
Abstract
We previously reported that mast cells (MCs) serve as a source of basic fibroblast growth factor (bFGF), a potent angiogenic and mitogenic polypeptide, suggesting that bFGF may mediate MC-related neovascularization and fibroproliferation. Unlike many other growth factors, bFGF lacks a classic peptide sequence for its secretion, and the mechanism(s) for its release remains controversial. Because MCs release a wide spectrum of bioactive products via degranulation, we hypothesized that MC degranulation may be a mechanism of bFGF release and used ultrastructural immunohistochemistry to test the hypothesis. We reasoned that if bFGF is released through degranulation, it should be localized to MC secretory granules. Human tissues with chronic inflammation and rat/mouse tissues with anaphylaxis were studied. In all tissue samples examined, positive staining (or immunogold particle localization) for bFGF in MCs was predominantly in the cytoplasmic granules. Moderate bFGF immunoreactivity was also found in the nucleus, whereas the cytosol and other subcellular organelles exhibited minimal immunogold particle localization. In contrast, no immunogold particle localization for bFGF was observed in lymphocytes or plasma cells. In rat/mouse lingual tissue undergoing anaphylaxis, immunogold particle localization for bFGF was found not only in swollen cytoplasmic granules but also in the extruded granules of MCs. Three different anti-bFGF antibodies gave similar immunogold particle localization patterns, whereas all controls were negative. These results provide morphological evidence suggesting that, despite the lack of a classic secretory peptide in its structure, bFGF is localized to the secretory granules in MCs and may be released through degranulation.
B
Several lines of evidence indicate that the release of bFGF at both the cellular and the tissue level is tightly controlled and that loss of such control may be deleterious. First, increased free bFGF has been linked to neoplasms. For example, increased levels of bFGF were found in cerebral spinal fluid (CSF) in 62% patients with brain tumors and correlated well with the vascular density of the tumors. In contrast, bFGF was not detectable in the normal controls (Li et al. 1994). High-level expression of bFGF occurs in primary Kaposi's sarcoma (AIDS-KS), and tumor cells from Kaposi's sarcoma constitutively produce and release bioactive bFGF in the absence of cell death (Bailer et al. 1995; Samaniego et al. 1995). Second, a switch to the export of bFGF has been implicated in malignant transition in vivo. In a transgenic mouse model of fibrosarcoma, Kandel et al. (1991) found that although bFGF is expressed in normal dermis and in the early mild lesion of fibromatosis, a switch to the export of intracellular bFGF occurs concomitantly both with the neovascularization characteristic of aggressive fibrosarcoma seen in vivo and with the tumorigenicity of cell lines derived from the lesion. Third, NIH 3T3 fibroblasts transfected with native bFGF exhibit no morphological alteration or tumorigenicity despite expression of high levels of bFGF (Blam et al. 1988; Rogelj et al. 1988). However, cells transfected with the bFGF gene fused with a secretory signal sequence undergo morphological alteration and display tumorigenicity. The transformed cells cause rapidly growing tumors within 10 days in 100% of syngenic and nude mice. In nude mice, the tumors are highly vascularized (Rogelj et al. 1988,1989). Similar results are obtained with the human endometrial adenocarcinoma HEC-1-B cell line (Coltrini et al. 1995). When introduced into Xenopus embryos, bFGF mRNA for a secretory form of bFGF has been shown to be 100-fold more potent in eliciting aberrant mesoderm formation than its native counterpart (Thompson and Slack 1992). Results from these studies indicate that the mechanism(s) of bFGF secretion exerts a critical control of the bioactivities of this multipotent growth factor.
