
Abstract
Keywords
A
Microscopic evaluation of apoptosis, although somewhat cumbersome, remains the most reliable system of detection. Although many probes are available for DNA staining in confocal microscopy (Suzuki et al. 1997), none allows continuous visualization of living cells undergoing apoptosis. The opportunity for direct imaging of viable cells throughout the process is relevant under several conditions in addition to strictly morphological studies. For example, direct visualization is crucial in mixed cultures or when only a very small percentage of cells enters apoptosis or when it is important to follow the time course of the process, which is often variable, depending on both the cell type and the apoptogen stimulus employed. In this regard, the importance of a multiprobe assessment of apoptosis, especially for cytometry, has been recently stressed (Frey 1997).
Calcein-AM proved to be both specific and sensitive for detection and tracking of apoptosis in living cells by confocal laser microscopy (Bussolati et al. 1995). This neutral vital dye is loaded and rapidly converted by cell esterases into its negative, impermeant fluorescent analogue. The nucleus-cytoplasm signal intensity ratio is approximately 3:1 and allows clear visualization of both structures. In cells undergoing apoptosis, early chromatin condensation is read as a sharp nuclear signal increase, and initial cell shrinkage is also visualized. When chromatin begins to fragment and eventually is segregated within blebs, the process can be tracked stepwise in real time. At the same time, it is possible to check membrane integrity, whose preservation is one of the most significant features of apoptosis with respect to necrosis, because in the presence of membrane defects calcein leaks out of the cell and the signal also vanishes in the presence of residual esterase activity (Morris 1990; Weston and Parish 1990). In other words, the entire process can be followed without “blind” points.
In this study we compared the performance of annexin V-FITC and calcein-AM as early apoptosis probes in confocal microscopy. Two different cell lines, PC12 and NIH3T3, were studied in a self-built flow chamber that allows long-term observation of living adherent cells under controlled microenvironmental conditions and repeated culture medium changes without modification of a preset observation field (Dall'Asta et al. 1997). Particular attention was devoted to ascertaining the relationship between annexin V-FITC positivity and maintenance of cell adhesion.
Materials and Methods
Cell Cultures and Treatment
Rat pheochromocytoma PC 12 cells (2 × 104cells/cm2) were grown on coverslips coated with poly-
Mouse NIH3T3 fibroblast cells (2 × 104 cells/cm2) were grown on coverslips in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% calf serum. For apoptosis induction (Bussolati et al. 1995), 24 hr after seeding coverslips were placed in the above-mentioned flow chamber and incubated in complete growth medium supplemented with 5 IU/ml
All reagents for cell culture were purchased from Life Technologies (Gaithersburg, MD). All other chemicals were supplied by Sigma Chemical (Milan, Italy).
Confocal Microscopy
Our images were acquired with a Multiprobe 2001 Molecular Dynamics confocal laser scanning microscope (CLSM) (Sunnyvale, CA) based on a Nikon diaphot inverted microscope and equipped with an argon laser. We employed × 40 NA 0.95 dry and × 100 NA 0.75 oil immersion Planapo lenses coupled to a 100-μm pinhole according to the cell type under investigation. Image processing was performed on a Silicon Graphics Personal Iris workstation (Image Space Software; Molecular Dynamics).
As stated above, our flow chamber allows repeated medium substitutions with stability of the observation field both laterally (x-y-axis) and vertically (z-axis).
Once plated in the flow chamber, cells maintained in normal growth medium do not show morphological signs of distress, adhesion defects, or reduction of cell number up to 24 hr of brightfield observation.
Probe Loading
After 4 hr of staurosporine and 12 hr of
After 10 min of loading an observation area was chosen in brightfield and section series was acquired with z-step set at 1 μm.
Samples were excited with the 488-nm laser line of CLSM and the emission recorded through a 510-nm primary beamsplitter and a 530 nm DF barrier filter. To assess the presence of propidium iodide in the nuclei, emission over 560 nm was also always checked.
After series acquisition, calcein acetoxymethylester (calcein-AM; Molecular Probes, Eugene, OR) was added to the culture medium in the flow chamber at a final concentration of 2 μM. The dye was taken from a stock solution (1 mM) in dimethylsulfoxide (DMSO). After 10 min of loading, the same field was scanned again with the same filter setting. The signal-to-noise ratio is very high and therefore confocal image acquisition is possible at very low laser power (well below 1 mW).
