
Abstract
PAR-2 is a second member of a novel family of G-protein-coupled receptors characterized by a proteolytic cleavage of the amino terminus, thus exposing a tethered peptide ligand that autoactivates the receptor. The physiological and/or pathological role(s) of PAR-2 are still unknown. This study provides tissue-specific cellular localization of PAR-2 in normal human tissues by immunohistochemical techniques. A polyclonal antibody, PAR-2C, was raised against a peptide corresponding to the amino terminal sequence SLIGKVDGTSHVTGKGV of human PAR-2. Significant PAR-2 immunoreactivity was detected in smooth muscle of vascular and nonvascular origin and stromal cells from a variety of tissues. PAR-2 was also present in endothelial and epithelial cells independent of tissue type. Strong immunolabeling was observed throughout the gastrointestinal tract, indicating a possible function for PAR-2 in this system. In the CNS, PAR-2 was localized to many astrocytes and neurons, suggesting involvement of PAR-2 in neuronal function. A role for PAR-2 in the skin was further supported by its immunolocalization in the epidermis. PAR-2C antibody exemplifies an important tool to address the physiological role(s) of PAR-2.
Keywords
T
The tissue distribution and hence the function of PAR-2 have only recently begun to be elucidated. Expression patterns derived from Northern analysis indicated an abundance of PAR-2 mRNA in kidney, stomach, pancreas, liver, colon, and small intestine (Nystedt et al. 1994; Böhm et al. 1996). Since the discovery of PAR-2, several in vitro studies have been reported describing the functional stimulation of PAR-2. The cellular localization of PAR-2 was first described for human keratinocytes (Santulli et al. 1995) and it was subsequently shown to be expressed in vascular endothelial cells (Hollenberg et al. 1996; Mirza et al. 1996), epithelial cells of the gastrointestinal tract (Böhm et al. 1996), gastric smooth muscle (Hollenberg et al. 1996), pancreatic acinar cells (Böhm et al. 1996), and lung adenocarcinoma cells (Böhm et al. 1996). The localization of PAR-2 protein in vivo, however, has not yet been described. Localization of PAR-2 protein in various tissues may help to construct hypotheses about its function, especially in comparison to the well-described PAR-1. For this purpose, we generated a polyclonal antibody, PAR-2C, directed against the tethered agonist peptide region of human PAR-2 (residues 37–53; SLIGKVDGTSHVTGKGV).
We used this antibody to immunophenotype a series of normal human tissues. In addition, an antibody to the human PAR-1 was used for tissue-specific comparisons. We report significant PAR-2 immunolocalization in epithelial cells and smooth muscle of vascular and nonvascular origin. We also provide in vivo evidence for PAR-2 expression in the brain, supporting a recent report demonstrating PAR-2 expression and function in rat hippocampal cultures (Smith–Swintosky et al. 1997).
Materials and Methods
Preparation of Antibodies
Peptides were linked through a terminal cysteine to maleide-activated KLH (Pierce; Rockford, IL) according to the manufacturer's instructions. For antibody generation to human PAR-2, a peptide containing the sequence SLIGKVDGTSHVTGKGVC (residues 37–53) was used. This sequence is present in both cleaved and uncleaved receptors. Unlinked peptide was removed by dialyzing against PBS (50-kD cutoff). New Zealand White rabbits were immunized by
WEDE15, an antibody specific to PAR-1, is a previously described IgG1 monoclonal antibody (MAb) produced in mice immunized with PAR-1 peptide KYEPFWEDEEKNES, corresponding to residues 51–64 of the human PAR-1 sequence (Hoxie et al. 1993). WEDE15 was used at a titer of 2 μg/ml for immunohistochemical staining.
Expression of PAR-1 and PAR-2 in Sf9 Cells
A 430-BP fragment of the murine PAR-2 cDNA was used as a probe for screening a human keratinocyte cDNA library (Clontech; Palo Alto, CA). A full-length clone of human PAR-2 was identified. Restriction sites were introduced immediately before and after the coding region of this clone by PCR and the cDNA was cloned in the Baculovirus expression vector pAcMP2. A human PAR-1 cDNA was similarly modified and cloned in the above vector. Each expression construct was co-transfected with baculogold AcNPV DNA (PharMingen; San Diego, CA) in Sf9 cells according to the manufacturer's instructions. Recombinant viral clones were amplified and titrated by infection of Sf9 cells.
