
Abstract
We designed a protocol to improve the immunohistochemical analysis of human brain structures, which overcomes the limited detection sensitivity, high background, and intense autofluorescence commonly associated with human tissue. This procedure was evaluated by using antibodies against major GABAA receptor subunits (α1, α2, α3, γ2) in autopsy and surgical specimens. Tissue blocks were briefly fixed by immersion and pretreated with microwave irradiation in sodium citrate buffer. Immunoperoxidase staining revealed a marked enhancement of cell surface immunoreactivity and reduction of background in microwave- irradiated tissue, irrespective of its origin. For confocal laser scanning microscopy, immunofluorescence staining was optimized with the tyramide signal amplification (TSA) technique. This procedure not only dramatically increased the sensitivity for antigen detection but also totally suppressed autofluorescence, thus revealing the cellular and subcellular distribution of GABAA receptor subunits. A distinct neuron-specific expression pattern of the α-subunit variants was observed in cerebral cortex and hippocampal formation, along with widespread expression of the γ2-subunit. Of particular interest was the prominent α2- and α3-subunit staining on the initial axon segment of pyramidal neurons. This protocol represents a major improvement for high-resolution studies of human brain tissue aimed at investigating morphological alterations underlying neurological diseases.
Keywords
A
Immunohistochemical methods have been significantly improved with the development of two novel approaches for increasing the sensitivity of antigen detection. The first technique, antigen retrieval through microwave irradiation, was designed primarily for histopathological analysis of formalin-fixed, paraffinembedded sections of human tissue (Shi et al. 1991). The high-temperature heating allows the retrieval of antigens masked by fixation. We have recently modified this procedure to enhance immunohistochemical staining of type A γ-aminobutyric acid (GABAA) receptors in rat brain tissue (Bohlhalter et al. 1996; Fritschy et al. 1998). The other approach, tyramide signal amplification (TSA), was first introduced for immunoassays (Bobrow et al. 1989,1991) and later adapted for immunohistochemistry at the light microscopic level (Adams 1992; Berghorn et al. 1994; Merz et al. 1995; Shindler and Roth 1996; Strappe et al. 1997; Van heusden et al. 1997; von Wasielewski et al. 1997), fluorescence in situ hybridization (Kerstens et al. 1995; Raap et al. 1995; De Haas et al. 1996; Macechko et al. 1997; Schmidt et al. 1997; Speel et al. 1997; van Gijlswijk et al. 1997), and electron microscopy (Mayer and Bendayan 1997). This technique employs horseradish peroxidase (HRP) bound, for example, to secondary antibodies to catalyze the in situ deposition of biotinylated tyramide. The biotin moiety is subsequently used for attachment of an avidin-conjugated fluorochrome or of avidin-conjugated HRP, which then catalyzes a color reaction. TSA has been reported to increase sensitivity five- to 1000-fold compared to conventional immunohistochemistry (Adams 1992; Werner et al. 1996).
In the present study we have established a protocol based on these two procedures to investigate the cellular and subcellular localization of GABAA receptor subunits in the human brain. These experiments were conducted as part of a larger project aimed at determining whether alterations in the expression of GABAA receptors contribute to the pathophysiology of epilepsy (Loup et al. 1997). GABAA receptors form hetero-oligomeric chloride channels assembled from a family of subunits encoded by at least 17 genes (α 1-6, β1-3, γ1-3, δ, ∊, ρ1-3) (for review Macdonald and Olsen 1994; Sieghart 1995; Mohler et al. 1997). Multiple GABAA receptor subtypes differing in subunit composition have been identified in rodent brain and allocated to distinct neuronal populations (Gao and Fritschy 1994; Gao et al. 1995; Fritschy and Mohler 1995; McKernan and Whiting 1996). Using antigen retrieval immunohistochemistry, a remarkable specificity in the synaptic targeting of GABAA receptor subunits was observed in hippocampal and neocortical neurons (Fritschy et al. 1998). It is not known, however, whether a similar organization of GABAA receptor subtypes occurs in the human brain.
The protocol presented here involves microwave irradiation to enhance antigen detection in immersionfixed human brain tissue obtained at autopsy or surgery. GABAA receptor subunits were then visualized by immunofluorescence staining in cerebral cortex and hippocampus using subunit-specific antibodies in combination with an optimized TSA technique. The results demonstrate a dramatic increase in sensitivity and a complete suppression of autofluorescence signals, allowing detailed high-resolution analysis of the subcellular distribution of major GABAA receptor subunits in both autopsy and surgical specimens.