It has been well documented that cell injury results in bFGF release. However, injury also results in release of many other intracellular components and growth factors that are normally secreted, such as transforming growth factorb (TGF-β) and plateletderived growth factor (PDGF). Although the exact role(s) of bFGF remains unknown, its wide tissue distribution, broad spectrum of target cells, and mutation lethality in transgenic animals strongly suggest its important roles in normal homeostasis. It is reasonable to believe that release mechanisms other than cell injury play the major role in controlling the export of bFGF in normal homeostasis. A study by Mignatti et al. (1991) shows that bFGF can be released from single uninjured cells and mediates functions in an autocrine manner. Aortic smooth muscle cells release bFGFlike mitogen under shear stress (Sterpetti et al. 1994). In bFGF-transfected COS-1 cells, an energy-dependent, non-ER/Golgi pathway is reported to direct the export of bFGF (Florkiewicz et al. 1995). We and others demonstrated that mast cells express bFGF and serve as a major source of tissue bFGF in several disease conditions (Qu et al. 1995; Reed et al. 1995). Mast cells have been linked for almost two decades to neovascularization and fibroproliferative diseases. However, the exact contribution of mast cells to these disease processes remains largely unknown. Previous studies by Reed et al. and by us strongly suggest that bFGF mediates angiogenesis and fibroproliferation associated with mast cells (Qu et al. 1995; Reed et al. 1995). Because mast cells release several bioactive substances by degranulation, we hypothesize that bFGF can be released by mast cells through degranulation. In the present study we tested this hypothesis by immunolocalization of bFGF in secretory granules in both resting and activated mast cells. We found that bFGF immunoreactivity was localized in the cytoplasmic granules of mast cells in human tissues from lung, synovium, nasal mucosa, and skin, and in lingual tissues from rats and mouse. Immunogold particle localization for bFGF was also present in the extruded secretory granules of mast cells in rat/mouse tongue undergoing passive anaphylactic reaction. The results provide morphological evidence for bFGF release through a noninjury mechanism.
Materials and Methods
Tissue Processing
Fresh synovium (n = 4), lung (n = 3), and nasal mucosal membrane (n = 2) samples were obtained during surgery from patients with rheumatoid arthritis, pulmonary fibrosis, and nasal polyposis, respectively. All samples were divided into two parts and processed for light and electron microscopy separately. For light microscopy (LM), the samples were fixed in neutralized buffered formalin for 24-30 hr at 4C, dehydrated, and embedded in paraffin.
For electron microscopy (EM), tissue samples were sliced into 1-3-mm3 pieces and fixed with 4% paraformaldehyde-0.5% glutaraldehyde in 0.1 M Sörensen's phosphate buffer for 4 hr at 4C. From each case, some sample pieces were also postfixed in 1% OsO4 for 1 hr at room temperature (RT). After extensive washes in the phosphate buffer, the samples underwent ethanol dehydration to propylene oxide and then were embedded in Araldite 502 (Electron Microscopic Sciences; Ft Washington, PA).
In Vivo Mast Cell Degranulation
To induce degranulation in vivo, direct lingual anaphylactic assay was performed on female Brown Norway rats weighing 250-300 g (n = 4). Rats were anesthetized with isoflurane/O2, delivered with an anesthetic vaporizer, and about 15 μg goat anti-rat IgE antibody in 100 μl PBS was injected into the tongue along the midline. The control rats (n = 3) were injected with an equal amount of nonimmune goat IgG. Anesthetized rats were sacrificed 1 or 2 hr after the injection by
In vivo mast cell degranulation in mice was induced using a passive cutaneous anaphylactic assay previously described by Wershil et al. (1991). Briefly, 20 μl serum-free Dulbecco's modified Eagle's medium (DMEM) containing 100 ng of mouse monoclonal IgE anti-dinitrophenyl (DNP) (Sigma; St Louis, MO) was injected into the ear pinna of 3-month old Balb-c/CBYJ male mice (Jackson Labs; Bar Harbor, ME) anesthetized with isoflurane/O2. After 30 hr, the mice were challenged
Histochemical and Immunohistochemical Staining
Paraffin-embedded tissues were cut into 2-3-μm sections and laid on poly-
Postembedding Immunogold Labeling for Electron Microscopy
Ultrathin sections were cut at 50-70 nm, collected on 200- mesh nickel grids coated with Coat-Quick “G” (Electron Microscopic Sciences), and immunolabeled with three different antibodies specific for bFGF, using a two-step procedure. All immunogold steps were carried out at RT in 70-μl reagent drops on moisture-resistant parafilm (American National Can; Greenwich, CT). The grids were first treated with hyaluronidase at 2 mg/ml in 0.1 M acetate buffer, pH 5.2, for 15 min to unmask antigens (Qu et al. 1995). After two 5-min washes in Tris-buffered saline (TBS), pH 7.5, containing 0.1% Triton X-100, nonspecific binding sites were blocked by incubation for 20 min in blocking solution as described above. Then the grids were incubated in a primary anti-bFGF antibody (see Table 1) in the blocking solution for 2 hr, followed by two 5-min washes and one 30-min wash with TBS and 7.5- and 5-min washes in TBS buffer, pH 8.2. The grids were incubated for 1 hr with a goat antirabbit IgG antibody conjugated with 5- or 10-nm gold particles (Goldmark Biologicals; Philipsburg, NY) in TBS buffer, pH 8.2. After several washes in TBS and dH2O, the grids were air-dried and then counterstained with aqueous uranyl acetate. Controls included substitution of nonimmune IgG for primary antibodies, preabsorption of the primary antibodies with recombinant bFGF, or omission of the primary antibody.