Analysis of PAR-1 and PAR-2 Expression by Flow Cytometry
Monolayer cultures of Sf9 cells were infected with PAR-1- or PAR-2-encoding virus at a multiplicity of infection of 2 and were incubated for 72 hr at 27C. Cells were dislodged, washed once in PBS, and 5 × 105 cells were resuspended in staining buffer (PBS with 1% BSA and 0.02% sodium azide) with the PAR-2C or WEDE15 antibody at 10 μg/ml. Cells were stained for 1.5 hr on ice, washed with ice-cold PBS, and resuspended in staining buffer containing 5 μg/ml FITC-conjugated goat anti-rabbit (PAR-2C-stained cells) or 5 μg/ml FITC-conjugated goat anti-mouse antibody (WEDE15 stained cells) (Cappel; Durham, NC), and 10 μg/ml propidium iodide. The stained cells were analyzed on a FACScan flow cytometer (Becton Dickinson; Mountain View, CA). The live cells were gated.
Cell Culture
Human neonatal epidermal keratinocytes and human dermal fibroblasts (AG1523) were cultured as previously described (Santulli et al. 1995). Cells were plated on gelatin-coated Lab-Tek chamber slides (Fisher Scientific; Pittsburgh, PA) and grown to 70–80% confluence. Cells were washed twice with PBS and subsequently fixed in 10% neutral buffered formalin for 2 hr at room temperature (RT). Cells were then washed three times in PBS and stored at 4C until immunocytochemical staining.
Human Platelets
Human platelet concentrate (105 platelets/(μl) was purchased from Biological Specialty Corporation (Colmar, PA). Platelets were collected from drug-free human donors by plasmaphoresis on a Cobe Spectra using ACD (acid citrate dextrose solution). Human platelets were routinely fixed in 4% paraformaldehyde for 2 hr at RT and then processed for paraffin embedding.
Immunohistochemistry
Human multitissue blocks were purchased from Biomeda (Foster City, CA) and Dako (Carpenteria, CA). Tissue sections were mounted on slides, deparaffinized, and hydrated. Slides were immersed in Target reagent (Dako) heated two times for 3 min at high power in an 800-W commercial microwave. After the slides cooled, the endogenous peroxidase was blocked by 3.0% H2O2 for 10 min. Tissue slides were processed through an avidin–biotin blocking system according to the manufacturer's instructions (Vector Labs; Burlingame, CA) and then placed in PBS. Platelets were embedded in paraffin, cut onto slides, and then routinely dewaxed, hydrated, and then placed in PBS.
All reagent incubations and washes were performed at RT. Normal blocking serum was then placed on all slides for 10 min (Vector Labs). After brief rinsing in PBS, primary antibodies were placed on the slides for 30 min (Table 1). The slides were washed and then the biotinylated secondary antibodies, goat anti-rabbit (PAb) or horse anti-mouse (MAb) were placed on the tissue sections for 30 min (Vector Labs). After rinsing in PBS, the avidin–biotin–HRP complex reagent (Vector Labs) was added for 30 min. The slides were washed and treated with the chromogen 3,3′-diaminobenzidine (Biomeda; Foster City, CA) two times for 5 min and rinsed in dH2O, counterstained in Mayer's hematoxylin, dehydrated, and then coverslipped. The slides were visualized and photographed using an Olympus BX50 light microscope.
