
Abstract
β-Dystroglycan (β-DG) is a dystrophin-associated glycoprotein that is expressed in skeletal muscle and other tissues. In the retina, dystrophin is present in the outer plexiform layer (OPL), where it is enriched under the photoreceptor cell membrane. In this study we determined the immunocytochemical localization of β-DG at both light and electron microscopic levels. β-DG immunoreactivity was detected at the inner limiting membrane, OPL, and around blood vessels. Immunoelectron microscopy detected β-DG immunoreactive products under the photoreceptor cell membrane, which are the same regions of dystrophin localization. In addition, β-DG was detected under the Müller cell membrane that is attached to the paravitreous or perivascular basement membrane. Our results suggest that β-DG may interact with dystrophin in photoreceptor membranes. However, β-DG-related interactions between Müller cells and basement membranes appear to be independent of dystrophin and raise the possibility that β-DG interacts with other molecules. We speculate that β-DG plays a role in maintaining the structural relationship between photoreceptor and bipolar cells or between Müller cells and basement membranes.
D
Recent studies have indicated an oligomeric transmembrane complex in muscle cells that binds dystrophin and utrophin and interacts with several extracellular matrix proteins such as laminin, merosin, and agrin (Ibraghimov–Beskrovnaya et al. 1992; Matsumura et al. 1992; Ervasti et al. 1993; Gee et al. 1993, 1994; Bowe et al. 1994; Campanelli et al. 1994; Yamada et al. 1994). This dystrophin-associated glycoprotein (DAG) complex comprises five transmembrane proteins of 35 kD (35DAG), 43 kD (β-dystroglycan, β-DG), 50 kD (adhalin), and 25 kD (25DAP), one extracellular membrane-associated glycoprotein of 156 kD (α-dystroglycan, α-DG), and one intracellular membrane-associated protein of the syntrophin family.
The human dystroglycan gene maps to chromosome 3p21 (Ibraghimov–Beskrovnaya et al. 1993) and encodes a common precursor protein for α- and β-DG. α- and β-DG are cleaved during or shortly after translation (Ibraghimov–Beskrovnaya et al. 1992). Therefore, both DG isoforms are co-expressed in a variety of tissues such as brain and lung. It is assumed that α-DG interacts with laminin in the extracellular matrix (Gee et al. 1993), whereas β-DG binds to the cysteine-rich dystrophin domain (Suzuki et al. 1992).
It is generally accepted that the retinas of individuals with DMD are functionally and morphologically normal. However, recent studies have described abnormal electroretinograms (ERGs) in individuals with DMD, indicating a reduced amplitude of the b-wave under conditions of dark adaptation (Pillers et al. 1993; Sigesmund et al. 1994). Accordingly, it is possible that abnormal signal transmission might occur in the outer plexiform layer (OPL). Immunocytochemical studies also support a relationship between dystrophin and an abnormal ERG, because dystrophin is expressed in the OPL (Schmitz et al. 1993; Ueda et al. 1995). In the rat retina, we have previously detected dystrophin under the rod spherule membrane by immunoelectron microscopy (Ueda et al. 1995). In addition, dystrophin is observed in cone cells of mouse retina (Ueda et al. 1997). Recently, Montanaro et al. (1995) detected α- and β-DG in the retina of normal and dystrophin-deficient mice (mdx mice), and their distributions were similar to those of dystrophin and utrophin. Drenckhahn et al. (1996) also detected β-DG in the OPL of human retina. In the present study, we examined β-DG localization in the rat retina using confocal and immunoelectron microscopy.
Materials and Methods
Animals and Tissue Preparations
Three adult male Wistar rats were used in the present study. The animals were anesthetized with ethyl ether and sodium pentobarbital and then perfused with 2% paraformaldehyde in 0.1M phosphate buffer (PB), pH 7.3. Their eyeballs were immediately enucleated and divided into anterior and posterior hemispheres. The retinas were gently removed and immersed in the same fixative for 60 min. After treatment with 30% sucrose in PBS, they were embedded in OCT compound and frozen at −80C.
Antibody
An anti-β-DG antibody (NCL-43DAG) was purchased from Novocastra (Tyne, UK). This antibody recognizes 15 of the last 16 amino acids at the C-terminus of the human dystroglycan sequences (PKNMTPYRSPPPYVP-PCOOH.
