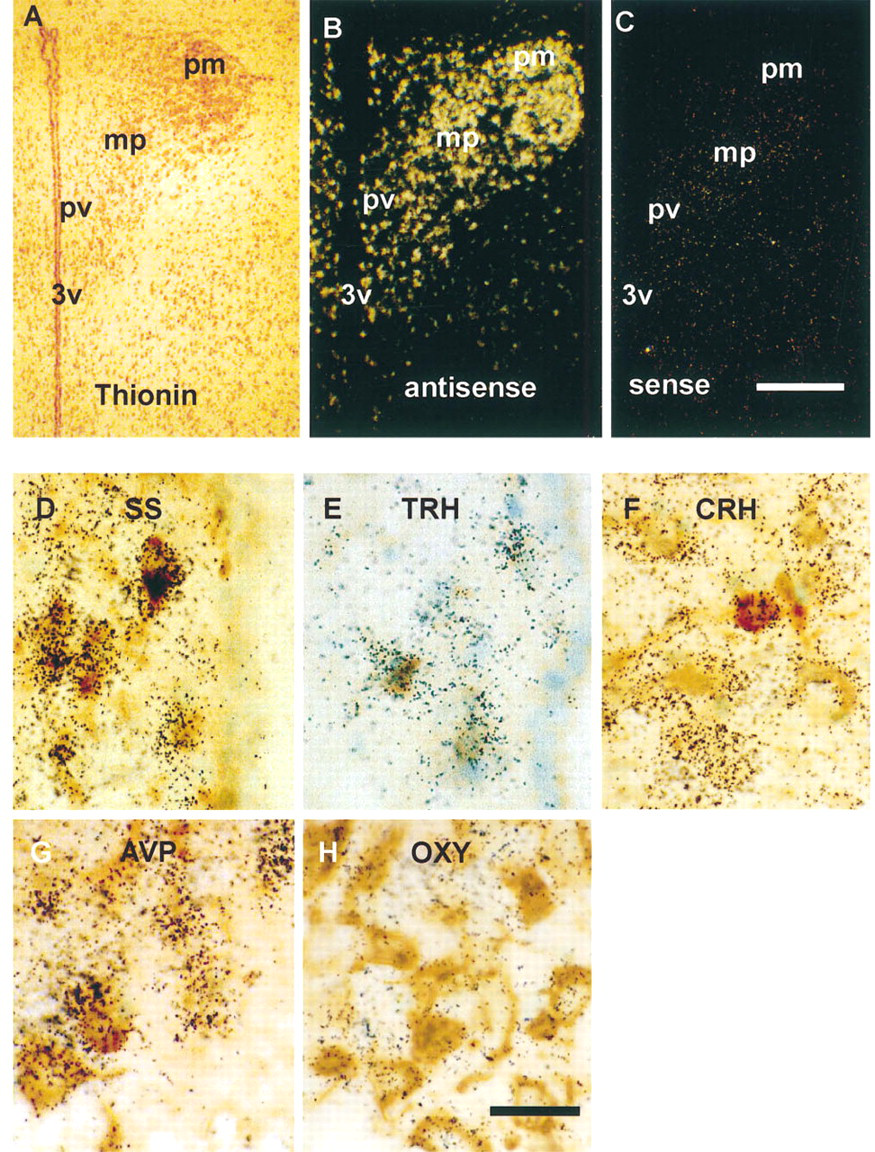
RESP18 expression in the hypothalamic paraventricular nucleus of the rat. (
Western Blot Analysis
Punches (1.5-mm diameter; 4-mm-thick sections) of rat hypothalamic nuclei were processed as previously described for rat tissues (Schiller and Darlington 1996). Briefly, tissue punches were brought to 10% (w/v) in 20 mM NaTES, 10 mM mannitol, pH 7.4, containing a protease inhibitory cocktail supplemented with 100 μg/ml N-acetyl-leucine-leucine-norleucinal (TM), freeze/thawed three times, and centrifuged at 1000 × g for 5 min to remove cell debris (Schiller and Darlington 1996). Supernatants were centrifuged at 436,000 × g for 15 min and pellets were resuspended in a volume of TM buffer containing 1% Triton X-100 equal to the supernatant. Protein concentrations were measured by BCA protein assay (Pierce; Rockford, IL) and extracts were analyzed by Western blot.
Differential centrifugation of homogenates of hypothalamic nuclei was as previously described for analysis of stable cell lines (Bloomquist et al. 1994b). Briefly, tissues were homogenized (10% w/v) in buffer (0.25 M sucrose, 1 mM MgCl2, 1 mM Na2EDTA, 10 mM HEPES, pH 7.4) containing protease inhibitors as above. Homogenization continued until a least 50% of the cells were lysed, as assessed after staining with 0.4% trypan blue. Remaining intact cells, nuclei, and debris were removed by centrifugation at 1100 × g for 5 min at 4C. The supernatant was then centrifuged at 4400 × g for 15 min (10K rpm), then at 17,400 × g (20K), 39,200 × g (30K), and 436,000 × g (100K). Pellets were re-suspended in TM buffer and aliquots were boiled for 5 min in 1 × Laemmli sample buffer. SDS-PAGE and Western blot analysis were carried out as previously described (Bloomquist et al. 1994b). Low range prestained molecular weight markers (Amersham) were used for slab gels (16.4% acrylamide, 0.60% N,N'-methylenebisacrylamide). Proteins were transferred to Polyscreen membranes (DuPont–NEN) for 1500 mAmp/hr. Blots were incubated with one of the following antisera: RESP18 (JH1162), BiP, synaptotagmin, synaptophysin. Antisera were visualized using the ECL detection system according to the manufacturer's protocol (Amersham).
Results
We set out to determine if RESP18 was expressed in AVP, OXY, CRH, TRH, somatostatin, POMC, neuropeptide-Y, and catecholaminergic neurons in the different regions of the rat brain using in situ hybridization for RESP18 mRNA and immunocytochemistry for the neuropeptides. In general, RESP18 mRNA was found in all neurons that immunostained these peptides. However, RESP18 mRNA was not restricted to these neurons and message was found in neurons that did not immunostain for these peptides.