Primary antibodies used to characterize cell-specific markers
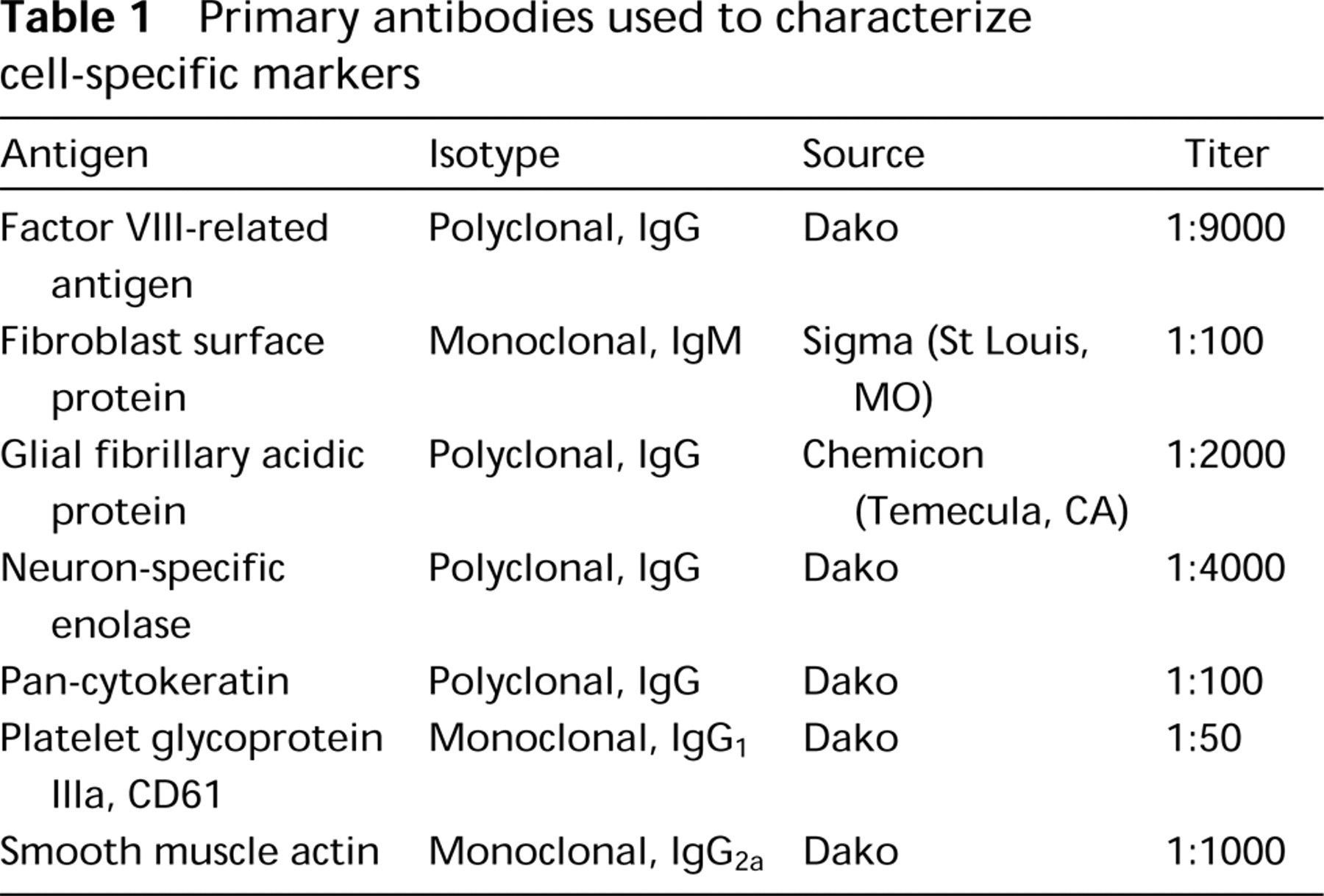
Controls were run for each tissue. Negative controls included replacement of the primary antibody with the antibody diluent (Zymed Laboratories; San Francisco, CA) and the use of the same species isotype nonimmunize serum. Specificity of the antibodies for PAR-2 and PAR-1 was confirmed by preincubation overnight at 4C with their respective antigen in a 20-fold molar excess of antigen to antibody. In addition, the PAR-2C antibody was preincubated with peptide corresponding to the PAR-1 amino terminal sequence SFLLRNPNDKYEPF relative to the antigen sequence used to generate PAR-2C. Positive control antibodies included relevant cellular markers (Table 1).
Results
Specificity of PAR-2C
The specificity of PAR-2C for PAR-2 was first examined using cells expressing recombinant human PAR-2. Flow cytometry analyses performed on Sf9 cells infected with recombinant baculoviruses containing either human PAR-2 or PAR-1 cDNA demonstrated that PAR-2C was selective for PAR-2 and did not bind to PAR-1 (Figure 1). In addition, the MAb WEDE15 recognized PAR-1 and did not crossreact with PAR-2.