Materials and Methods
Tissue Collection
Brains obtained at autopsy were dissected into 7- to 12-mm-thick blocks from cerebral cortex or hippocampal formation. The subjects, three men, had died at 40, 69, and 73 years of age and had no known history of neurological or psychiatric disease. The interval between death and tissue processing ranged from 8 to 16 hr. Specimens of cerebral cortex or hippocampus were resected from 15 patients with medically intractable frontal or temporal lobe epilepsy and collected immediately on resection. These procedures were performed in accordance with the Helsinki Declaration of 1975 and were approved by the Ethics Committee of the University Hospital Zurich. Tissue blocks were rinsed in PBS, pH 7.4, and immersion-fixed for 6-8 hr at 4C in a mixture of 4% freshly dissolved paraformaldehyde and 15% saturated picric acid in 0.15 M phosphate buffer, pH 7.4 (Somogyi and Takagi 1982).
Microwave Irradiation
Microwave pretreatment of the fixed tissue blocks was performed as described previously (Fritschy et al. 1998). Meninges and major blood vessels were removed to prevent damage to underlying tissue by shrinkage during the irradiation procedure. The tissue specimens were first incubated overnight in 0.1 M sodium citrate buffer, pH 4.5, at room temperature (RT). They were then transferred to a jar containing 80 ml of fresh buffer and irradiated in a household microwave oven at 650 W for 90-150 sec, depending on specimen size. Under these conditions it took approximately 60 sec for the fluid to start boiling. After cooling to RT, the tissue blocks were cryoprotected in 10, 20, and 30% sucrose in PBS over a period of 3-4 days, frozen at −28C in isopentane, and stored at −80C. Subsequently, sections cut at a thickness of 40 μm in a cryostat were collected in PBS and preincubated in 1.5% H2O2 in PBS for 10 minutes at RT to block endogenous peroxidase activity. They were then washed three times for 10 min in PBS and processed for immunohistochemistry (see below). Adjacent sections were stained with cresyl violet for histological examination. To determine the effect of microwave irradiation on GABAA receptor subunit staining, cortical and hippocampal samples were divided into two blocks. One block was microwave-irradiated as described above and the other was processed without overnight incubation and microwave irradiation in citrate buffer, while all other steps were left unchanged. Specimens for each treatment protocol were stained in the same session.
Immunohistochemistry
The antibodies tested included the monoclonal antibody bd-24 recognizing the human GABAA receptor α1-subunit (Schoch et al. 1985; Ewert et al. 1990) and polyclonal guinea pig antisera recognizing the GABAA receptor α2-, α3-, and γ2-subunits. The preparation and characterization of the polyclonal antibodies, raised against synthetic peptides derived from rat cDNA sequences, have been described previously (see Fritschy and Mohler 1995 for details). For each subunit, the amino acid sequences used as antigens were found to be identical in rat and human cDNAs.
Free-floating sections were processed for immunoperoxidase staining using the ABC method of Hsu et al. (1981). Briefly, the sections were incubated overnight at 4C with primary antibodies diluted in 50 mM Tris-saline at pH 7.4 containing 2% normal goat serum and 0.05% Triton X-100. The dilutions of the antibodies were as follows: α1-subunit (monoclonal antibody bd-24) 0.2 μg/ml; α2-subunit (affinity-purified) 1.3 μg/ml; α3-subunit (affinity-purified) 1.8 μg/ml; and γ2-subunit (crude serum) 1:1500. Sections were then washed three times for 10 min with Tris-saline and incubated for 30 min with biotinylated secondary antibodies raised in goat (diluted 1:300 in Tris-saline containing 2% normal goat serum and 0.05% Triton X-100). After washing three times for 10 min with Tris-saline, sections were reacted with the ABC complex for 30 min (Vectastain Elite kit; Vector Laboratories, Burlingame, CA), washed again three times for 10 min, and finally incubated with 0.05% diaminobenzidine tetrahydrochloride (Sigma; St Louis, MO) and 0.001% hydrogen peroxide diluted in Tris-saline at pH 7.7. The staining reaction was carried out at RT for 5-15 min and was stopped by transferring the sections into ice-cold PBS. Sections were then washed three times for 10 min with PBS, mounted on gelatin-coated slides, air-dried, dehydrated with an ascending series of ethanol, and coverslipped out of xylene. Control experiments for staining specificity included replacement of primary antibodies with nonimmune serum and preabsoption of the antibodies with 3-5 μg/ml of their respective peptide antigen (Fritschy and Mohler 1995). No specific staining was seen in either case.