Immunoblotting
Immunoblotting of β-DG was performed as described previously (Ueda et al. 1997). Rat retinas and skeletal muscles were separately homogenized in lysis buffer, pH 7.6, containing 50 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 1.5 mM aprotinin (Sigma; St Louis, MO), 21 mM leupeptin (Sigma), 1 mM phenylmethyl sulfonyl fluoride (Sigma), 0.02% sodium azide, and 0.1% Triton X-100. They were centrifuged at 10,000 × g for 60 min at 4C, and both super-natants and pellets were boiled with sample buffer (10% SDS, 125 mM Tris-HCl, pH 6.8, 30% glycerol, 5% 2-mer-captoethanol, and 0.02% bromophenol blue) for 5 min at 100C. Subsequently, the samples were loaded onto 10% sodium dodecyl sulfate-polyacrylamide gels for electrophoresis and blotted on Immobilon PVDF membranes (Millipore; Bedford, MA). The blots were pretreated with 5% skim milk in Tris-buffered saline containing 0.05% Tween-20 (TBST) overnight at 4C, and then placed in the following solutions: anti-β-DG antibody (diluted 1:10) in TBST for 60 min at room temperature; goat anti-rabbit IgG antibody conjugated to biotin (diluted 1:300; Amersham, Buckinghamshire, UK) for 60 min; and streptavidin-biotinylated horseradish peroxidase complex (diluted 1:3000; Amersham) for 60 min. The reaction products were visualized with an enhanced chemiluminescence system (Amersham).
Immunofluorescence Microscopy
Immunocytochemical procedures were performed as described previously (Ueda et al. 1995). In brief, the frozen retinas embedded in OCT compound were sliced at 10-μm thickness in a cryostat and placed on gelatin-coated slide glasses. The sections were incubated with 2% bovine serum albumin (BSA) containing 15 mM sodium azide for 30 min to block nonspecific binding. Then they were incubated with the monoclonal anti-β-DG antibody at 1:10 dilution, followed by goat anti-mouse IgG antibody coupled with Texas Red (Molecular Probes; Eugene, OR) at 1:200 dilution for 60 min. They were observed with a confocal laser scanning microscope (Leica TCS 4D; Heidelberg, Germany). For immunocontrols, cryosections were incubated with PBS instead of the primary antibody and followed in the same way.
Conventional Electron Microscopy
Some retinas were routinely fixed with 2.5% glutaraldehyde in PB for 60 min and 1% osmium tetroxide in PB for 60 min. After being rinsed in PBS, they were dehydrated in a graded series of ethanol concentrations and embedded in Quetol-812 (Nissin EM; Tokyo, Japan). Ultrathin sections were prepared, counterstained with uranyl acetate and lead citrate, and observed with a Hitachi H-600 electron microscope (Tokyo, Japan).
Immunoelectron Microscopy
The immunoperoxidase technique was used for immunoelectron microscopy. The 6–10μm cryosections, which had been immunostained with anti-β-DG antibody, were subsequently incubated with rabbit anti-mouse IgG antibody conjugated to biotin and then with horseradish peroxidase conjugated to streptavidin (Nichirei; Tokyo, Japan). After being rinsed in PBS, they were fixed again by 0.25% glutaraldehyde in PB for 10 min. Then they were rendered visible by metal-enhanced DAB (Pierce; Rockford, IL) and treated with 1% osmium tetroxide in PB for 30 min. After routine dehydration in a graded series of ethanol and acetone concentrations, they were embedded in epoxy resin by the inverted gelatin capsule method. Finally, ultrathin sections were counterstained with uranyl acetate and lead citrate and observed in an electron microscope.
Results
Immunoblotting Analysis
To check the specificity of the anti-β-DG antibody, immunoblotting was performed with skeletal muscle supernatants (SM), retina supernatants (RS), and retina pellets (RP). Figure 1 shows immunoblotting of rat skeletal muscle and retina extracts with anti-β-DG antibody. All lanes clearly show a 43-kD molecular weight band (arrowhead), which was most intense in the RP lane. Additional bands were also observed in all lanes. A 114-kD band was labeled intensely in the RS and RP lanes but was only faintly detectable in the SM lane. A 65-kD band was observed in all lanes, and a 29-kD band was observed only in the RP lane.
