
Abstract
Tumor necrosis factor (TNF) induces apoptotic death of hepatocytes in the galactosamine (GalN)-sensitized mouse liver after 5 hr. In our study, the most remarkable sign of the early stage of apoptosis was the focal rupture of the outer mitochondrial membrane. Parts of the inner membrane extended through the gap of the outer membrane, whereas the rest of the inner membrane still formed the cristae. This feature appeared in hepatocytes before chromatin condensation. With the diaminobenzidine technique for localization of cytochrome oxidase activity, the reaction product was detectable by light and electron microscopy. Ten percent of the hepatocytes were apoptotic, with condensed chromatin and high enzyme activity, 37% were pre-apoptotic, without chromatin condensation but high enzyme activity, and 53% had neither condensed chromatin nor a remarkable reaction product of cytochrome oxidase activity. Fas (APO-1, CD95) molecules on the plasma membrane of hepatocytes increased and were represented immunohistochemically in cells without chromatin condensation. DNA strand breaks were also detectable before chromatin aggregation. The results of this study indicate that mitochondria play a pivotal role in pre-apoptotic hepatocytes, together with an increase of the Fas molecule on the plasma membrane and with the occurrence of DNA strand breaks in the nucleus.
T
Recently, the participation of mitochondria in the apoptotic cascade was described: The pre-apoptotic disruption of the mitochondrial transmembrane potential occurred before cells exhibited nuclear DNA fragmentation (Kroemer et al. 1997). Human mutant cell lines that lacked mitochondrial DNA could still be induced to undergo apoptosis (Jacobson et al. 1993). Using cytochrome c as electron donor/acceptor, O'Donnell et al. (1995) were able to measure mitochondrial respiratory enzymes in TNF α-mediated apoptosis in cell cultures. Krippner et al. (1996) reported that the activity of cytochrome oxidase appeared to be normal but that the content of cytochrome c was greatly diminished in the apoptotic program of Jurkat cells.
The aim of this study was to investigate morphological changes and functional characteristics of mitochondria in pre-apoptotic and apoptotic mouse hepatocytes. Applying immunohistochemical methods, we examined whether Fas expression and DNA fragmentation into strand breaks occurred before chromatin condensation was seen.
Materials and Methods
Recombinant murine TNFα was generously provided by Dr. G.R. Adolf (Boehringer Institute; Vienna, Austria).
Animals
This study was performed in compliance with the German Animal Protection Law under a permit issued by the state government (Freiburg, Germany). Six- to 8-week-old male BALB/c mice were obtained from the animal facility of the University of Konstanz (Germany) and kept on a standard laboratory diet with free access to water. Sixteen hours before the beginning of the experiments food was withdrawn. GalN (700 mg/kg) (Roth Chemicals; Karlsruhe, Germany) was given IP in a volume of 200 μl saline. Recombinant murine TNF (10μg/kg) was injected into the tail vein in a volume of 300μl saline containing 0.1% HSA 20 min after GalN administration. The administration of only TNF or only GalN served as a negative control for the induction of apoptosis. Five hours after treatment, animals were anesthetized by sodium pentobarbital 150 mg/kg
Fixation
For light and electron microscopic studies, the livers were fixed via the portal vein with a fixative containing 0.25% glutaraldehyde and 2% sucrose in 100 mM piperazine-N,N'-bis (2-ethanesulphonic acid) (Pipes) buffer at a pH value of 7.4 for 5 min. After fixation, 50- μm sections were cut with a microslicer (Dosaka EM; Kyoto, Japan) and collected in 100 mM Pipes buffer at a pH value of 7.4.
Electron Microscopy
Incubation for Cytochrome Oxidase Activity. For demonstration of cytochrome oxidase activity we applied a modified 3,3-diaminobenzidine-tetrahydrochloride (DAB) technique (Angermüller and Fahimi 1981). Sections were incubated for 60 min at 37C in a medium containing 15 mg DAB and 0.05% cytochrome c in 100 mM Pipes buffer, pH 7.2. Control incubation was performed in the same medium also containing 50 mM NaCN as an inhibitor of cytochrome oxidase activity. Sections were postfixed with aqueous osmium tetroxide, dehydrated in graded ethanol, and embedded in Epon 812. Ultrathin sections were counterstained with lead citrate for 1 min and examined in a Philips EM 301 electron microscope.
Light Microscopy
Immunohistochemical Localization of Fas Receptor. Eponembedded semithin sections were etched with 10% NaOH in absolute ethanol for 30-45 min (Litwin et al. 1984). Deplasticized sections were rehydrated, incubated in 3% H2O2 for 5 min to inhibit endogenous peroxidase activity, washed in PBS, transferred into protein blocking buffer (5% goat serum and 1% BSA in PBS buffer, pH 7.4) for 30 min, washed again, and incubated in a humidified chamber with the monoclonal hamster anti-mouse Fas antibody (Jo2; Pharmingen; Hamburg, Germany) for 12 hr at a dilution of 1:1000. After a short rinse in PBS buffer, semithin sections were incubated with the second antibody, a peroxidase-labeled goat antihamster IgG antibody (Pharmingen), diluted 1:50, for 90 min. For the visualization of peroxidase activity we used 3-amino-9-ethyl-carbazole as chromogen. Sections were counterstained with hematoxylin for 1 hr. Applying the same procedure, semithin sections of the liver of untreated animals served as negative control. An incubation only with the second antibody, goat anti-hamster IgG, served as a further control.
