
Abstract
Increased epidermal growth factor receptor (EGF-R) gene expression and functional protein levels correlate with the metastatic potential of human colon carcinoma (HCC) cells in nude mice. The purpose of this study was to determine whether the production of liver metastases by HCC cells depends on the EGF-R activation status and whether different organ microenvironments influence this activation. Using two independent monoclonal antibodies specific for the activated (i.e., tyrosine-phosphorylated) EGF-R, increased immunoreactivity was observed in HCC cells growing as metastatic lesions in the livers of athymic nude mice. Staining was observed throughout these lesions, both peripherally and centrally. In contrast, little or no immunoreactivity for activated EGF-R was observed in primary tumors growing orthotopically in the cecum or ectopically in the subcutis of nude mice. Immunohistochemistry for total EGF-R levels (irrelevant of activation status) demonstrated similar levels of immunoreactivity in HCC tumors growing in the cecum, subcutis, or liver of nude mice, indicating that total EGF-R levels are not altered after growth in these different microenvironments. Controls included immunohistochemistry for total and activated EGF-R levels in HCC cells growing in vitro under serum-free or EGF-stimulated conditions and A431-epidermoid carcinoma growing in nude mice. Western blot analyses confirmed the specificity of the antibodies for the activated EGF-R. These results suggest that the production of liver metastasis by HCC cells depends in part on the response of tumor cells to organ-derived growth factors and hence the activation of specific cell surface tyrosine kinase receptors.
S
As Paget (1889) proposed and as our recent biological and molecular evidence demonstrates, the organ microenvironment (the “soil”) influences the implantation, invasion, survival, and growth of particular metastatic tumor cells (the “seeds”). Using a human colon cancer model system, Giavazzi et al. (1986) demonstrated that the formation of metastatic lesions in the livers of nude mice was determined not by the ability of the HCC cells to reach the liver but by the survival and proliferative capacity of the metastatic cells in the liver parenchyma. These data suggest that the differential expression of certain growth factor receptors by HCC cells and their response to liver-derived paracrine growth factors represent one potential mechanism by which distinct metastatic colon cancer cells extravasate, survive, and proliferate in the liver parenchyma. Recent studies from our laboratory support this premise (Radinsky 1993,1995; Radinsky et al. 1995b; Singh et al. 1997). Indeed, highly metastatic HCC cells from surgical specimens of Dukes' Stage D tumors or liver metastases responded to mitogens associated with liver regeneration induced by hepatectomy in athymic nude mice (Gutman et al. 1994). Common to the regulation of both liver regeneration and proliferation of normal colon epithelial cells is transforming growth factor-α (TGF-α) (Mead and Fausto et al. 1989; Grupposo et al. 1990; Markowitz et al. 1990; Michalopoulos and DeFrances 1997). TGF-α exerts its effects through interaction with the 170-kD transmembrane glycoprotein EGF-R, which contains a tyrosine kinase (TK) domain in the cytoplasmic region of the protein. A series of rapid responses are stimulated by the binding of TGF-α to the EGF-R, including dimerization and activation of the EGF-R TK, resulting in autophosphorylation of tyrosine residues within the EGF-R itself and within a number of other proteins participating in the signal transduction cascade (Carpenter 1987). After 10–12 hr of continuous stimulation by EGF or TGF-α, cells are committed to synthesize DNA and to divide.
The purpose of this study was to determine whether the production of liver metastases by HCC cells depends on the EGF-R activation status and whether different organ microenvironments influence this activation. Using two independent mouse monoclonal antibodies specific for activated (i.e., tyrosine-phosphorylated) EGF-R, we immunohistochemically analyzed primary and metastatic tumors resulting from implantation of highly metastatic HCC cells into the cecum, subcutis, spleen, and livers of nude mice. Our results demonstrate increased immunoreactivity in HCC metastatic lesions in the livers but not in HCC primary tumors growing in the cecum or subcutis of athymic nude mice. These findings suggest that the production of liver metastasis by HCC cells depends, in part, on the response of tumor cells to organ-derived growth factors and hence the activation of specific cell surface tyrosine kinase receptors.