Results
Localization of bFGF in Rat and Mouse Tissue After Cutaneous Anaphylaxis
On intralingual injection of anti-Ig E antibody or
Antibodies used for the study
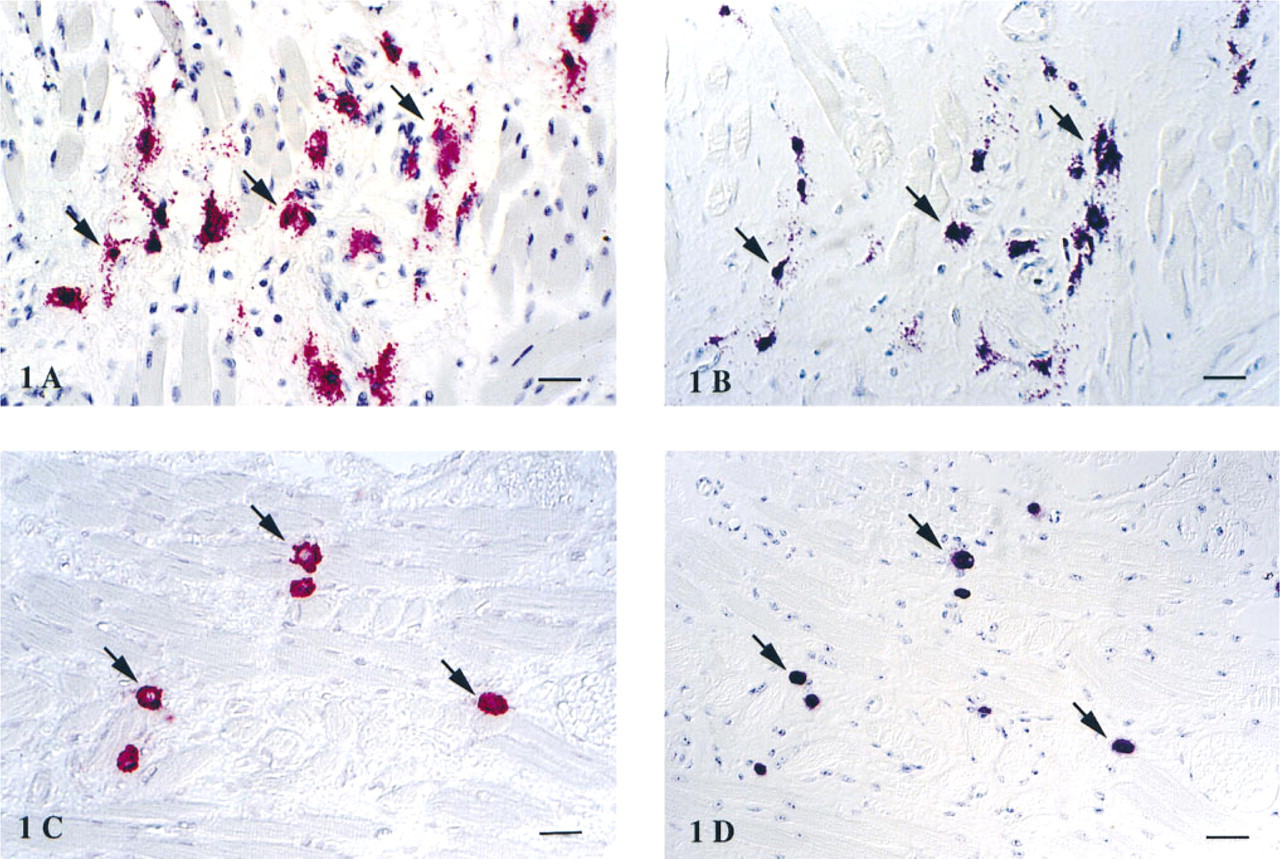
To identify the subcellular compartment associated with bFGF-like immunoreactivity, tissue samples were examined by EM. Tissue mast cells were identified by metachromasia after toluidine blue staining of semithin sections and by characteristic ultrastructure, such as typical plicated membrane folds, conspicuous membrane- bound cytoplasmic granules, and nonlobed nucleus (Figure 2). Immunogold particle localization of bFGF was found predominantly in the cytoplasmic granules of MCs (Figure 2). However, no particular distribution pattern of the bFGF immunoreactivity within the cytoplasmic granules was observed. Similarly, moderate bFGF immunoreactivity was also found in the nucleus, whereas the cytosol and other subcellular organelles were essentially devoid of immunogold particles for bFGF in MCs. Only background level of immunogold particle localization was found in cytoplasm and mitochondria (Figure 3A). Moderate to little immunogold particle localization was present in the cytoplasm and nucleus of fibroblasts and endothelial cells (data not shown). In contrast, no immunogold particle localization for bFGF was observed in lymphocytes or plasma cells.