Immunohistochemical Detection of DNA Strand Breaks
For in situ labeling of DNA breaks in nuclei, we used LR White-embedded semithin sections and applied the method of Gavrieli et al. (1992) using the in situ cell death detection kit, POD (Boehringer; Mannheim, Germany). The incubation of the semithin sections started with 20μg/ml proteinase K in Tris-HCl for 15 min. Sections were rinsed with PBS and incubated with the TUNEL [terminal dexoynucleotidyl transferase (TdT)-mediated dUTP-biotin nick end-labeling] reaction mixture in a humidified chamber for 60 min at 37C. After washing again, samples were incubated under the same conditions with the converter POD (anti-fluorescein antibody conjugated with peroxidase as reporter enzyme) for 30 min. Peroxidase was visualized by DAB as chromogen substrate tablets (DAB tablets; Kem-En-Tec, Copenhagen, Denmark). For a negative control, a semithin section of the liver of a GalN/TNF-treated animal was incubated only with dUTP, followed by incubation with the converter POD.
Quantification of Pre-apoptotic and Apoptotic Cells
Animals treated only with GalN or TNF did not show preapoptotic or apoptotic changes. Therefore, we counted the nuclei or cells only of GalN/TNF-treated mice. In LR Whiteembedded semithin sections apoptotic nuclei stained by hematoxylin/eosin were determined. A total of 1400 nuclei were randomly counted and the percentage of the number of nuclei with condensed chromatin was calculated. In Eponembedded semithin sections incubated for demonstration of cytochrome oxidase activity, pre-apoptotic and apoptotic Mitochondrial Alterations in Apoptotic Hepatocytes hepatocytes were identified by the strong brown reaction product. A total of 1200 cells were counted randomly and the percentage of the number of pre-apoptotic and apoptotic hepatocytes was calculated.
Results
We investigated death factor-induced hepatocellular apoptosis in the livers of GalN/TNF-treated mice cytochemically and immunohistochemically 5 hr after treatment. The liver was sparsely infiltrated by lymphocytes. Therefore, additional leukocyte mediators were unlikely to contribute to hepatocellular apoptosis. Apoptotic cell death is very rare in the healthy adult mouse liver. Approximately 1-5 apoptotic cells/10,000 hepatocytes are found in untreated mice (for review see Schulte-Hermann et al. 1995). In our model, hematoxylin/eosin-stained LR White semithin sections revealed 10% apoptotic hepatocytes, identified by chromatin condensation. Neither GalN nor TNF administration induced hepatocellular apoptosis.
The DAB method for demonstration of cytochrome oxidase activity revealed that this last enzyme of the respiratory chain was very active in mitochondria of pre-apoptotic and apoptotic hepatocytes when we offered cytochrome c as electron donor in the incubation medium (Figures 1A and 1B). Before chromatin condensation was seen, alterations occurred in mitochondria. The outer membrane of the mitochondria was scarified, and the inner membrane had expanded through this gap to form a large bulge (Figures 2B-2D). In that part of the mitochondrion in which the outer membrane was intact, cristae stained by the DAB reaction product were tightly folded by the inner membrane, and the bulge resembled a vacuole without or with fewer cristae (Figure 1A). When chromatin condensation had taken place, the same alterations were found in mitochondria (Figures 1B and 2A). Cytochrome oxidase was very active in all mitochondria of pre-apoptotic and apoptotic hepatocytes (Figures 1A, 1B, 2A, 3A, and 4A). It should be emphasized that 47% of the hepatocytes showed these characteristic alterations. In GalN/TNF-treated animals, 53% of the hepatocytes showed normal mitochondrial morphology with less cytochrome oxidase activity (Figures 3A, 3B, and 4A). Neither GalN nor TNF administration caused any bulges of the inner mitochondrial membrane, but the administration induced a remarkable increase in the number of cytochrome oxidasepositive mitochondria in comparison to the above mentioned 53% with normal mitochondrial structure (Figures 3A-3D and 4A-4C). In control sections incubated with NaCN together with cytochrome c, the DAB reaction product was not detected in mitochondria.