Materials and Methods
Cells and In Vitro Culture Conditions
A431 human epidermoid carcinoma cells were obtained from the American Type Culture Collection (Rockville, MD). The human colon cancer line KM12C was established in culture from a Dukes' Stage B2 surgical specimen (Morikawa et al. 1988a,b). KM12C cells were injected into the cecum of nude mice and liver metastases were isolated, established in culture, and designated as KM12SM. The metastatic potential of the KM12SM cell line has been previously established (Morikawa et al. 1988a,b; Radinsky et al. 1995). All tumor cell lines were maintained on plastic in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), sodium pyruvate, nonessential amino acids,
Animals and Production of Tumors
Male athymic nude mice were obtained from the animal production area of the NCI-Frederick Cancer Research Facility (Frederick, MD). The mice were maintained under specific pathogen-free conditions and were used for experiments at 8 weeks of age. Animals were maintained in facilities approved by the American Association for Accreditation of Laboratory Animal Care and in accordance with current regulations and standards of the US Department of Agriculture, Department of Health and Human Services, and National Institutes of Health.
To produce tumors, tumor cells were harvested from sub-confluent cultures by 2–3-min treatment with 0.25% trypsin and 0.02% EDTA. Trypsinization was stopped with medium containing 10% FBS and the cells were washed once in serum-free medium and resuspended in Hanks' balanced salt solution (HBSS). Only single-cell suspensions (1 × 106 cells/ 0.05 ml of HBSS) with greater than 90% viability were used for injections into the cecum, spleen, or subcutis of nude mice (Morikawa 1988a,b; Singh et al. 1997). The mice were sacrificed 4–10 weeks thereafter and the size of the primary tumors and the number of liver nodules were determined. Histopathology studies confirmed the nature of the disease.
Western Blot Analyses
Tumor cells were washed and scraped into PBS (0.15 M NaCl, 2.7 mM NaHPO4 7H2O), 1.5 mM KH2PO4, pH 7.3) containing 5 mM EDTA and 1 mM Na-orthovanadate and centrifuged, and the pellet resuspended in lysis buffer [20 mM Tris-HCl, pH 8.0, 137 mM NaCl, 10% glycerol (v/v), 2 mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 20 μM leupeptin, 0.15 U/ml aprotinin], sonicated, and centrifuged to recover insoluble protein. Lysis buffer with 1% Triton X-100 (v/v) was added to the pellet, and Triton X-100-soluble protein was separated by centrifugation and diluted in sample buffer (62.5 mM Tris-HCl, pH 6.8, 2.3% NaDodSO4 (w/v), 100 mM dithiothreitol, and 0.005% bromphenol blue) and boiled. The proteins (20 μg/lane) were resolved on 7.5% NaDodSO4-PAGE and transferred onto 0.45-μm nitrocellulose membranes. The filters were blocked with 3% bovine serum albumin (w/v) in TBS (20 mM Tris-HCl, pH 7.5, 150 mM NaCl), probed with either polyclonal sheep anti-human EGF-R (1:1000) (Upstate Biotechnology; Lake Placid, NY), monoclonal anti-activated EGF-R (1:1000) (Chemicon International; Temecula, CA), or monoclonal anti-phosphotyrosine (MAb 4G10) (1:2000) (Upstate Biotechnology) in TTBS [0.1% Tween 20 (v/v) in TBS], incubated with horseradish peroxidase-conjugated donkey anti-sheep IgG (1:2000) (Sigma Immunochemicals; St Louis, MO) or sheep anti-mouse IgG (1:2000), respectively, in TTBS. The blots were re-probed with rabbit anti-α-actin (1:100) in TTBS (Sigma Immunochemicals) followed by donkey anti-rabbit IgG (1:2000) in TTBS (Amersham; Arlington Heights, IL). Protein bands were visualized by the ECL detection system (Amersham).
