
Abstract
Ca2+/calmodulin-dependent protein kinases I and II, initially identified in brain on the basis of their ability to phosphorylate synapsin I, have been implicated in the regulation of Ca2+-dependent synaptic neurosecretion. Specific recombinant and synthetic peptide antibodies were used to examine the distribution of CaM kinases I and II in the rat pancreas and other tissues. The CaM kinase I antibodies detected a doublet of cytosolic proteins of ∼38 and ∼42 kD by immunoblot. CaM kinase I was observed in glucagon-containing A-cells at the periphery of the islet of Langerhans but had little or no overlap with pancreatic polypeptide or somatostatin cells. In contrast, CaM kinase II was localized to somatostatin-containing D-cells. CaM kinase I co-localized with glucagon secretory granules. CaM kinase II was not associated with the somatostatin granule but rather was enriched in areas of the cells that contained relatively little somatostatin. Because glucagon secretion is Ca2+-dependent, it is attractive to speculate that CaM kinase I may play a regulatory role in glucagon secretion. Glucagon and somatostatin cells both utilize intracellular Ca2+ for signaling. Therefore, specific CaM kinases may act as effectors of Ca2+ in these different cell types.
C
Most of the intracellular effects of calcium are mediated by binding to calmodulin. In turn, the calcium/ calmodulin complex interacts with many classes of effector molecules, including the calcium/calmodulin-dependent protein kinases (CaM kinases), resulting in their activation (Braun and Schulman 1995). Some CaM kinases have single substrates, such as myosin light chain kinase and CaM kinase III, which phosphorylate myosin P light chain and the protein synthesis factor elongation factor 2, respectively. CaM kinases I, II, and IV are multifunctional enzymes, each with a variety of substrates. The best characterized is CaM kinase II; its biochemical properties, such as regulation of neurotransmitter release through phosphorylation of synapsin I on sites 2 and 3 are well understood (Lin et al. 1987; Nichols et al. 1990).
Like CaM kinase II, CaM kinase I catalytic activity and mRNA have been found in all tissues examined; the highest levels of the enzyme are present in brain, adrenals, liver and lung (Nairn and Greengard 1987; Picciotto et al. 1996). Immunological studies have determined that neuronal CaM kinase I is a soluble protein that is present in cell bodies, axons and at the synapse (Picciotto et al. 1995). It is regulated by an autoinhibitory mechanism that is relieved by calmodulin binding (Yokokura et al. 1995; Goldberg et al. 1996); it is also regulated by phosphorylation by a distinct CaM kinase I kinase that is part of a cascade analogous to the MAP kinase system (Lee and Edelman 1994). Full-length CaM kinase I cDNA clones have been isolated from rat brain, rat fetal lung, and human promyelocyte (HL-60) libraries (Picciotto et al. 1993; Cho et al. 1994; Haribabu et al. 1995). Clones for two full-length isoforms, α and β, have recently been isolated from an embryonic rat brain cDNA library (Yokokura et al. 1997). The predicted molecular weights of the encoded proteins correspond to two major immunoreactive polypeptides of approximately 42 and 38 kD which, along with several minor polypeptides, have been observed in rat cortex (Picciotto et al. 1995).
The substrate recognition motif for CaM kinase I, Hyd-Xaa-Arg-Xaa-Xaa-(Ser/Thr)-Xaa-Xaa-Xaa-Hyd (Lee et al. 1994), is only somewhat more restrictive than that of CaM kinase II (Arg-Xaa-Xaa-Ser/Thr) (Czernik et al. 1987). Several in vitro substrates have been identified, including synapsin I, synapsin II, the cAMP response element binding protein (CREB), and the cystic fibrosis transmembrane conductance regulator (CFTR) (Nairn and Greengard 1987; Sheng et al. 1991; Picciotto et al. 1992). However, the wide distribution of this kinase in mammalian cells suggests that additional in vivo substrates are likely to exist.
In this study we compared the distributions of two effectors of calcium signaling, CaM kinases I and II, in gastrointestinal tissues using an immunocytochemical approach. The results indicate that the two enzymes have distinct cellular distributions with little overlap. The greatest differences are observed in the pancreatic islet, in which CaM kinase I immunoreactivity is concentrated in glucagon-containing cells and CaM kinase II immunoreactivity is found exclusively in somatostatin-containing cells. We also compared the localization pattern of CaM kinase I to previously described localization patterns of CFTR (Marino et al. 1991; Ameen et al. 1995) and CREB (Bito et al. 1996), seeking evidence for co-localization that would suggest that either or both are substrates in vivo.
Materials and Methods
Antibodies
CC77 is an affinity-purified rabbit antibody to CaM kinase I and is expressed in E. coli as a glutathione S-transferase fusion protein (Picciotto et al. 1995). It was used for immunoblotting at a concentration of ∼100 pg/ml and for immunofluorescence at ∼2 ng/ml. CC102 is an affinity-purified rabbit antibody to a synthetic peptide corresponding to the carboxyl terminus of CaM kinase I (CVEPGSELPPAPPPSSRAMD) (Cho et al. 1994). It was used for immunoblotting at a concentration of ∼200 pg/ml and for immunofluorescence at ∼4 ng/ml. G301 is an affinity-purified rabbit antibody to a synthetic peptide corresponding to residues 281–302 of the α-subunit of rat brain CaM kinase II (Yamagata et al. 1991). It was used for immunoblotting at a concentration of ∼200 ng/ml and for immunofluorescence at a concentration of ∼3 μg/ml. Antibody CUREPPMoAb#5, raised against rat pancreatic polypeptide, was provided by Dr. John Walsh (CURE/Gastroenteric Biology Center, Antibody/ RIA Core; NIH grant #DK41301) (Wong et al. 1995) and diluted ∼1:3000. Monoclonal anti-glucagon mouse ascites fluid clone K796B10 was purchased from Sigma (St Louis, MO) and diluted 1:10,000. Mouse monoclonal anti-somatostatin was purchased from Biomeda (Foster City, CA) and diluted 1:50.