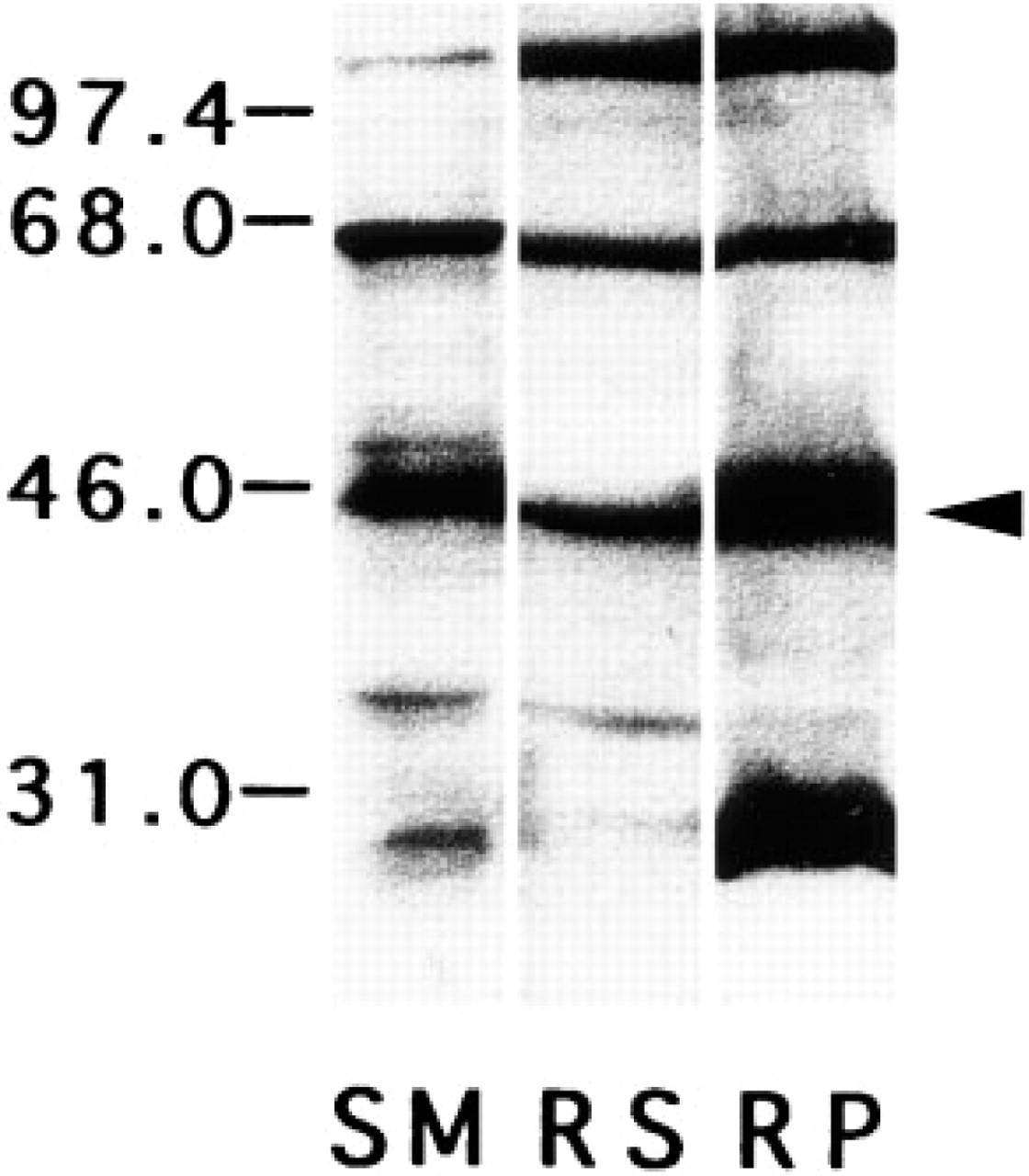
Immunoblotting of extracts from the skeletal muscle and the retina with anti-β-dystroglycan antibody. All lanes clearly show a 43-kD molecular weight band (arrowhead). In addition, the SM lane shows an approximately 65-kD molecular weight band, and the RS lane shows approximately 65-kD and 114-kD molecular weight bands. The RP lane shows four immunoreactive bands of approximately 29 kD, 43 kD, 65 kD, and 114 kD. RS, retinal supernatant; RP, retinal pellet; SM, skeletal muscle supernatant.
Confocal Laser Scanning Microscopic Analysis
β-DG distribution in the retina was examined with immunofluorescence staining and observed by confocal laser scanning microscopy. β-DG was detected in the inner limiting membrane (ILM) and around blood vessels of rat retinas. In addition, it was detected in the OPL like distinctive foci (Figure 2A), as well as dystrophin (Schmitz et al. 1993; Ueda et al. 1995). These β-DG staining patterns were the same as reported previously (Montanaro et al. 1995; Drenckhahn et al. 1996). Immunocontrol sections showed no labeling (Figure 2B). In the obliquely cut sections, larger immunostaining dots were definitely recognized at higher magnifications (Figure 2C). Small dots may represent rod spherules and large ones cone pedicles. These findings were consistent with that of the dystrophin immunostaining pattern (Schmitz et al. 1993; Ueda et al. 1997). Therefore, β-DG has both synaptic and nonsynaptic localization.