Immunohistochemical Analyses
A431 epidermoid carcinoma and KM12SM HCC cells were grown in culture to 60% confluence on sterile slides, starved overnight in 0% serum-containing medium, treated with 50 ng/ml EGF for 20 minutes, and fixed in 4% paraformaldehyde/PBS (v/v) for 10 min. The slides were treated for 12 min with 3% hydrogen peroxide in methanol (v/v) and then incubated in protein blocking solution [5% normal human serum/0.5% normal goat serum in PBS (v/v)] for 15 min. The slides were then incubated overnight at 4C overnight in a humidified chamber with either mouse MAb anti-EGF-R (1:50) (Biogenex; San Ramon, CA), which detects total EGF-R proteins, or mouse anti-activated MAb EGF-R (1:50) (Zymed Laboratories; San Francisco, CA) which detects only the activated (tyrosine-phosphorylated form) of EGF-R. It does not crossreact with other phosphorylated proteins or with inactivated EGF-R (Campos-Gonzalez and Glenney 1991; Sorkin et al. 1992; Duello et al. 1994; Dinney et al. 1997). The samples were then rinsed three times with PBS, incubated in protein-blocking solution for 10 min, and then incubated for 60 min with anti-mouse peroxidase-conjugated secondary antibody (1:200) (Boehringer Mannheim; Indianapolis, IN) in protein-blocking solution and treated with the chromagen substrate 3,3′diaminobenzidine (Research Genetics; Huntsville, AL), rinsed, and mounted in Permount before photography.
At necropsy, tumor tissue from KM12SM HCC growing in the cecum, subcutis, and liver of nude mice, or from A431 epidermoid carcinoma growing in the subcutis of athymic mice, was cut into 5-mm pieces, placed in OCT compound (Miles Laboratories; Elkhart, IN) in 1-inch aluminum caps, and snap-frozen in liquid nitrogen. Frozen sections (8–10 mm) were picked up on silane-treated slides (Fisher Scientific; Pittsburgh, PA) and air-dried for 30 min. The slides were fixed in cold acetone for 10 min, treated for 12 min with 3% hydrogen peroxide in methanol (v/v), and then incubated in protein blocking solution [5% normal human serum/0.5% normal goat serum in PBS (v/v)] for 15 min. Three different primary antibodies were used for analysis of tissue sections: (a) a mouse monoclonal anti-EGF-R antibody (1:50) (Biogenex) that reacts with total EGF-R protein levels and does not distinguish between activated and inactivated EGF-R; (b and c) two independent mouse monoclonal anti-activated EGF-R antibodies (1:50) (Chemicon International or Zymed) that react specifically with the activated and phosphorylated human EGF-R and do not react with other phosphorylated proteins (Campos-Gonzalez and Glenney 1991; Sorkin et al. 1992; Duello et al. 1994; Dinney et al. 1997). The sections were incubated with the primary antibody in a humidified chamber for 15–18 hr at 4C, rinsed three times with PBS, and incubated in protein-blocking solution for 10 min. Sections were then incubated for 60 min with anti-mouse peroxidase-conjugated secondary antibody [1:200 (v/v)] for 1 hr at ambient temperature. Positive reaction was visualized by incubating the slides with stable 3,3′-diaminobenzidine for 10–20 min. The sections were rinsed with distilled water and counterstained with Gill's hematoxylin for 1 min. The sections were dried, mounted in Permount, examined in a brightfield microscope, and images digitized using a Sony 3CCD color video camera (Sony of America; Montvale, NJ) and a personal computer equipped with Optimas Image Analysis Software (Optimas; Bothell, WA). Composite images were printed on a Sony dye-sublimation printer.
