
Abstract
The formation of vascular stroma plays an important role in the pathophysiology of malignancy. We describe the use of in situ hybridization in our laboratory as a tool to study the role of vascular permeability factor/vascular endothelial growth factor in the angiogenesis associated with malignancy.
T
Vascular permeability factor (VPF), also known as vascular endothelial growth factor (VEGF), is a multifunctional angiogenic cytokine expressed by many tumors (reviewed in Dvorak et al. 1995). VPF/VEGF binds to endothelial cells by at least two specific cell surface tyrosine kinase receptors, flt-1 and KDR (de Vries et al. 1992; Terman et al. 1992). VPF/VEGF induces microvascular hyperpermeability with a strength some 50,000 times that of histamine, leading to extravasation of plasma proteins and consequent proangiogenic stromal changes (Dvorak et al. 1979; Senger et al. 1983). VPF/VEGF is also an endothelial cell mitogen (Ferrara and Henzel 1989; Gospodarowicz et al. 1989; Keck et al. 1989; Leung et al. 1989) and alters the synthetic profile of endothelial cells, inducing expression of proteins such as collagenase and plasmino-gen activators, which may play a role in angiogenesis (Pepper et al. 1991; Unemori et al. 1992). Angiogene-sis and tumor growth can be suppressed by inhibition of VPF/VEGF or its receptors in certain experimental models (Kim et al. 1993; Millauer et al. 1994). In situ hybridization studies have been important in defining the role of VPF/VEGF in the formation of vascular stroma in malignancy.
Materials and Methods
In Situ Hybridization
In situ hybridization (ISH) was performed on 6-μm frozen sections of tissues that had been fixed in 4% paraformaldehyde in PBS, pH 7.4, for 2–4 hr at 4C and then transferred to 30% sucrose in PBS overnight at 4C and frozen in OCT compound (Miles Diagnostics; Elkhart, IN) and stored at −70C or, alternatively, on 4-μm-thick sections of archival formalin-fixed, paraffin-embedded tissue.
Briefly, slides with paraffin sections were passed through xylene and graded alcohols; 0.2 M HCl; 10 mM Tris/1 mM EDTA with 3 μg/ml proteinase K; 0.2% glycine; 4% paraformaldehyde in PBS, pH 7.4; 0.1 M triethanolamine containing 1/200 (v/v) acetic anhydride; and 2 X SSC. Slides were hybridized overnight at 50C with 500,000 counts of gel-purified
Anti-sense single-stranded 35S-labeled VPF/VEGF RNA probe and its sense control were designed by Brygida Berse and have been described previously (Brown et al. 1992). The anti-sense probe hybridizes specifically with a region of VPF/ VEGF mRNA common to all known VPF/VEGF splicing variants. 35S-Labeled single-stranded anti-sense and sense RNA probes for the VPF/VEGF receptors flt 1 and KDR were designed by Robert W. Jackman and have been described previously (Brown et al. 1993a,b).
Immunohistochemistry
Preparation of an affinity-purified rabbit antibody to the N-terminal 26 amino-acid peptide of human VPF/VEGF has been previously described (Dvorak et al. 1991). The antibody bound VPF/VEGF in ELISA assays and on immunoblots (Sioussat et al. 1993). Immunohistochemistry (IH) was performed on 5-μm frozen sections of the same blocks used for ISH, using an avidin-biotin-peroxidase conjugate protocol. Normal rabbit IgG diluted to an equivalent protein concentration was used as a control in place of the primary antibody.
Staining for factor VIII-related antigen utilized a rabbit polyclonal antibody (Dako; Santa Barbara, CA) with an avidin-biotin-peroxidase conjugate protocol.