Immunoblot Analysis
Tissues were cut into 5–10 mm cubes in ice-cold minimal Eagle's Medium (Gibco; Grand Island, NY) and were homogenized with a mechanical homogenizer in the presence of protease inhibitors (Gorelick et al. 1987). A postnuclear supernatant was formed by sedimenting unbroken cells and nuclei for 20 min at 600 X g. This extract was boiled in 1% sodium dodecyl sulfate (SDS) and subjected to SDS-PAGE (Laemmli 1970). Separated proteins were transferred to Immobilon for 2400 volt-h (Gershoni and Palade 1983). Membranes were treated with 5% nonfat dry milk (Carnation; Los Angeles, CA) in 0.1 M NaCl, 0.01 M Tris-HCl, pH 7.4, 0.1% Triton X-100, 0.05% NaN3 to block nonspecific binding, then incubated with CC77 or CC102 for 2 hr at room temperature. Antibody detection used an enhanced chemiluminescence kit (Amersham; Arlington Heights, IL).
Immunofluorescence Localization
Rats were perfused with DMEM, followed by 2% formaldehyde (made from paraformaldehyde) in 0.075 M NaPO4, pH 7.4. Rats were treated in accordance with local institutional guidelines. Tissues were removed, fixation was continued for a total of 1 hr, and the tissues were cryopreserved in 15% sucrose in PBS overnight at 4C, then cryosectioned to approximately 3 μm in thickness. After incubation in 3% goat serum in PBS for 1 hr, then for 2 hr with primary antibody, FITC-goat anti-rabbit immunoglobulin F(ab')2 (1:200) (Biosource; Camarillo, CA) or TRITC-goat-anti-mouse F(ab')2 (1:300) (Biosource) was used to detect the antigen.
Slides were photographed on a Zeiss Axiophot microscope (Thornwood, NY) or a Zeiss Axiovert 10 microscope equipped with a Bio-Rad (Richmond, CA) MRC-600 confocal attachment.
Results
Immunoblot Survey of CaM Kinase I in Non-neuronal Tissues
Rat tissues were homogenized and postnuclear super-natants were subjected to immunoblot analysis using antibodies CC77 and CC102 (Figure 1). In the pancreas, CC77, raised against a fusion protein encoding the entire 42-kD α-isoform, (Cho et al. 1994; Yokokura et al. 1997), recognized ∼42- and ∼38-kD bands of approximately equal intensity, consistent with previous data (Picciotto et al. 1995). Two major immunoreactive bands of approximately 42 and 38 kD were observed in various ratios in all tissues examined except for the parathyroid, in which only an ∼38-kD band was observed. Synthetic peptide antibody CC102, raised against the COOH terminus of the α-isoform, preferentially detected the larger band in all tissues examined. In addition, an ∼38-kD band was also observed in the pancreas, stomach, adrenal and submandibular glands. Interestingly, in the parathyroid gland, this antibody recognized an ∼42-kD band even though CC77 did not. In some of the tissues examined the antibodies recognized other unidentified nonspecific bands. CC77 reacted with minor higher molecular weight bands in stomach and parathyroid. Antibody CC102 detected prominent high molecular weight bands in the stomach and minor bands in the adrenal gland. These observations are consistent with the presence of at least two isoforms of CaM kinase I in various ratios in all of the non-neuronal tissues surveyed.