Immunofluorescence staining was performed using the protocol described in Fritschy et al. (1998). Briefly, sections were incubated overnight with primary antibodies (α2-subunit 2.5 μg/ml; α3-subunit 3 μg/ml; and γ2-subunit 1:1000). They were then incubated with affinity-purified goat secondary antibodies (Jackson Immunoresearch; West Grove, PA) coupled to dichlorotriazinylaminofluorescein (DTAF; 1:100), carbocyanine (Cy2; 1:100), or indocarbocyanine (Cy3; 1: 300).
Immunofluorescence Staining with Tyramide Signal Amplification
The TSA kit used for the amplification procedure was obtained from NEN Life Science Products (Brüssels, Belgium). Free-floating sections were incubated overnight at 4C with primary antibodies diluted in 50 mM Tris-saline at pH 7.4 containing 4% normal goat serum and 0.05% Triton X-100. Several concentrations were tested, and the optimal dilutions of antibodies were as follows: α1-subunit (monoclonal antibody bd-24) 0.04 μg/ml; α2-subunit, 0.15 μg/ml; α3-subunit 0.09 μg/ml; and γ2-subunit 1:10,000. Subsequent steps were performed at RT. Sections were washed three times for 10 min with Tris-saline and incubated for 60 min with biotinylated secondary antibodies raised in goat (diluted 1:500 in Tris-saline containing 4% normal goat serum and 0.05% Triton X-100). After washing three times for 10 min in TNT buffer (0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, 0.05% Tween-20), sections were blocked with TNB buffer (0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, 0.5% NEN Blocking Reagent) for 30 min. They were then incubated for 30 min with streptavidin-HRP diluted at 1:2000 in TNB. After washing three times for 10 min in TNT, the sections were incubated with biotinylated tyramide in amplification diluent. Several concentrations (1:50, 1:75, 1:150) and incubation times (3 min, 7 min, 10 min) were tested. The best results were obtained with a biotinylated tyramide dilution of 1:75 and an incubation time of 10 min. Thereafter, sections were rinsed three times for 10 min in TNT and then TNB. They were then incubated for 30 min in streptavidin conjugated either to Cy2 or Cy3 (diluted 1:1000 in TNB). Finally, sections were washed three times for 10 min with PBS, mounted on gelatin-coated slides, air-dried, and coverslipped with buffered glycerol.
Data Analysis and Photography
Sections were analyzed with a Zeiss Axiophot microscope (Jena, Germany) equipped for brightfield and epifluorescence microscopy. Photomicrographs were taken with Kodak T-max 100 film (Rochester, NY). The same sections were also analyzed by confocal laser scanning microscopy (TCS 4D; Leica, Heidelberg, Germany) using Imaris software (Bitplane; Zurich, Switzerland) for image processing. Digital images were printed with a Fuji Pictrography 3000 digital image printer (Tokyo, Japan).
Results
Antigen Retrieval by Microwave Irradiation
The influence of microwave irradiation on GABAA receptor subunit immunoreactivity (IR) was evaluated in autopsy and surgical specimens of cerebral cortex or hippocampus using the monoclonal antibody bd-24, which recognizes the α1-subunit (Schoch et al. 1985; Ewert et al. 1990), and subunit-specific antisera against the α2-, α3-, and γ2-subunits. With the standard immunoperoxidase protocol, bd-24 yielded a highly specific and differentiated staining pattern for the α1-subunit, as described previously in the human hippocampal formation (Houser et al. 1988). The polyclonal antisera, however, produced strong background staining that largely masked the weak specific signals, as shown in Figure 1A for the α3-subunit in frontal cortex. Microwave crowave irradiation resulted in a marked increase of the signal-to-noise ratio by enhancing cell surface staining and decreasing nonspecific background staining. Thus, in microwave-irradiated tissue, the α3-subunit IR was found to label pyramidal neurons located in the deeper layers of the frontal cortex, revealing their dendrites extending towards superficial layers (Figure 1B). Whereas this pattern was largely conserved in frontal and entorhinal cortex, clear differences in staining intensity became apparent in the laminar distribution of α3-subunit IR. In frontal cortex there was intense and diffuse staining of the neuropil in Layer II and the superficial portion of Layer III, with more moderate staining in Layers V and VI (Figure 1B). In contrast, in the entorhinal cortex staining was intense in the deeper layers and light to moderate in Layers II and III. In addition, the cell islands of Layer II were devoid of α3-subunit IR (Figures 1C and 1D).