Results
Western Analyses of Total EGF-R and Activated EGF-R in A431 Carcinoma Cells Growing in Culture
A431 epidermoid carcinoma cells express high levels of EGF-R (approximately 2 × 106 receptors per cell) and produce TGF-α in an autocrine manner, which results in activation of the EGF-R TK and hence auto-phosphorylation (Ullrich et al. 1984). Western blot analysis of A431 cell lysates demonstrated high levels of total EGF-R and activated EGF-R under different culture conditions (Figure 1). A431 cells growing in vitro under serum-free conditions or in the presence of EGF for 15 min exhibited high levels of activated-EGF-R with MAbs specific for either phosphotyrosine or activated human EGF-R (Figure 1, Lanes A and C). Importantly, anti-activated EGF-R antisera showed specificity only for the phosphorylated EGF-R but not for other phosphorylated proteins, as indicated by the lack of lower molecular weight immunoreactive proteins, whereas anti-phosphotyrosine antisera demonstrated strong immunoreactivity to the EGF-R and to many low molecular weight proteins (Figure 1; compare Lanes A and C). Pretreatment of these cells with the TK inhibitor (30 mM) specific for EGF-R for 1 hr, followed by treatment with EGF (15 min), abrogated this effect (Figure 1; compare Lane D with Lanes A-C).
Immunohistochemical Analysis of Total EGF-R and Activated EGF-R in A431 Carcinoma and KM12SM HCC Cells Growing in Culture
The total EGF-R content and level of activated EGF-R (tyrosine-phosphorylated form) was analyzed with immunohistochemistry in KM12SM HCC cells growing in culture under different treatment conditions. Highly metastatic KM12 SM cells treated with EGF demonstrated enhanced immunostaining with the anti-activated EGF-R MAb compared with control untreated cells (Figure 2; compare Figures 2G and 2F). No change in total EGF-R levels was observed under the identical conditions (Figure 2E; data not shown). These results indicate enhanced EGF-R autophosphorylation after treatment with EGF, which is detectable at the single-cell level with this assay. Similarly, analysis of A431 epidermoid carcinoma cells growing in culture showed high levels of immunoreactivity indicative of total EGF-R and activated EGF-R correlating directly with the Western blot analyses (Figures 2A–2C). These data demonstrate the feasibility and specificity of anti-total EGF-R and anti-activated EGF-R immunohistochemical analyses.