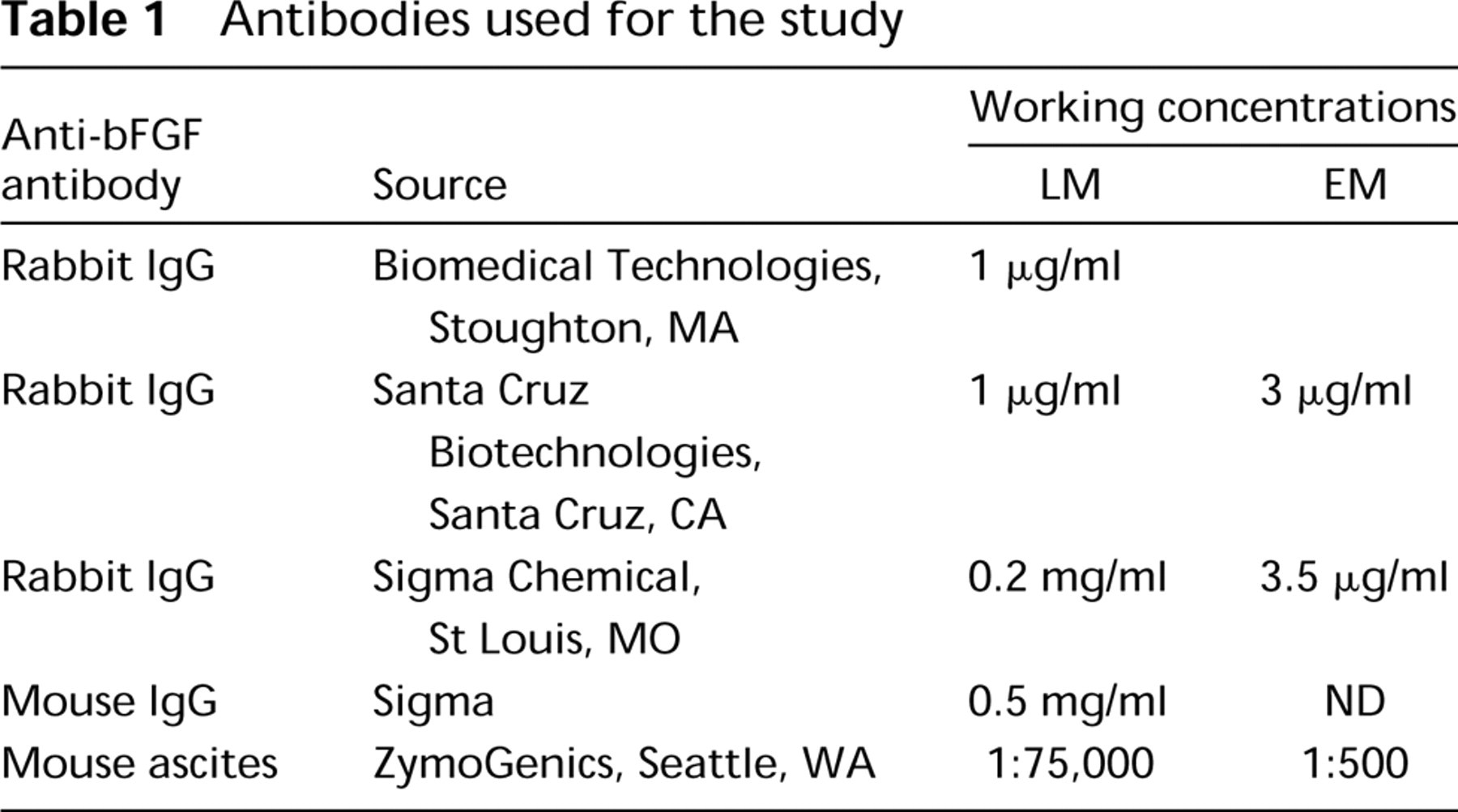
Localization of mast cell-associated bFGF by immunohistochemistry. Paired sequential sections at 2-μm thickness of lingual tissue from rat with anaphylaxis (
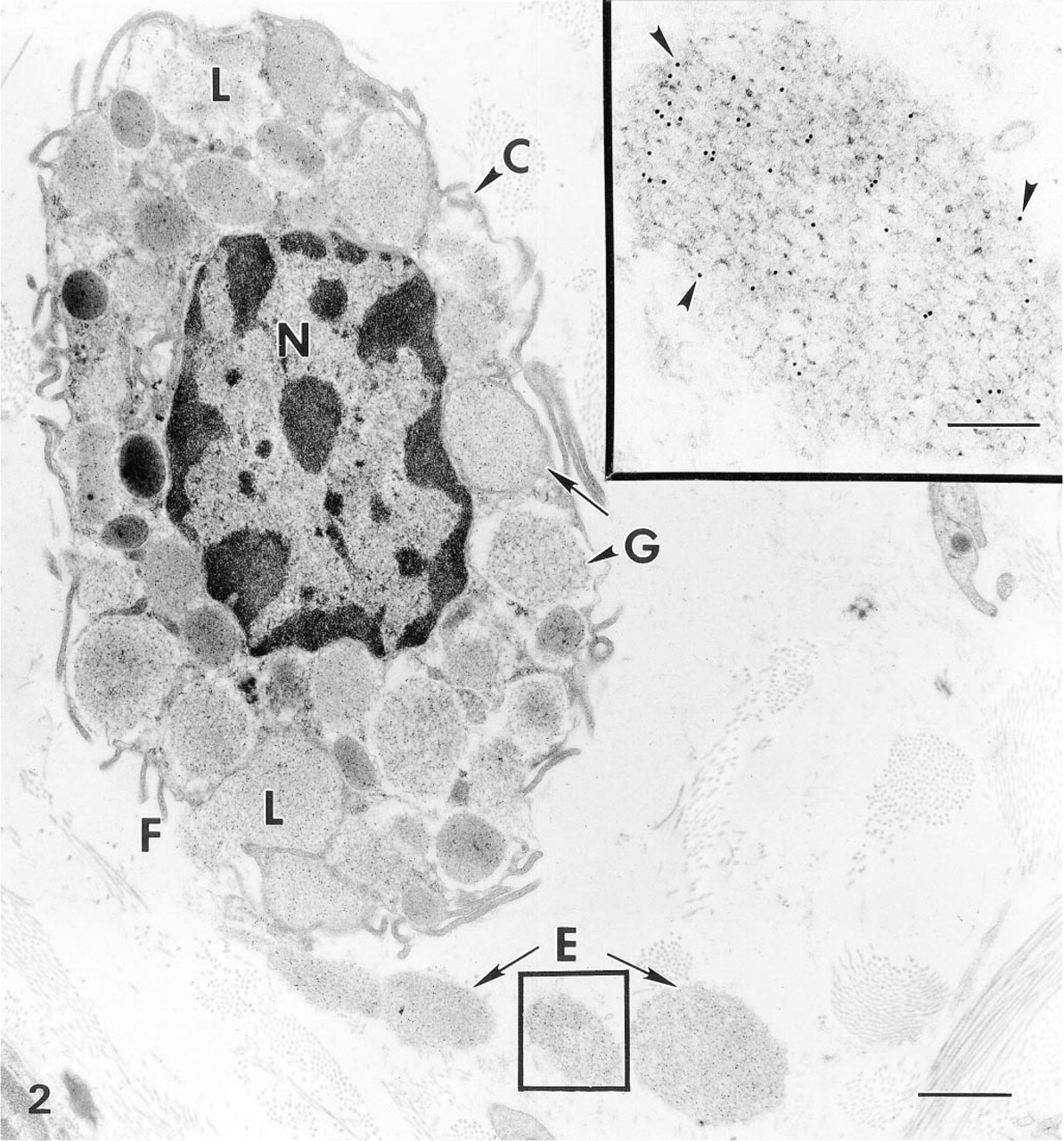
Immunogold particle localization of bFGF in MC of mouse lingual tissue stimulated with anti-IgE antibody for 2 hr in vivo: Characteristic MC membrane folds (F), granules (G), and round nucleus (N). In addition, this MC shows degranulation features such as channel formation (C), swollen granules (L), and granule extrusion (E). Immunogold particle localization for bFGF is present in both the cytoplasmic granules (G) and extruded granules (E). Bar = 1.2 μm. (
After degranulation, mast cells in the anaphylactic tissue exhibited morphological changes characteristic of degranulation, such as degranulation channel formation, swollen granules, shed surface folds, and granule extrusion (Figures 2 and 3A). The extruded granules were membrane-free and exhibited reduced matrix density. Extensive granule extrusion was observed (Figure 3A). Immunogold particle localization of bFGF was present in cytoplasmic secretory granules of the degranulating mast cells. Notably, bFGF immunogold particle localization was also present in the extruded granules (Figures 2 and 3A).