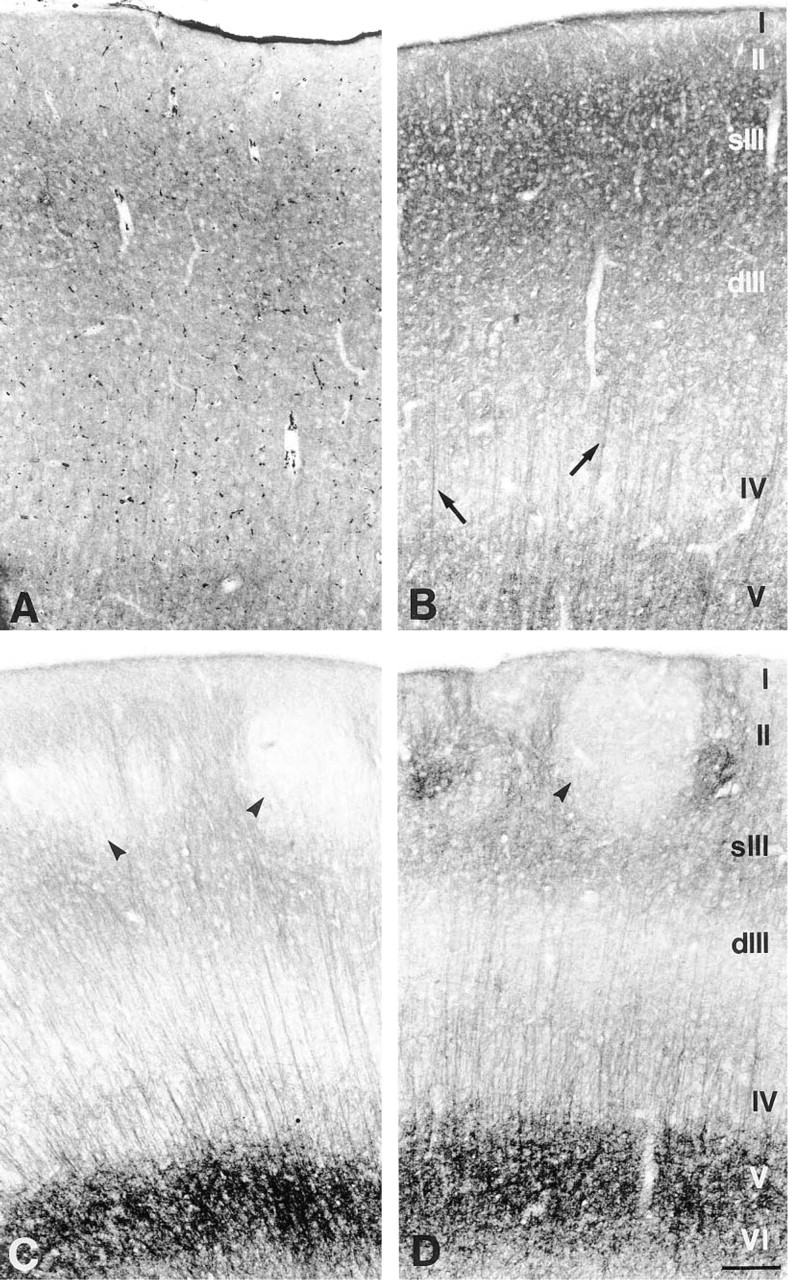
Effect of microwave irradiation on GABAA receptor α3-subunit IR in sections processed for immunoperoxidase staining. (
Pretreatment of fixed tissue with microwave irradiation proved to be equally effective for tissue obtained both at autopsy and at surgery, indicating that a postmortem interval of up to 16 hr did not significantly reduce GABAA receptor subunit IR. This is shown in the entorhinal cortex by comparing the GABAA receptor α3-subunit IR in tissue resected from a patient with intractable temporal lobe epilepsy (Figure 1C) and in control tissue obtained at autopsy (Figure 1D). In the surgical specimen, staining in layers II and III of the entorhinal cortex was slightly decreased compared with the autopsy sample (Figure 1C). This finding is most likely associated with the underlying chronic seizure disorder and may reflect neuronal cell loss in Layers II and III of the entorhinal cortex (Du et al. 1993). In the postmortem tissue sample, the α3-subunit IR was abundant, with a characteristic laminar distribution, although the patient had suffered a prolonged agonal state with progressive renal failure and the postmortem interval was 8 hr (Figure 1D). Furthermore, when the same region was compared in the three specimens obtained at autopsy, no significant difference in the intensity or specificity of GABAA receptor subunit staining was detected. For tissue obtained both at autopsy and at surgery, the best results were obtained when the 7-12-mm-thick tissue blocks were fixed for less than 8 hr and the irradiation time did not exceed 150 sec.