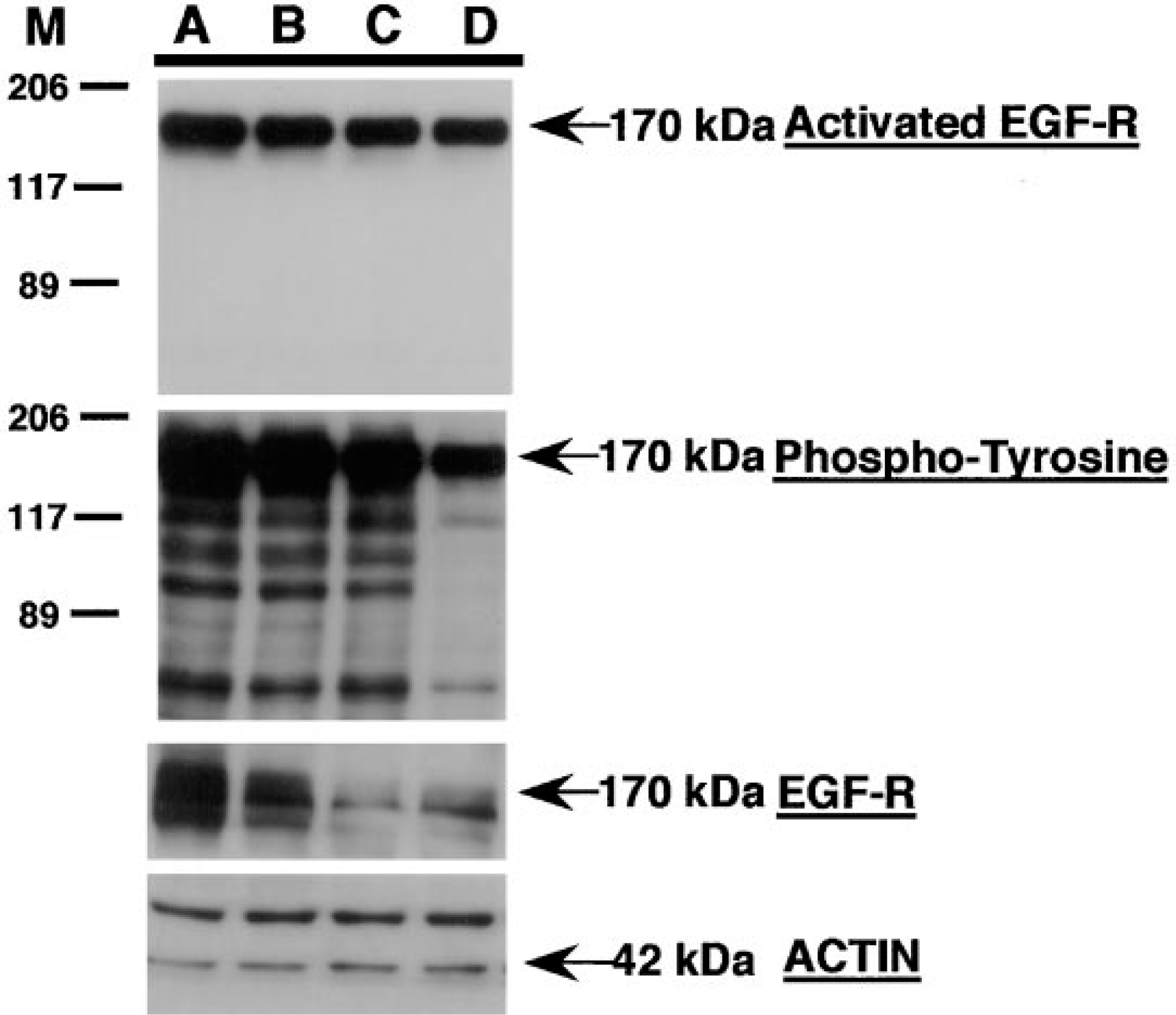
Western blot analysis of A431 epidermoid carcinoma cells for total and activated EGF-R. A431 epidermoid carcinoma cells growing in vitro under serum-free conditions were stimulated for 20 min with EGF in the presence or absence of an EGF-R TK-specific inhibitor (Dinney et al. 1997). Cells were washed, lysed, and soluble proteins in the lysate (30 mg/lane) were separated by 7.5% SDS-PAGE, transferred to nitrocellulose, and sequentially probed with antisera specific for activated EGF-R, phosphotyrosine, total EGF-R, and actin (see Materials and Methods). The immunoreactive protein bands were detected by incubating the blot with the corresponding peroxidase-conjugated IgG and were visualized with the ECL system. A431 cells grown in (
Immunohistochemical Analysis of Total EGF-R and Activated EGF-R in Tumor Sections of KM12SM HCC Cells Growing in the Subcutis, Cecum, and Liver of Athymic Nude Mice
We next determined if different organ microenvironments can influence the activation status of EGF-R in HCC cells growing as tumors in nude mice. Little or no immunoreactivity was detected in KM12 SM tumor sections from subcutaneous or cecal environments with two independent anti-activated EGF-R MAbs (Figures 3F-3G). In contrast, specific immunoreactivity for anti-activated EGF-R was detected at higher levels, both peripherally and centrally, in KM12 SM HCC liver metastases (Figure 3H). Immunoreactivity for total EGF-R was observed in each tumor specimen, with no detectable differences between the different sites, suggesting that the expression of total EGF-R in KM12 SM HCC cells is not regulated by the microenvironment (Figures 3B-3D). Analysis of A431 epidermoid carcinoma grown in the subcutis of athymic nude mice demonstrated high immunoreactivity for both total and activated EGF-R (Figures 3A and 3E). These results indicate increased immunoreactivity for activated EGF-R in HCC liver metastases but not in HCC primary tumors growing in the cecum or subcutis of athymic nude mice. These findings suggest that the production of liver metastasis by HCC cells depends, in part, on the response of tumor cells to organ-derived growth factors (e.g., TGF-α) and hence the activation of specific cell surface tyrosine kinase receptors.