Results and Discussion
VPF/VEGF Expression in Invasive Carcinomas
We first used in situ hybridization studies to study expression of VPF/VEGF in an animal carcinoma model (Dvorak et al. 1991). Whereas VPF/VEGF mRNA was detected only in tumor cells, immunoreactive VPF/VEGF protein could be detected in both tumor cells and blood vessels. These data suggested that VPF/VEGF was being synthesized by tumor cells and bound by endothelial cells. VPF/VEGF has been shown to bind to endothelial cells by at least two specific cell surface tyrosine kinase receptors, flt 1 and KDR (de Vries et al. 1992; Terman et al. 1992).
The first human tumor we studied for the expression of VPF/VEGF, flt-1, and KDR mRNAs was the highly vascular renal cell carcinoma (Brown et al. 1993a). VPF/VEGF mRNA was strongly expressed by the malignant epithelial cells of renal cell carcinoma (Figures 1A and 1B). No labeling was seen with control sense probes, and background was low (Figures 1C and 1D). Expression of mRNAs of the VPF/VEGF receptors flt-1 and KDR was strong in endothelial cells in the small vessels adjacent to carcinoma (Figures 1E and 1F). Immunohistochemical staining with anti-VPF/VEGF antibodies revealed staining of both tumor cells and blood vessels (Figure 2A). This provided additional support for a model in which VPF/VEGF was expressed by tumor cells and bound to VPF/VEGF receptors expressed on endothelial cells. Strong expression of VPF/VEGF was also seen in the malignant epithelium of invasive high-grade transitional cell carcinomas of the bladder in this study.
In an effort to extend these findings to other common human malignancies, we studied adenocarcinomas of the colon, stomach, small intestine, and pancreas (Brown et al. 1993b). The patterns of expression were the same as seen in renal cell carcinomas, i.e., tumor cells strongly expressed VPF/VEGF mRNA and endothelial cells in the closely associated vessels in the vascular stroma strongly expressed flt-1 and KDR. Anti-VPF/VEGF antibodies stained both tumor cells and vessels. In later studies, the same patterns of expression of VPF/VEGF and its receptors were noted in invasive ductal carcinoma of the breast (Brown et al. 1995), endometrial adenocarcinoma (Guidi et al. 1996), ovarian carcinoma (Abu-Jawdeh et al. 1996), and invasive squamous cell carcinoma of the uterine cervix (Guidi et al. 1995).
In situ hybridization with radioactive probes can be evaluated in a semiquantitative manner by counting the number of grains in the photographic emulsion overlying the cells. For example, if large numbers of grains are counted over malignant cells in invasive carcinoma but few grains are counted over normal epithelium, then expression is strong in the tumor compared to the normal tissue. Many blocks in our studies were chosen to contain both tumor and normal tissue, thereby allowing direct comparison of mRNA expression in the same section. In almost all cases we have studied, expression of VPF/VEGF and its receptors has been far stronger in carcinomas than in adjacent normal tissue.
In summary, VPF/VEGF and its receptors are strongly expressed in many common invasive human carcinomas, and probably play an important role in the formation of vascular stroma in these malignancies. This makes VPF/VEGF an especially attractive target for antiangiogenic therapy, because such therapy could be effective across a broad range of common human carcinomas.
VPF/VEGF Expression in Dysplasia and Carcinoma In Situ
In situ hybridization is an even more critical tool in the study of the early steps in neoplastic progression. When large invasive carcinomas are resected, a pathologist can often select a piece of tumor that is not important for diagnostic purposes to be optimally prepared for molecular studies such as Northern analysis. However, small biopsy specimens must be routinely processed through paraffin in their entirety for diagnosis. It is in these biopsies that the precursor lesions of invasive carcinoma, i.e., dysplasia and in situ carcinoma, can be found.