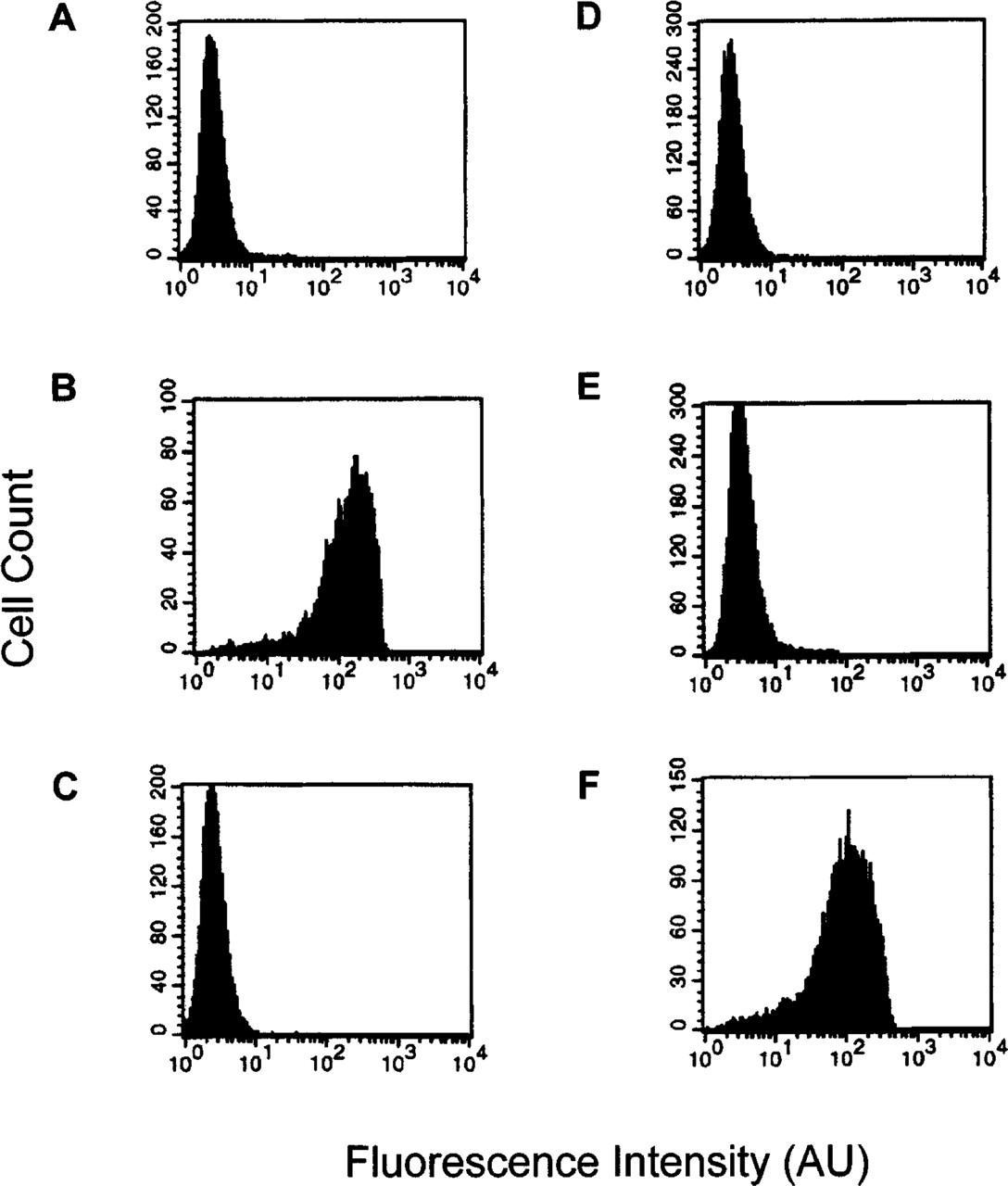
PAR-2C does not crossreact with PAR-1. Sf9 cells were infected with human PAR-2 encoding virus (
Negative immunohistochemical controls were devoid of any detectable immunolabeling. Preincubation of PAR-2C and the PAR-2 antigen also resulted in absence of immunolabeling. Furthermore, preincubation of PAR-2C with the PAR-1 antigen did not affect PAR-2C immunolabeling, providing additional evidence that PAR-2C does not crossreact with PAR-1. The positive control antibodies labeled according to the manufacturer's specifications.