Messenger RNA for RESP18 was found at high levels throughout the PVN (Figures 1A and 1B). In situ hybridization using sense probes for RESP18 established the background signal in the PVN (Figure 1C) and other parts of the brain (not shown). In the periventricular division of the PVN, RESP18 mRNA was found in neurons that immunostained for somatostatin (Figure 1D), TRH (Figure 1E), and CRH (not shown). RESP18 mRNA was also found in neurons that immunostained for CRH in the medial (Figure 1F), dorsal, and lateral parvocellular divisions. RESP18 mRNA was found in vasopressin and oxytocin neurons of the posterior (Figures 1G and 1H) and anterior magnocellular division of the PVN.
RESP18 mRNA was also found at high levels in all neurons of the supraoptic nuclei (Figure 2). RESP18 mRNA was highly expressed in the magnocellular neurons that immunostained for oxytocin (Figures 2A–2C) and AVP (Figures 2D–2F). There was a high coincidence of oxytocin and AVP staining with elevated levels of RESP18 mRNA (Figures 2C and 2F). RESP18 immunostaining was strong in all magnocellular neurons of the SON and accessory nuclei in a manner similar to the magnocellular neurons of the PVN (not shown).
RESP18 mRNA was highly expressed in the arcuate nucleus (Figure 3). Most of the neurons in the arcuate nucleus contained RESP18 mRNA (Figure 3A). Using in situ hybridization for RESP18 combined with POMC immunostaining, we found high levels of RESP18 mRNA expression in POMC neurons (Figures 3B and 3C). RESP18 mRNA was also found in arcuate neurons that do not immunostain for POMC and correspond to dopamine neurons by location. RESP18 mRNA was also found in the pars tuberalis and a few scattered cells in the median eminence (Figure 3A).
RESP18 mRNA was expressed at lower levels in hippocampal and brainstem neurons. Because neuropeptide Y is expressed in these regions, the distributions of RESP18 mRNA and NPY immunostaining were compared (Figure 4). RESP18 mRNA was found in neurons that immunostained for neuropeptide Y in the dorsal tegmental nucleus of the pons (Figure 4A), in NPY-containing neurons scattered in the hilar area of the dentate gyrus (Figure 4B), in all layers of CA3, and in the stratum pyramidale and oriens of CA1, and in NPY-containing neurons in the lateral reticular nucleus of the medulla oblongata (Figure 4C). Each NPY-containing neuron identified contained RESP18 mRNA.
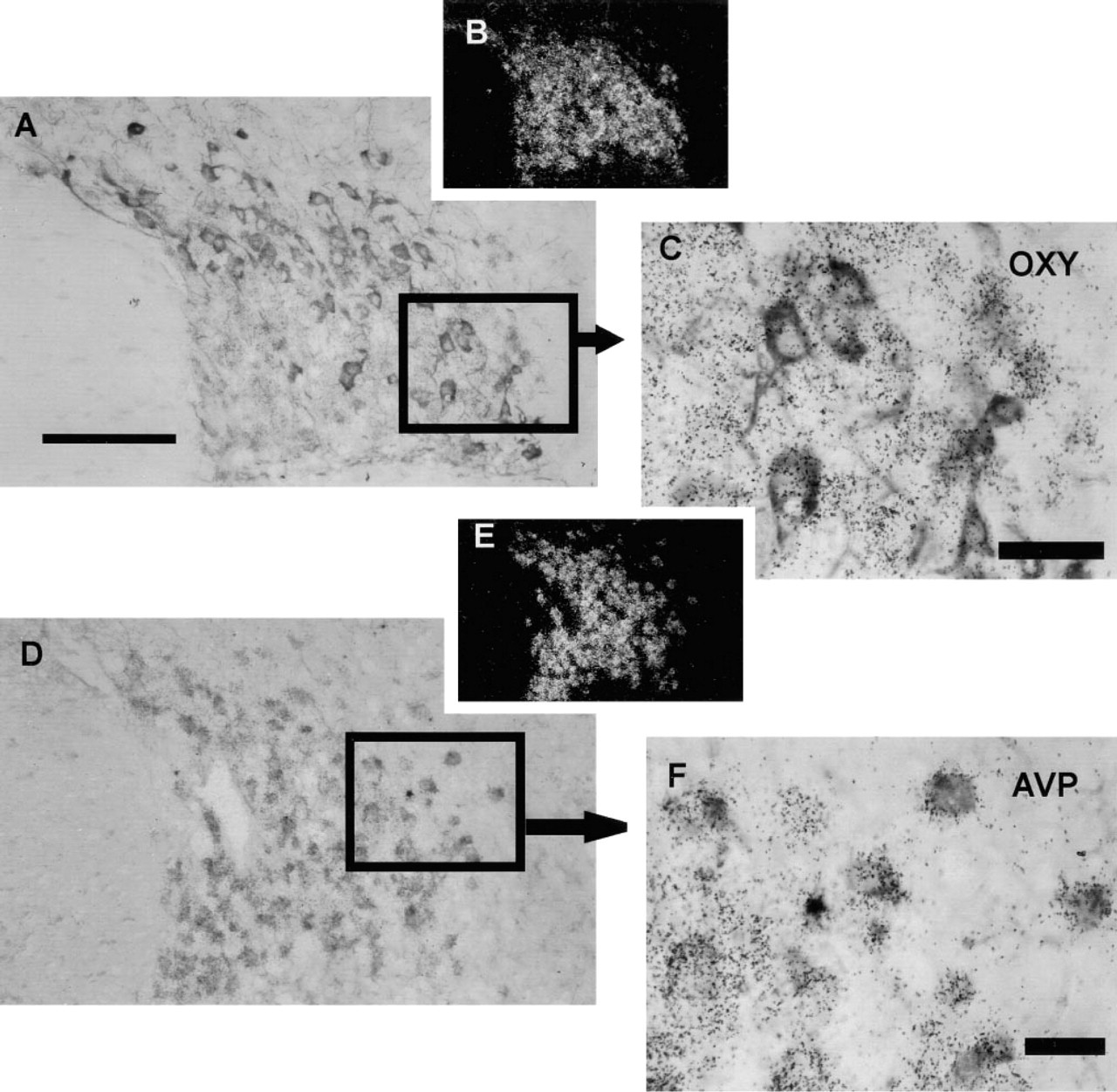
RESP18 expression in the supraoptic nucleus of the rat. Lightfield (
RESP18 mRNA was localized to brainstem regions rich in catecholaminergic neurons. In particular, RESP18 mRNA was localized to the region of the A1, A2, A5, and A6 catecholamine cell groups of the brainstem. Immunostaining for DβM, one of the enzymes required for biosynthesis of norepinephrine, was used to identify noradrenergic neurons in these regions. We found RESP18 mRNA in neurons of the ventrolateral reticular nucleus of the medulla oblongata (A1, Figure 5A), the nucleus of the solitary tract (A2, Figure 5B), the ventrolateral pons (A5, Figure 5C), and locus ceruleus (A6, Figure 5D). All neurons that immunostained for DβM contained RESP18 mRNA. However, RESP18 mRNA was not restricted to catecholaminergic neurons in these nuclei, as non-catecholaminergic neurons also contained RESP18 mRNA (Figure 5, white arrows).