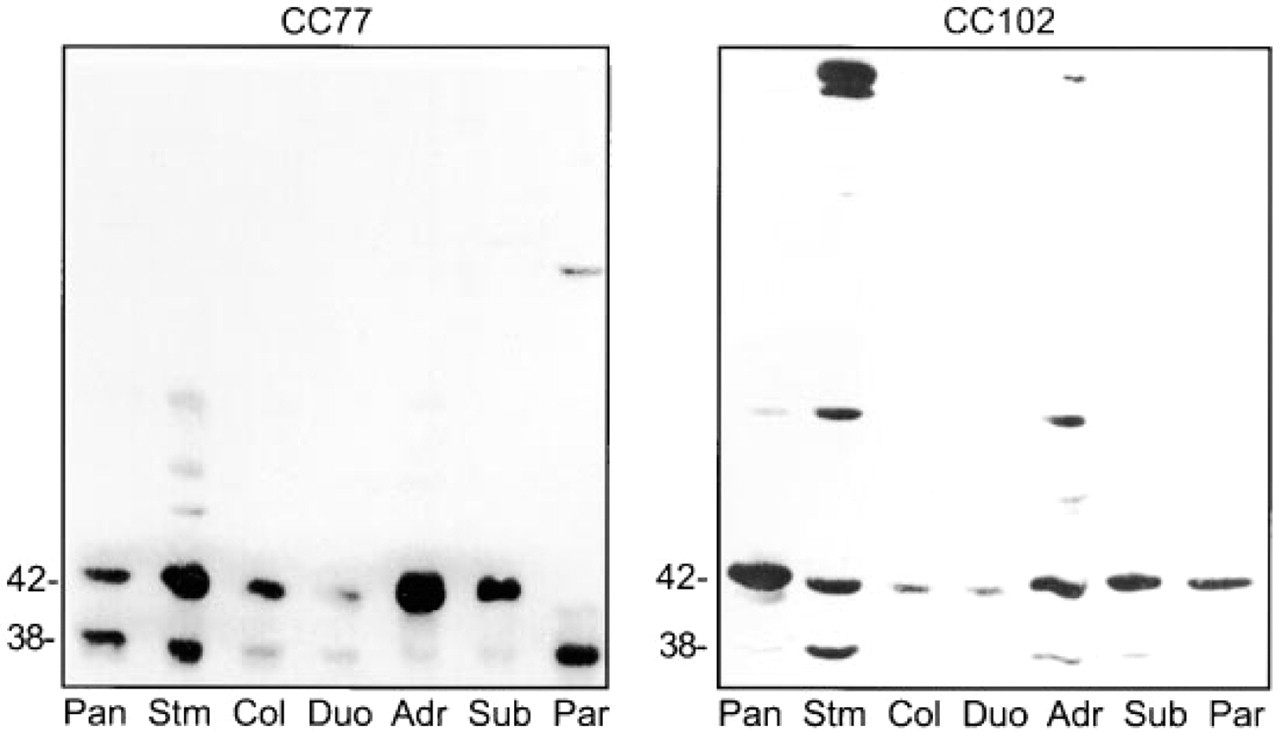
Immunoblot of CaM kinase I in non-neuronal tissues. Rat tissue postnuclear supernatants (55 μg protein/lane) were separated by SDS-PAGE (8% acrylamide) and transferred to Immobilon. The membranes were probed with the affinity-purified antibody CC77 or CC102. The immunoreactive bands were detected by enhanced chemiluminescence. Pan, Pancreas; Stm, stomach; Col, colon; Duo, duodenum; Adr, adrenal; Sub, submandibular gland; and Par, parathyroid.
Immunocytochemical Localization of CaM Kinase I in Non-neuronal Tissues
Immunofluorescence microscopy was used to examine the distribution of CaM kinase I in tissues outside the central nervous system. The choroid plexus has previously been shown to be immunoreactive with CC77 and to have high levels of CaM kinase I (Picciotto et al. 1995). CC77 labeling was present throughout the epithelial cells of the choroid plexus (Figure 2A), consistent with the soluble localization previously described (Picciotto et al. 1995). CC102 staining (Figure 2B) showed a similar pattern but with lesser intensity than CC77. No staining was observed in the absence of primary antibody (Figure 2C).
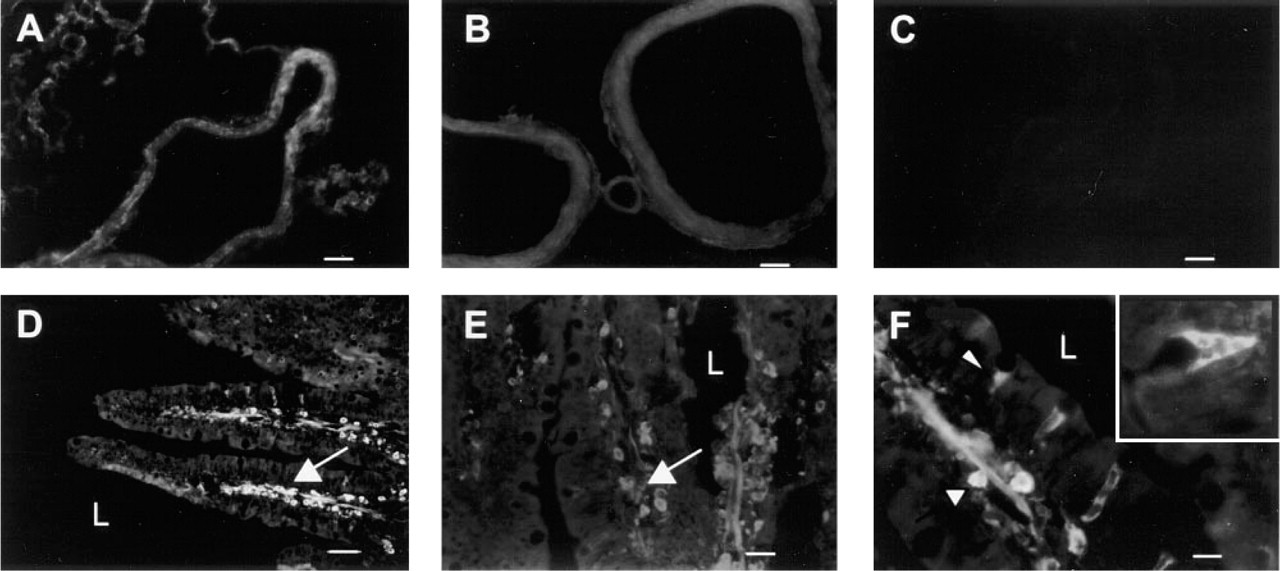
Immunofluorescence localization of CaM kinase I in non-neuronal tissues. Rat tissues were fixed with paraformaldehyde, processed as frozen sections, and labeled using antibody C77 (
In the duodenum, the most prominent staining by antibodies CC77 (Figure 2D) and CC102 (Figures 2E and 2F) was in mononuclear cells of the lamina propria. There was also prominent staining of either capillary endothelium or lacteals at the core of the villus. A distinct population of mucosal cells were labeled by antibodies CC77 (data not shown) and CC102 (Figure 2F). CaM kinase I immunoreactivity was restricted to the basal region of these cells; phase microscopy demonstrated that they were mucous-secreting goblet cells. Only goblet cells in the mid-villous region, not those at the tip, were immunoreactive. Notably, CaM kinase I immunoreactivity was detected neither in the apical membrane region of intestinal mucosal cells where CFTR is found (Ameen et al. 1995) nor associated with nuclei, as would be consistent with colocalization with CREB. The latter is consistent with previous findings that the intracellular location of CaM kinase I is cytosolic in all parts of the brain that were examined (Picciotto et al. 1995) and in cultured hippocampal neurons (Bito et al. 1996).