To further characterize the specificity of this antibody for endogenous PAR-2, staining patterns were evaluated in human dermal fibroblasts and human epidermal keratinocytes. We have previously demonstrated that both cell types express PAR-1 mRNA and possess functional PAR-1. However, PAR-2 is expressed exclusively in keratinocytes and not in fibro-blasts (Santulli et al. 1995). This pattern of receptor expression was maintained in the cell cultures examined. Human epidermal keratinocytes exhibited positive staining for PAR-2 (Figure 2A), whereas human dermal fibroblasts were negative for PAR-2 (Figure 2B). Both cell types were positive for PAR-1 as well as the respective cell-specific markers cytokeratin and fibroblast surface protein, which labeled keratinocytes and fibroblasts, respectively (data not shown).
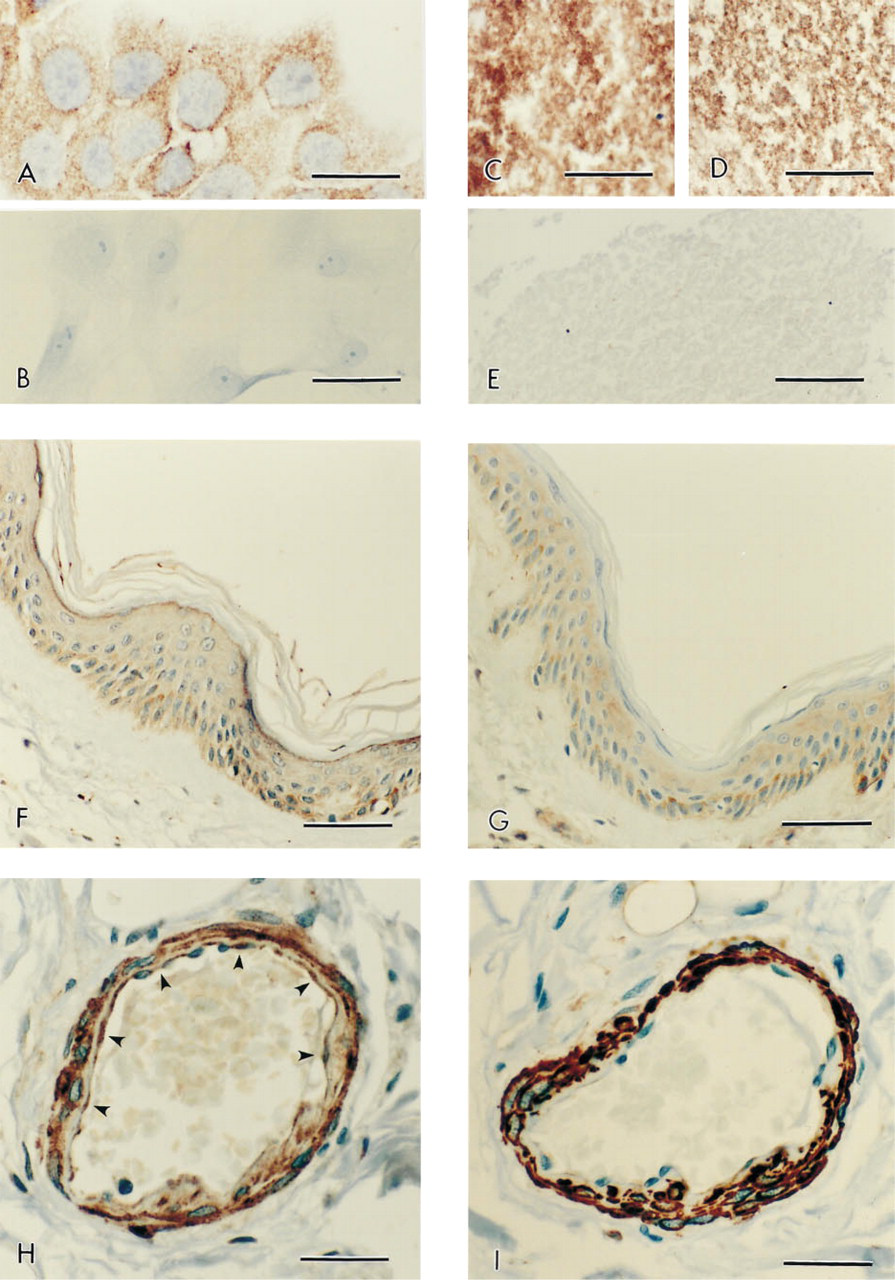
Examples of immunolabeling results for PAR-2 and several other immunohistochemical markers on representative human tissues. (
Human platelets represent another cell type that expresses PAR-1 but does not express PAR-2 (Hwa et al. 1996). To further demonstrate the specificity of PAR-2C labeling, platelets were immunolabeled with PAR-2C. Figure 2C demonstrates intense immunolabeling for the positive control antibody, murine anti-human platelet glycoprotein IIIa (CD61) in the human platelets. Figure 2D demonstrates positive immunolabeling of PAR-1. As seen in Figure 2E, however, human platelets did not immunolabel with PAR-2C.