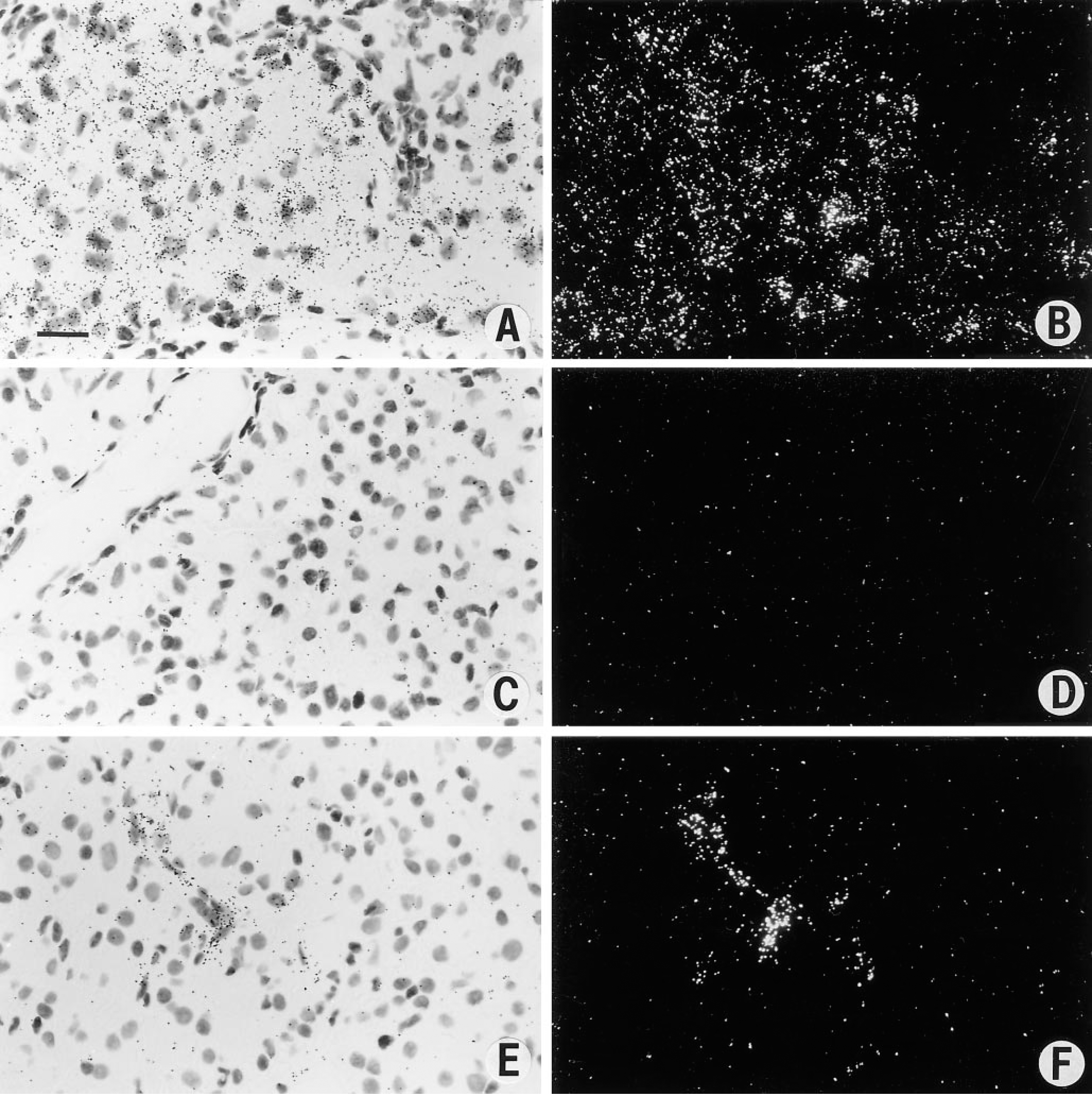
In situ hybridization studies of renal cell carcinoma. (
In situ hybridization can be performed on archival paraffin-embedded material, although in our experience with less sensitivity than when used on fixed frozen sections. In our hands, in situ hybridization is optimally performed on tissue immersed in freshly made 4% paraformaldehyde immediately after surgical removal, fixed for 2–4 hr at 4C, cryoprotected with sucrose, and embedded for frozen sectioning, with all steps using solutions that are ribonuclease-free. In contrast, in specimens undergoing routine histological processing for paraffin sections, the time before a tissue is placed into fixative, the time the tissue is in fixative, and exposure to RNA-degrading enzymes have not been controlled. In situ hybridization may therefore underestimate mRNA expression in archival paraffin-embedded tissues, but meaningful data can still be derived.