Increased Sensitivity and Elimination of Autofluorescence by TSA
For immunofluorescence staining, the microwave irradiation procedure alone was found to be inadequate because the strong autofluorescence occluded the weak GABAA receptor subunit IR. This is illustrated for the γ2-subunit in the CA2 region (Figure 2A) and in the dentate gyrus and hilus of the hippocampus (Figure 2D). The TSA procedure was then tested in sections pretreated with microwave irradiation. This resulted in a striking increase in immunofluorescence staining intensity for the four GABAA receptor subunits investigated and in the elimination of autofluorescence, as illustrated in Figures 2C and 2E. In the hippocampus, the pyramidal cell layer and the dendritic layers, as well as the interneurons, appeared intensely stained in sections processed with TSA, as shown for the γ2-subunit (Figure 2C). Prominent staining of this subunit was also seen in the dentate gyrus, with the somata of granule cells being clearly outlined and the molecular layer strongly labeled (Figure 2E). In addition, cell somata in the hilus of the hippocampus were intensely stained, together with their long processes (Figure 2E). A comparable improvement in the detection of GABAA receptor subunits was observed for the α2 and α3 antisera and for bd-24 recognizing the α1-subunit. The distribution of the α2–subunit IR was similar to that of the γ2-subunit, except that it was not detected in interneurons. The α3-subunit IR was particularly abundant in the CA1 region of the hippocampus, whereas the α1–subunit IR was most prominent in many interneurons distributed throughout the hippocampal formation. For all antibodies used, TSA led to a dramatic increase in sensitivity. For example, a 1:10,000 dilution of the γ2 antiserum yielded intense staining, whereas the signals were barely detectable with a 1:1000 dilution in conventional immunofluorescence staining.
The second major effect of TSA was suppression of autofluorescence, which was directly correlated with the duration of incubation with biotinylated tyramide. Although autofluorescence signals were reduced but still present after 3 min of incubation (Figure 2B), they had disappeared after 10 min of incubation with biotinylated tyramide (Figure 2C). The enhanced detection of GABAA receptor subunits and the elimination of autofluorescence were equally effective, regardless of the age at which the subjects had died or undergone surgery or of the premortem and postmortem conditions. Even in the oldest subjects (69 and 73 years of age) with the highest amount of lipofuscin granules, autofluorescence was totally suppressed.