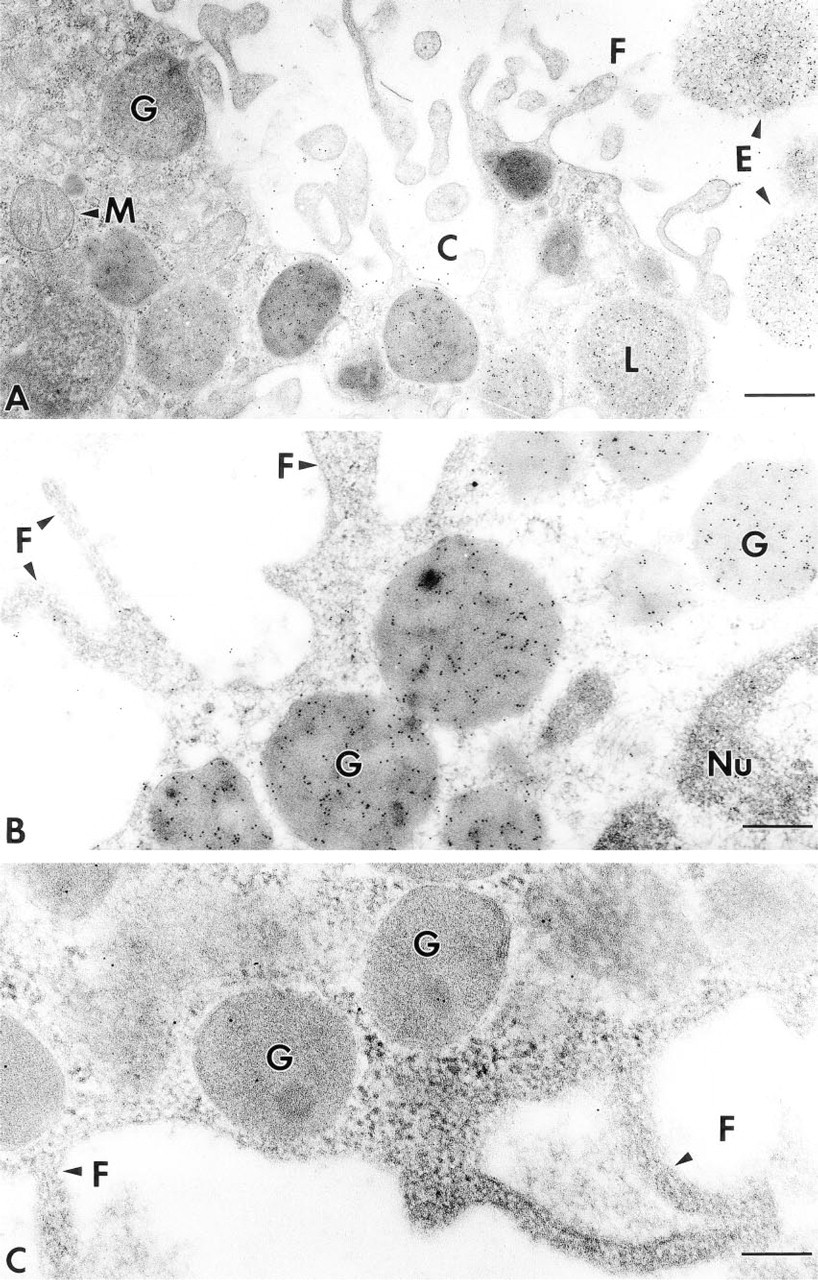
Immunogold particle localization for bFGF in MCs. (
Localization of bFGF in Human Tissues
In all samples examined, immunogold particle localization for bFGF was present in mast cells. Specific bFGF staining in the human tissues at the LM level was reported in detail our previous article (Qu et al. 1995). To determine subcellular localization of bFGF-like molecules, tissue samples were examined by immunoelectron microscopy. Tissue mast cells were identified by their metachromasia and morphological features as described above, and by positive labeling of the cytoplasmic granules by anti-tryptase antibody (data not shown). When examined by EM after immunogold labeling with anti-bFGF antibodies, immunogold particle localization in mast cells was found in all tissue specimens, and all mast cells examined demonstrated immunoreactivity to the anti-bFGF antibody. The localization was predominantly present in the cytoplasmic secretory granules and nucleus (Figure 3B). Immunogold particle localization of bFGF did not appear to assume any particular distribution pattern within the cytoplasmic granules. No apparent immunogold particle localization of bFGF was found in the cytoplasm or cell plasma membrane (Figure 3B).
Controls to Confirm the Specificity of bFGF Staining
To confirm the specificity of bFGF staining present in mast cells, cornu ammon 1 and cornu ammon 2 regions from primate and rat hippocampus known to contain bFGF were used as a positive control tissue. Previous studies by several independent research groups showed that both mRNA for bFGF and immunoreactivity, as well as biological activity, were present in neurons in these regions (Emoto et al. 1989; Eckenstein et al. 1994). Strong immunolabeling by antibFGF antibody was observed in the nucleus of cells in these regions (data not shown), consistent with previous findings reported by others (Emoto et al. 1989; Eckenstein et al. 1994). All anti-bFGF antibodies tested gave similar immunogold particle localization patterns at tissue and subcellular levels in both human, rat, and mouse tissues. Preabsorption of anti-bFGF antibody (Sigma F-3393) with recombinant human bFGF greatly reduced the positive labeling intensity in both cytoplasmic granules and nucleus of MCs (Figure 3C). Omission of primary antibody or substitution of a bFGF-specific antibody with nonimmune IgG or an irrelevant antibody from the same species as the specific antibody completely abolished the immunogold particle localization (data not shown). Specific immunogold particle localization for bFGF was not affected by preabsorption of any protein-containing solution with heparin beads, suggesting that the immunogold particle localization was not due to exogenous bFGF from the solution/reagents used.