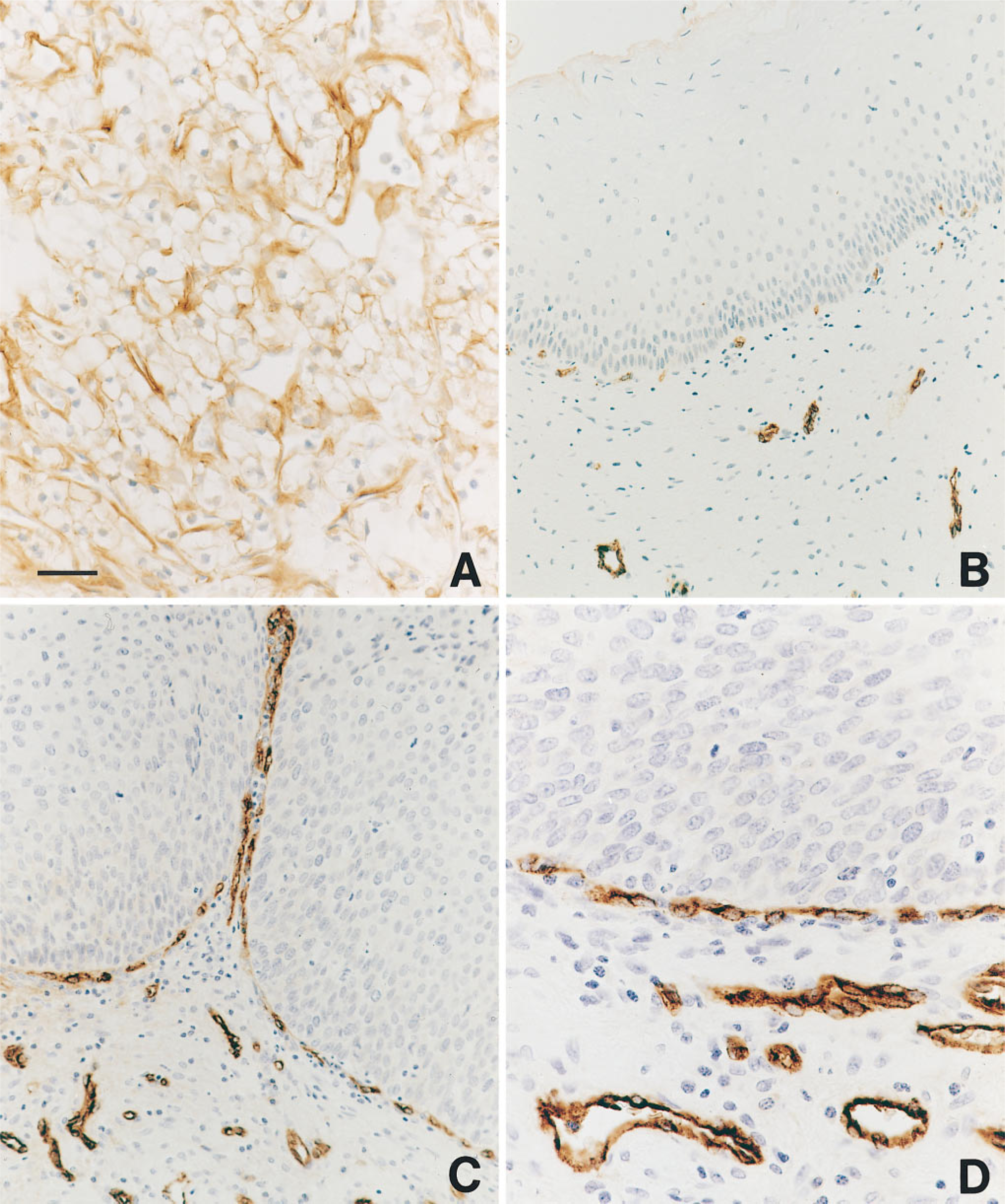
Immunohistochemical staining. (
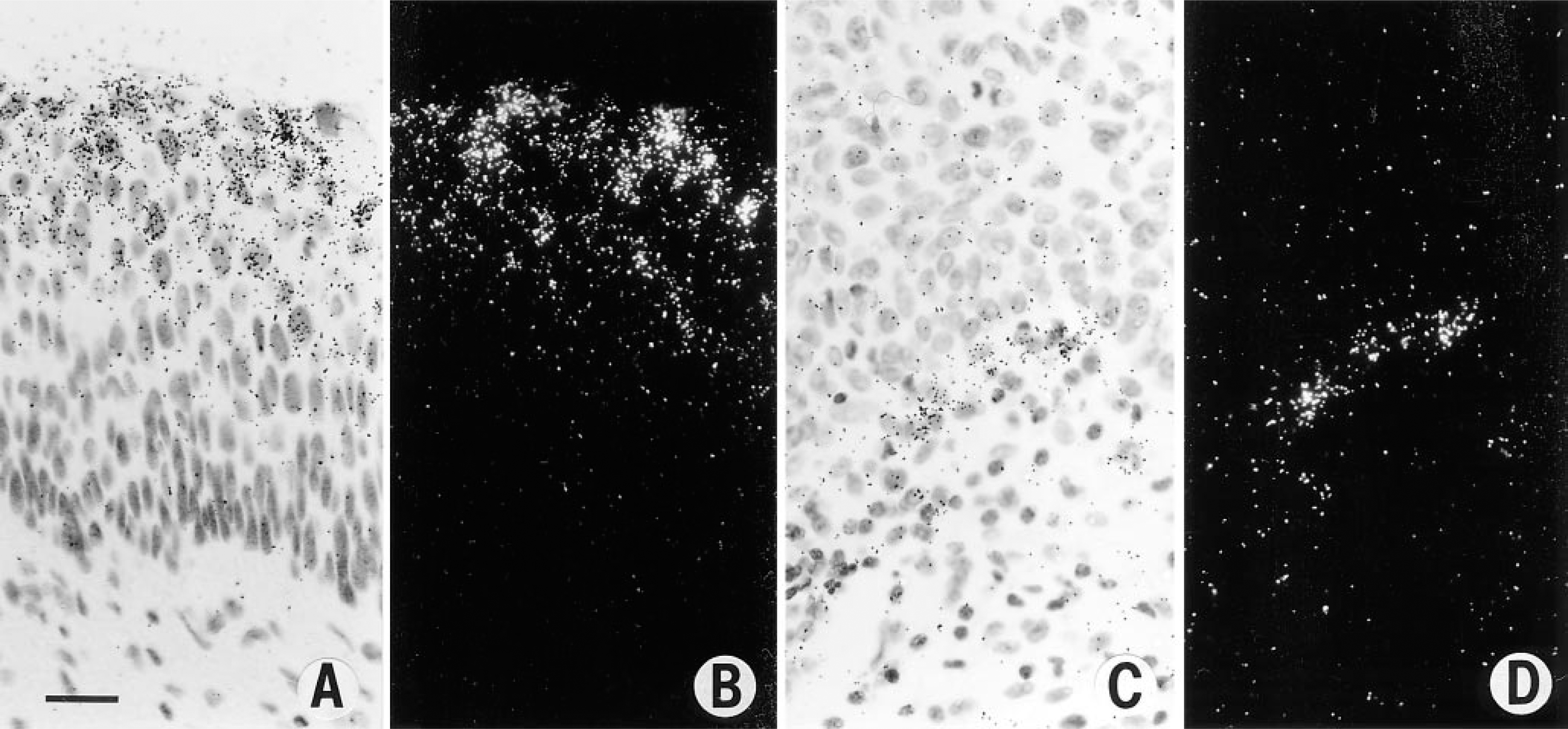
In situ hybridization studies. (
In a study of archival paraffin-embedded biopsies of the uterine cervix (Guidi et al. 1995), strong expression of VPF/VEGF mRNA was seen not only in invasive squamous cell carcinoma but also in high grade squamous dysplasia (Figures 3A and 3B). Therefore, expression of VPF/VEGF was upregulated before tumor invasion in the cervix. Furthermore, the strong expression of VPF/VEGF mRNA in high-grade dysplasia was associated with increased numbers of microvessels in the underlying stroma, which in some cases formed a nearly continuous rim at the junction of epithelium and stroma (Figures 2B–2D), and the endothelial cells on these vessels strongly expressed the VPF/VEGF receptors flt-1 and KDR (Figures 3C and 3D). Therefore, angiogenesis is initiated before tumor invasion in the cervix, and VPF/VEGF may play an important role in this process.