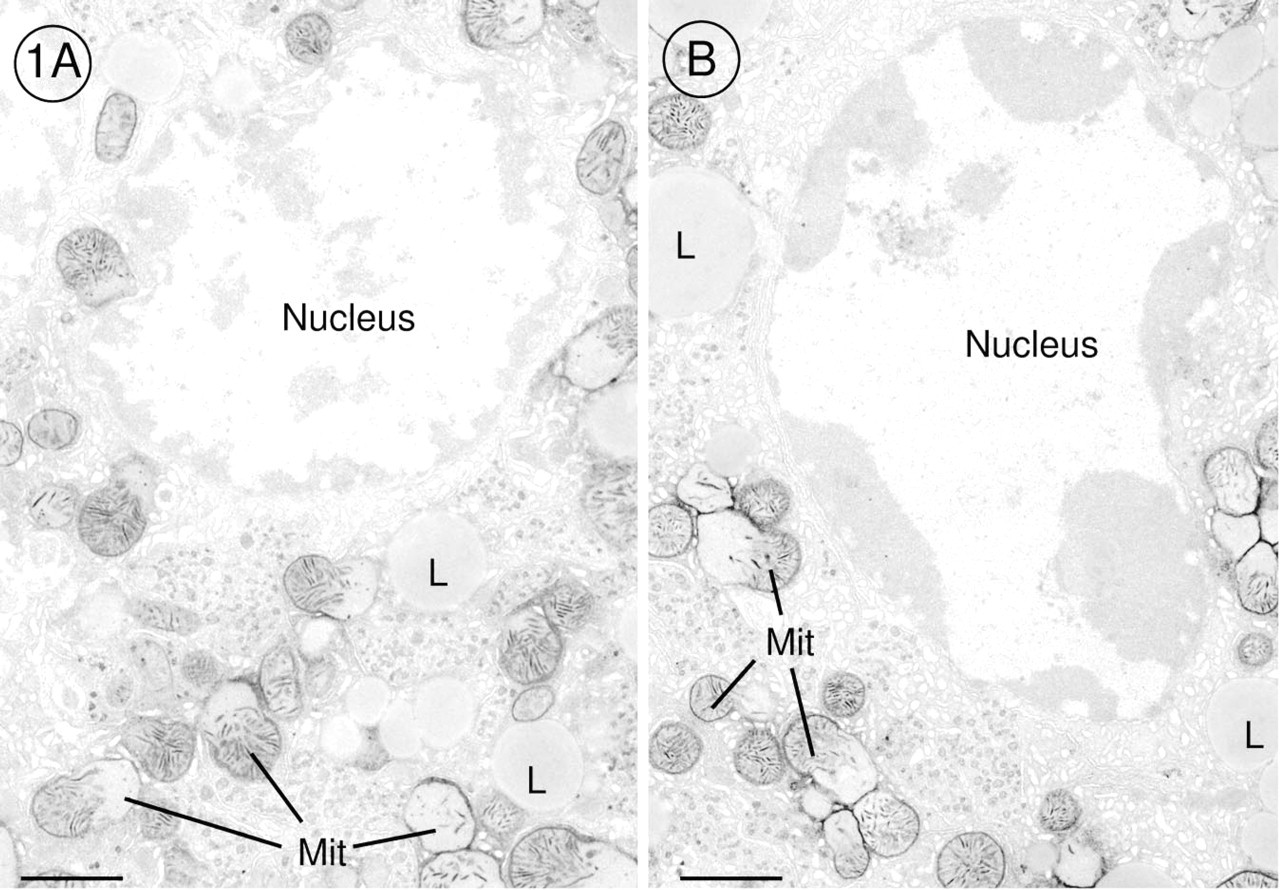
Electron micrographs of hepatocytes of GalN/TNF-treated mouse liver incubated for demonstration of cytochrome oxidase activity by the DAB method. (
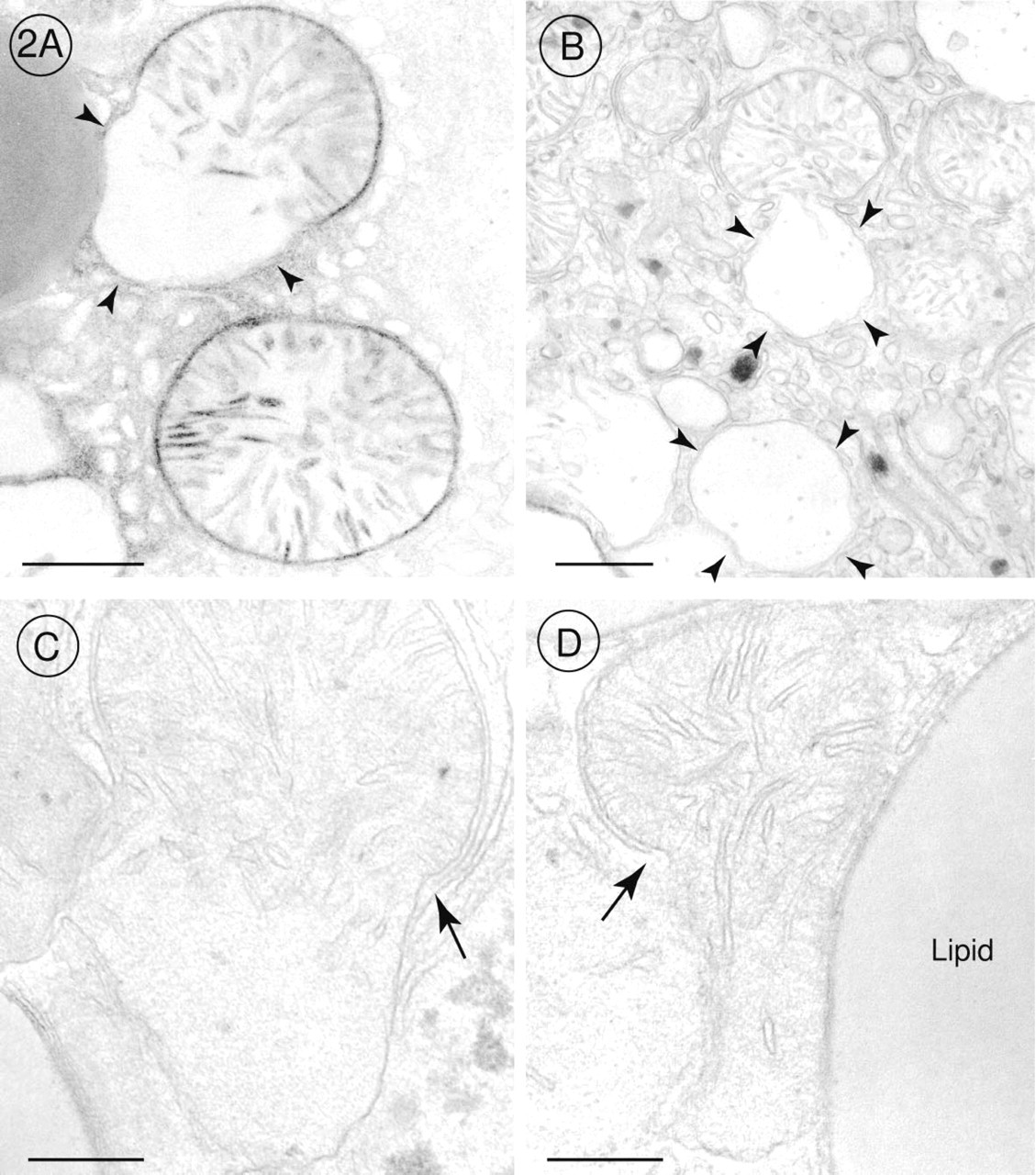
Higher magnifications of electron micrographs of hepatocytes of GalN/TNF-treated mice. (