Discussion
In this article we present morphological evidence suggesting that mast cell-derived bFGF can be released through degranulation, a classical exocytotic process characteristic of noninjured, properly stimulated mast cells. Because bFGF is often found extracellularly and may be readily available to mast cells, one can argue that mast cells could take up extracellular bFGF and store it in their granules. Two findings appear to refute such an explanation. First, unlike epithelial cells of the tongue and airways that express both bFGF and FGF receptor-1, −2, and −3 (Hughes and Hall 1993), mast cells exhibit no apparent immunostaining for these receptors (our unpublished observation). Second, exogenous bFGF uptake by cells in culture via either FGF receptors or heparin sulfate proteoglycans is directed into multivesicular bodies in endothelial cells (Gleizes et al. 1995), which are morphologically distinct from secretory granules and were not seen in mast cells examined. Although crossreaction of the anti-bFGF antibodies to other members of the FGF family with secretory peptides cannot be completely ruled out, the identical immunogold particle localization patterns given by different anti-bFGF antibodies and the abolishment of the selective localization by preabsorbing the antibody with excess bFGF indicate that this is unlikely. Previous studies by Reed at al. and by us have also shown that both tissue MCs and MC lines contain mRNA for bFGF (Reed et al. 1995; Qu et al., 1995,1998). Finally, because of high bFGF-heparin affinity, exogenous bFGF from BSA and normal sera used for blocking nonspecific binding may bind to mast cell granules and give rise to false but highly specific immunogold particle localization. However, the bFGF level in the blocking solution used was undetectable using a commercial ELISA kit with detection sensitivity at 5 pg/ml (unpublished data). Moreover, the blocking solution was absorbed with heparin acrylamide beads before use. Therefore, possible contamination of tissue sections by exogenous bFGF is unlikely.