Similar findings were seen in ductal carcinoma in situ (DCIS) of the breast (Brown et al. 1995). Strong expression of VPF/VEGF mRNA was seen in the epithelial cells of DCIS. The number of microvessels has been reported to be increased in DCIS, and rimming of microvessels around DCIS, similar to the pattern seen in cervical dysplasia, has also been described (Guidi et al. 1994). Furthermore, the endothelial cells of these vessels strongly expressed mRNA of the VPF/ VEGF receptors flt-1 and KDR.
Therefore, at least in the cervix and breast, strong expression of VPF/VEGF, flt-1, and KDR begins before tumor invasion and coincides with angiogenesis. When these tumors do invade, they may be invading a highly vascular stroma that they have induced rather than “normal” host stroma. Therefore, the formation of vascular stroma may also play a role in tumor invasion. Such findings by in situ hybridization provide insights into the process of neoplastic progression which would be difficult to obtain by other means.
VPF/VEGF Expression in Metastatic Carcinoma
In situ hybridization studies have also provided intriguing information on tumor metastases. For example, in foci of metastatic breast cancer, tumor cells strongly express VPF/VEGF (Figures 4A and 4B), and there is associated vascular stroma formation with strong endothelial cell expression of VPF receptors (Figures 4C and 4D). Thus, the formation of vascular stroma also occurs at sites of metastasis and VPF may play an important role in the process. Therefore, the tumor is not growing in the native tissue of the metastatic site but rather in a vascular stroma it has induced. The ability to form a vascular stroma may be important to the survival and growth of tumor cells at sites of metastases.
Regulation of VPF/VEGF Expression
In addition to identifying the specific cell types responsible for synthesis of VPF or its receptors in tumors, in situ hybridization studies have also provided clues to factors which may regulate VPF expression. VPF/VEGF expression in glioblastoma was noted to be strongest near areas of necrosis, and in vitro experiments in the same study confirmed hypoxia as an important factor that can regulate VPF expression (Shweiki et al. 1992). We have seen similar strong expression of VPF/VEGF expression adjacent to areas of necrosis in all of the carcinomas we have studied. Careful observation of the patterns of mRNA expression can also provide clues to factors that may regulate expression.