Discussion
Many human tumors and cell lines express increased levels and amplification of EGF-R. These include breast cancer (Sainsbury et al. 1995), gliomas (Liber-man et al. 1985), lung cancer (Harris and Neal 1989), bladder cancer (Neal et al. 1985), tumors of the female genital tract (Gullick et al. 1986), the A431 epidermoid carcinoma (Ullrich et al. 1984), and colon carcinoma (Gross et al. 1991). These findings suggest that the overexpression of EGF-R plays an important role in the growth of certain tumors.
We have previously shown that the expression of EGF-R directly correlates with the ability of human colon carcinoma cells to produce liver metastases (Radinsky et al. 1995). Highly metastatic HCC cells derived from surgical specimens of late stage HCCs (Dukes' Stage D) or liver metastases expressed 10–20-fold higher steady-state mRNA levels for EGF-R compared with low metastatic cell lines derived from early-stage HCCs (Dukes' Stage A or B). Immunohistochemical staining of these highly metastatic KM12SM cells demonstrated uniform and intense staining for total EGF-R. In contrast, low metastatic parental KM12C cells showed heterogenous EGF-R staining, with less than 10% of cells staining positive (Radinsky et al. 1995). Data from our laboratory further suggested that HCC cells expressing increased EGF-R levels have a selective advantage in the production of liver metastases in athymic nude mice (Radinsky et al. 1995; Singh et al. 1997). Collectively, these data support the hypothesis that the formation of metastatic lesions in the livers of nude mice is determined not by the ability of the HCC cells to reach the liver