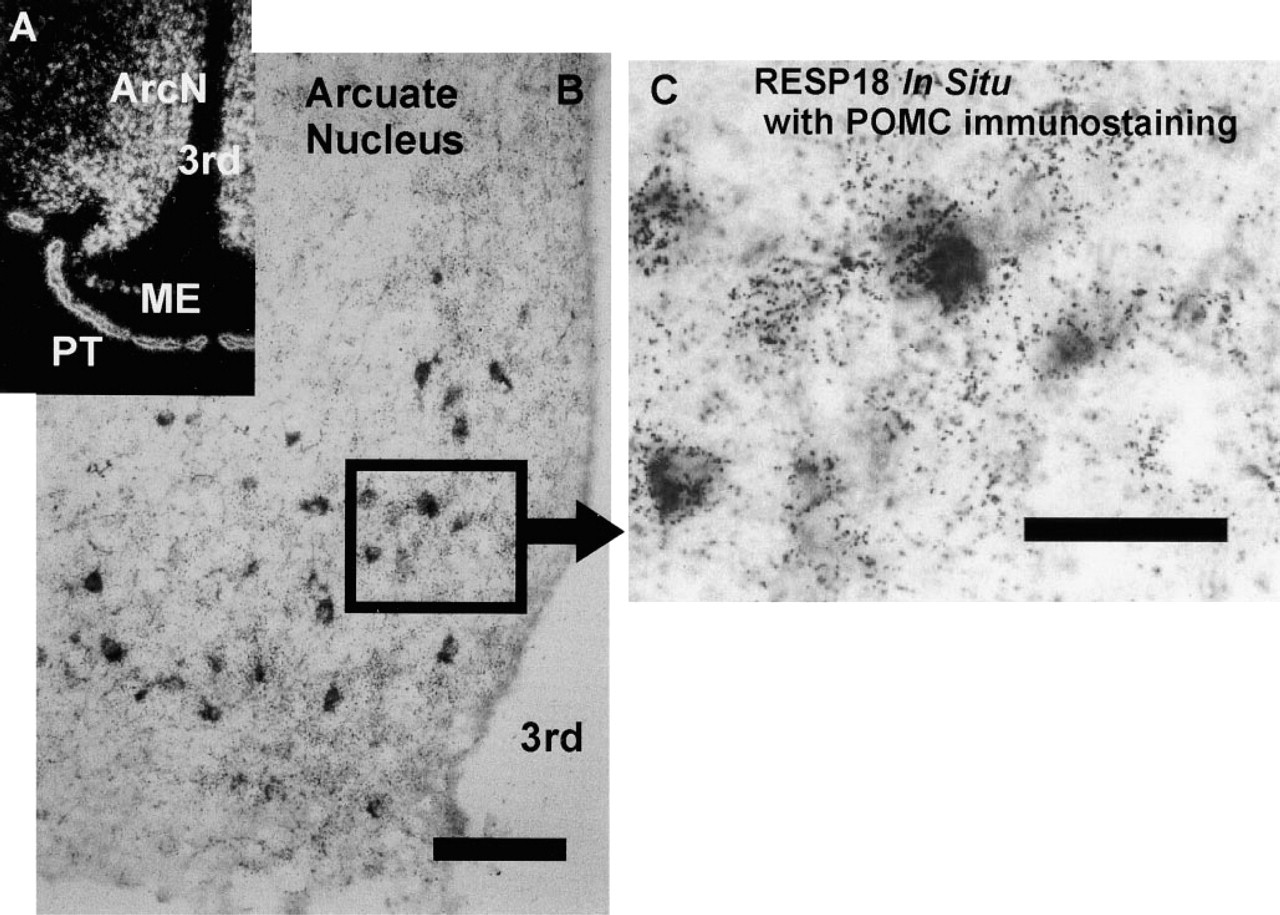
RESP18 expression in the arcuate nucleus of the rat. (
RESP18 expression is regulated at a post-transcriptional level. Although RESP18 mRNA is abundant throughout the PVN and arcuate nucleus (Figure 6), not all of the neurons in these regions could be immunostained for RESP18 protein (Figure 6). This discrepancy was observed using antisera directed towards two different epitopes in RESP18 (JH1162 or JH1163). In the PVN, RESP18 immunostaining was observed most readily in neurons of the posterior and anterior magnocellular PVN and in neurons of the periventricular division (Figures 6A–6D). This pattern of RESP18 immunostaining matches the distribution of AVP and OXY neurons in the magnocellular divisions and somatostatin neurons in the periventricular division. The medial parvocellular (Figures 6A, 6C, and 6D; mp), and lateral parvocellular (not shown) divisions that contain most of the CRH neurons exhibited little or no RESP18 immunostaining (Figure 6C). This suggests that RESP18 protein levels are low in the parvocellular divisions compared to the magnocellular divisions.
In the arcuate nucleus, RESP18 immunostaining was found only in isolated neurons, whereas RESP18 mRNA was abundant throughout the nucleus (Figures 6E and 6F); the location of RESP18 immunopositive neurons corresponds to POMC neurons. Other parts of the arcuate nucleus contained little or no RESP18 immunostaining, including the medial and ventral areas known to contain dopaminergic neurons. This suggests that RESP18 protein levels are high in POMC neurons and low in all other neurons in the ArcN. This lack of correlation of RESP18 mRNA and RESP18 protein in the PVN and ArcN indicates that expression of RESP18 is controlled at the post-transcriptional level.