Tissue and Cellular Distribution of PAR-2
The distribution of PAR-2 immunolocalization in human tissues is summarized in Table 2. PAR-2 was abundant in the skin, vasculature, gastrointestinal tract, lung, brain, prostate, and uterus. Overall, PAR-2 exhibited marked expression in most epithelial and smooth muscle cells.
Immunolocalization of PAR-2 in normal human tissues a
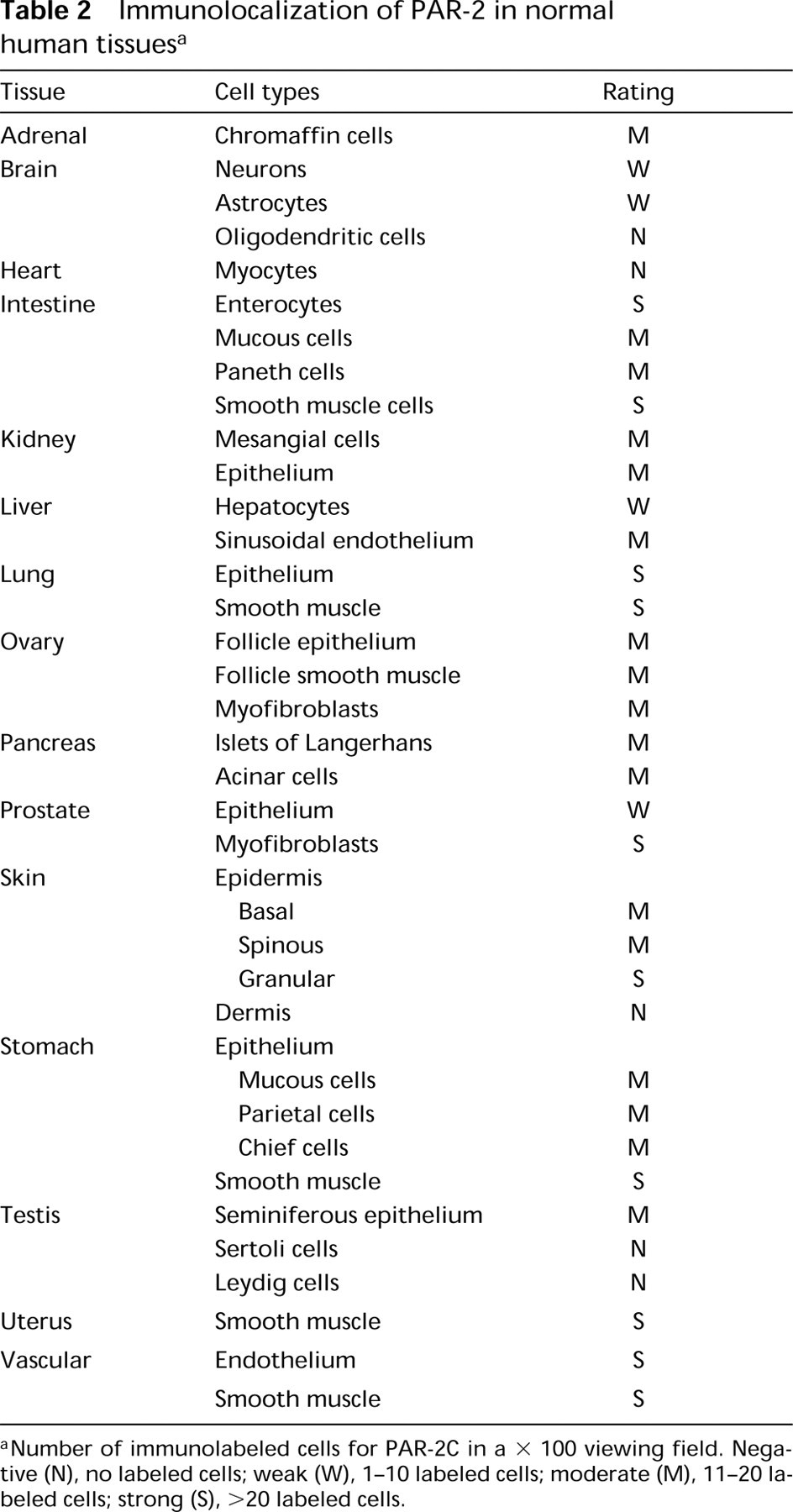
Number of immunolabeled cells for PAR-2C in a × 100 viewing field. Negative (N), no labeled cells; weak (W), 1–10 labeled cells; moderate (M), 11–20 labeled cells; strong (S), >20 labeled cells.