but by the survival and proliferative capacity of the metastatic cells in the liver parenchyma. Hence, the differential expression of certain growth factor receptors by HCC cells and their response to liver-derived paracrine growth factors may be one mechanism by which distinct metastatic colon cancer cells survive and proliferate in the liver parenchyma.
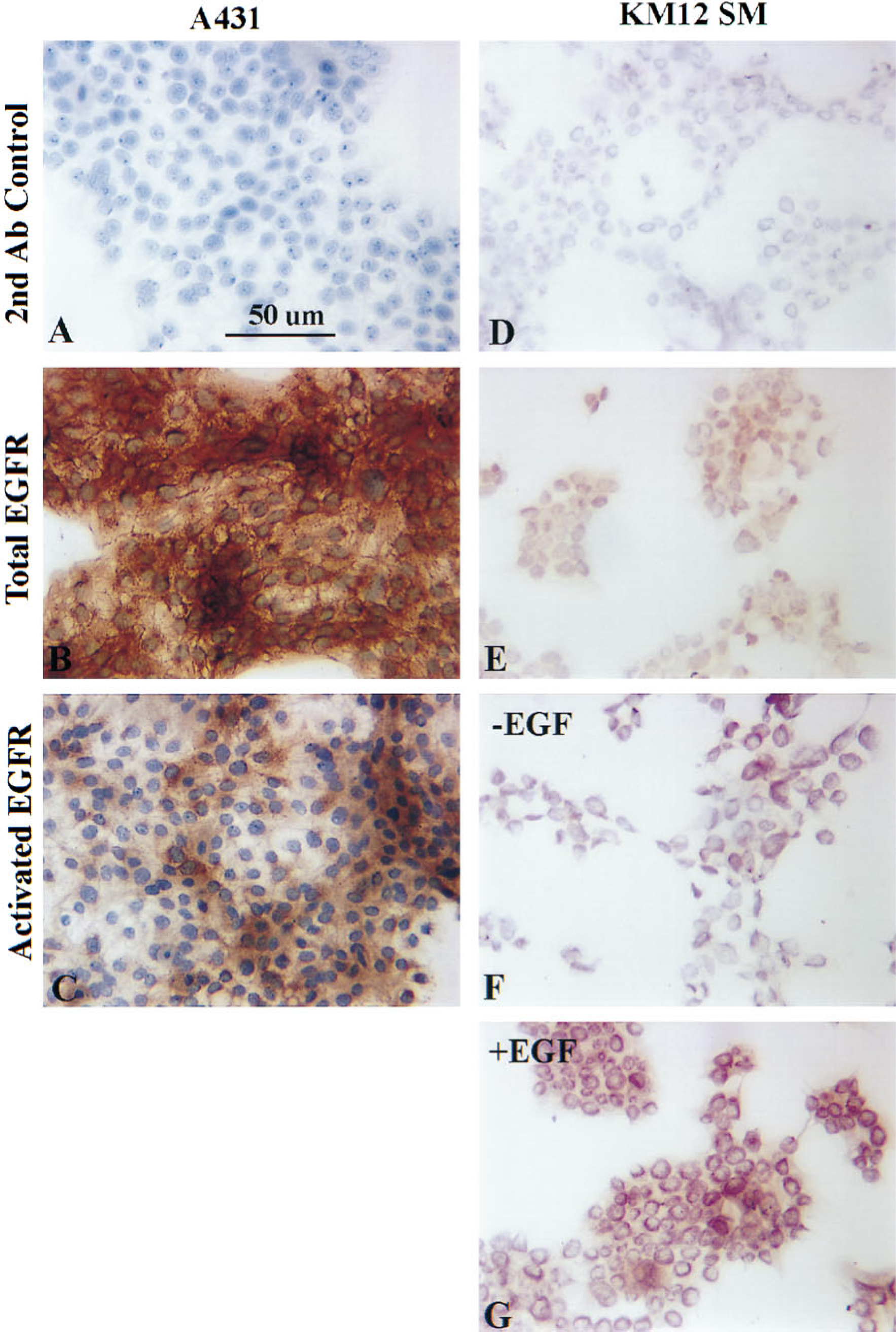
Immunohistochemical staining of A431 epidermoid carcinoma and KM12SM HCC cells growing in culture for total and activated EGF-R. A431 and KM12SM cells growing in culture were serum-starved overnight, treated with 50 ng/ml EGF for 20 min, and fixed in 4% paraformaldehyde/PBS (v/v). The cells were immunolabeled with either anti-EGF-R MAb (1:50) (Biogenex) or anti-activated EGF-R Mab (1:50) (Zymed), followed by incubation with a peroxidase-conjugated secondary antibody (1:200). Samples were treated with the chromagen DAB, rinsed, counterstained with hematoxylin solution Gill No. 3, and mounted in Permount before photography. (
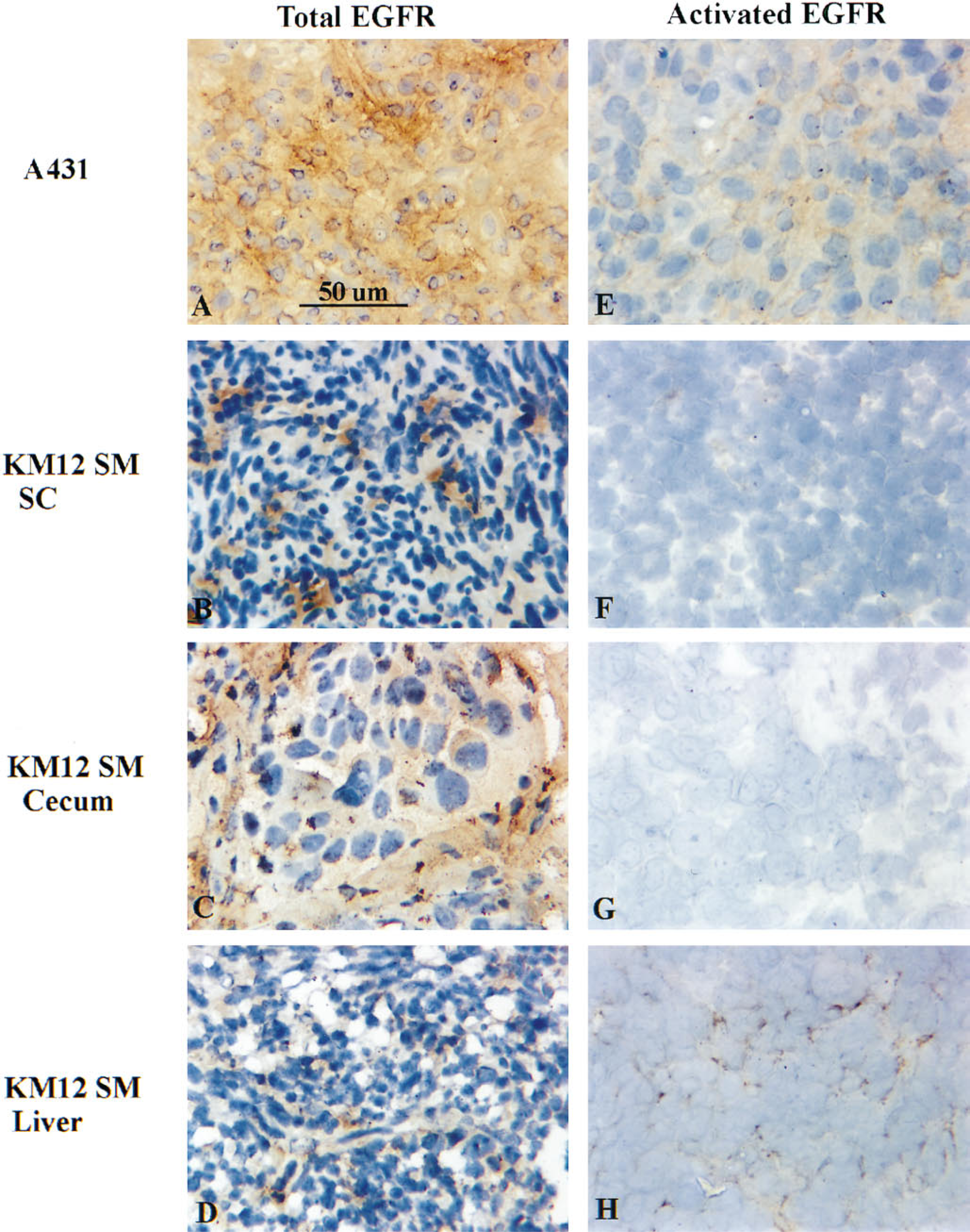
Immunohistochemical analyses of total and activated EGF-R in tumor sections of KM12 SM cells growing in the subcutis, cecum, and livers of nude mice. Frozen tumor sections were immunostained with (
Our present results demonstrate preferential activation of the EGF-R in HCC metastatic lesions in the livers of athymic nude mice, using two independent antibodies to the activated form of EGF-R. Little or no immunoreactivity for the activated EGF-R was observed in tumor sections from the cecal and subcutaneous sites. This finding cannot be attributed to an absence of EGF-R expression