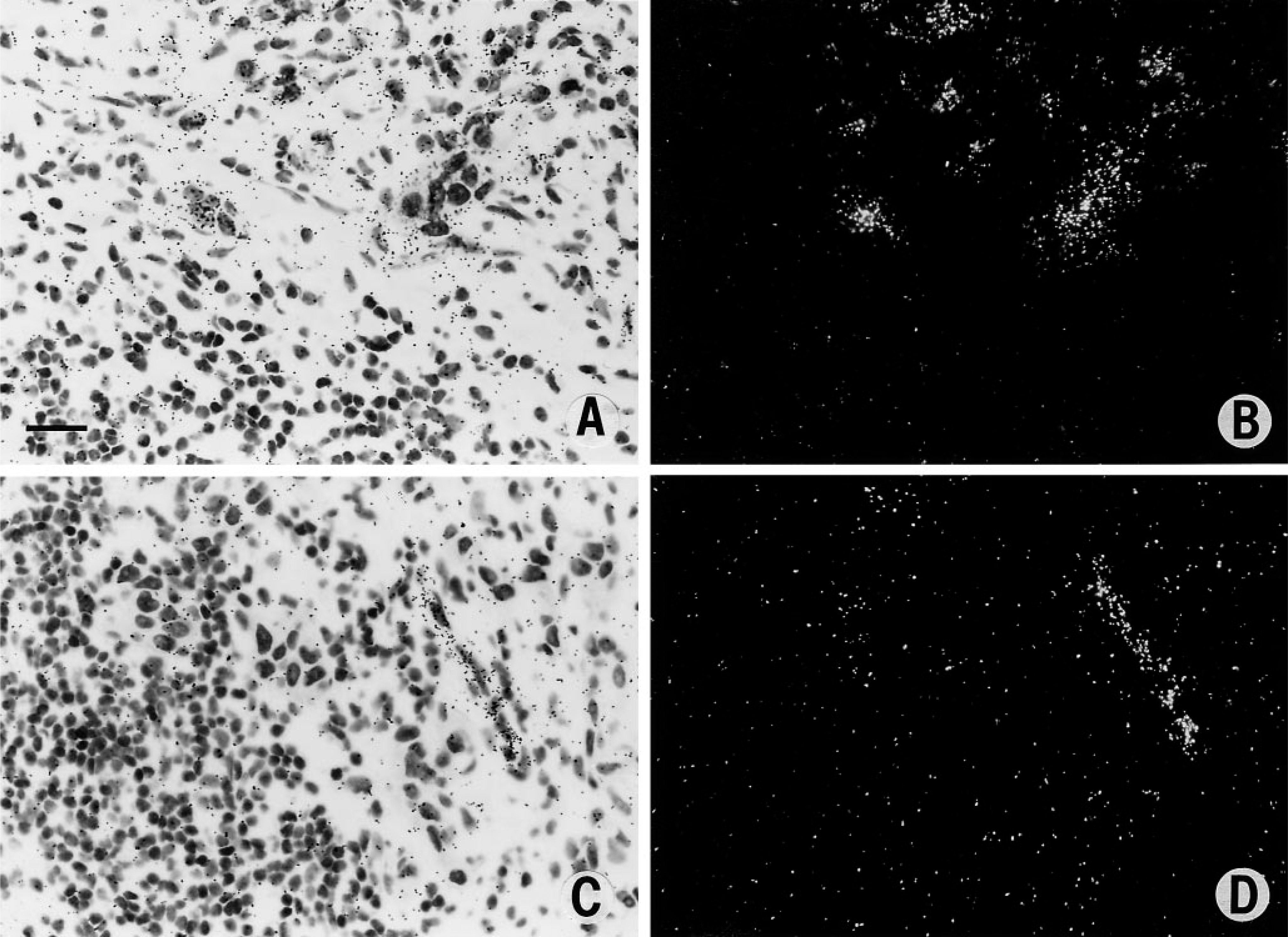
In situ hybridization studies. (
Conclusions
In situ hybridization studies have helped to establish VPF/VEGF as an important angiogenic factor in many common human carcinomas. VPF/VEGF may play an important role not only in invasive tumors but also in preinvasive lesions and in metastases.