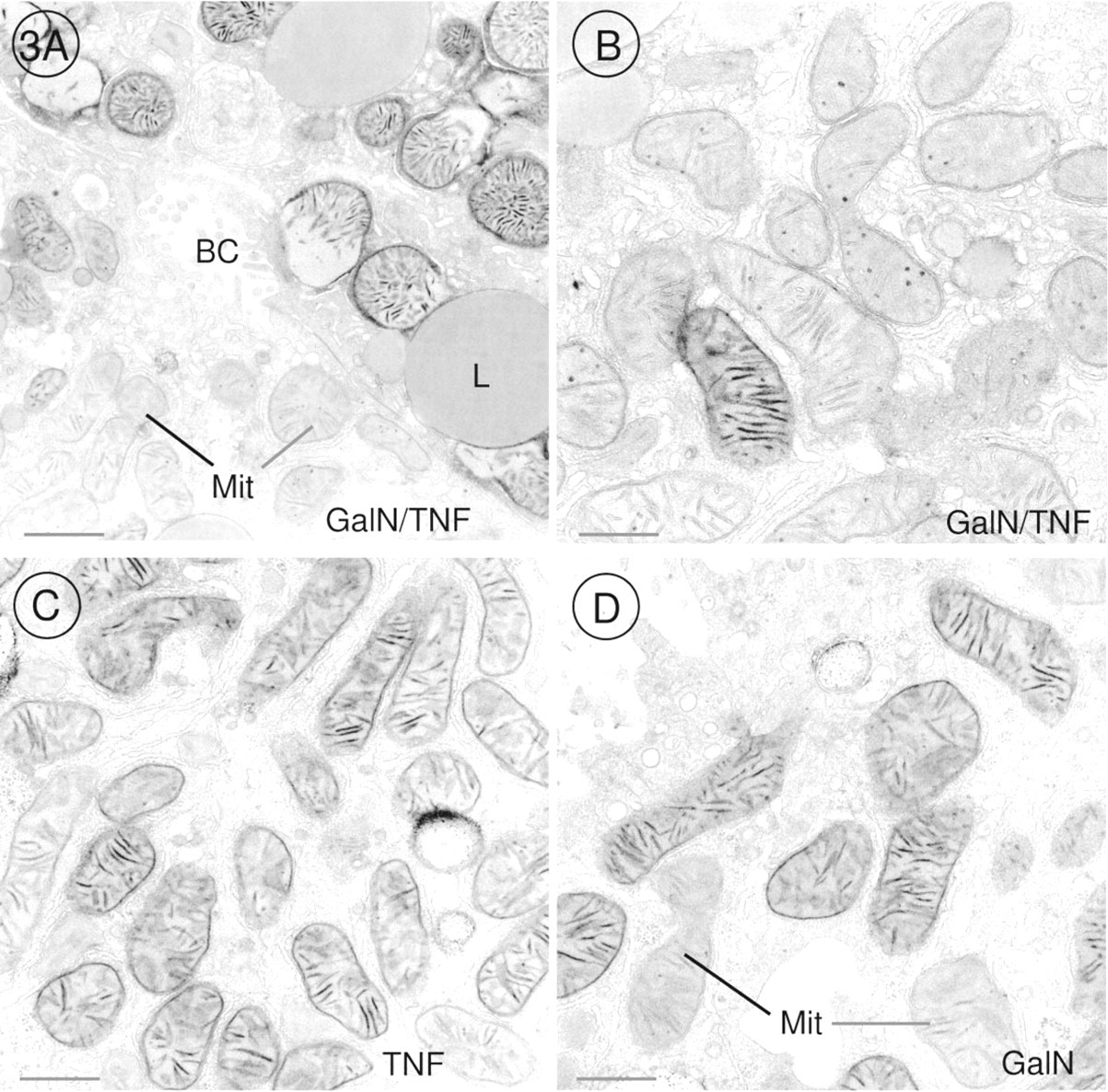
Electron micrographs of hepatocytes incubated for demonstration of cytochrome oxidase activity. (
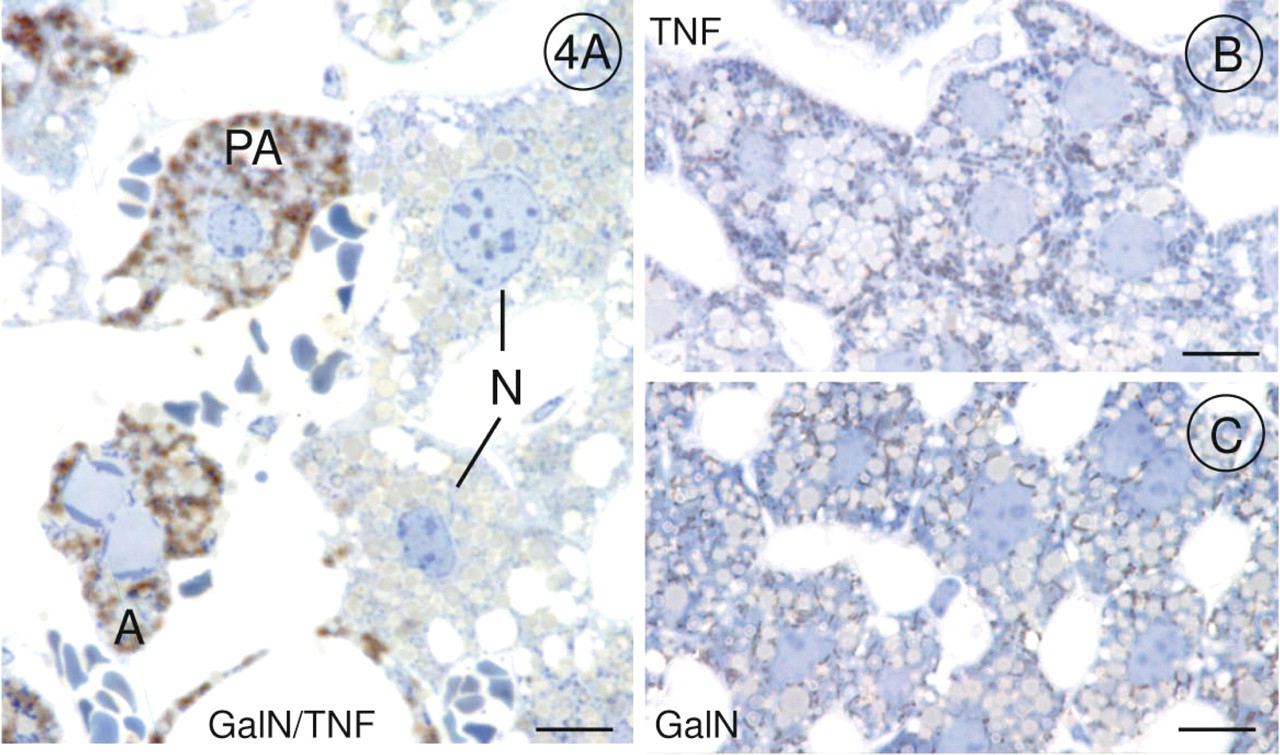
Light micrographs of semithin Epon sections incubated for demonstration of cytochrome oxidase activity and stained with toluidine blue; (