RESP18 immunostaining is confined to the perikarya of RESP18-positive neurons. By comparing the pattern of immunostaining of several peptide hormones with the immunostaining of RESP18, we found that RESP18 protein was not located in neuronal processes that anatomically correspond to neuronal axons and terminals. In the arcuate, supraoptic, and paraventricular nuclei, immunostaining for POMC, oxytocin, and somatostatin could be seen in perikarya, axons, and terminals (Figures 7A, 7C, and 7E, dark arrows). However, RESP18 immunostaining was confined to cell bodies and the proximal region of dendritic processes in the same nuclei (Figures 7B, 7D, and 7F, white arrows). In no instance did RESP18 antisera visualize long processes (axons) or terminal boutons as demonstrated by the immunostaining of POMC, oxytocin, or somatostatin. These findings suggest that the intracellular distribution of RESP18 protein in neurons differs from that of POMC, oxytocin, or somatostatin.
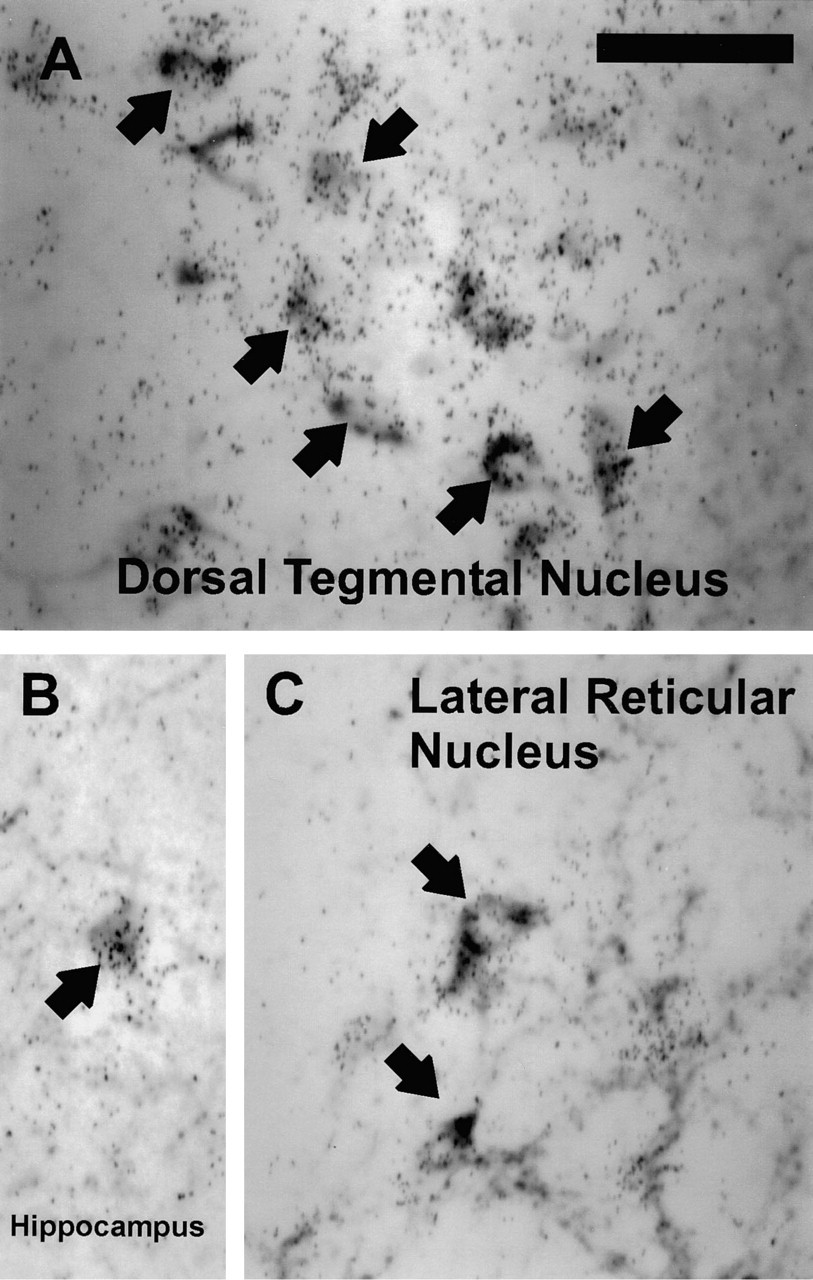
RESP18 mRNA expression in neuropeptide Y (NPY) neurons in the (
Western blot analysis of total protein extracts prepared from punches of the PVN, SON, and arcuate nuclei identified 18-kD RESP18 as the major form present (Figure 8A). Only small amounts of the 22-kD isoform of RESP18 were detected in arcuate nucleus extracts. Homogenates of hypothalamic punches were fractionated by differential centrifugation and analyzed by Western blot (Figure 8B). Heavy chain-binding protein (BiP), an endoplasmic reticulum marker, was observed as the expected 78-kD band concentrated in the 10K pellet and to a lesser extent in the 20K pellet of the SON, PVN, and arcuate nucleus. The distribution of RESP18 was similar to that of BiP, concentrated in the 10K pellet and less so in the 20 K pellet, suggesting that RESP18 is found in the endoplasmic reticulum. The small amount of 22-kD RESP18 in the arcuate sample was recovered primarily in the 20K pellet. Synaptotagmin, a marker for large dense-core vesicles, small dense-core vesicles, synaptic vesicles, and plasma membrane sheets, was observed as the expected 65-kD band in all of the pellets and in the soluble fraction. Although overlapping, the distribution of synaptotagmin differed from that of RESP18. Some degradation of synaptotagmin was observed despite the presence of protease inhibitors. The presence of an integral membrane protein such as synaptotagmin in the soluble fraction likely reflects a light buoyant density lipid-containing fraction that rises to the surface during centrifugation.
Synaptophysin, a marker for synaptic vesicles, some of which may be associated with plasma membrane sheets in nerve terminals, was observed as the expected 38-kD band in the 10K and 30K pellets, with less signal in the 20K fraction. The distribution of RESP18 most closely resembled that of BiP, suggesting that RESP18 is located in the endoplasmic reticulum of neurons in these nuclei.