on the tumor cells, because positive staining for total EGF-R was demonstrated in serial tumor sections from both the cecal and subcutaneous environments and from liver metastatic lesions. These observations, coupled with the results of Western blotting and immunohistochemical analyses of the cells grown in culture after treatment with EGF, confirmed the specific nature of the immunoreactivity.
These results demonstrate that the production of liver metastases by HCC may depend, in part, on the activation status of EGF-R, and support the hypothesis that organ-specific metastasis of HCC to the liver may be in mediated by the HCC cells' responses to organ-derived growth factors, resulting in the activation of specific cell surface tyrosine kinase receptors. Furthermore, these results point to EGF-R as a potential target for therapeutic intervention in the treatment of HCC and suggest that the activated form of EGF-R may be a more accurate prognostic indicator than total EGF-R. The latter awaits experimental proof from the in-depth analysis of HCC clinical samples. However, recent studies from our laboratory of human bladder carcinoma growing orthotopically in athymic nude mice have demonstrated the potential use of the anti-activated EGF-R antibodies as clinical indicators for in vivo evaluation of the therapeutic activity of different tyrosine kinase inhibitors (Dinney et al. 1997).
Footnotes
Acknowledgements
Supported in part by grant R29-CA67952 (RR) and by Cancer Center Support Core grant CA16672 from the National Cancer Institute, National Institutes of Health, and by the Gustavus and Louise Pfeiffer Research Foundation (RR).