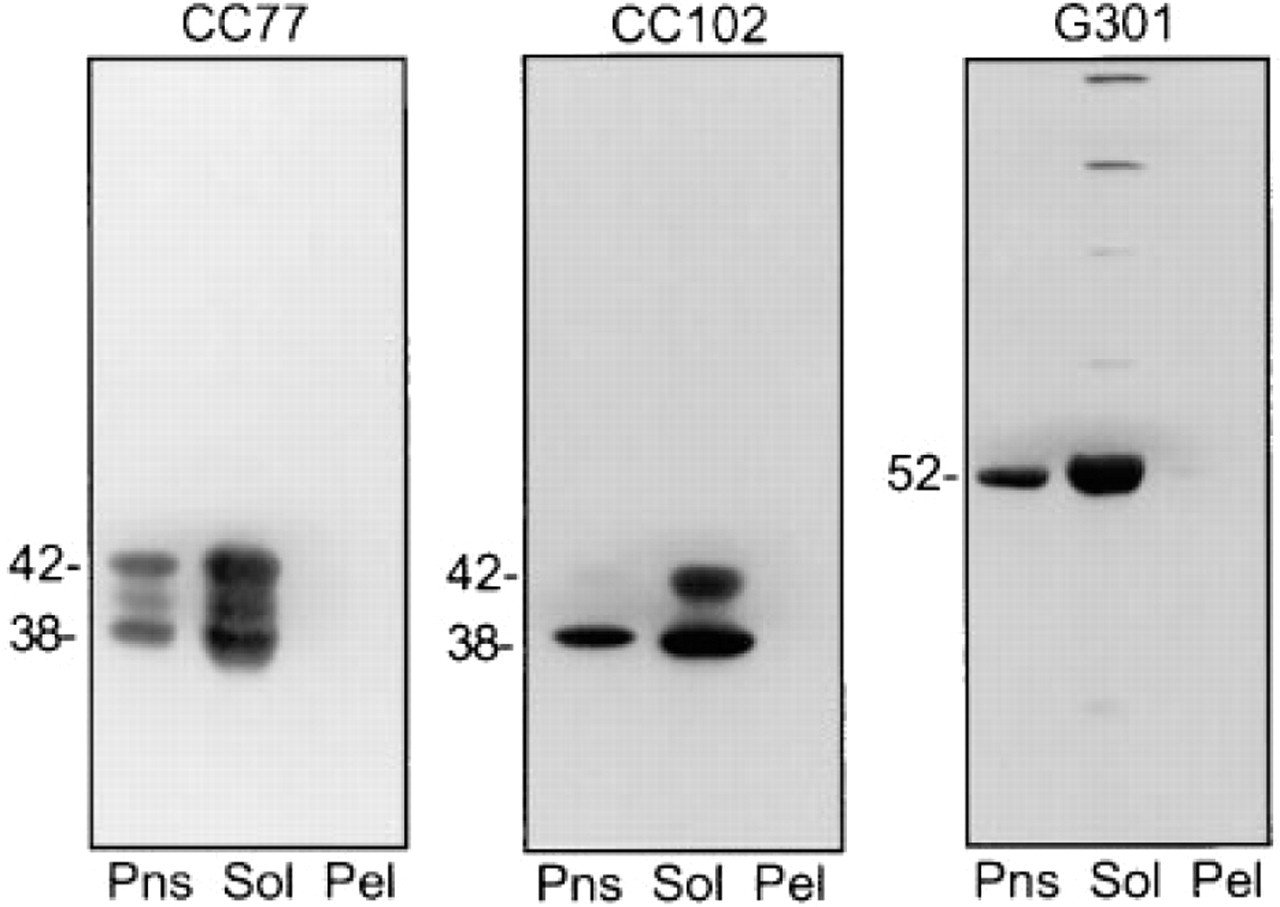
Immunoblot of CaM kinases I and II in rat pancreas particulate and soluble fractions. Pns, postnuclear supernatant; Sol, 100,000 x g supernatant; Pel, 100,000 x g pellet. Gels (8% polyacrylamide) were processed as described for Figure 1 (55 μg protein/lane).