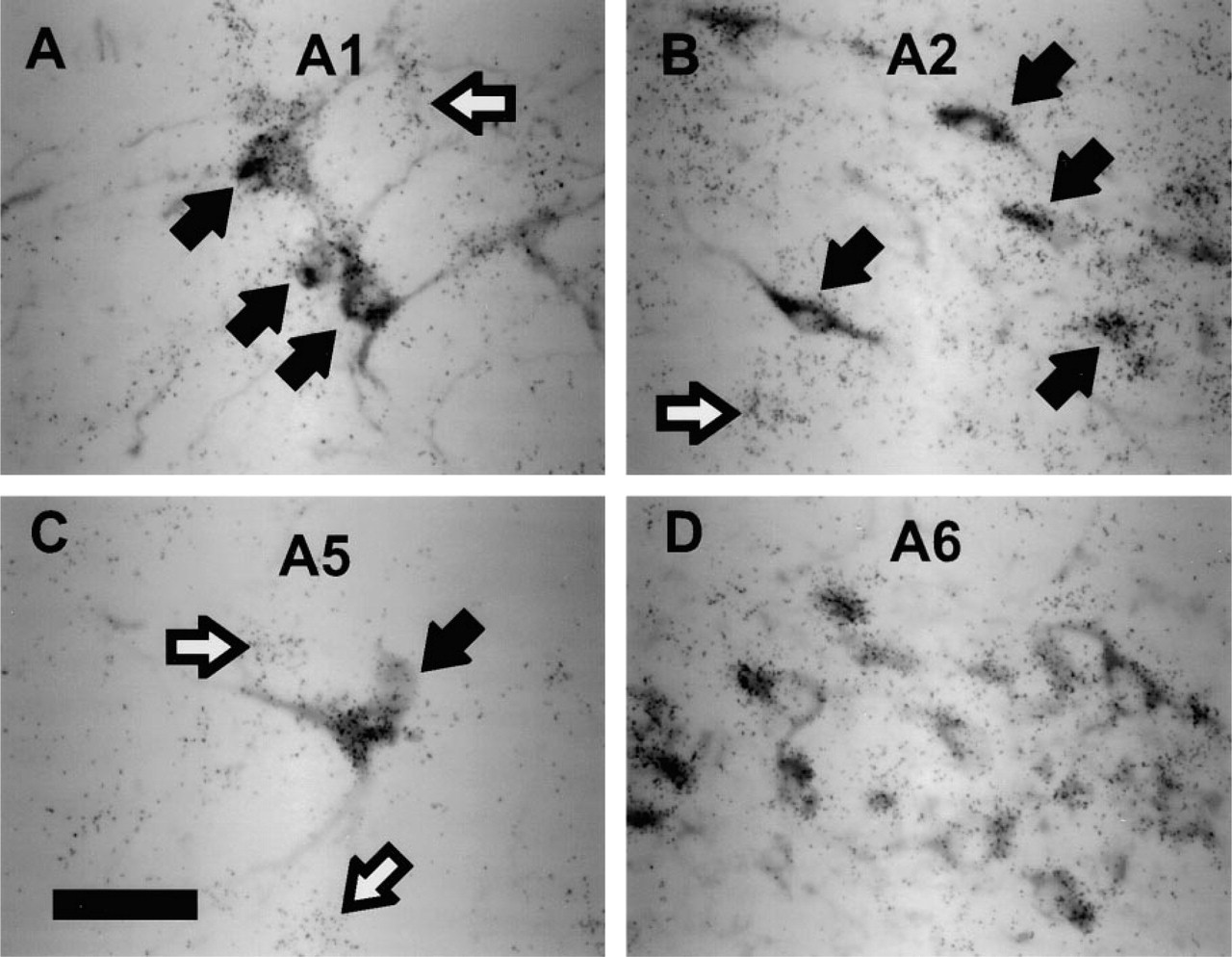
RESP18 expression in catecholaminergic neurons. RESP18 in situ hybridization in neurons that were immunostained with DβM antiserum in the (
Because the magnocellular neurons in the supraoptic and paraventricular nuclei are readily visualized with antisera to RESP18 and have terminals situated in the neural lobe of the pituitary, homogenates of neural lobe were subjected to differential centrifugation (Figure 8C). High molecular weight isoforms of RESP18 (22 kD and 24 kD) were much more prevalent in neural lobe homogenates than in homogenates of SON, PVN, or arcuate nucleus (Figures 8B and 8C). The subcellular fractionation analysis is consistent with the localization of 18-kD RESP18 to the endoplasmic reticulum of hypothalamic neurons. The presence of 22-and 24-kD RESP18 isoforms in the neural lobe of the pituitary indicates that the higher molecular weight isoforms are found in distal compartments of the secretory pathway.