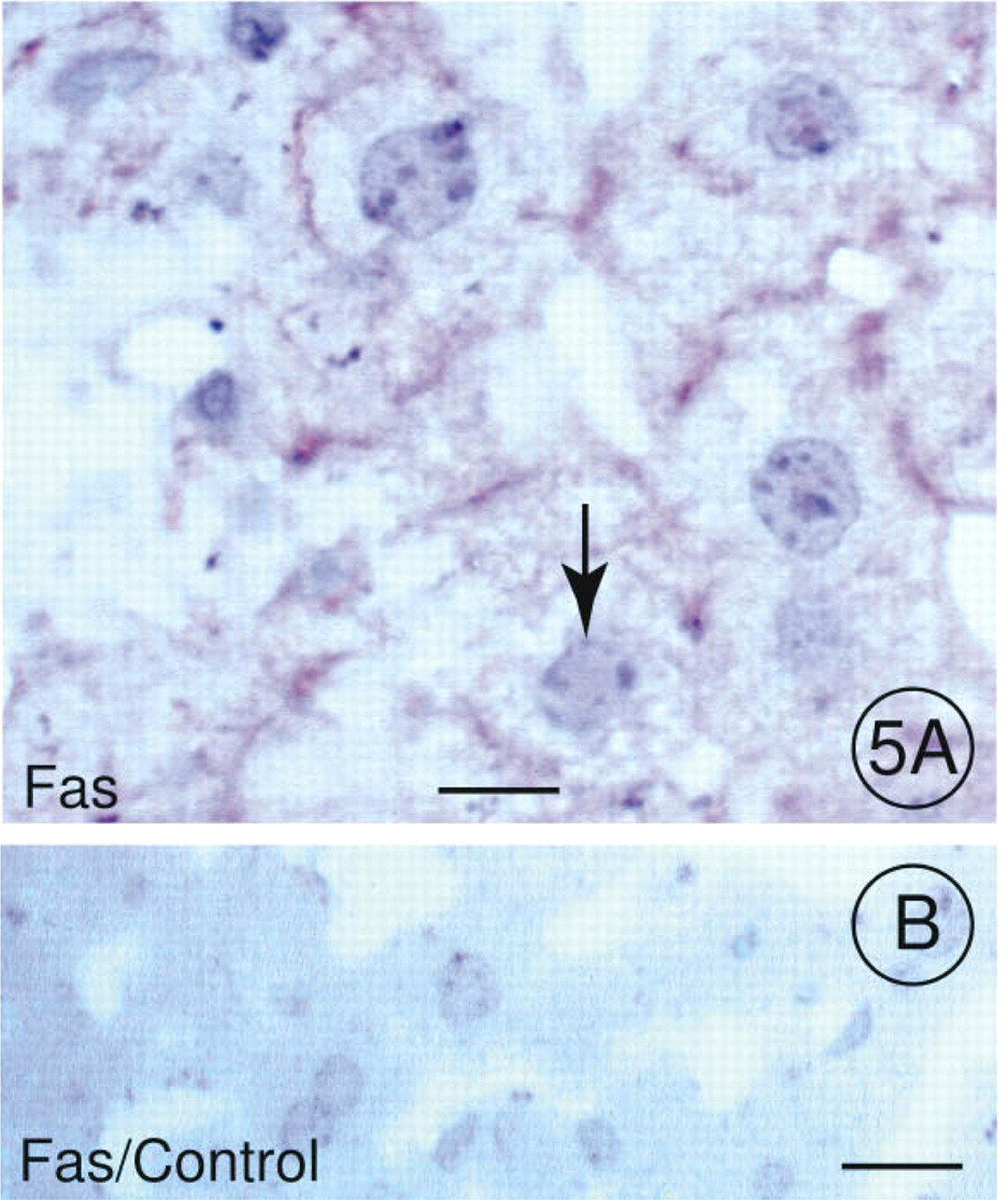
Immunohistochemical Localization of Fas
The Fas receptor is present in cell membranes of several tissues, including normal liver (Schulte-Hermann et al. 1995). However, GalN/TNF treatment stimulated the expression of Fas on the plasma membrane of hepatocytes. As shown in Figure 5A, the Fas antibody specifically recognized the Fas receptor on the hepatocellular membrane before chromatin condensation, whereas sections of an untreated control animal did not show any reaction product when incubated with the Fas antibody (Figure 5B).