In another experimental setting, NIH3T3 cells were treated for 12 hr in
In all experiments, PC12 and NIH3T3 cells incubated in the absence of apoptogenic stimuli were dye-loaded and used as controls. No toxic effects were observed under these conditions at the experimental times. For each cell type, experiments were replicated three times with comparable results.
Results
PC12 Cells
Xia et al. (1995) have reported a steep increase in apoptotic PC 12 cell numbers starting in the third hour of incubation with staurosporine. Pilot experiments carried out in our laboratory have shown that after 2 hr of incubation the addition of annexin V-FITC does not yield positive labeling, whereas, after 4 hr, a clear-cut membrane staining becomes evident in some cells (Figure 1A). All the cells in the culture are negative for propidium iodide staining.
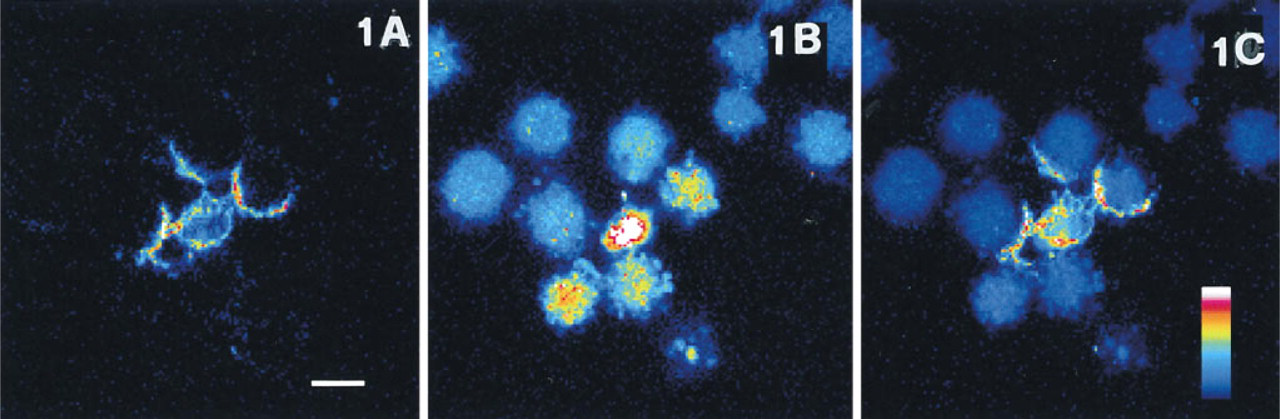
Confocal section showing PC12 cells incubated for 4 hr in the presence of 1 μM staurosporine and loaded for 10 min with 1 μg/ml annexin V-FITC and 2 μg/ml propidium iodide.
According to our experimental protocol, we added calcein-AM to the culture medium and the entire cell population in the field became appreciable within 10 min (Figure 1B). A heterogeneous pattern could be observed, corresponding to the asynchronous progression of the population through the process. Only a few cells show normal probe distribution, and many elements have intense nuclear fluorescence due to chromatin condensation. Cells that have further progressed present a variable amount of blebs containing highly condensed chromatin. The membrane integrity of the cell remnants is preserved, as can be inferred by the persistence of calcein signal in cytoplasm.
To compare the behavior of the two probes, we have merged the annexin V-FITC and the calcein images (Figure 1C). The latter was evenly attenuated to 60% of the original intensity, because the exceedingly higher efficiency of calcein would overwhelm the weaker signal from annexin V-FITC and render it unappreciable. It is evident that annexin V-FITC labeled only the cells in the early phases (i.e., chromatin condensation, initial blebbing), whereas other cells that have further proceeded (i.e., intense blebbing) are annexin V-FITC negative, as are apparently normal cells.