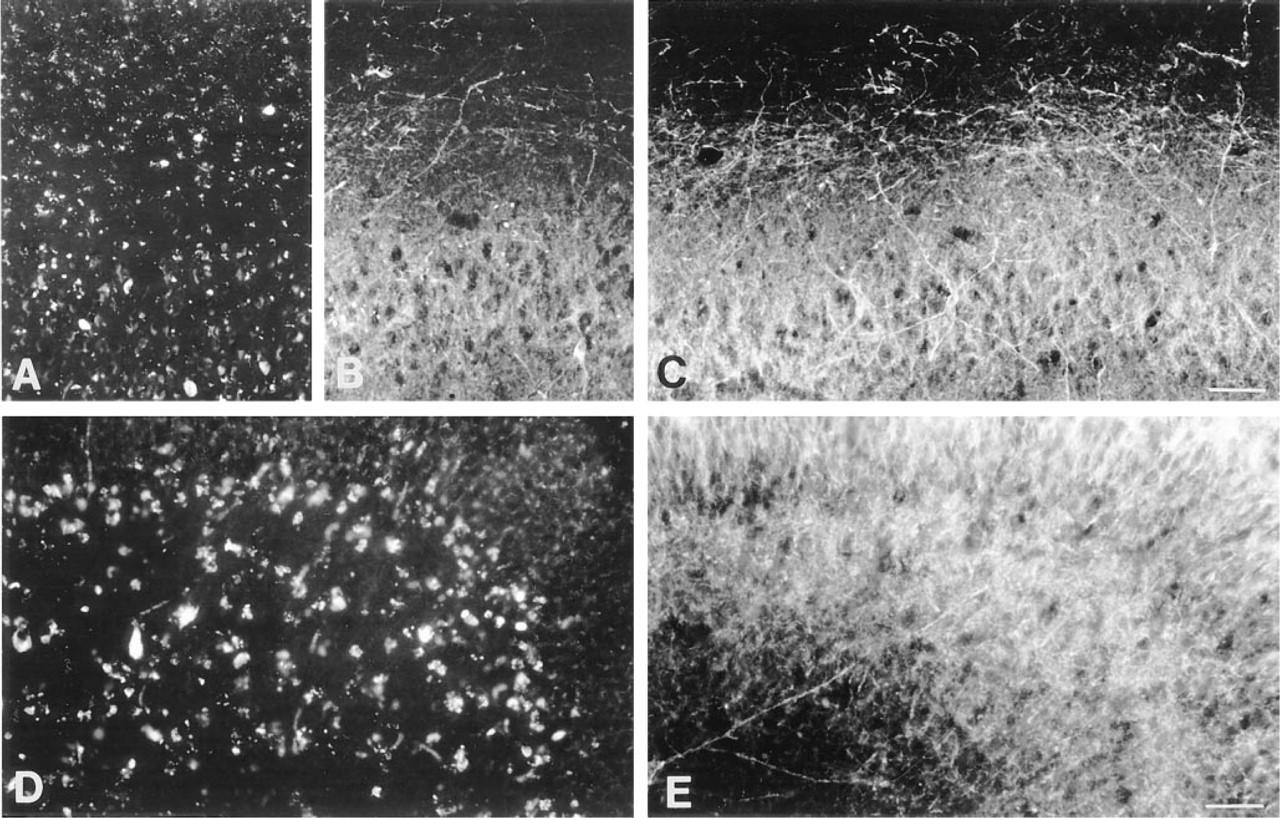
Effect of TSA on immunofluorescence staining of the GABAA receptor γ2-subunit. The tissue was obtained at autopsy from a 73-year-old man after a 16-hr postmortem interval, fixed by immersion for 8 hr and pretreated with microwave irradiation. (
Analysis of the Subcellular Distribution of GABAA Receptor Subunits
The strong immunofluorescent signals and the elimination of autofluorescence produced with TSA enabled us to investigate GABAA receptor subunit staining at the subcellular level with confocal laser scanning microscopy. Individual neurons were outlined along their surface by many puncta, most likely representing synaptic receptors, as shown for the γ2-subunit in CA1 pyramidal neurons (Figure 3A) and for the α2-subunit in granule cells of the dentate gyrus (Figure 3B). Particularly strong staining of the α1-subunit was observed in mossy cells (Figure 3C), revealing the characteristic “clusters of spheres” that form the excrescences of this hilar cell type described by Amaral (1978).
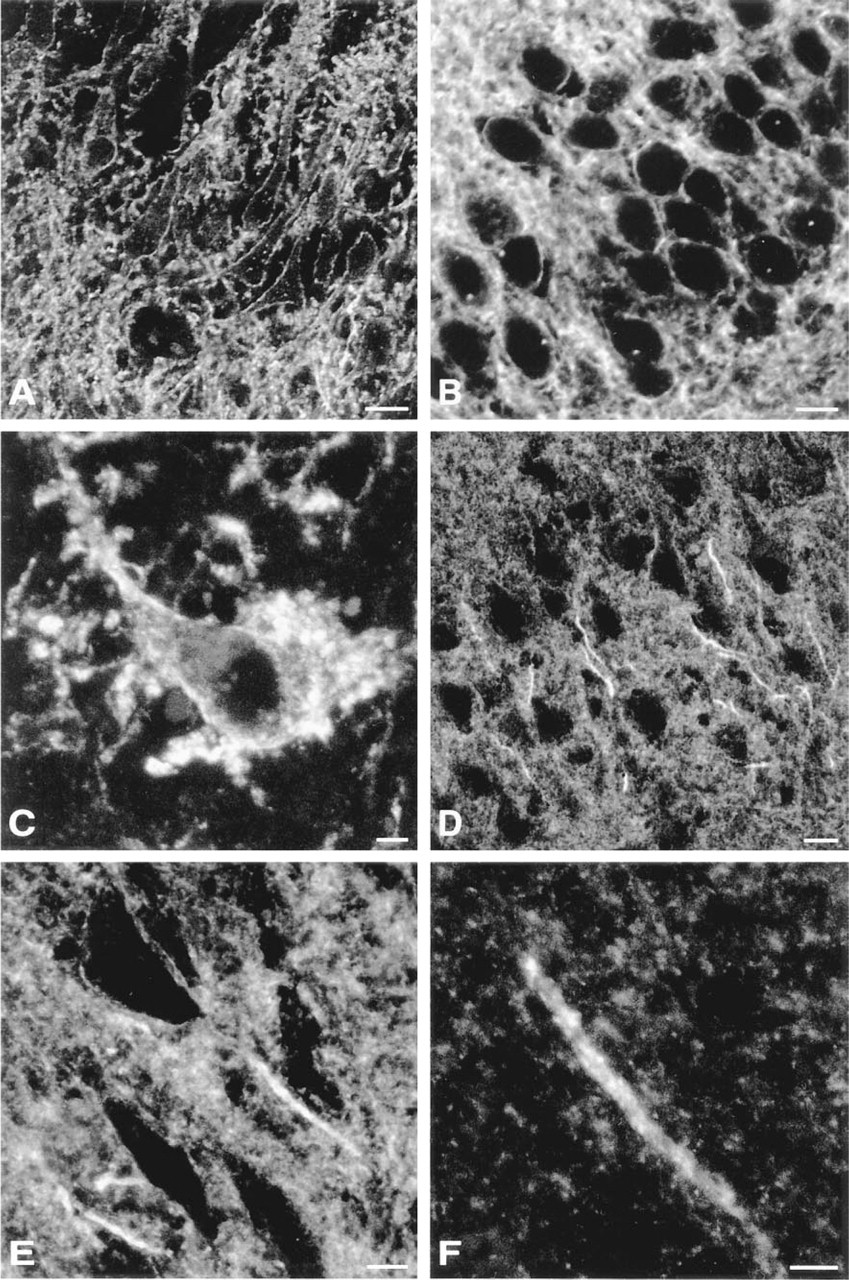
Digital images from confocal laser scanning microscopy illustrating the subcellular distribution of the GABAA receptor α1–, α2-, and γ2-subunit IR in hippocampal specimens obtained at autopsy. The sections were pretreated with microwave irradiation and processed for immunofluorescence with TSA. (
The most striking observation, however, was the intense α2-subunit IR on the axon initial segment (AIS) of pyramidal neurons in the hippocampus (Figure 3D-3F) and cerebral cortex (not shown). In contrast, the α3-subunit IR was mostly localized on the AIS of neocortical pyramidal neurons. The AIS of principal cells receives prominent GABAergic input from chandelier cells (Kosaka 1980; Somogyi et al. 1983). In rat brain, GABAA receptors were first reported at these synapses in ultrastructural studies using the postembedding immunogold technique (Nusser et al. 1996; Somogyi et al. 1996). Recently, these receptors were also visualized in lightly fixed cryostat sections of rat brain (Fritschy et al. 1998). Their detection in postmortem human brain tissue confirms the sensitivity and selectivity of the TSA