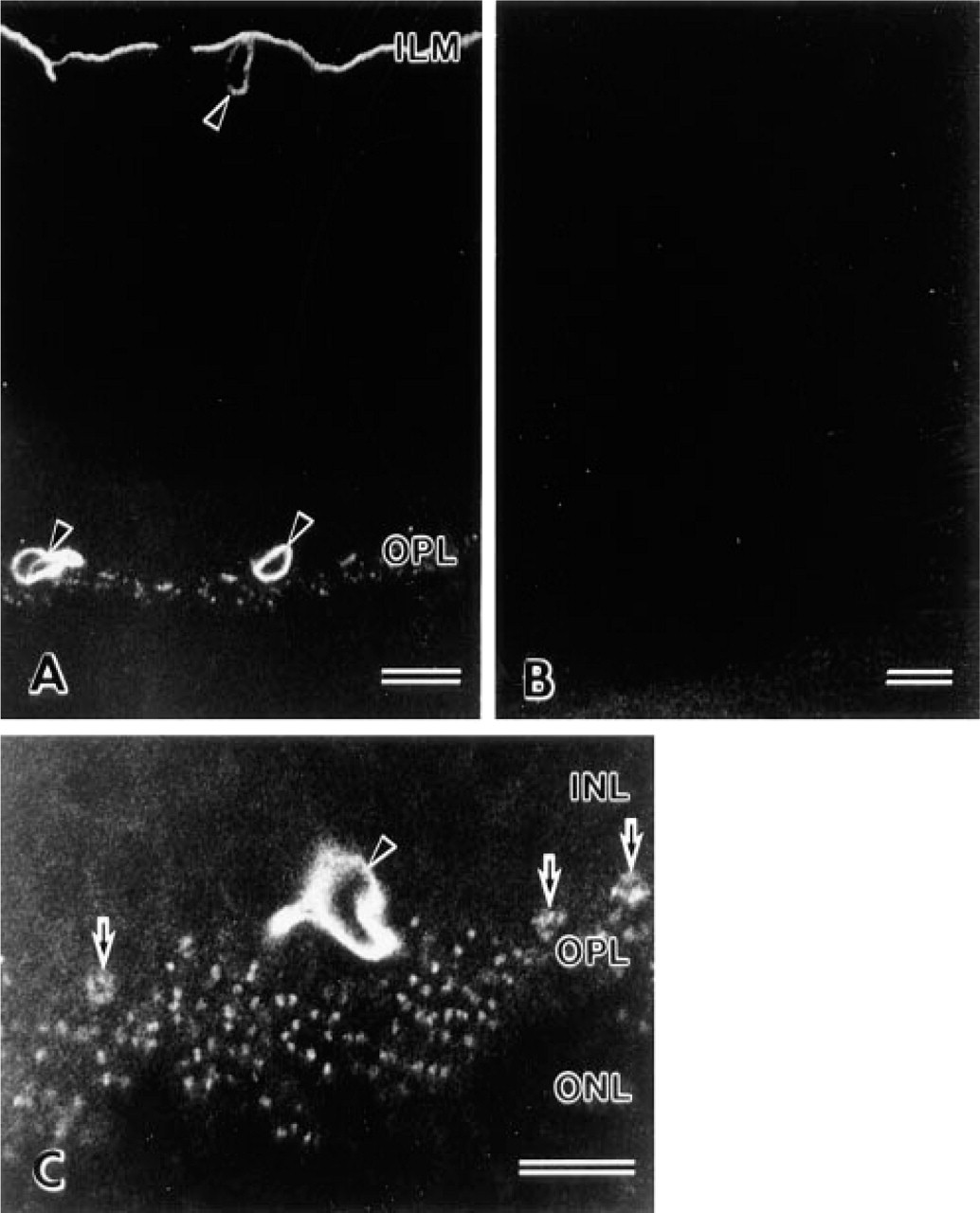
Confocal images of β-dystroglycan distribution in the rat retina. (
Electron Microscopic Analysis
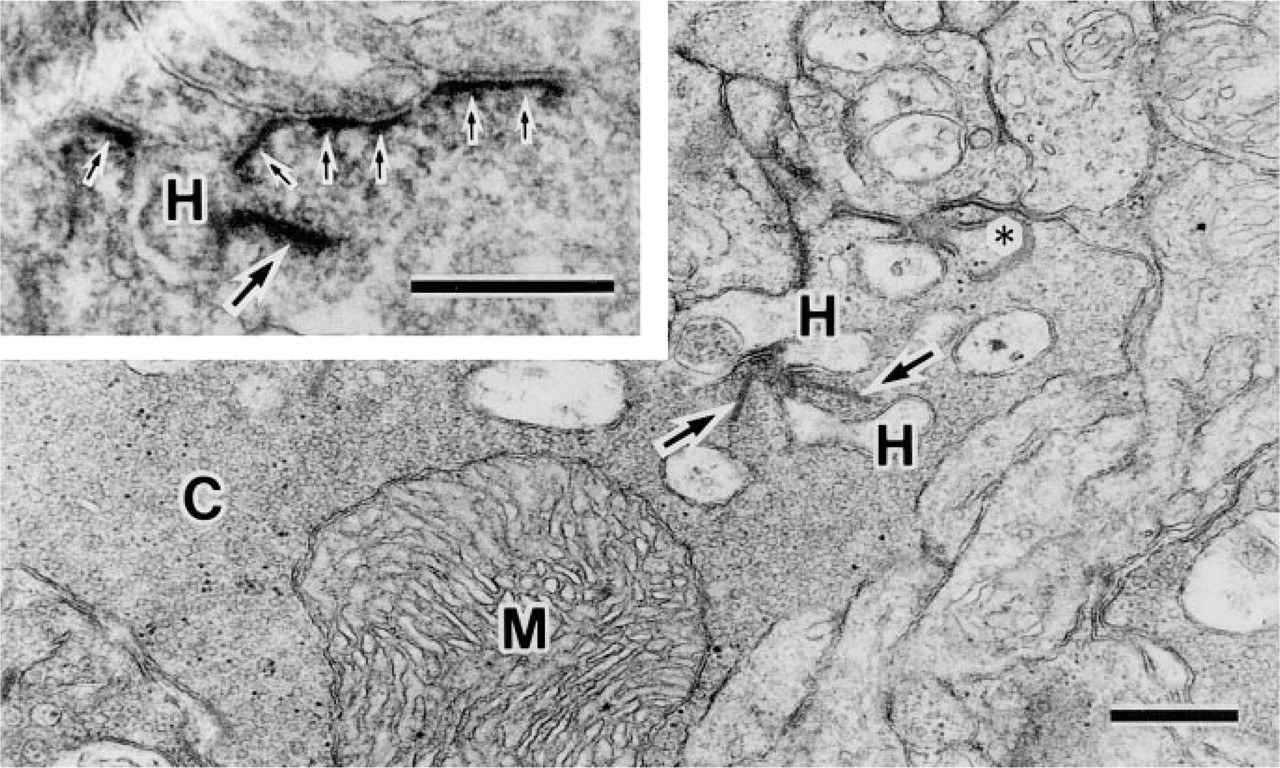
β-DG localization in the retina was investigated at the ultrastructural level. The ILM of the retina consists of paravitreous basement membranes and Müller cell membranes, and electron-dense regions are present under the Müller cell membranes (Figure 3, inset). β-DG-immunoreactive products were discontinuously detected under the Müller cell membrane (Figure 3), and their localization was consistent with the electron-dense regions. Müller cells always encircled the blood vessels in the retina and attached to the basement membrane around