NIH3T3 Cells
We have previously shown that
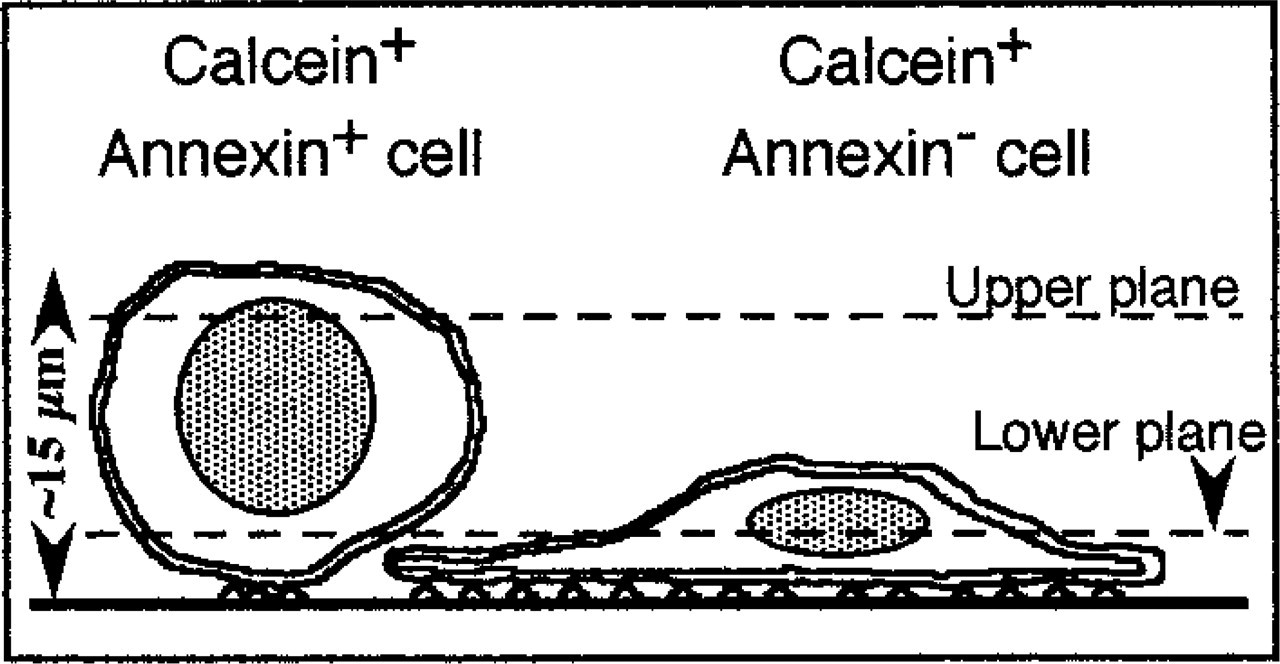
Schematic side view of the situation imaged in Figure 3. Dashed lines represent upper and lower section planes that correspond to images
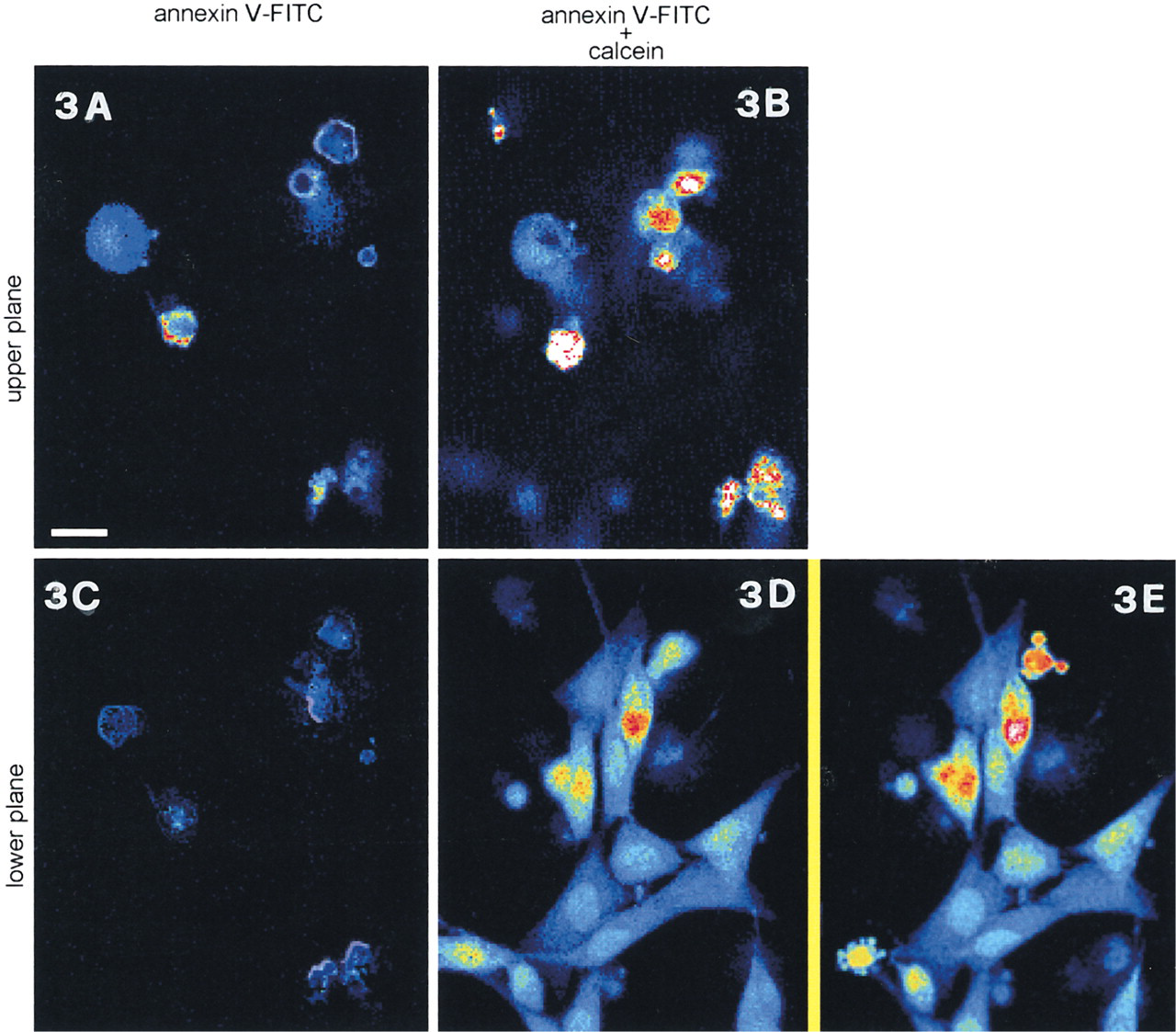
NIH3T3 cells were incubated for 12 hr in the presence of 5IU/ml L-asparaginase and then loaded for 10 min with 1 μg/ml annexin V-FITC and 2 μg/ml propidium iodide. Confocal sections of the same microscopic field at two different scansion levels are shown.
In cells harvested by trypsin digestion after 12 hr of treatment with
Discussion