CaM Kinase I and II Are Soluble Proteins in Pancreas
Rat pancreas postnuclear supernatant was centrifuged at 100,000 X g to obtain a soluble and a particulate pellet fraction. Each of these was examined for immunoreactivity with CaM kinase I (CC77 and CC102) and CaM kinase II (G301) by immunoblotting (Figure 3). CaM kinase I was recovered in the soluble fraction but not in the pellet, and therefore is a soluble cytosolic protein in the pancreas, consistent with its predominantly cytosolic localization in the cortex of the brain (Picciotto et al. 1995). CC77 detects approximately equal amounts of the 42- and 38-kD isoforms of CaM kinase I in the pancreas postnuclear supernatant. In addition, in some experiments a third band of ∼40 kD was observed. This is most likely to be a degradation product, but the possibility that it represents another isoform has not been ruled out. CC102 preferentially detected the smaller isoform in the postnuclear supernatant and the soluble fraction. G301 detected an ∼52-kD soluble protein in the pancreas, consistent with the molecular weight of the α-subunit of CaM kinase II.
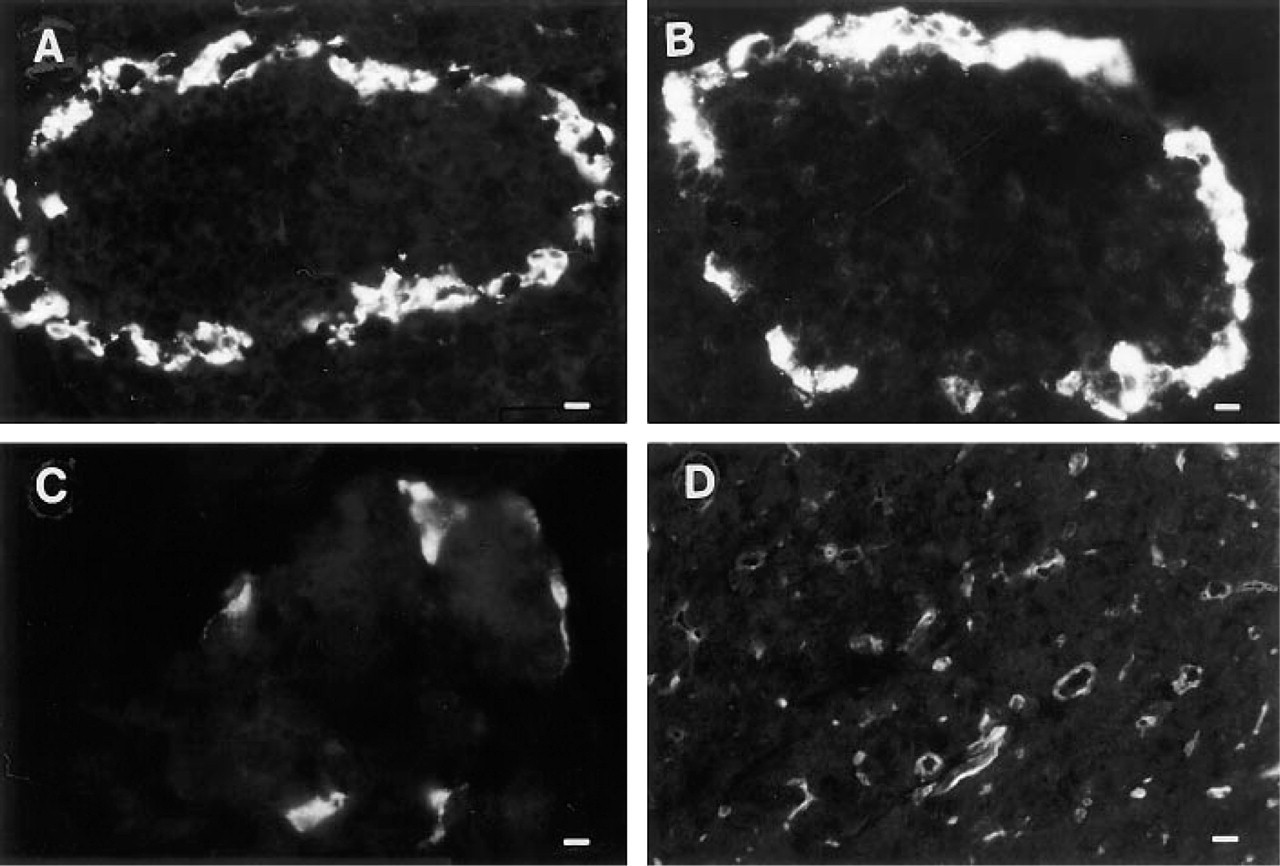
Immunofluorescence localization of CaM kinase I and II in rat pancreas. (
Immunocytochemical Localization of CaM Kinases I and II in Pancreas
The localization of CaM kinase I and CaM kinase II in the pancreas was determined by immunofluorescence microscopy (Figure 4). Both CaM kinases were predominantly expressed in the islet of Langerhans. Antibodies to CaM kinase I (CC77, Figure 4A, and CC102, Figure 4B) detected a signal at the periphery of the islet, with the majority of cells at the mantle of the islet being immunoreactive. A cytoplasmic pattern of labeling was observed; labeling was excluded from the nucleus. Little or no CaM kinase I staining was seen in acinar cells or in the pancreatic duct system. Immuno-absorption with the fusion protein antigen completely blocked CC77-generated immunofluorescence in the pancreatic islet and greatly diminished CC102 staining (data not shown). G301, the antibody specific for CaM kinase II, labeled a subset of cells in the mantle of the islet, but these cells were less numerous than the CaM kinase I-positive cells (Figure 4C). With a slightly higher antibody concentration and longer photographic exposure times, CaM kinase II was also observed to be prominent in the apical region of pancreatic ducts (Figure 4D).