the endothelial cells or vascular smooth muscle fibers (Figure 4A). They had electron-dense regions beneath the cell membrane as well as paravitreous ones. β-DG immunoreactive products were detected under the Müller cell membrane that is attached to the perivascular basement membrane (Figure 4B) but was not detected beneath the sarcolemma of the vascular smooth muscle fibers (data not shown).
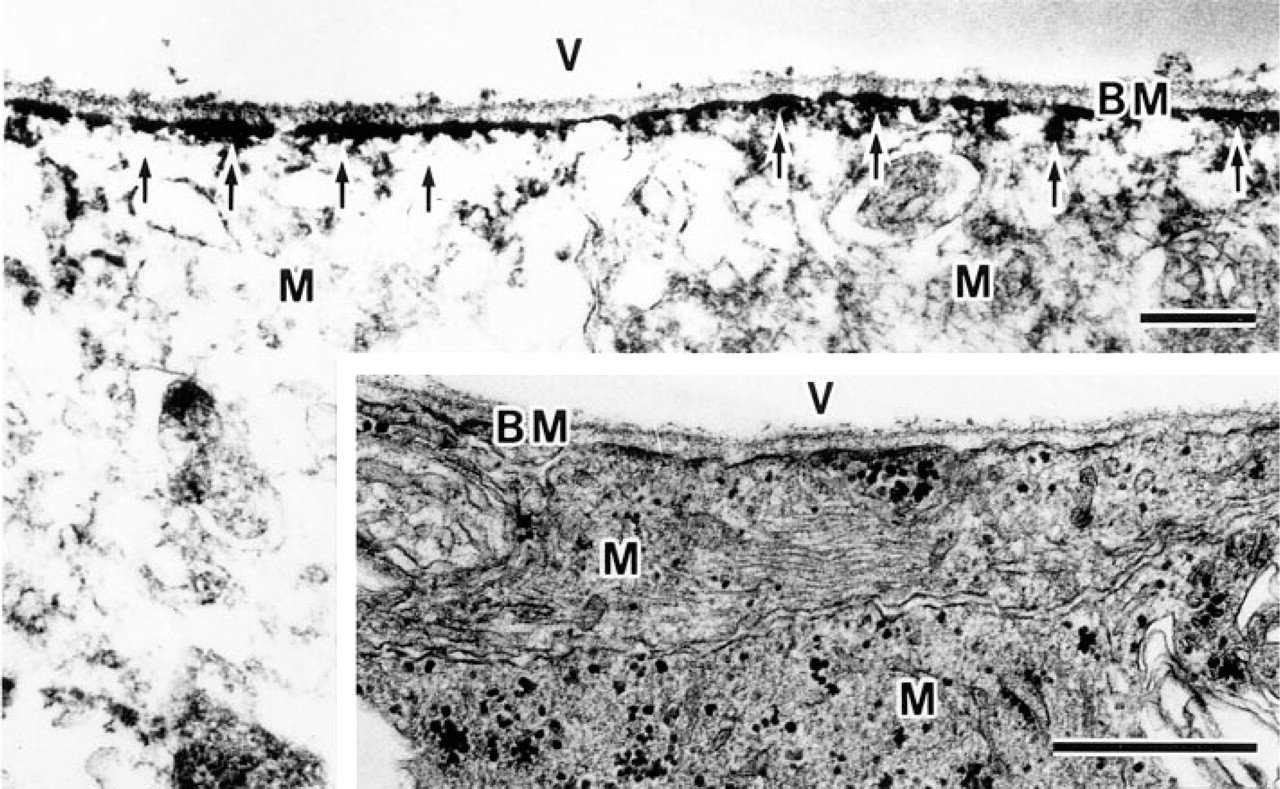
Electron micrographs of the inner limiting membrane. Immunoreactive products of β-dystroglycan (arrows) are observed under the Müller cell membrane attached to the basement membrane. (Inset) Conventional section. BM, basement membrane; M, Müller cell; V, vitreous body. Bars = 0.5 μm.