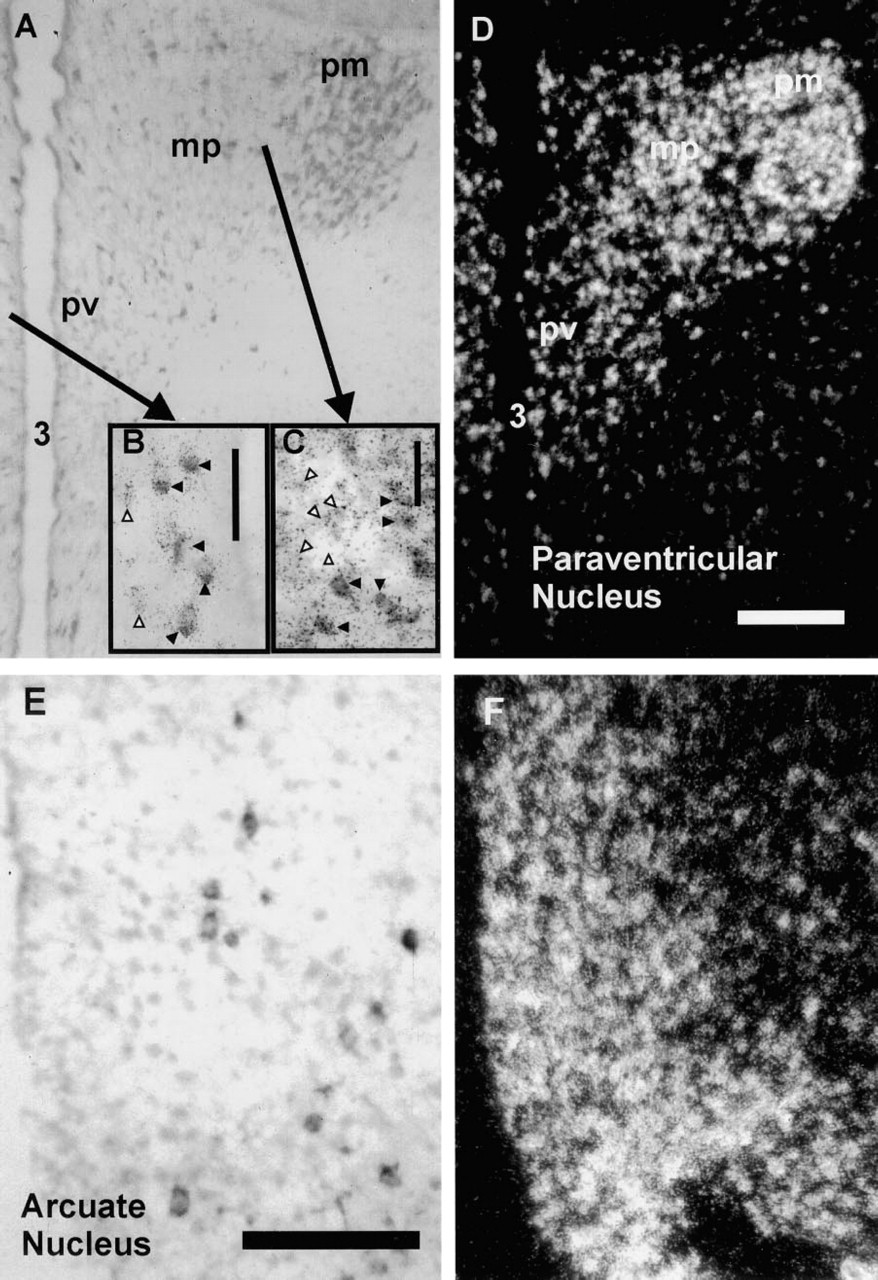
Comparison of sites of RESP18 protein and mRNA expression. RESP18 immunostaining (
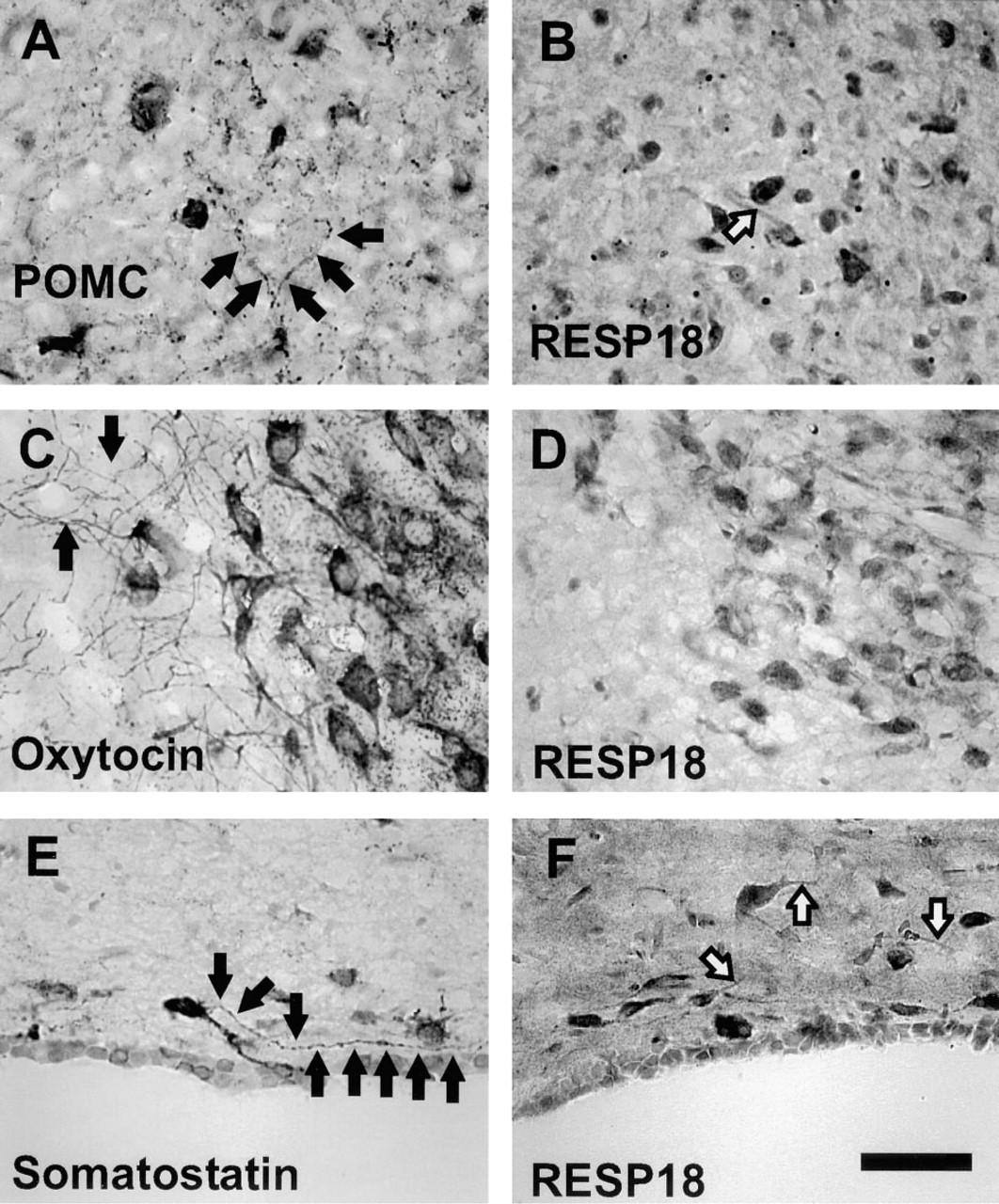
Comparison of RESP18, POMC, oxytocin, and somatostatin immunostaining. RESP18 immunostaining was compared to POMC immunostaining in the arcuate nucleus (
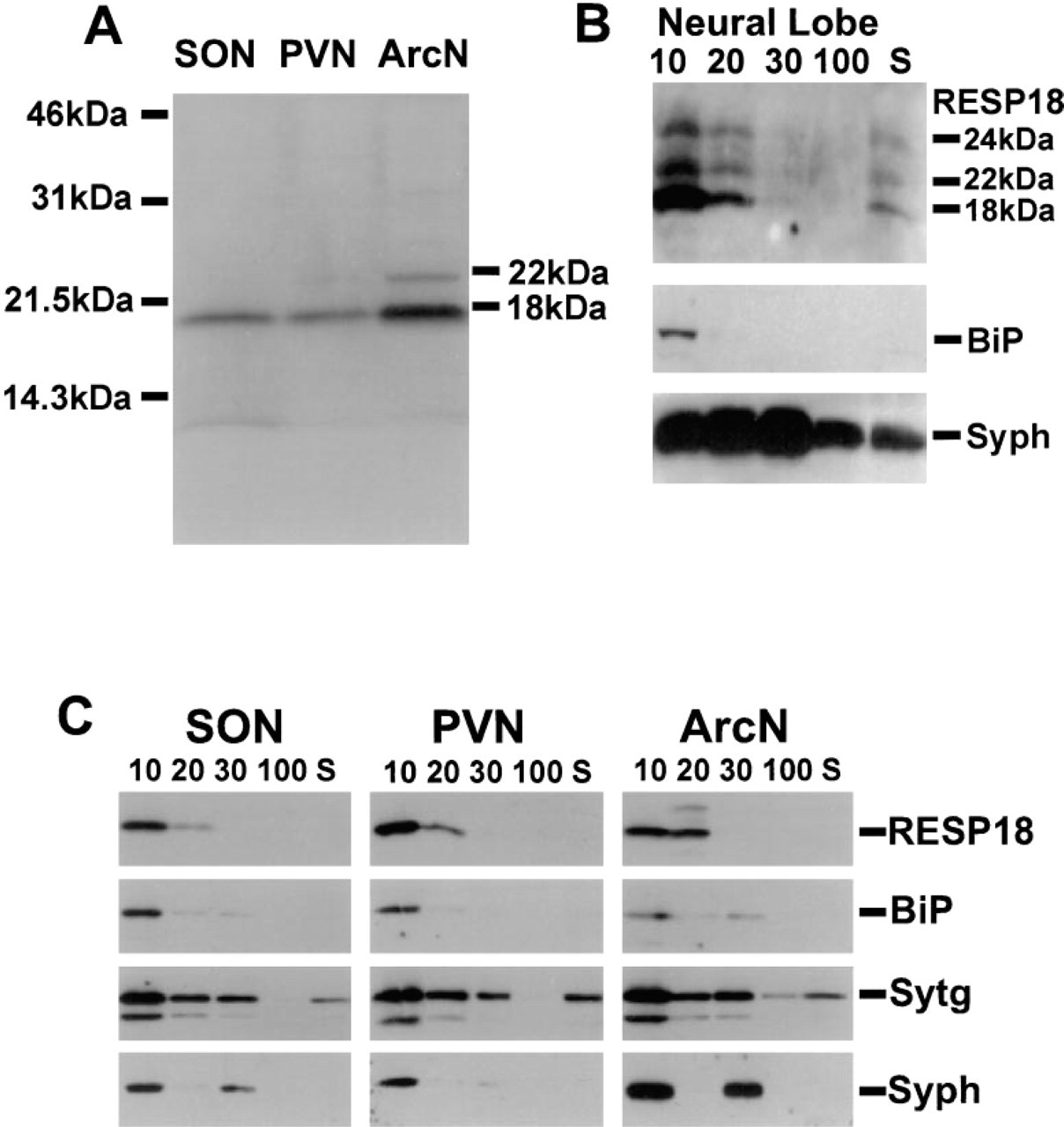
Western blot analysis of RESP18. (
Discussion
In this study we have shown that RESP18 mRNA is expressed in neurons that secrete peptide hormones and catecholamines. In particular, RESP18 is expressed in CRH, OXY, and AVP neurons of the medial parvocellular PVN and in CRH, TRH, and somatostatin neurons of the periventricular PVN. These neurons project to the hypophyseal portal circulation in the median eminence, where these neurohumoral agents regulate pituitary secretion (Swanson and Sawchenko 1983; Zimmerman and Nilaver 1984). Some parvocellular CRH, AVP, and OXY neurons expressing RESP18 project into and regulate autonomic areas of the brain-stem (Armstrong 1995). RESP18 expression was observed in magnocellular AVP and OXY neurons of the PVN, SON, and accessory nuclei. These neurons project to the posterior pituitary and are involved in salt and water homeostasis and in milk letdown (Swanson and Sawchenko 1983).