Morphological evaluation of apoptosis is still the less biased procedure available; it should therefore precede any quantitative assessment of the process in previously untested cells.
Annexin V-FITC is presently employed, both in cytometry and microscopy, as an early marker for apoptosis. This method has been proven reliable in suspended cells, whereas its efficiency in adherent cultures is uncertain and only one report is available on this specific matter (van Engeland et al. 1996).
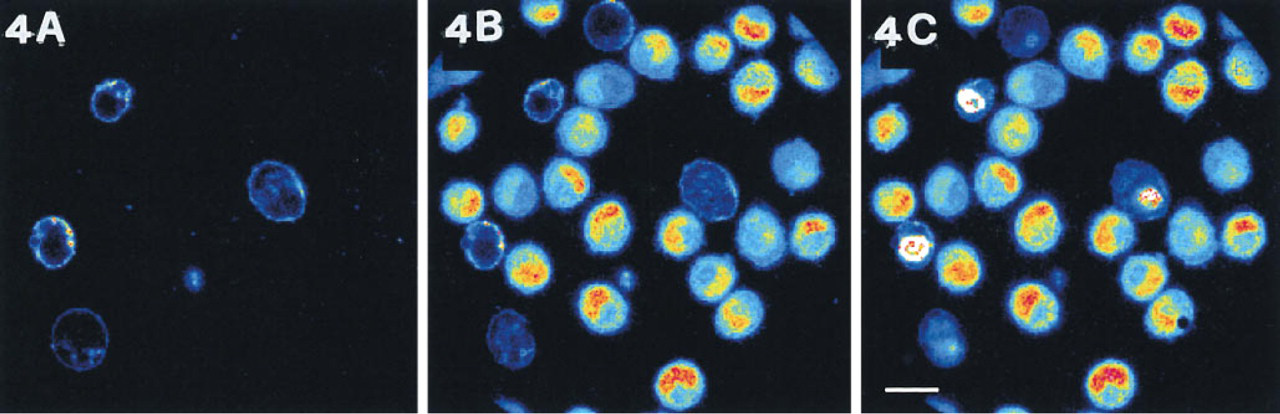
(A) Confocal image of NIH3T3 cells treated for 12 hr with L-asparaginase, trypsinized and loaded in suspension for 10 minutes with 1 μg/ml annexin V-FITC and 2 μg/ml propidium iodide.