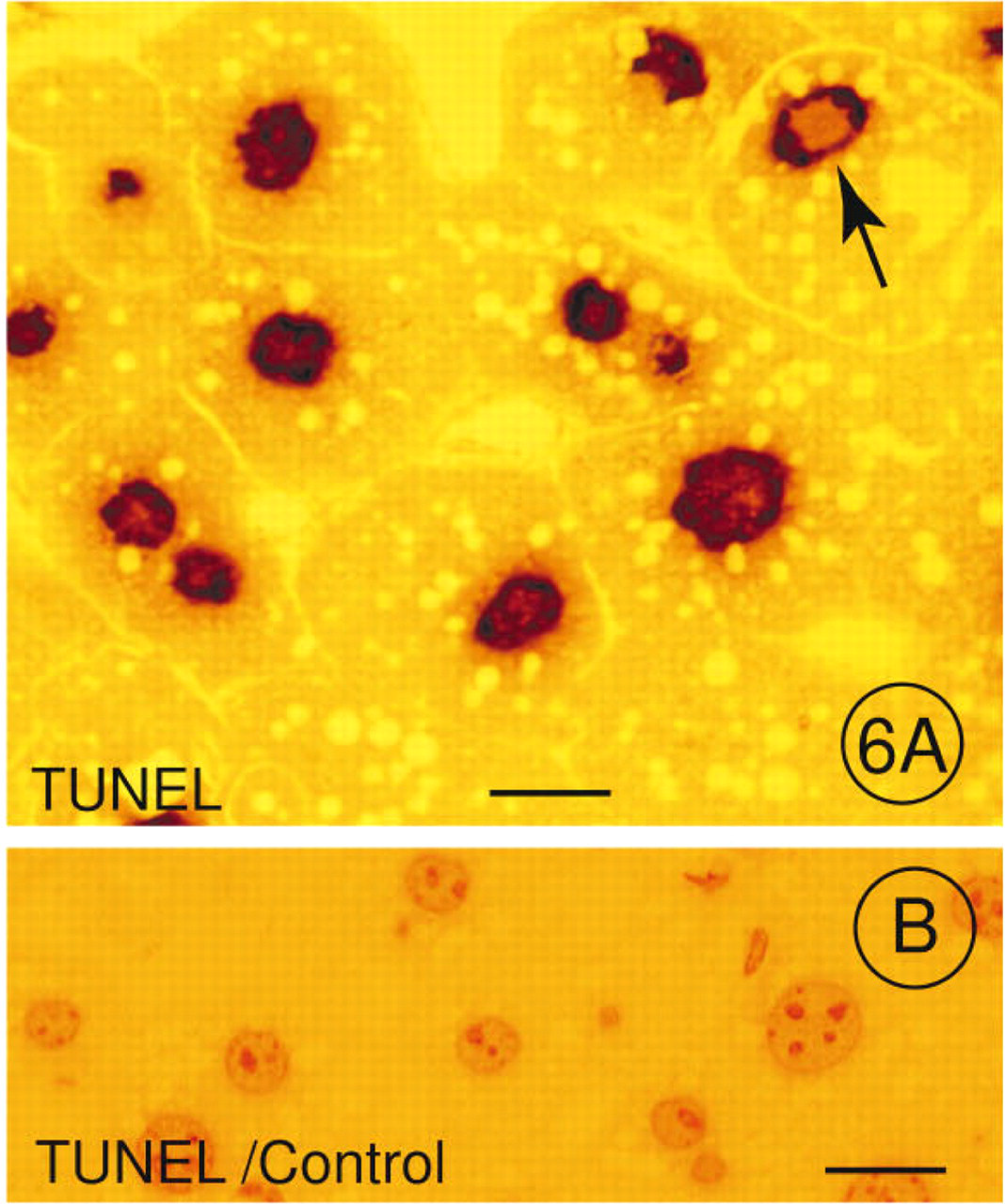
Immunohistochemical Detection of DNA Strand Breaks
The TUNEL method end-labeled DNA double-strand breaks and single-strand breaks in hepatocellular nuclei of LR White-embedded semithin sections. Figure 6A shows the polymerization of dUTP to the 3′-hydroxyl end of fragmented DNA, resulting in a brown reaction product that was detected in most nuclei of hepatocytes (Figure 6A). The negative control of a semithin section from a treated animal incubated only with dUTP failed to show any reaction product (Figure 6B).
Discussion
In this study we investigated functional characteristics and earliest morphological alterations of TNF-induced hepatocellular apoptosis in the GalN-sensitized mouse liver.
Before the known morphological signs of apoptosis, such as chromatin condensation, cell shrinkage, and fragmentation were detectable (Kerr et al. 1972; Faa et al. 1992; Wyllie and Duvall 1992; Majno and Joris 1995; Patel and Gores 1995; Feldmann 1997), we observed ultrastructurally a focal rupture of the outer mitochondrial membrane as the first indication of the morphological apoptotic program. In the past, electron microscopic observations had mainly focused on nuclear alterations, whereas changes of the mitochondrial ultrastructure were less often and controversially discussed. In apoptotic cells, mitochondria are described as normal (Searle et al. 1987; Cossarizza et al. 1995; Majno and Joris 1995; Kimura et al. 1997) or slightly swollen (Krippner et al. 1996). Petit et al. (1995) observed a reduction in the number of cristae and a destruction of the surrounding membrane in mitochondria of apoptotic thymocytes. We were able to identify two different features in mitochondria of preapoptotic and apoptotic hepatocytes: (a) the outer membrane was scarified, and through this gap the inner membrane expanded, forming a large bulge predominately without cristae; and (b) where the outer mitochondrial membrane was still intact, the inner membrane was folded into tightly packed cristae. If mitochondria were cut through the portion with the tightly packed cristae, they appeared in a normal mitochondrial morphology; if they were cut through the bulge, they resembled a great vacuole hardly recognizable as a mitochondrion.