RESP18 mRNA was highly expressed in the arcuate nucleus. Because the majority of cerebral POMC neurons are located in the arcuate nucleus (Bloom et al. 1978; Watson et al. 1978; Gee et al. 1983), we performed in situ hybridization for RESP18 combined with POMC immunostaining and detected high levels of RESP18 mRNA expression in POMC neurons. RESP18 mRNA was also found in arcuate neurons that do not immunostain for POMC and which correspond to dopaminergic neurons by location (Simerly 1995). These neurons project to the median eminence and secrete POMC-derived peptides and dopamine into the hypophyseal portal circulation. Dopamine regulates prolactin secretion in the anterior pituitary and POMC expression in the intermediate pituitary (Gibbs and Neil 1978; Mains et al. 1990).
RESP18 is also expressed in the A1, A2, A5, and A6 catecholamine groups (Lindvall et al. 1974; Loewy et al. 1979; Aston–Jones et al. 1995), which are implicated in autonomic and pituitary function and in sleep arousal (Aston–Jones and Bloom 1981; Sawchenko et al. 1985; Byrum and Guyenet 1987). Because catecholamine neurons have been shown to produce many different neuropeptides and neurohormones, such as neuropeptide Y (Figure 5) (Everitt et al. 1984; Sawchenko et al. 1985; Blessing et al. 1986; Aston–Jones et al. 1995), RESP18 may function in association with peptides co-expressed with catecholamines.