procedure. In our study, the α2-subunit IR revealed the AIS as forming 20-40-mm-long tail-like structures outlined by many intensely stained hot spots as shown for the AIS of CA3 pyramidal neurons (Figures 3E and 3F). Typically, the staining started at some distance (10-20 mm) from the base of the soma. These features are similar to those reported for the rat, indicating that the subcellular distribution of the GABAA receptor α2-subunit in pyramidal neurons is largely conserved across species.
Discussion
The protocol described in this report combines microwave pretreatment of fixed tissue and the TSA technique to overcome two major obstacles associated with immunohistochemical studies of neurotransmitter receptors in human brain tissue. Microwave irradiation significantly increases the signal-to-noise ratio by enhancing cell surface IR and reducing nonspecific staining, whereas TSA results in a striking boost in sensitivity for antigen detection and eliminates autofluorescence originating from lipofuscin pigments. Our preliminary analysis using this protocol reveals a differential and neuron-specific expression pattern of the major GABAA receptor subunits in cerebral cortex and hippocampal formation at both the regional and the cellular level.
Effects of Microwave Irradiation
Antigen retrieval by microwave irradiation has been reported to be effective for a large variety of antigens masked in paraffin-embedded and vibratome sections of formalin-fixed archival tissue (for reviews, see Werner et al. 1996; Shi et al. 1997). However, a universal procedure applicable for every antibody is not available. Our protocol, which has been optimized for studying human brain tissue, differs in two major ways from those described previously (Evers and Uylings 1994a,b,1997; Yachnis and Trojanowski, 1994). First, the duration of microwave irradiation for visualization of GABAA receptor subunits did not exceed 2.5 min vs 10-15 min reported for other antigens (Werner et al. 1996; Shi et al. 1997). Second, the tissue blocks were immersed in aldehyde for less than 8 hr to minimize deleterious effects on tissue antigenicity (Fox et al. 1985), whereas archival tissue was fixed for up to several years (Werner et al. 1996; Shi et al. 1997). These results point to a relationship between fixation time and microwave irradiation time, as reported in a study examining human tonsil tissue (Munakata and Hendricks 1993).