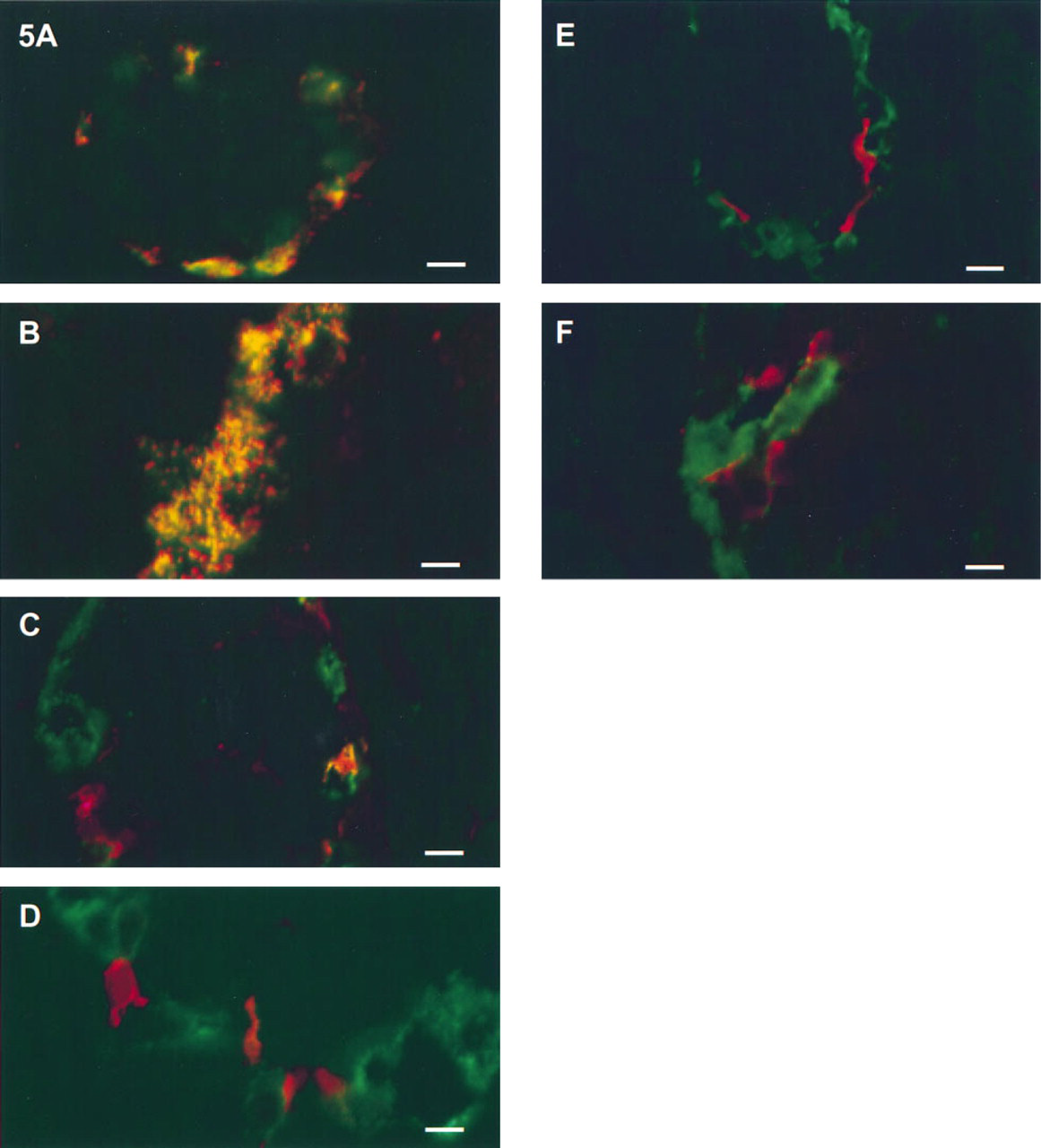
CaM Kinase I Co-localizes with Glucagon in the Islet of Langerhans
Double label studies using islet cell markers were performed to identify the cells immunoreactive for CaM kinase I. Glucagon immunoreactivity was observed in the majority of the cells at the periphery of the islet, and CaM kinase I was co-localized to this cell population (Figure 5A). At higher magnification it was apparent that CaM kinase I had a granular cytoplasmic distribution and co-localized with glucagon granules in the A-cells (Figure 5B). Simultaneous labeling for pancreatic polypeptide and CaM kinase I revealed that these two antigens primarily reside in different cell types (Figure 5C and 5D) but rarely do they co-localize to the same cell (Figure 5C). Double labeling for somatostatin and CaM kinase I revealed no overlap between the two antigens (Figure 5E and 5F).