Electron micrographs of conventional (
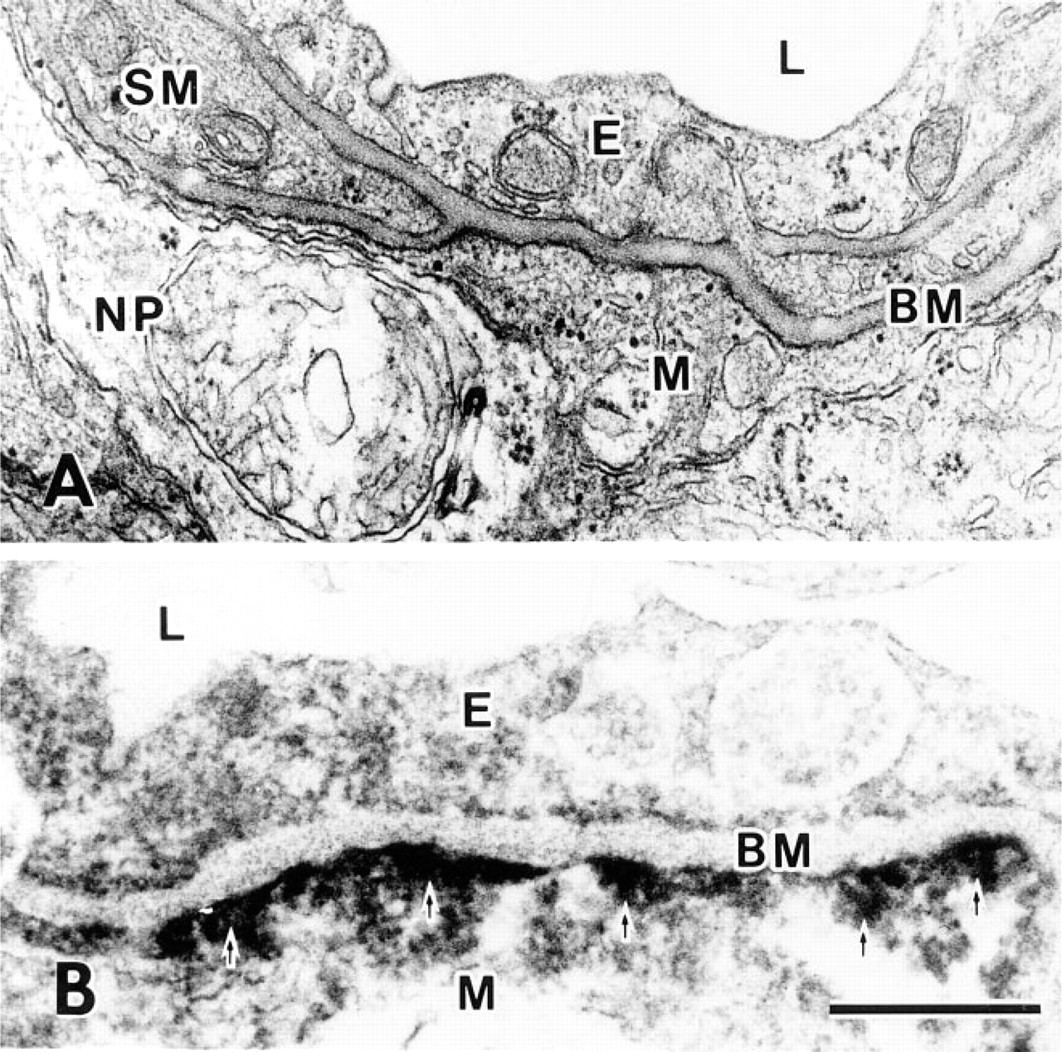
β-DG localization in the rod spherule was compared with conventional sections (Figure 5). Two bipolar cell processes were separated by a rod spherule projection (Figure 5A). It was noted that electron-dense regions were present under the rod cell membrane facing bipolar cells but not facing horizontal cells. In a corresponding immunoelectron micrograph, β-DG immunoreactive products were detected around two bipolar cell processes (Figure 5B) but not around horizontal cell processes or synaptic ridges (Figure 5C). β-DG localization is consistent with the submembranous electron-dense regions, and its distribution is the same to dystrophin (Ueda et al., in press). The synaptic ribbon was not different from that of control sections, even though it had high electron density. Accordingly, it is impossible to state that β-DG is expressed at synaptic ribbons.