Mast cells have been implicated in a wide spectrum of diseases characterized by fibroproliferation and neovascularization (for review see Rother et al. 1990; Galli et al. 1993). Notable examples include pulmonary fibrosis (Schulman 1993), rheumatoid arthritis (Gruber and Kaplan 1993), scleroderma, and GVHD (Claman 1990). Active mast cell degranulation has been noted at least in scleroderma (Claman 1990; Irani et al. 1992). Studies have directly linked mast cell degranulation to fibrotic and angiogenic changes in vivo. For example, Gordon and Galli (1994) have demonstrated that mast cell degranulation elicited by passive cutaneous anaphylaxis resulted in increased collagen gene expression by mouse fibroblasts. Such an increase occurs before apparent influx of neutrophils and other inflammatory cells, indicating that factors derived from the resident cells are responsible. Although the PCA animal model was used to demonstrate the release of bFGF through mast cell degranulation in the present study, extensive studies by Dvorak et al. have shown that a slow degranulation process, piecemeal degranulation, is also present and is clinically relevant to diseases characterized by fibroproliferation and neovascularization (for review see Dvorak 1992).
In addition to cell injury, bFGF release has been described in two other experimental systems in which noninjury release is also contemplated (Mignatti et al. 1991; Sterpetti et al. 1994). Because bFGF is localized in the cytoplasmic secretory granules of mast cells that also contain other secretory proteins such as tryptase, the intracellular trafficking and release mechanism of bFGF in mast cells may differ from those described in two other cell types. Therefore, it appears that different types of cells may use different transport-export systems for bFGF release. Unlike the studies described above, our findings suggest that bFGF is sorted into the classical endoplasmic reticulum/Golgi route. Results from a study by Rubartelli et al. (1990) have indicated that interleukin-1 β, a multifunctional cytokine that also lacks a classical secretory peptide, is also targeted into the classical protein secretory pathway. It is still unknown from the present study how bFGF is targeted into the classical secretory pathway. We believe that there are several plausible mechanisms. The first potentially explanatory finding emerges from a study of another prominent member of the heparin binding growth factor family, acidic FGF, which also lacks a classical secretory sequence. Preliminary results from the study by LaVallee et al. (1996) indicate that intracellular aFGF forms a complex with a protein with high homology to synaptotagmin, a synaptic vesicle protein involved in exocytosis by regulating neurotransmitter uptake, vesicle targeting, and fusion with presynaptic membranes (for review see Elferink and Scheller 1995). Interestingly, the Type II synaptotagmin mRNA was also found in cornu ammon 1-3 areas of the hippocampus of the brain where bFGF is localized (Emoto et al. 1989; Marqueze et al. 1995; Williams et al. 1996). Synaptotagmin is also found in the anterior pituitary of rat, hamster, and guinea pig, a location in which bFGF is present (Redecker et al. 1995). “Secretory lysosomes” may offer a second plausible explanation. Compelling evidence has suggested that cells of the hematopoietic lineage may use special sorting and secretory mechanisms in which the lysosome functions as a regulated secretory granule (for review see Griffiths 1996). In these cells, the secretory lysosomes may transport the proteins that lack a classical secretory sequence as a sorting signal directly into the ER/Golgi route by fusing with different compartments of the pathway. Finally, mast cells are rich in heparin and chondroitin sulfate, which are localized in the secretory granules. It is possible that bFGF employs these proteoglycans as transporting vehicles owing to their high bFGF binding affinities. Although results from the present study cannot provide the answer to how bFGF is sorted into the classical secretory pathway, they strongly suggest that, despite the lack of a classic secretory peptide in its structure, bFGF is localized to the secretory granules in MCs and is released together with the granules via degranulation.
Footnotes
Acknowledgements
Supported by NIH grant EY 10572, by a grant from the Gerlinger Fund, and by an unrestricted grant from Research to Prevent Blindness, New York. JTR is a senior scholar supported by Research to Prevent Blindness, New York. SRP is a recipient of an RPB Special Scholar Award, New York.
We wish to acknowledge the helpful critical review and important insights freely given this manuscript by Dr Ann Dvorak.