In the skin, PAR-2 was observed in the basal, spinous, and granular cell layers of the epidermis (Figure 2F). PAR-1 was localized in the basal and spinous cell layers of the epidermis (Figure 2G). However, in contrast to the immunolocalization of PAR-2, PAR-1 was not expressed in the granular layer of the epidermis. In addition, stromal fibroblasts of the dermis were negative for PAR-2 but were positive for PAR-1, consistent with the results in cultured human dermal fibroblasts (data not shown).
PAR-2 was present throughout the vasculature, with strong labeling in both the vascular smooth muscle and the endothelium. As an example, Figures 2H and 2I demonstrate the presence of PAR-2 and smooth muscle actin, respectively, in a human artery from the intestinal submucous plexus. The endothelium of the artery showed intense staining for PAR-2 (arrowheads) and was similarly positive for factor VIII immunolabeling (data not shown). PAR-1 demonstrated similar expression patterns to those of PAR-2 in the vasculature (data not shown).
PAR-2 immunoreactivity was evident in the epithelium of many tissues. Most notable were the tissues of the gastrointestinal tract. In the intestine, PAR-2C labeling extended from the surface to the crypts of the epithelium (Figure 2J). In addition, PAR-2 expression in nonvascular smooth muscle was demonstrated throughout the gastrointestinal tract.
Nonvascular smooth muscle cells as well as stromal myofibroblasts labeled positively for PAR-2 in several tissues in addition to those of the gastrointestinal tract. As shown in Figure 2K, both bronchial epithelium and smooth muscle cells of human lung stained positive for PAR-2. For comparison, Figure 2L demonstrates positive labeling for smooth muscle actin. PAR-2 was also observed in the endothelium and epithelium of the alveolar sacs.
PAR-2 immunoreactivity was also clearly demonstrated in the brain. Neurons showed positive labeling for PAR-2 (Figure 2M) and several astrocytes showed weak immunolabeling (data not shown). Interestingly, not all neurons were positive, indicative of mixed populations of PAR-2-reactive cells. Figures 2N and 2O confirm the immunolabeling of astrocytes (arrowheads) with the antibody to glial fibrillary acidic protein and neurons (arrows) with the antibody neuron-specific enolase, respectively, in the brain.
Discussion
The physiological and/or pathological role of PAR-2 is unknown. Because the endogenous activating protease for PAR-2 has yet to be clearly defined, it has been difficult to identify specific functional roles of PAR-2 in vivo. The aim of this study was to provide a thorough characterization of the tissue-specific cellular localization of PAR-2 protein in vivo. In this report, the tissue distribution of PAR-2 in normal human tissues was described using a polyclonal antibody that selectively recognizes PAR-2. PAR-2 expression was highly expressed in most epithelial cells, including the vascular endothelium. PAR-2 was also abundant in vascular and nonvascular smooth muscle. Expression in stromal cells was tissue-specific. In addition, selective expression in cerebral neurons and astrocytes was noted.