In contrast to the morphological description of mitochondria in apoptotic cells, their functional role has been emphasized by many authors. The reduction in the mitochondrial transmembrane potential (Δ Φ m) that causes opening of mitochondrial permeability transition pores is an early event in the apoptotic process (Castedo et al. 1995; Cossarizza et al. 1995; Petit et al. 1995; Zamzami et al. 1995,1996). Those “apoptotic” mitochondria contain a protein of about 50 kD in the mitochondrial intermembrane space, which is released when the outer membrane is disrupted (Susin et al. 1996). Enari et al. (1995) described similar results using a cell lysate containing a substance that induces apoptosis. The authors combined this substance with the purified nuclei of intact cells and observed chromatin condensation and DNA fragmentation of the nuclei. The release of this protein was significantly inhibited by Bcl-2 protein (Kroemer et al. 1997). Bcl-2 is a proto-oncogene that blocks programmed cell death (Hockenbery et al. 1990,1991). It is located in the perinuclear membrane and throughout the cytoplasm, and is predominantly associated with the outer mitochondrial membrane (Monaghan et al. 1992). Recently, a functional association between Bcl-2 protein and cytochrome c was postulated by several authors (Kluck et al. 1997; Yang et al. 1997). Overexpression of Bcl-2 protein prevents the outflow of cytochrome c from mitochondria, thereby blocking caspase-3 activation (Kluck et al. 1997; Yang et al. 1997). The apoptotic protease cascade leading to caspase-3 activation is initiated by the formation of the Apaf-1 (apoptotic protease activating factor-1)/caspase-9 complex in the presence of cytochrome c and dATP. This event leads to caspase-9 activation, which in turn cleaves and activates caspase-3 (Li et al. 1997; Zou et al. 1997). Activated caspases play a central role in the biochemical apoptotic pathway (Nicholson and Thornberry 1997).
Light micrographs of deplasticized semithin Epon sections. (
LR White-embedded semithin section of the liver of a GalN/TNF-treated mouse. (
It appears that mitochondrial apoptotic factors and all structures involved in the pre-apoptotic ruptures of the Δ Ψm are present in mitochondria lacking mitochondrial DNA. Therefore, apoptosis is controlled by the nuclear rather than by the mitochondrial genome (Jacobson et al. 1993; Marchetti et al. 1996). The enzymes of the respiratory chain appear not to be involved in the apoptotic process (O'Donnell et al. 1995; Krippner et al. 1996). The data presented in our study indicate that mitochondrial cytochrome oxidase in apoptotic cells can still function when cytochrome c is offered as electron donor. Krippner et al. (1996) demonstrated that the activity of cytochrome oxidase appears to be normal in Fas-mediated apoptotic cells but that inhibition and inactivation of cytochrome c comprise the potentially fatal components of the apoptotic program. This suggests that the function of the respiratory chain can be disrupted by a lack of cytochrome c. Even in cytochrome c-containing mitochondria of apoptotic cells, cytochrome oxidase is unable to function when cytochrome c is inactivated by a putative cytochrome c inhibitor (Krippner et al. 1996). Asoh et al. (1996) reported that defects in mitochondria enhanced Fas gene expression. In our experiments, the Fas receptor increased and was identified immunohistochemically on the plasma membrane of hepatocytes before chromatin condensation was morphologically detectable. In untreated animals the Fas molecule was not determined, although Schulte-Hermann et al. (1995) described the appearance of the Fas receptor in normal liver. Hence, although the transcriptional inhibitor GalN normally blocks hepatic protein synthesis by 50% within 2 hr after administration to mice (Leist et al. 1995), Fas translation was still measurable. However, a functional role of Fas in TNF-induced hepatocellular apoptosis in mice has been excluded by experiments showing that lpr mice, which bear a functional mutation of Fas, were as sensitive to TNF as the corresponding wild-type (Leist et al. 1996). Therefore, Fas appears to be expressed at early time points during liver injury, independently of its functional contribution to the apoptotic process.
DNA strand breaks are a typical sign of apoptosis. Using the same GalN/TNF-model as we did, Leist et al. (1995) demonstrated hepatic oligonucleosomal DNA fragmentation by showing a typical DNA ladder on agarose gel. Using the TUNEL method, we were able to demonstrate DNA strand breaks before chromatin condensation. Therefore, DNA fragmentation occurs very early in the apoptotic process.
In summary, our study demonstrates characteristic morphological alterations in mitochondria as a very early hallmark of the hepatocellular apoptotic program. Before chromatin condensation occurs, the outer mitochondrial membrane is focally disrupted and the inner membrane protrudes through this gap. High levels of cytochrome oxidase activity are observed cytochemically in pre-apoptotic and apoptotic hepatocytes. Fas molecules on the plasma membrane of hepatocytes increase and are also present in cells without chromatin condensation. The appearance of DNA strand breaks is detectable before nuclear chromatin becomes aggregated in dense crescents beneath the nuclear envelope.
Footnotes
Acknowledgements
Supported by grants An 192/1-3 and Ti 169?4-1 from the Deutsche Forschungsgemeinschaft (Bonn Bad-Godesberg).
We thank Prof H.D. Fahimi for helpful discussion. The excellent technical assistance by Ms Barbara Lenschow and Ms Annette Stradtmann is gratefully acknowledged.