Calcein-AM provides morphological evidence of chromatin condensation and segregation in blebs, together with functional information about plasma membrane competence (Bussolati et al. 1995).
Employing confocal microscopy, we have compared these two different morphological approaches in two cell lines with different adhesion features: PC12 and NIH3T3. The former are not intrinsically adherent and also, on poly
As expected, a significant percentage of PC 12 cells exhibit both PS exposure and condensed chromatin after 4 hr of incubation with staurosporine. Rather surprisingly, however, other cells have further progressed through the apoptotic process, as demonstrated by calcein pattern, but are negative for annexin staining. It appears unlikely that phospholipid asymmetry, if lost at the very beginning of the process, could be restored later, because of the depletion of energetic resources occurring in the late stages of apoptosis (Bevers et al. 1996). It is more conceivable that PS outward flipping does not concern the overall population, as already reported in other cell lines (Frey 1997). Another possibility is that membrane changes have occurred that hinder annexin V binding. Compared with control cultures (not shown), a marked decrease in cell number is detected in treated populations, pointing to an early detachment of many elements in the initial phases of the process. Annexin V-FITC-positive cells may therefore represent only a residual portion of those cells that have entered the apoptotic pathway. In any event, because no reports are available about the fate of annexin V-FITC labeling in advanced apoptosis, heterogeneity must be taken into consideration when an analysis based on annexin V-FITC tagging is carried out on previously untested cells.
The same experiment carried out on NIH3T3 cells confirms that at any given time annexin V does not detect the entire cell population proceeding through different steps of the apoptotic pathway. Only cells undergoing important morphological changes in the adhesion surface are annexin-tagged. In contrast, cells progressing through apoptosis but maintaining a “flat” aspect are annexin V-negative, although it is not possible to forecast if these cells will become positive later on. These results indicate that in NIH3T3 cells PS exposure is prevented if cell shape is substantially preserved. Chen et al. (1997) have recently shown that cell shape, regardless of the type of the matrix protein or the integrin mediating adhesion, governs whether a cell grows or becomes apoptotic. Because this represents an identification mark for macrophages (Wyllie et al. 1980), the exposure of PS might become relevant in adherent cells only after a loss in shape control or detachment from substrate.
However, cytoskeletal changes and PS exposure are independent although strictly coupled (van Engeland et al. 1997). Indeed, in many physiological events, such as mitosis, dramatic changes of cytoarchitecture occur without loss of phospholipid asymmetry. Moreover, temporary cell detachment does not entail immediate annexin V-FITC labeling, although calcein shows that apoptosis has been triggered and is proceeding (Figure 4). Interestingly, the harvesting of these apoptotic NIH3T3 cells entails gross membrane damage to some elements, which results in propidium iodide positivity, calcein negativity, and PS labeling by an-nexin V-FITC on the cytoplasmic side of the membrane, results consistent with those obtained in different cells (Pellicciari et al. 1997). At the same time, in all the other cells chromatin condensation, as revealed by calcein, has already occurred whereas PS exposure has not yet appeared. Therefore, at least in NIH3T3 cells, annexin V-FITC can not be regarded as a reliable early marker for apoptosis.
In summary, our results show that identification of apoptosis in adherent cells by annexin V-FITC can introduce important biases whose entity and degree depend on the cell type under examination and on the time elapsed after stimulus application. In contrast, calcein allows checking of almost all of the pertinent morphological indexes (i.e., cell shrinkage, chromatin condensation, nuclear fragmentation, blebbing, preservation of membrane integrity) and therefore is not affected by the asynchronous progression of the population throughout apoptosis.
Footnotes
Acknowledgements
Supported by CNR target project “Biotechnology.” The confocal apparatus is a facility of the Centro Interfacoltà Misure of the University of Parma.