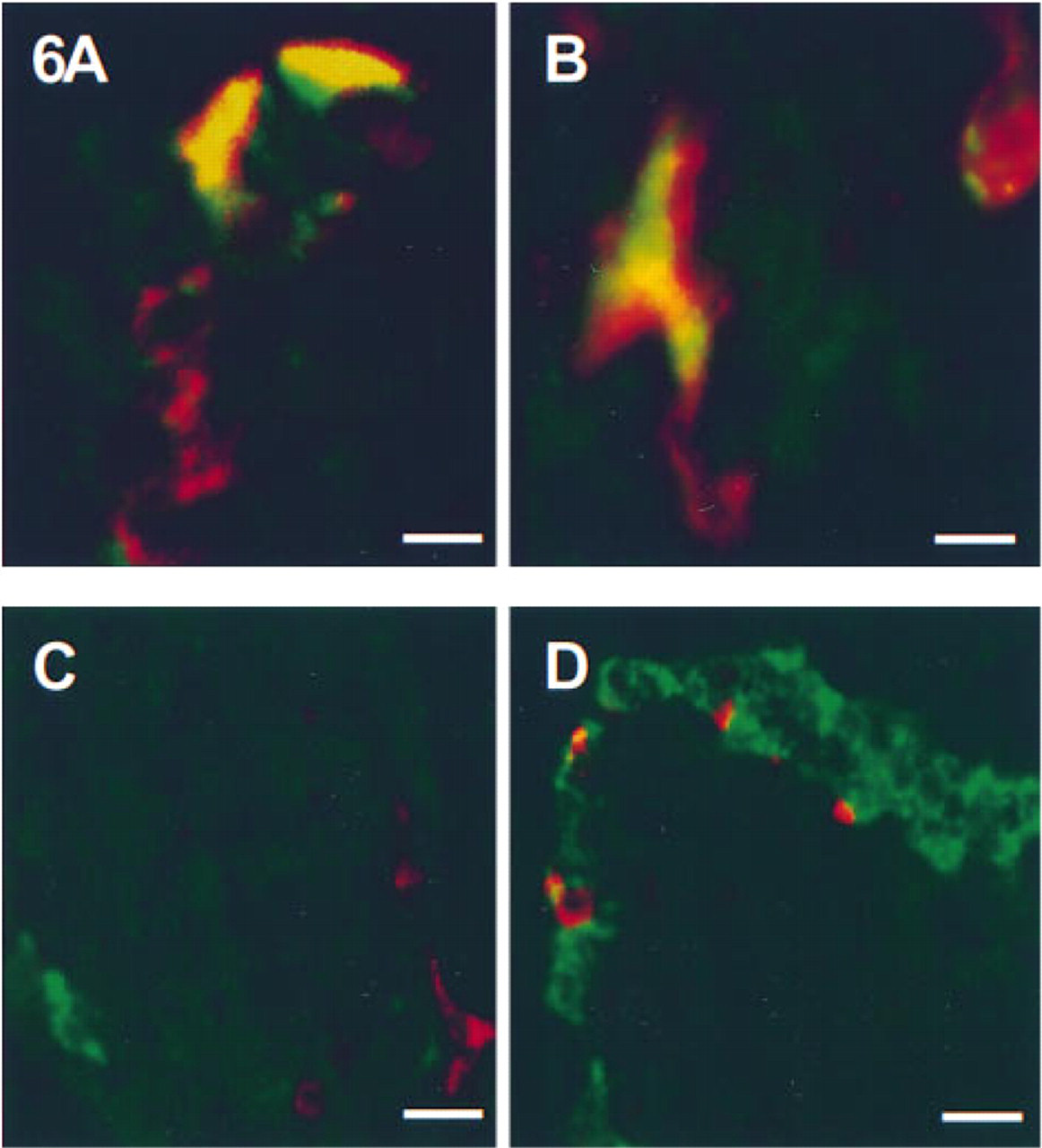
CaM Kinase II Co-localizes with Somatostatin in the Islet of Langerhans
Double label studies with rabbit antibody 301 and mouse monoclonal anti-somatostatin demonstrated that CaM kinase II in the islet is restricted to D-cells (Figures 6A and 6B). Although CaM kinase II was localized to virtually all cells with somatostatin immunoreactivity, the intensity of CaM kinase labeling was variable. Somatostatin was prominent at the cell periphery and in the processes that extend from these neuroendocrine cells; CaM kinase II was concentrated at the center of the cell. There was no co-localization of CaM kinase II with pancreatic polypeptide (Figure 6C) or with glucagon (Figure 6D).
Discussion
The cellular and subcellular distributions of CaM kinase I were examined in rat gastrointestinal tissues. Unexpectedly, the most prominent immunoreactivity was observed in the pancreatic islets of Langerhans. Double label immunolocalization demonstrated that CaM kinase I was localized to the glucagon-containing A-cells and occasional pancreatic polypeptide cells, and was not observed in insulin or somatostatin cells. Within A-cells, CaM kinase I was specifically localized to the glucagon-containing secretory granule. Glucagon secretagogues increase intracellular Ca2+, which leads to vesicle release. Because glucagon secretion is Ca2+-dependent it is attractive to speculate that CaM kinase I may play a regulatory role in glucagon secretion. It may integrate input from spatial and temporal changes in intracellular Ca2+ concentration, such as the Ca2+ oscillations that have been observed to occur spontaneously in a clonal glucagon-secreting cell line derived from hamster endocrine pancreas (Bode et al. 1994).