Several reports have implicated PAR-2's involvement in vascular function (Hollenberg et al. 1996; Mirza et al. 1996). We found expression throughout the vascular endothelium, consistent with results obtained from in vitro studies using cultured endothelial cells that demonstrated mRNA for PAR-2 in addition to functional coupling of PAR-2 to calcium mobilization and cell proliferation (Mirza et al. 1996). Furthermore, PAR-2 activation has been shown to stimulate an endothelium-dependent nitric oxide-mediated relaxation of isolated vascular tissue (Hwa et al. 1996; Magazine et al. 1996) as well as a marked depressor response in vivo (Hwa et al. 1996). In the absence of endothelium, no direct vascular smooth muscle responses, either dilatory or contractile in nature, have been observed. Such studies have suggested that vascular smooth muscle does not express functional PAR-2. Our results, however, do not support such implications. Instead, we observed significant PAR-2 expression in vascular smooth muscle associated with all blood vessels examined. Moreover, PAR-2 activation induces calcium mobilization in cultured human vascular smooth muscle cells (unpublished observation). The significance of these observations is unclear. However, one can propose that PAR-2 may mediate other functions known to involve vascular smooth muscle, including responses to tissue injury such as migration, proliferation, or cytokine release. In this regard, recent studies implicate mast cell tryptase as a putative activating protease for PAR-2 (Molino et al. 1997). Mast cells reside in most tissues and, when activated, release substantial concentrations of tryptase in a localized manner. Activation of PAR-2 in such a microenvironment could contribute to smooth muscle-associated inflammatory responses. Clearly, our results suggest that further studies are warranted to explore the role of PAR-2 in vascular function.
PAR-2 expression was also evident in a variety of nonvascular tissues. Overall, a high correlation of tissue-specific PAR-2 expression in the epithelium and either the respective smooth muscle or the stromal layer was observed. Expression of PAR-2 throughout the gastrointestinal tract, both in the epithelium and the underlying smooth muscle, supports a role for PAR-2 in gastrointestinal function and is consistent with the results of Böhm et al. (1996), who demonstrated a similar localization pattern by in situ hybridization. The dense staining at the surface epithelium along the gastrointestinal tract is of special interest because the epithelial cells are likely to become exposed to trypsin, a putative physiological activator of PAR-2. The expression of PAR-2 within the smooth muscle of the gastrointestinal tract supports previous studies using isolated tissues that indicated a role for PAR-2 in regulating gastric motility (Al-Ani et al. 1995). Specific localization of PAR-2 in bronchial smooth muscle and epithelium was also observed and suggests a possible role for PAR-2 in maintaining airway function, perhaps contributing to airway responsiveness in asthma or other respiratory diseases. Co-expression in epithelial and smooth muscle cells of the same tissue suggests that PAR-2 may have a dual activity, perhaps in balancing the overall functions of the respective tissue.
One notable exception to such a distribution pattern was skin. Expression of PAR-2 only within the epidermis and not the dermis of skin provides a unique example of cell type-specific localization within a single tissue. The expression of PAR-2 within the epidermis suggests that PAR-2 may modulate epidermal structure and function in processes of wound healing, psoriasis, or skin-derived tumors. In vitro studies have indicated that stimulation of PAR-2 in human keratinocytes inhibits both their growth and their differentiation (Derian et al. 1997), which is consistent with the pronounced PAR-2 immunolabeling in the granular layer of the epidermis.
The expression of PAR-2 in the brain was a rather unexpected finding, although a recent report supports this observation (Smith–Swintosky et al. 1997). We observed expression of PAR-2 in some but not all neuronal cells and astrocytes. PAR-2 mRNA has been identified in neurons by RT in situ PCR (G. Nuovo, personal communication). The role of PAR-2 in the central and peripheral nervous system remains to be determined. PAR-2 activation might be associated with inflammatory responses after ischemic injury or trauma.
The results of the immunolocalization studies reported here compile a detailed profile of PAR-2 expression in different tissues and suggest a provocative role(s) for this novel receptor. Although this receptor is structurally similar to PAR-1, its physiological function(s) appears to be somewhat different. Clearly, additional studies are needed to define the significance of this receptor in normal or pathophysiological states to place PAR-2 as a distinctive member of this new class of proteolytically activated G-protein-coupled receptors.
Footnotes
Acknowledgements
We thank Christina J. Rogahn for excellent technical assistance, Jack Kauffman for preparation of the human platelets, Drs Charlotte Keenan and Barbara Kulwich for their morphological assistance, and Quality Photo Labs (Voorhees, NJ) for their high-quality photographic reproductions.