By comparing the visualization of RESP18 protein and mRNA, it became clear that expression of RESP18 is controlled at both the transcriptional and the post-transcriptional level. Neurons containing similar amounts of RESP18 mRNA contain vastly different amounts of RESP18 protein (Figure 6). We observed abundant RESP18 mRNA expression throughout the PVN and arcuate. However, only magnocellular neurons and some periventricular neurons in the PVN showed detectable RESP18 immunostaining. RESP18 immunostaining was low or at background levels in most of the parvocellular division, the area containing most of the CRH neurons. In the arcuate nucleus, RESP18 immunostaining was robust in the large neurons that correspond to POMC-containing cells. However, little or no immunostaining for RESP18 was detected in the area of the arcuate that contains dopaminergic neurons, even though the signal for RESP18 mRNA was strong in this region (Figure 6). We have observed the same phenomenon in the pituitary, adrenal medulla, and pancreatic islets, in which the signal for RESP18 mRNA is intense and uniform throughout the glands but immunostaining is found in only a subset of the cells (Darlington et al. 1996b). This finding suggests that translation of the RESP18 mRNA or the half-life of the RESP18 protein is regulated in a cell-specific manner. In AtT-20 cells, the half-life of newly synthesized endogenous RESP18 is only 18 min (Schiller et al. 1995). Regulation of translation or protein turnover would allow RESP18 neurons to respond rapidly to physiological stimuli.
The RESP18 protein appears to be restricted to the perikarya, and RESP18 does not appear in long fibers of passage (axons) or terminal boutons, as shown by immunocytochemistry. By comparing the immunostaining pattern of POMC, oxytocin, and somatostatin to that of RESP18 in the same nuclear regions, we found that RESP18 antisera immunostained perikarya in a manner similar to the immunostaining of POMC in the arcuate, oxytocin in the supraoptic nucleus, and somatostatin in the paraventricular nucleus. However, RESP18 antisera did not immunostain terminal boutons as did POMC and somatostatin antisera or long fibers (axons) as did oxytocin antisera, suggesting that RESP18 protein is not in axons or terminals. Analysis of RESP18 protein expression in homogenates of SON, PVN and arcuate identified 18-kD RESP18 in the same subcellular fractions as the endoplasmic reticulum marker BiP. Higher molecular mass isoforms (22 and 24 kD) were identified in the neural lobe of the pituitary (Figure 8) in the same subcellular fractions as markers for synaptic vesicles, suggesting that some RESP18 protein can appear in terminals (Figure 8) but not enough to immunostain (Darlington et al. 1996b). Higher molecular mass isoforms of RESP18 have been detected in rat anterior pituitary, intermediate pituitary, testis, thyroid, and plasma (Schiller and Darlington 1996; Darlington et al. 1996b). In the anterior pituitary, the increase in molecular mass appears to be due to O-glycosylation because treatment of extracts with neuraminidase reduced the levels of 24-kD RESP18 (Schiller and Darlington 1996). Transfected cell lines that overexpress RESP18 secrete a higher molecular mass RESP18 isoform, suggesting that RESP18 is modified, possibly by O-glycosylation, as it moves through the Golgi (Schiller and Darlington 1996). In this study, the appearance of 22- and 24-kD RESP18 in neural lobe homogenates suggests that a small amount of RESP18 protein exits the endoplasmic reticulum and enters distal compartments of the secretory pathway. This finding is consistent with the prominent perikaryal staining observed for RESP18 and is also supported by subcellular fractionation of hypothalamic nuclei. RESP18 protein was found in the 10K pellet coinciding with BiP, an endoplasmic reticulum marker. The 22-kD isoform of RESP18 was found in the 20K pellet, suggesting that it traversed more of the secretory pathway (Bloomquist et al. 1994b).
RESP18 protein is largely restricted to the endoplasmic reticulum of AtT-20 cells (Schiller et al. 1995). A similar localization of neuronal RESP18 was suggested by its prominent immunostaining in perikarya (Figure 7). Western blot analysis was used to search for the higher molecular mass isoforms of RESP18 that are formed only when RESP18 enters the Golgi complex (Schiller et al. 1995). Only small amounts of the 22-kD isoform of RESP18 were detected in arcuate nucleus extracts. We used differential centrifugation to compare the distribution of RESP18 to that of known markers for endoplasmic reticulum (BiP), large dense core vesicles (synaptotagmin), and synaptic vesicles (synaptophysin). Subcellular fractions localized RESP18 in the endoplasmic reticulum in punches of the PVN and SON. These punches contain the cell bodies of AVP and OXY neurons that project to the neural lobe of the pituitary for secretion of AVP and OXY. Subcellular fractions of the neural lobe showed 22- and 24-kD isoforms of RESP18, indicating that RESP18 is found in more distal compartments of the secretory pathway.
In conclusion, we have found that RESP18 is located in neurons that synthesize neuropeptides (AVP, OXY, CRH, TRH, somatostatin, POMC, neuropeptide Y) and catecholamines in the hypothalamus and brainstem. Furthermore, the post-transcriptional regulation of RESP18 expression varies from neuron to neuron, because RESP18 mRNA levels are uniformly high throughout the PVN and arcuate but immunodetectable RESP18 protein is not. Finally, immunohistochemical studies and differential centrifugation both indicate that the RESP18 protein is largely retained in the endoplasmic reticulum, with only a small amount of RESP18 entering the distal parts of the secretory pathway.
Footnotes
Acknowledgements
Supported by grants from NIH GM46540 and NIDA DA-00266, DA-05540.
We thank Jay Baraban for the use of the cryostat, Richard Johnson for making plasmids, and Marie Bell for laboratory assistance and her endless pursuit of photographs. We appreciate the donation of antisera for synaptotagmin from Dr Richard Scheller, for vasopressin (VA4) and oxytocin (VA10) from Dr H. Gainer, for TRH from Dr Eduardo Nillni, and for somatostatin from Dr Brian Noe.