The distributions of CaM kinases I and II were compared in the rat pancreas and were found to be distinct. CaM kinase II was detected at high levels in the pancreatic islet but in a different cell population. It was specifically found in somatostatin-secreting D-cells, but there was no evidence for co-localization of somatostatin and CaM kinase II to the same intracellular compartment within the D-cell.
A regulatory role for CaM kinase II in insulin secretion has been observed, and recently the ∼2 isoform has been identified on the insulin secretory granule of B-cells (Hughes et al. 1993; Easom et al. 1997; Möhlig et al. 1997). However, using antibody 301, a peptide antibody raised against a sequence that is highly conserved among all major classes of isoforms (Tobimatsu and Fujisawa 1989), we did not detect CaM kinase II in the B-cell by immunofluorescence analysis in this study. This could be either because it is present at a concentration that is below the threshold of detection of the antibody at the concentration used, because the epitope is shielded, or because it does not recognize the CaM kinase II isoform(s) present in B-cells. We consider the latter possibility unlikely because more than 20 isoforms have been described to date and no significant variability in this region has been found (Braun and Schulman 1995; Tombes and Krystal 1997). In addition, the corresponding sequence of the ∼2-isoform is identical to the 301 antigen with the exception of one conservative amino acid substitution (α291K/∼2292R) (Lin et al. 1987; Ngheim et al. 1993).
CaM kinase I is apparently encoded for by only one gene in both rat (Picciotto et al. 1993) and human (Haribabu et al. 1995). The CaM kinase I antibodies CC77 and CC102 specifically recognized several proteins by immunoblot in a tissue-specific manner. CaM kinase I purified from rat brain is comprosed of 42-, 39-, and 37-kD proteins (Nairn and Greengard 1987). A recent study has demonstrated the existence of three isoforms, α, β, and γ, that show considerable homology throughout their catalytic and regulatory regions (Yokokura et al. 1997). The three isoforms differ at their N-terminus and α and β differ at their C-termi-nus (the C-terminus of γ has not yet been defined). The predicted molecular weights of α and β are 41,643 and 38,463 daltons, respectively. Therefore, it is likely that the protein bands that we are detecting represent different isoforms of CaM kinase I. Although both antibodies labeled bands of approximately 42 and 38 kD, they demonstrated tissue specificity, as illustrated in the pancreas and the parathyroid.
Confocal immunofluorescence co-localization of CaM kinase I with endocrine cell markers in the rat pancreatic islet. (
Confocal immunofluorescence co-localization of CaM kinase II with endocrine cell markers in the rat pancreatic islet. (
CaM kinase I has been found to efficiently phosphorylate CFTR, a chloride ion channel, although the physiological relevance of this is unknown (Picciotto et al. 1992; Yokokura et al. 1995). This study was undertaken in part to determine if the location of CaM kinase I might correspond to the distribution of this putative in vivo substrate. However, CaM kinase I was localized neither to the apical membrane of pancreatic duct cells nor to small intestinal mucosal cells that express high levels of the CFTR protein (Ameen et al. 1995). Therefore, these findings do not support a role for CaM kinase I in the regulation of CFTR in these tissues. They do not, however, rule out the possibility that CaM kinase I is present in amounts too small to be detected by the methods used, or that it phosphorylates CFTR in only a subset of the tissues in which it is expressed.
Another in vitro substrate for CaM kinase I is the cAMP response element binding protein (CREB), a nuclear phosphoprotein that regulates transcription (Dash et al. 1991; Sheng et al. 1991). Calcium-dependent phosphorylation of CREB by a CaM kinase cascade that includes nuclear CaM kinase IV has been implicated in the induction of CRE-dependent genes. Using antibody CC77, Bito et al. (1996) have localized CaM kinase I to the nucleus of hippocampal neurons after four divisions in culture, i.e., before synaptogenesis. After nine divisions (following synaptogenesis) CaM kinase I was no longer in the nucleus but was abundant in the cytoplasm, where it may function to regulate gene expression at a specific stage of development. We did not observe CaM kinase I in the nucleus of any of the epithelial cell types examined in this study. However, we have not ruled out the possibility that transient translocation of CaM kinase I to the nucleus may occur in cell types other than hippocampal neurons, regulating transcription by phosphorylation of CREB in concert with CaM kinase IV.
In summary, CaM kinases I and II are found in distinct cell types in the endocrine pancreas that are specialized for the secretion of glucagon and somatostatin, respectively. CaM kinase I is associated with the glucagon secretory granule. On occasion it was observed in pancreatic polypeptide cells but not in acinar or duct cells. In contrast, CaM kinase II was observed predominantly in D-cells of the islet and in the pancreatic duct system. Because CaM kinases I and II are found in different islet cells and have different intracellular localizations, they are likely to have different substrates and different effector mechanisms.
Footnotes
Acknowledgements
Supported by Merit Review Awards from the Veterans Administration to FSG and LMM.
We thank Lillemor Wallmark for performing the immunofluorescence experiments, Michael Lipcan and Steven Rhee for the immunoblots, and Gloria Bertuzzi for technical assistance.