The enhancement of cell surface IR observed for GABAA receptor subunits suggests that microwave irradiation facilitates the access of antibodies to their antigen, notably within synaptic clefts (Fritschy et al. 1998). This is in keeping with the fact that the antibodies employed in this study were raised against epitopes localized in the extracellular N-terminal domain of GABAA receptor subunits (Fritschy and Mohler 1995). Although the mechanisms underlying the reduction of nonspecific staining by microwave irradiation are not known, two factors appear to be involved. First, sites of nonspecific binding of secondary antibodies probably are neutralized, as suggested previously (Ainley and Ironside 1994; Fritschy et al. 1998). Second, endogenous peroxidase activity is blocked, as indicated by the lack of staining of major blood vessels (Figure 1). However, autofluorescence is only slightly reduced with microwave treatment alone (Figure 2).
Microwave irradiation was equally effective for both autopsy and surgical specimens, indicating that GABAA receptor subunit antigens are stable for several hours after death. Furthermore, the quality of staining is comparable to that reported for perfusionfixed rat brain tissue (Fritschy et al. 1998). Therefore, the application of this protocol to human brain tissue will permit detailed studies of neuropathological alterations with a degree of resolution and sensitivity that was previously achievable only in animal studies.
Effects of TSA on Immunofluorescence Staining
As a result of the remarkable increase in sensitivity induced by TSA, low concentrations of primary anti-bodies are sufficient for strong immunofluorescence staining. This represents a clear advantage, considering that polyclonal antisera may contribute substantially to background staining (Fritschy et al. 1998). The following variables in the TSA procedure were critical in determining the intensity of the staining signal: the concentration of the primary antibody, as stated above, the concentration of biotinylated tyramide, and the incubation time with biotinylated tyramide (i.e., duration of the enzymatic reaction leading to in situ deposition of tyramide). The second significant benefit provided by TSA was the elimination of autofluorescence originating from lipofuscin pigments, a major drawback in working with human brain tissue. In our study, autofluorescence was suppressed in all tissue specimens regardless of the age of the subjects. Although the mechanism leading to the elimination of autofluorescence remains unexplained, incubation of the sections in biotinylated tyramide for at least 10 min appeared to be essential (Figure 2). Therefore, the TSA procedure is ideally suited for visualizing proteins of relatively low abundance, such as ion channels or neurotransmitter receptors, and represents a major improvement for studies with both epifluorescence microscopy and confocal laser scanning microscopy.
Analysis of the Distribution of GABAA Receptor Subunits
To date, high-affinity monoclonal antibodies such as bd-24 and bd-17 (recognizing the α1- and the β2/β3- subunits, respectively) have provided the best results for immunohistochemical visualization of GABAA receptors in the human brain (Houser et al. 1988; Faull et al. 1993; Hendry et al. 1994). It is clearly important, however, to analyze the remaining major a-subunit variants (α2, α3, α5) because they represent receptor populations with distinct pharmacological profiles and with a characteristic distribution pattern in rodent brain (Fritschy and Mohler 1995). Furthermore, determination of the γ2-subunit distribution is of particular relevance, considering that this subunit, which is found in approximately 95% of GABAA receptors in rat brain (Benke et al. 1996), is essential for formation of the binding site for classical benzodiazepines. The preliminary results presented here provide the first demonstration of the α2- and α3-subunit IR in human brain. A number of similarities were observed in the expression pattern of these subunits compared to the rat. For example, in both rat and human, the α3-subunit predominates in deep cortical layers and is found in dendrites extending towards the pial surface, whereas the α2-subunit is very abundant throughout the hippocampal formation in pyramidal cells and dentate gyrus granule cells. The similarity extends to the subcellular distribution of receptor subtypes, with the α2-subunit IR being enriched in the AIS of hippocampal and neocortical pyramidal cells. However, clear differences were also observed. For example, the α3-subunit is particularly intense in the CA1 region of the human hippocampus whereas it is virtually absent in the rat (Fritschy and Mohler 1995).
The detection of presumptive synaptic GABAA receptors in human brain, as revealed by the intense punctate staining outlining cell bodies and dendrites and by the prominent α2- and α3-subunit IR on AIS of pyramidal cells, demonstrates both the sensitivity achieved with our protocol and the fact that these proteins are stable enough to withstand postmortem delays and tissue processing. A dysfunction in the GABAergic transmitter system is believed to contribute to the pathophysiology of epilepsy (for review see Olsen and Avoli 1997). Therefore, this protocol will permit the detection of potential changes in the synaptic distribution of GABAA receptors in human epileptic brain tissue, thereby providing further insights into the mechanisms underlying this common neurological disorder.
Footnotes
Acknowledgements
Supported by the Théodore OTT Fund.
We are grateful to Dr Hanns Möhler for his continuous support and we thank Drs Thomas Bächi and Matthias Höchli for their competent help with confocal laser scanning microscopy.