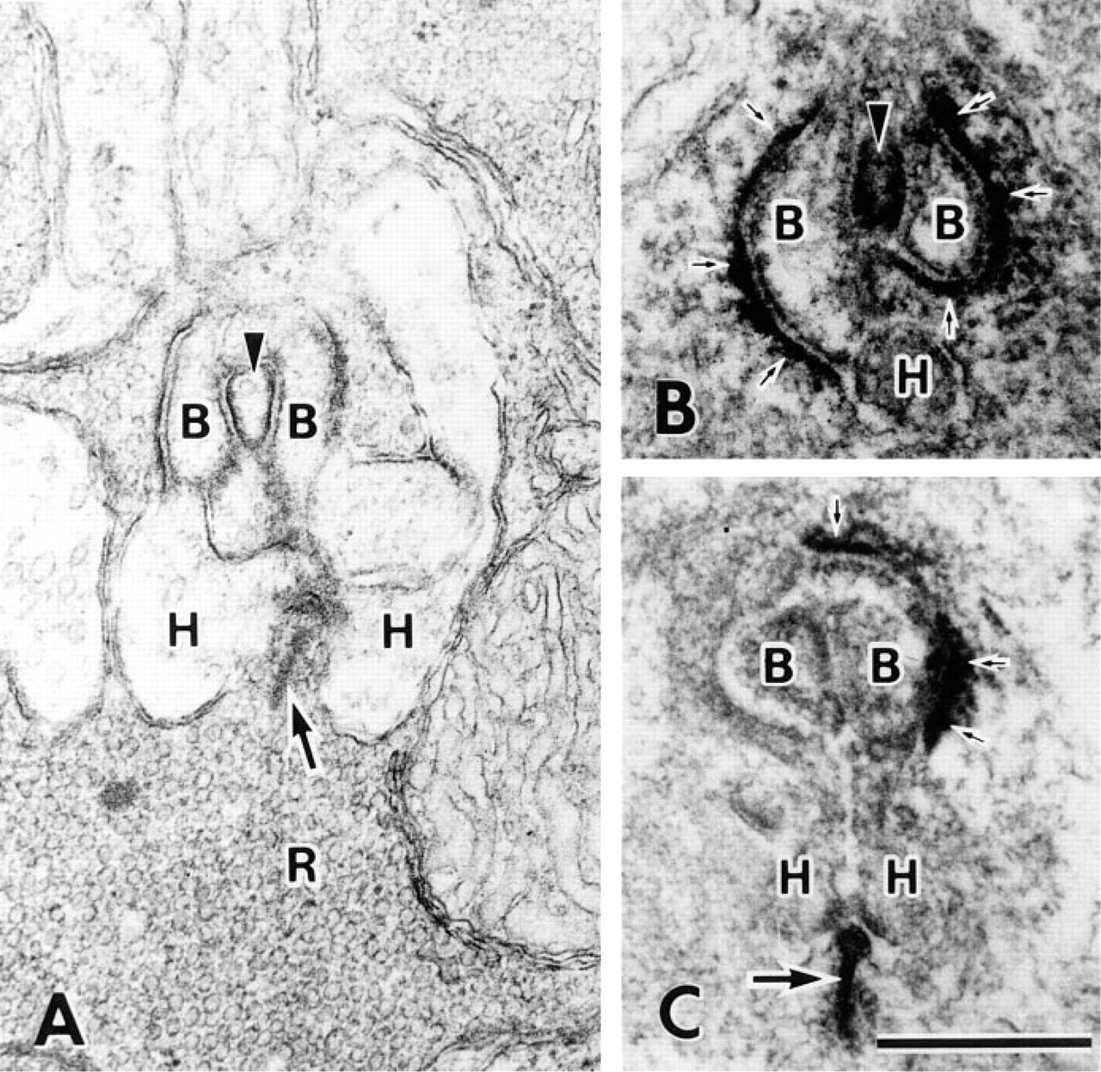
Electron micrographs of the rod synapse. (
Cone cells have two synapses, an invaginated synapse and a flat synapse (Figure 6). Invaginated synapses have horizontal cell processes and synaptic ribbons, but flat synapses have neither. Cone cell membranes facing the bipolar cell processes showed high electron density at both synapses as well as at the rod cell membrane. The inset to Figure 6 shows β-DG immunoreactivity at a flat synapse under the cone cell membrane.
Electron micrographs of the cone cell. Conventional section shows both invaginated and flat synapses (asterisk). (Inset) Immunoreactive products of β-dystroglycan (small arrows) are observed under the cone cell membrane of the flat synapse. Large arrows indicate synaptic ribbons. C, cone cell; M, mitochondrion. Bars = 0.5 μm.
Discussion
In the present study, β-DG was detected in the rat retina using immunoblotting and immunocytochemistry. Immunofluorescence stainings showed that β-DG was expressed in the INL, OPL, and around blood vessels. At the ultrastructural level, β-DG was localized under Müller cell membranes that are attached to basement membranes, and under photoreceptor cell membranes facing bipolar processes. Considering these locations, β-DG may function in a number of membrane-membrane or membrane-extracellular matrix interactions.
A monoclonal anti-β-DG antibody showed some immunoreactive bands in addition to a 43-kD molecular weight band in the rat retinal extracts. It is assumed that β-DG interacts with α-DG and dystrophin/utrophin to connect cell membranes to the extracellular matrix in skeletal muscles. Therefore, larger molecular weight bands might indicate complexes of β-DG and other proteins, and a small molecular weight band in the retinal pellet might indicate a novel β-DG isoform. However, it is not known whether the β-DG immunoreactive products represent β-DG tightly associated with other proteins or spliced novel isoforms encoded by the same gene.
In this study, β-DG was clearly detected under Müller cell membranes that were attached to paravitreous or perivascular basement membranes. Müller cells, the main glial cells in the vertebrate retina, penetrate all retinal layers perpendicularly. The plasma membrane of Müller cells is characterized by orthogonal arrays of particles (OAPs) that are revealed by the freeze-fracture replica method (Landis and Reese 1981; Wolburg and Berg 1988). Although OAPs are present in all surface membrane of the Müller cells, pericapillary endfeet and paravitreous endfeet have a much higher OAP density than non-endfoot membranes (Wolburg and Berg 1988). In vascularized mammalian retina, K+ efflux occurs from several cell regions having a high density of K+ channels: paravitreous endfeet, pericapillary endfeet, and apical end-foot-like villi (Newman and Reichenbach 1996). Therefore, it has been hypothesized that the OAP may be the structural manifestation of specific K+ channels (Wolburg and Berg 1988). β-DG distribution is very similar to that of OAP, and it may contribute to OAP structure because β-DG is a transmembrane protein. However, to date, dystrophin and/or utrophin have not been detected in Müller cells. Therefore, β-DG may bind proteins other than dystrophin or utrophin and may be associated with the K+ channel.
In our previous study, dystrophin was detected in rod and cone cells (Ueda et al. 1995, 1997), at submembranous dense regions, including rod spherule projections (Ueda et al. in press). The present study showed that β-DG was also localized at submembranous dense regions in photoreceptor cells, including rod spherule projections. These projections have been reported to be of functional importance in lower vertebrates because they receive feedback input from horizontal cell processes, but their function has not been elucidated in mammalian retinas (Linberg and Fischer 1988). Recently, the first C-terminal missense of dystrophin in a DMD patient raised the absence of b-waves in dark-adapted ERG (Lenk et al. 1996). The dystrophin C-terminal portion is the dystroglycan binding domain (Ozawa et al. 1995; Suzuki et al. 1992,1994,1995) and is present in all dystrophin isoforms. Therefore, taken together with our findings, these projections may be important structures for a physiological function, and dystrophin and/or β-DG may be associated with signal transmission from photoreceptor to bipolar cells.
Recent studies have suggested that dystroglycan might function as a receptor for basement membrane components during epithelial morphogenesis (Durbeej et al. 1995) or as a mediator between the decidua and trophoblasts (Yotsumoto et al. 1996). However, it is likely that the DAG complex mechanically connects the sarcolemma to the basement membrane in the skeletal muscle fiber (Ohlendiek 1996). At present, it is reasonable to speculate that the DAG complex mainly plays a role in maintaining the structural relationship between Müller cells and basement membranes or between photoreceptor and bipolar cells.
In summary, β-DG is localized under the Müller cell membrane attached to the paravitreous or perivascular basement membrane. In addition, it is localized at submembranous dense regions in the photoreceptor cells, along with dystrophin. However, further studies are needed to determine the function of β-DG in the retina.
Footnotes
Acknowledgements
We thank Dr Bruce D. Trapp (Department of Neuro-sciences, The Cleveland Clinic Foundation) for checking our manuscript. We also thank Ms Y. Kato and Ms K. Ariizumi for excellent assistance.